
Abstract
EstroG-100®, a functional ingredient blend that has been used for menopausal health in women. However, detailed studies on its effects and their underlying mechanisms have been lacking. Herein, we evaluated the antifatigue effects of EstroG-100 using C2C12 cells exposed to oxidative stress and mice subjected to a forced swimming test. EstroG-100 significantly increased the viability and adenosine triphosphate (ATP) content of C2C12 cells under oxidative stress, while notably inhibiting the generation of reactive oxygen species. EstroG-100 tended to prolong the forced swimming time in mice. It also significantly inhibited the serum lactate dehydrogenase activity, increased intramuscular glycogen content, and upregulated the expression of peroxisme proliferator-activated receptor (PPAR)-δ and uncoupling protein (UCP)3 genes involved in β-oxidation of fatty acids. Furthermore, EstroG-100 significantly increased the activities of antioxidant enzymes, including catalase, superoxide dismutase, glutathione S-transferase, and glutathione levels, and decreased the malondialdehyde levels in the liver. These results indicate that EstroG-100 may contribute to recovery from physical fatigue by enhancing the antioxidant activity to alleviate oxidative stress and improving muscle function via modulation of glycogen and fatty acid metabolism.
INTRODUCTION
Fatigue, defined as a feeling of excessive tiredness, energy depletion, and exhaustion, is a common symptom in daily life that negatively affects exercise capacity, domestic life, and work outcomes. Fatigue is classified as mental and physical fatigue; the former refers to cognitive or perceptual aspects of fatigue, whereas the latter impairs exercise ability. 1 Physical fatigue is caused by the overproduction of reactive oxygen species (ROS) and malondialdehyde (MDA) because of imbalances between the oxidant and antioxidant systems during excessive exercise. ROS overaccumulation imposes oxidative stress, which damages cells and tissues, leading to a build-up of metabolites, such as inorganic phosphates, lactic acid, adenosine diphosphate (ADP), calcium ions, and magnesium in the blood, impairing muscle function and causing physical fatigue.2,3 Accordingly, antioxidants can alleviate physical fatigue caused by exercise-related oxidative injury.
In normal menopause, which is related to decreased estrogen levels and other hormonal changes, mental and physical fatigue can accompany other menopausal symptoms, such as cold sweats, flushing, and sleep disorders; these symptoms interact and exacerbate fatigue. 4 In a cross-sectional study of 300 women, 85.3% of postmenopausal and 46.5% of perimenopausal women, but only 19.7% of premenopausal women, reported symptoms of physical or mental exhaustion. 5 Besides reducing the quality of life in middle-aged women, menopausal fatigue has adverse financial impacts, increasing the likelihood of early retirement, which can have considerable socioeconomic repercussions. 6 Therefore, safe and effective functional ingredients that improve menopausal fatigue need to be discovered.
EstroG-100® is a standardized extract derived from three plants. EstroG-100 and its major constituents, namely cinnamic acid, shanzhiside methylester, and nodakenin, possess antidepressant, anticancer, anti-inflammatory, and antioxidant properties and exhibit diverse physiological effects, including improvement of bone density and flushing.7–12 In three clinical trials in peri- and postmenopausal women, the group taking EstroG-100 showed statistically significant improvements in several menopausal symptoms, including total score in the Kupperman Menopausal Index and fatigue.13–15 However, despite the clinically relevant effects of EstroG-100, detailed studies of its effects on fatigue and the underlying mechanisms have been lacking.
This study was conducted with the aim of elucidating the antifatigue effects of EstroG-100 and the underlying mechanisms. We investigated the effects of EstroG-100 on oxidative stress in an H2O2-induced C2C12 cell model and analyzed changes in tissue and blood levels of fatigue-related factors in mice subjected to a forced swimming test (FST).
MATERIALS AND METHODS
Preparation of EstroG-100
EstroG-100 was prepared by mixing the dried roots of Cynanchum wilfordii, Phlomis umbrosa, and Angelica gigas at a weight ratio of 1:1:1.08 (w/w/w). Water, corresponding to 5–10 times the total weight of the raw materials (w/w), was added, and the mixture was extracted at temperatures above 95°C for over 4 h. The extract was filtered, concentrated under reduced pressure to 30 ± 5 Brix using a rotary evaporator, and subsequently dried to obtain the final extract.
C2C12 cell culture and differentiation
C2C12 cells were acquired from the American Type Culture Collection (Manassas, MD, USA), and cultured at 37°C, in 5% CO2 atmosphere in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, USA) containing 10% fetal bovine serum (TCB, USA) and 1% penicillin/streptomycin. To induce differentiation of myotubes, cells were seeded and cultured until approximately 80% confluence. The medium was then replaced with differentiation medium consisting of DMEM supplemented with 2% horse serum (HS; Gibco, USA), and cells were maintained in this medium for 3 days.
Measurement of cell viability
To mimic the oxidative muscle stress encountered during fatigue, we utilized an H2O2-treated C2C12 cell model. This model is widely accepted as a valid in vitro system for evaluating muscle fatigue-related oxidative damage. 16 The MTT assay was used to investigate the protective effects of EstroG-100 against cytotoxicity and H2O2-induced cell injury in C2C12 cells. The cells (5 × 104 cells/well) were seeded in 48-well plates and incubated for 24 h, after which the medium was exchanged for 2% HS to induce differentiation for 3 days. To investigate the cytotoxicity of EstroG-100, the cells were treated with different concentrations of EstroG-100 (25, 50, 100, and 200 μg/mL) and incubated for another 24 h, before adding 0.5 mg/mL 3-(4,5-dimethyl-2thiazolyl)-2,5-diphnyl-2H-tetrazolium bromide (MTT; Sigma-Aldrich Chemical Co., USA) to each well and incubating at 37°C for 3 h. Next, the medium was removed, the formazan produced was dissolved in dimethylsulfoxide, and the absorbance at 450 nm was measured using a microplate reader (SYNERGY H1; BioTek, USA). The cytoprotective effects of EstroG-100 were investigated in cells differentiated for 3 days and pretreated with EstroG-100 at different concentrations for 1 h. Subsequently, oxidative damage was induced by 1.2 mM H2O2 (Sigma-Aldrich Chemical Co.). After 24 h incubation, the cytoprotective effects were estimated using the MTT assay.
Measurement of ROS generation
Intracellular ROS generation was measured using dichloro-dihydro-fluorescein diacetate. C2C12 cells (2.5 × 104 cells/well) were seeded in white 96-well plates and incubated for 24 h. The medium was replaced with differentiation medium, and the cells were cultured for 3 days to induce differentiation. Afterward, cells were pretreated for 1h with EstroG-100 and 30 μM 2′,7′-dichlorofluorescin diacetate (Sigma-Aldrich Chemical Co.), a nonfluorescent probe. After washing with phosphate-buffered saline, the cells were exposed to 600 μM H2O2 (Sigma-Aldrich Chemical Co.) for 2 h to induce oxidative stress without causing significant cytotoxicity, and intracellular ROS generation was determined by measuring fluorescence (excitation wavelength 485 nm/emission wavelength 530 nm) using a microplate reader (SYNERGY H1; BioTek).
Measurement of intracellular ATP content
To investigate the effects of EstroG-100 on decreased adenosine triphosphate (ATP) levels due to H2O2-induced oxidative stress, we performed an ATP assay. C2C12 cells (5 × 104 cells/well) were seeded in a 48-well plate. After 24 h, the medium was exchanged for the differentiation medium, differentiation was induced for 3 days, and the cells were pretreated for 1 h with different concentrations of EstroG-100. Oxidative stress was induced with 1.2 mM H2O2 (Sigma-Aldrich Chemical Co.). After 24 h incubation, the cells were collected and the ATP content was measured using an ATP assay kit (Abcam, Cambridge, MA, USA).
Experimental animals
Five-week-old male ICR mice were obtained from SaeRon Bio (Uiwang, Korea) and housed under standardized environmental conditions (temperature: 22 ± 2°C, humidity: approximately 50%, and a 12 h light/dark cycle). After a 7-day acclimation period, the animals were randomly assigned to one of three groups (n = 10 per group): control, EstroG-100 50 mg/kg, and EstroG-100 200 mg/kg. FST was conducted based on the method originally described by Moriura et al. 17 and later modified by Matsumoto et al. 18 A transparent acrylic tank (90 × 45 × 45 cm³) was filled with distilled water to a depth of 38 cm and maintained at 34 ± 1°C. A unidirectional water flow of 7.5 L/min was created using a circulation pump to increase swimming resistance. Prior to the experiment, mice were habituated to the FST once daily between 1:00 and 3:00 PM for 3 consecutive days. EstroG-100 (50 or 200 mg/kg) was administered orally 1 h before the test. The swimming task continued until the mouse’s nose remained below the water surface for 5 sec, indicating fatigue. Exhaustion was defined as a loss of coordinated movement and inability to resurface within 10 sec. This testing procedure was repeated daily for 14 days, and the swimming duration was recorded immediately after each session. All experimental procedures were approved by the Institutional Animal Care and Use Committee of Kyung Hee University (approval number: KHGASP-19–104) and were conducted in accordance with the institutional guidelines.
Assessment of biochemical markers
After the FST, the mice were sacrificed using isoflurane, blood was collected via cardiac puncture, and the organs, including the liver and muscles, were harvested. The serum was isolated from the collected blood, and an ELISA (BioVision Inc., Milpitas, CA, USA) was performed to measure the lactate levels and lactate dehydrogenase (LDH) activity. Another ELISA kit (BioVision Inc.) was used to measure glycogen concentrations in the muscle tissue.
RNA extraction and quantitative real-time PCR
Total RNA was isolated from the muscle tissue using an RNeasy extraction kit (QIAGEN, Germantown, Maryland, USA). cDNA was synthesized using the iScript Select cDNA synthesis kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA) according to the manufacturer’s instructions. RT-qPCR was performed using a Step One Real-Time PCR system (Applied Biosystems, Foster City, CA, USA) according to the iQ SYBR Green Supermix (Bio-Rad Laboratories, Inc.) protocol. The PCR amplification was carried out under the following thermal cycling conditions: an initial denaturation at 95°C for 10 min, followed by 35 cycles consisting of denaturation at 95°C for 10 s, annealing at 57°C for 30 s, and extension at 72°C for 30 s. The sense and antisense primers were as follows: Glyceraldehyde-3-phosphate dehydrogenase (GAPDH), 5′-TGG CCT CCA AGG AGT AAG AAA C-3′ and 5′-CAG CAA CTG AGG GCC TCT CT-3′ (BC087743.1); PPAR-δ, 5′-CGC AAG CCC TTC AGT GAC AT-3′ and 5′-CGC ATT GAA CTT GAC AGC AAA-3′ (NM_011145); and UCP3, 5′-CCA GAG CAT GGT GCC TTC GCT-3′ and 5′-CTC GTG TCA GCA GCA GTG-3′ (MN_009464). The mRNA quantity was normalized against GAPDH and analyzed using the 2−ΔΔCt method. In the in vitro model, total RNA was isolated from C2C12 cells that had been differentiated for 3 days, followed by pretreatment with EstroG-100 for 1 h and subsequent exposure to 1.2 mM H2O2 for 24 h. The extracted RNA was then subjected to RT-qPCR analysis using the same protocol described above.
Analysis of antioxidant activity in mouse liver
The liver tissue was homogenized in NP-40 buffer, and the supernatant obtained after centrifugation (19,326 g at 4°C for 20 min) was collected. Total protein was quantified using the Bradford method. Finally, catalase (CAT), superoxide dismutase (SOD), and glutathione S-transferase (GST) activities were estimated using ELISA assay kits (BioVision Inc.). The concentrations of antioxidant glutathione (GSH) and MDA, an intermediate product of lipid oxidation in tissues, were also determined using ELISA kits (BioVision Inc.).
Statistical analysis
The in vitro experimental results are expressed as means ± standard deviations (SDs) from at least three independent experiments. Student’s t-test was used to analyze significant differences between groups. Results of in vivo experiments are presented as means ± SD (n = 10/group). The statistical analysis was performed using the SPSS 22.0 software (SPSS Inc., Chicago, USA). Differences among groups were analyzed using one-way analysis of variance followed by the Bonferroni test. Significance was defined as P < .05.
RESULTS
Effect of EstroG-100 on cell cytotoxicity and protection
Cytotoxicity and protective effects of EstroG-100 were measured using C2C12 cells. No cytotoxicity was observed at 25, 50, 100, or 200 μg/mL for 24 h (Fig. 1A). We also examined the protective effects of EstroG-100 in C2C12 cells exposed to H2O2-induced oxidative stress. The cells were exposed to 1.2 mM H2O2 after pretreating for 1 h with 25, 50, 100, or 200 μg/mL EstroG-100. Cell viability decreased to 63.06% in cells not treated with EstroG-100, but increased significantly in cells pretreated with 100 or 200 μg/mL EstroG-100 (Fig. 1B). Thus, EstroG-100 exhibited a cytoprotective effect against H2O2-induced oxidative stress.
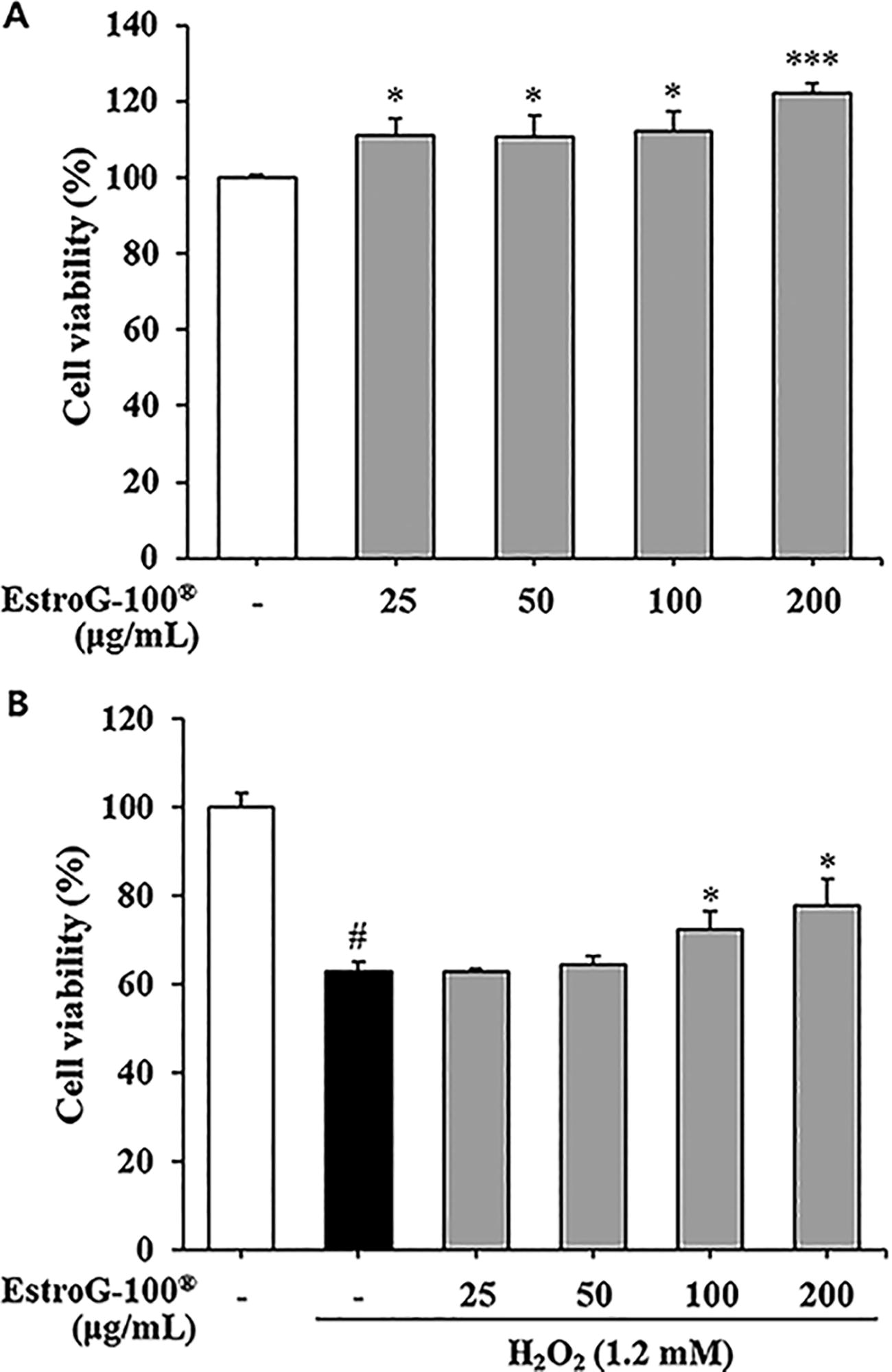
Effect of EstroG-100® on cell viability.
Effect of EstroG-100 on ROS generation
To assess the ROS scavenging activity of EstroG-100 under oxidative stress conditions without inducing cytotoxicity, we used 600 µM H2O2 in C2C12 cells. We confirmed that this concentration did not result in significant cytotoxicity after 2 h of exposure (Supplementary Fig. S1). ROS generation was markedly elevated in cells treated with only 600 μM H2O2 compared with untreated cells. However, pretreatment with 50, 100, and 200 μg/mL EstroG-100 significantly and dose-dependently inhibited ROS generation (Fig. 2). EstroG-100 exhibited strong ROS scavenging activity against H2O2-induced oxidative stress. These results indicated that the cytoprotective effect of EstroG-100 under H2O2-induced oxidative stress is related to the prevention of ROS generation.
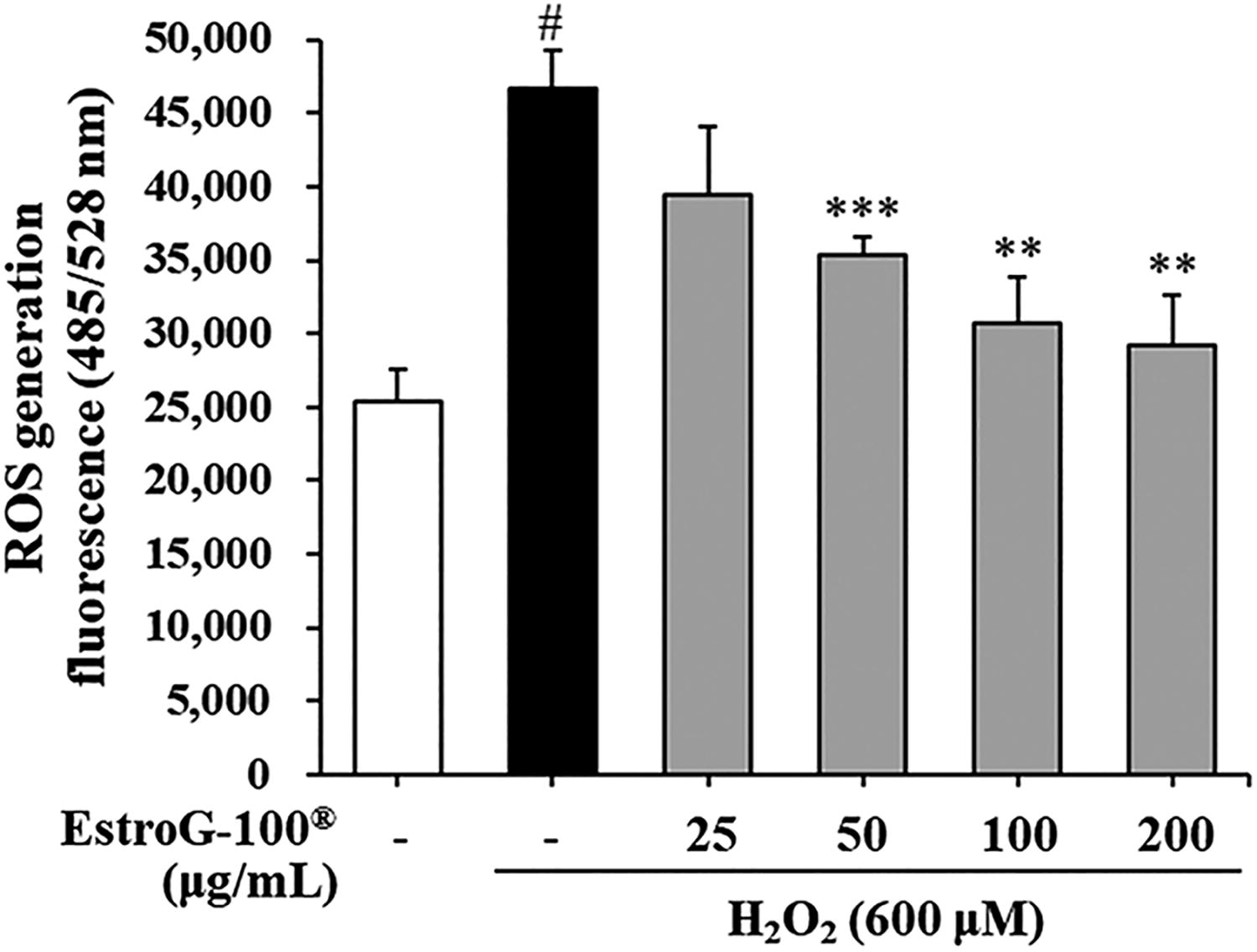
Effect of EstroG-100® on generation of reactive oxygen species (ROS) in hydrogen peroxide (H2O2)-induced C2C12 cells. All data are expressed as means ± standard deviations (n = 3). Statistical significance was compared with the control group at #P < .05 and with the H2O2 group at ***P < .001 and **P < .01 using the Student’s t-test. Groups were independently compared using the Student’s t-test. Statistical significance is indicated by P < .01 and P < .001.
Effect of EstroG-100 on intracellular ATP content
We measured intracellular ATP concentrations to investigate the ability of EstroG-100 to prevent reduced energy levels in C2C12 cells exposed to H2O2-induced oxidative stress. s. In cells treated with 1.2 mM H2O2, the relative ATP concentration decreased to 79.02%, whereas it increased significantly in cells pretreated with 25, 50, 100, and 200 μg/mL EstroG-100 (Fig. 3). Thus, EstroG-100 inhibited the oxidative stress-induced decrease in ATP.
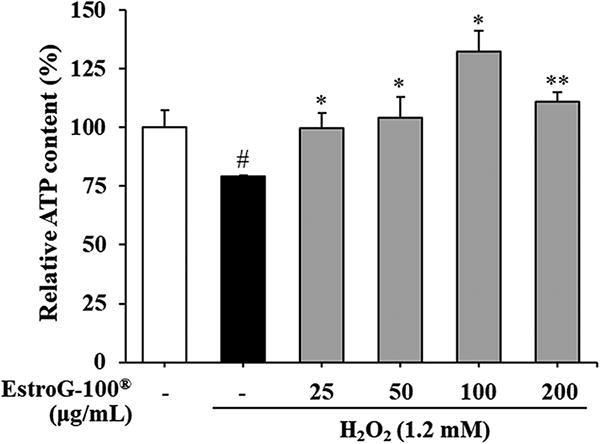
Effect of EstroG-100® on adenosine triphosphate (ATP) content in hydrogen peroxide (H2O2)-induced C2C12 cells. All data are expressed as means ± standard deviations (n = 3). Statistical significance was compared with the control group at #P < .05 and with the H2O2 group at **P < .01 and *P < .05 using the Student’s t-test. Groups were independently compared using the Student’s t-test. Statistical significance is indicated by P < .05 and P < .01.
Effect of EstroG-100 on forced swimming capacity
To evaluate the effects of EstroG-100 on muscle activity using a mouse model, we measured the time to immobility in FST at the start (day 0) and end (day 14) of the experiment (Fig. 4A). On day 0, no significant differences in the swimming time were observed compared with the control (23.46 ± 2.08 min), 50 mg/kg EstroG-100 (22.17 ± 3.11 min), and 200 mg/kg EstroG-100 (22.06 ± 2.46 min) groups. On day 14, the EstroG-100 groups (50 mg/kg: 24.27 ± 2.51 min; 200 mg/kg: 25.36 ± 3.24 min) showed increased swim time compared with the control group (22.43 ± 2.15 min), but the increase was not statistically significant (Fig. 4B).
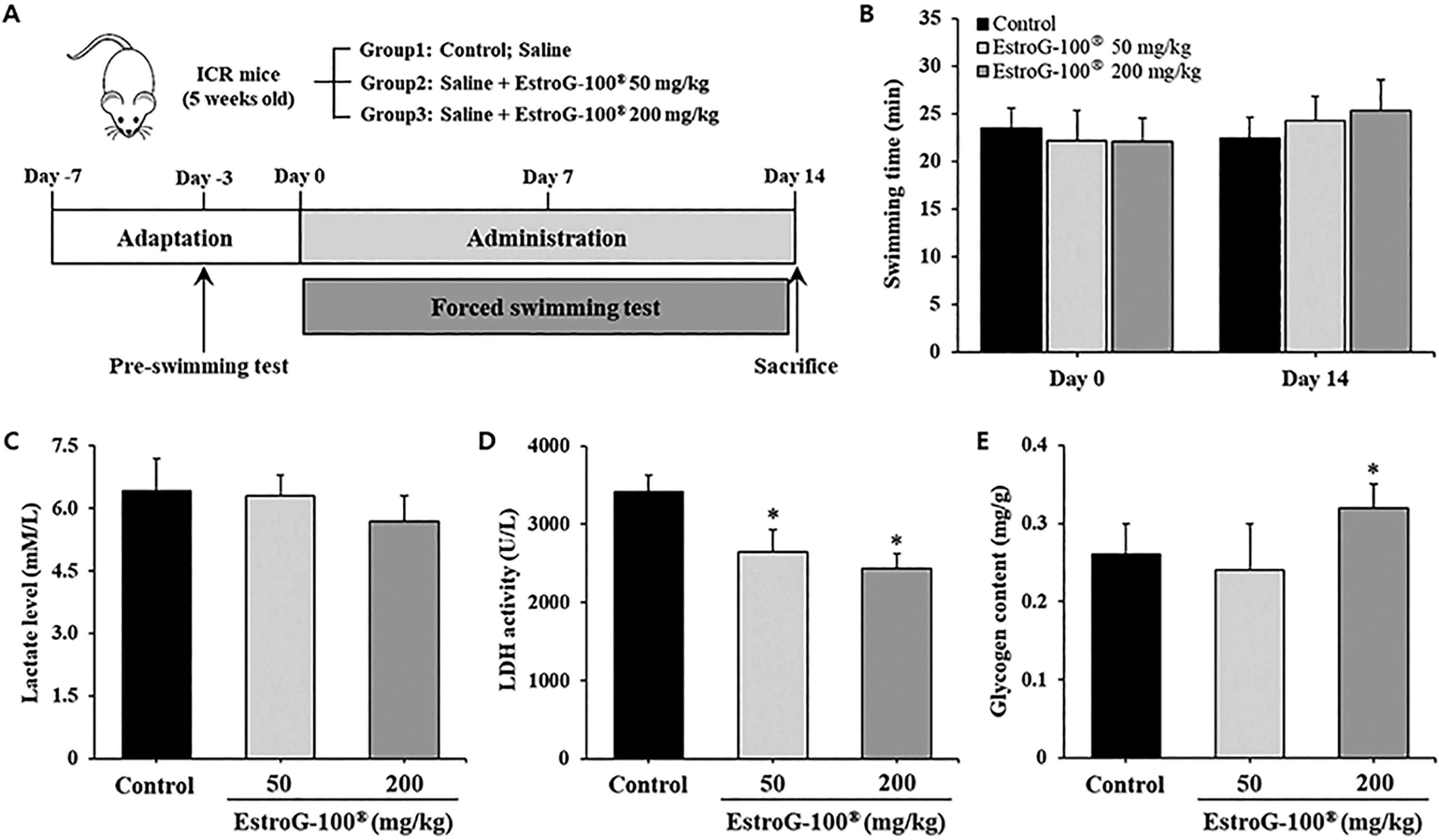
Effect of EstroG-100® on forced swimming test (FST)-induced fatigue in mice.
Effect of EstroG-100 on serum lactate levels and LDH activity
The lactate levels in mouse serum were 6.42 ± 0.78 mM/L in the control, 6.31 ± 0.49 mM/L in the 50 mg/kg EstroG-100, and 5.68 ± 0.62 mM/L in the 200 mg/kg EstroG-100 group, exhibiting 11.5% decrease in the 200 mg/kg EstroG-100 group compared with the control group, but this decrease was not significant (Fig. 4C). LDH activity was 3420 ± 275 U/L in the control, 2651 ± 194 U/L in the 50 mg/kg EstroG-100, and 2434 ± 207 U/L in the 200 mg/kg EstroG-100 group, decreasing significantly compared with the control group, by 22.5% and 28.8%, respectively, in the 50 mg/kg and 200 mg/kg EstroG-100 groups (Fig. 4D).
Effect of EstroG-100 on intramuscular glycogen content
The intramuscular glycogen content was 0.26 ± 0.04, 0.24 ± 0.06, and 0.32 ± 0.03 mg/g in the control, 50 mg/kg EstroG-100, and 200 mg/kg EstroG-100 groups, respectively. The 200 mg/kg EstroG-100 group showed a significant increase of 23.1% in the intramuscular glycogen content, demonstrating its prevention of glycogen loss (Fig. 4E).
Effect of EstroG-100 on muscle energy metabolism-related gene
To investigate the effects of EstroG-100 on intramuscular fatty acid oxidation control factors, we analyzed PPAR-δ mRNA expression. PPAR-δ mRNA expression was significantly upregulated in the 50 and 200 mg/kg EstroG-100 groups compared with the control group (Fig. 5A). A significant dose-dependent increase was noted in the mRNA expression for UCP3, an energy metabolism control factor, in the EstroG-100 groups (Fig. 5B).
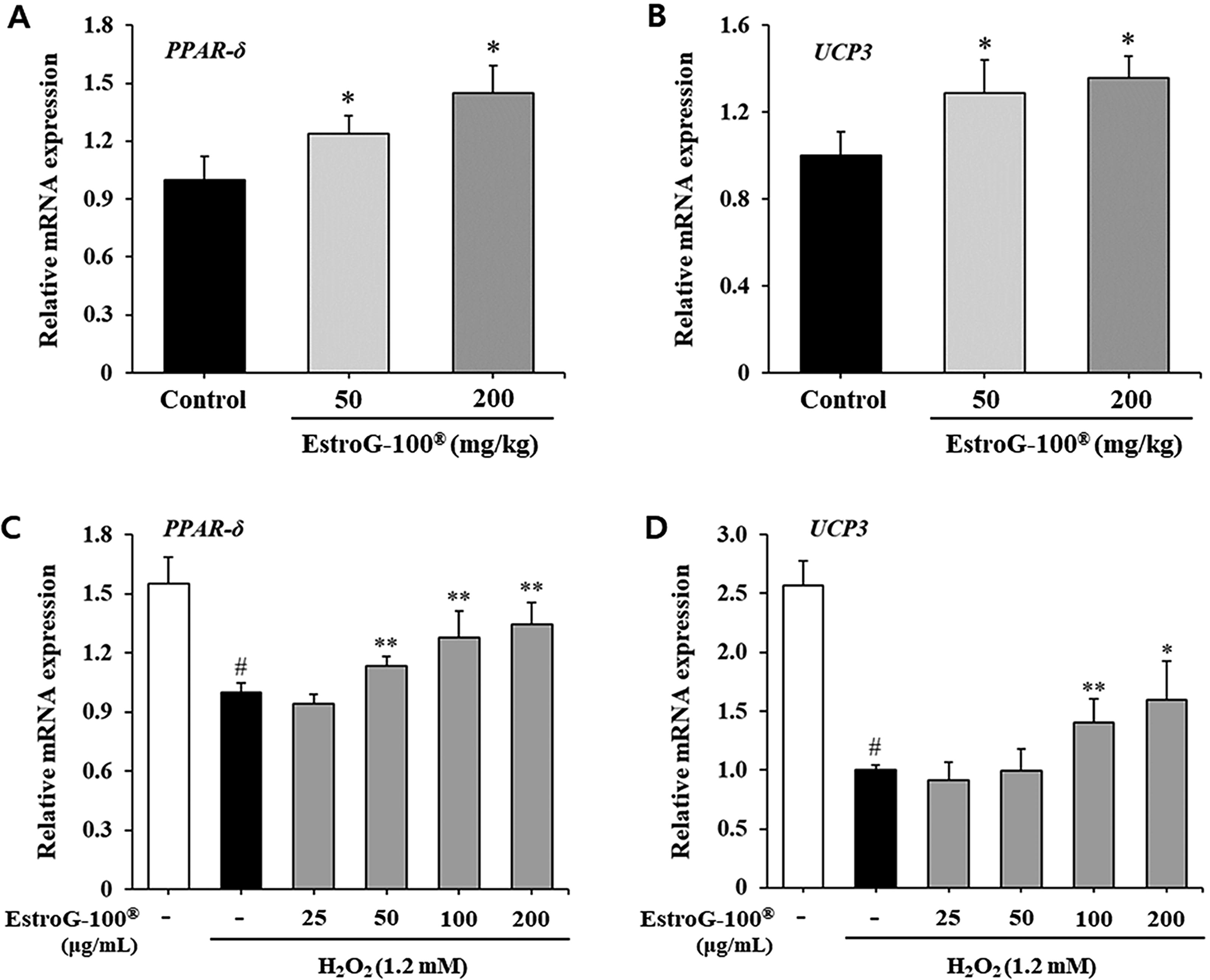
Effect of EstroG-100® on muscle energy metabolism-related factors.
To determine whether the observed in vivo transcriptional changes were also a direct result of cellular mechanisms, we examined the mRNA expression of PPAR-δ and UCP3 in C2C12 cells exposed to H2O2-induced oxidative stress. Exposure to 1.2 mM H2O2 for 24 h markedly suppressed the expression of both genes, which is likely attributable to cellular damage or apoptosis resulting from prolonged oxidative insult. In contrast, pretreatment with EstroG-100 restored and significantly upregulated the expression of PPAR-δ and UCP3 in a dose-dependent manner (Fig. 5C,D). Thus, EstroG-100 improved exercise fatigue and ability by regulating intramuscular energy factors against ROS-induced physical fatigue.
Effect of EstroG-100 on hepatic enzyme activities
To investigate the effects of EstroG-100 on hepatic antioxidant enzyme activity, we measured CAT, SOD, and GST activity. CAT activity was 15.28 ± 1.21, 17.54 ± 0.78, and 22.18 ± 1.54 U/mg in the control, 50 mg/kg EstroG-100, and 200 mg/kg EstroG-100 groups, respectively; thus, compared with the control group, the EstroG-100 groups showed significant increases of 14.8% and 45.2% in CAT activity. SOD activity was 14.12 ± 0.94, 20.31 ± 1.98, and 23.40 ± 1.73 U/mg in the control, 50 mg/kg EstroG-100, and 200 mg/kg EstroG-100 group, respectively; thus, compared with the control group, the EstroG-100 groups showed large significant increases of 43.8% and 65.7% in SOD activity. Finally, GST activity was much higher than CAT and SOD activity, at 38.41 ± 3.03, 42.36 ± 2.12, and 46.24 ± 3.25 U/mg in the control, 50 mg/kg EstroG-100, and 200 mg/kg EstroG-100 groups, respectively; thus, compared with the control group, the EstroG-100 groups showed dose-dependent significant increases of 10.3% and 20.4% in GST activity (Fig. 6). These results indicate that EstroG-100 is effective for alleviating fatigue by increasing activity of hepatic antioxidant enzymes in response to fatigue induced by forced swimming.
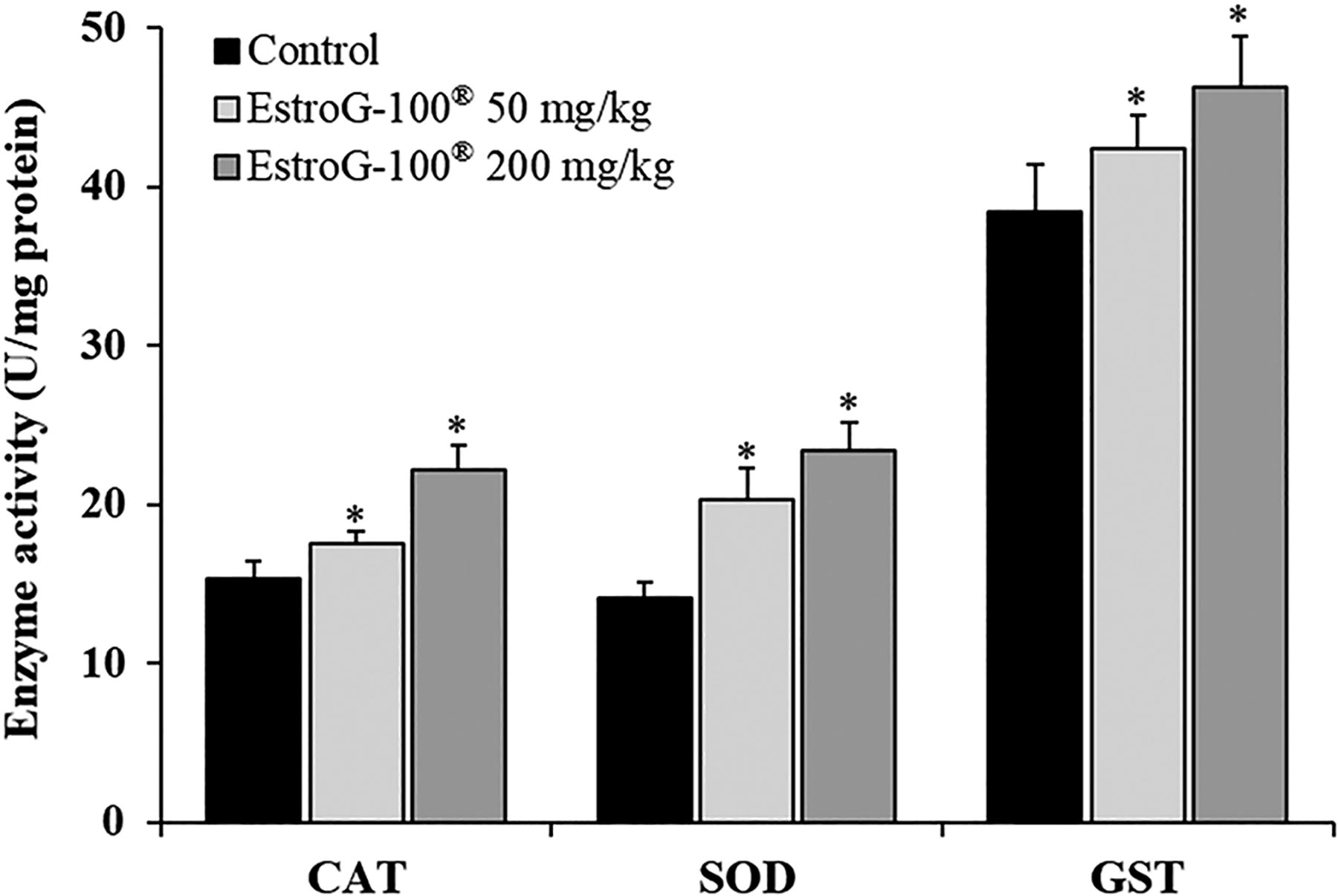
Effect of EstroG-100® on CAT, SOD, and GST enzyme activities in liver tissue in FST-induced mice. All data are expressed as means ± standard deviations for 10 mice in each group. Statistical analyses were performed using Bonferroni tests after one-way ANOVA with the SPSS software. Differences were considered statistically significant at P < .05. CAT, catalase; GST, glutathione S-transferase; SOD, superoxide dismutase.
Effects of EstroG-100 on GSH and MDA levels in the liver
The hepatic concentration of GSH, a nonenzymatic antioxidant, was 12.79 ± 1.22, 13.10 ± 0.98, and 15.77 ± 1.21 μmoles/mg protein in the control, 50 mg/kg EstroG-100, and 200 mg/kg EstroG-100 group, respectively; thus, a significant 23.3% increase in the high-dose group was observed compared with the control group. MDA concentration was 6.91 ± 0.35, 5.48 ± 0.17, and 4.34 ± 0.12 moles/g protein in the control, 50 mg/kg EstroG-100, and 200 mg/kg EstroG-100 group, respectively; thus, compared with the control group, a significant decrease of 20.7% and 37.2% occurred in the EstroG-100 groups (Table 1). Based on these findings, EstroG-100 is expected to protect the liver from damage by increasing the activity of antioxidant enzymes, while also increasing the concentration of nonenzymatic antioxidants.
Effect of EstroG-100® on GSH and MDA Levels in Mouse Liver

All data are expressed as means ± standard deviations for 10 mice in each group. Statistical analyses were performed using Bonferroni tests after one-way ANOVA using the SPSS software. Differences were considered statistically significant at P < .05.
ANOVA, analysis of variance; GSH, glutathione; MDA, malondialdehyde.
DISCUSSION
FST is used as a valid animal model to assess the antifatigue effects of various physiologically active substances. 19 Based on a 2-week FST, we confirmed a trend for increased swimming time in mouse groups treated with EstroG-100 (Fig. 4B). ATP and glycogen are essential for maintaining sustained muscle function during exercise. ATPase, which is involved in muscle contraction, ion balance, and relaxation, requires considerable ATP, whereas glycogen is an important energy source. As such, glycogen depletion harms muscle function and is a major cause of fatigue. 1 Fatty acids are also used as an energy source during exercise via energy-producing metabolic pathways, such as fatty acid β-oxidation. In these pathways, PPAR-δ is a transcription factor that regulates enzymes involved in the oxidation and transport of fatty acids in muscles, and UCP3 promotes fatty acid oxidation and protects mitochondria from ROS. 20 A study by Chen et al. demonstrated that naringin, a citrus-derived flavonoid, alleviates physical fatigue by enhancing mitochondrial function and antioxidant capacity, and significantly upregulates PPAR-δ and UCP3 expression. 21 In our study, EstroG-100 increased the intramuscular glycogen content (Fig. 4E) and significantly upregulated the expression of genes (PPAR-δ and UCP3) involved in fatty acid energy metabolism in mice (Fig. 5A,B). Consistent with these findings, EstroG-100 upregulated PPAR-δ and UCP3 expression (Fig. 5C,D) and mitigated ATP depletion (Fig. 3) in C2C12 cells under oxidative damage. These results suggest that EstroG-100 may exert antifatigue effects by promoting mitochondrial function and fatty acid oxidation.
Overproduction of ROS due to excessive exercise causes oxidative stress, which can lead to lipid peroxidation and oxidative damage to liver and muscle tissue. 22 As oxidative stress damages cell membranes, cytoplasmic enzymes, such as LDH, creatine kinase, aspartate aminotransferase, and alanine aminotransferase, leak into the blood. 23 LDH catalyzes the conversion of pyruvic acid to lactic acid in energy metabolism, and its activity increases during excessive exercise, promoting the conversion of pyruvic acid to lactic acid, and causing fatigue. 24 Levels of MDA, a major product of open-access radical-mediated injury and an intermediate in lipid peroxidation, increase in the liver during exercise. 25 In our study, EstroG-100 significantly decreased serum LDH activity (Fig. 4D) and significantly lowered hepatic MDA levels (Table 1). Moreover, EstroG-100 protected muscle cells (Fig. 1B) by inhibiting H2O2-induced ROS generation (Fig. 2). Thus, EstroG-100 could exhibit antifatigue effects by inhibiting oxidative stress-induced lipid peroxidation and cell injury.
Antioxidant defense systems protect the body from oxidative stress and are important for removing ROS and their metabolic products to maintain normal cell function. Enzymatic and nonenzymatic antioxidants provide the main defense against oxidative stress. The activities of antioxidant enzymes decrease after excessive exercise.26,27 CAT, SOD, and GST help protect cells by inhibiting the generation of open-access radicals and metabolic products produced under oxidative stress. GSH exhibits nonenzymatic antioxidant activity and is involved in detoxifying radicals through its antioxidant activity. 28 EstroG-100 significantly increased hepatic activity of CAT, SOD, and GST (Fig. 6), and at 200 mg/kg/day, EstroG-100 also significantly increased the GSH levels (Table 1). These results show that EstroG-100 can enhance hepatic antioxidant activity, thereby ameliorating oxidative stress in muscle cells and forming an environment that is conducive to normal muscle function. While EstroG-100 did not significantly enhance swimming time during the 2-week administration period, it did significantly modulate fatigue-related biochemical markers, including LDH activity, glycogen content, and antioxidant enzyme levels. These changes may indicate initial biological responses, and a longer treatment period may be necessary to determine whether they lead to behavioral improvements.
Our results indicate that EstroG-100 helps defend against oxidative stress by delaying energy depletion via glycogen and fatty acid oxidation and by promoting antioxidant enzyme activity. Thus, EstroG-100 is an effective intervention for improving physical fatigue. Our study provides preliminary mechanistic support for its antifatigue effects observed in clinical trials in menopausal women. However, because our study was focused on improving FST-induced physical fatigue in male mice, there are limitations in applying our results directly to fatigue in menopausal women. Furthermore, although this study demonstrated alterations in fatigue-related gene expression at the mRNA level, the lack of protein-level validation is a limitation. The upregulation of PPAR-δ and UCP3 offers initial mechanistic insight, but further investigation involving protein expression analysis and pathway-specific studies is needed to confirm the underlying mechanisms of EstroG-100. To better reflect the physiological conditions associated with menopausal fatigue, future research should also utilize animal models with altered female hormone levels, such as ovariectomized rats. These approaches will enable a more comprehensive understanding of the antifatigue mechanisms of EstroG-100 in menopausal women.
Finally, the doses of EstroG-100 administered to mice in this study were 50 and 200 mg/kg. When converted to a human equivalent dose using the body surface area normalization method suggested in the U.S. Food and Drug Administration’s Guidance Documents, these correspond to a dose of 243–972 mg for a 60 kg adult. 29 Based on this, a daily EstroG-100 intake of 243 mg or more would be expected to have antifatigue effects in humans.
AUTHORS’ CONTRIBUTIONS
Conceptualization: Y.-Y.H. and J.L. Methodology: Y.-Y.H. and J.L. Investigation: Y.-Y.H., J.-H.O., J.-M.K., B.-M.K., C.-S.P., Y.-W.L., and M.L. Data curation: Y.-Y.H. and M.L. Writing—original draft: Y.-Y.H. and J.L. All authors have read and approved the final version of the article.
Footnotes
AUTHOR DISCLOSURE STATEMENT
The authors declare no conflicts of interest.
FUNDING INFORMATION
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.