
Abstract
Hibiscus acid has antimicrobial activity against pathogenic bacteria; however, the mechanism of action against these bacteria has not been completely identified. The effect of hibiscus acid on the viability, motility, and cell membrane of multidrug-resistant enterohemorrhagic Escherichia coli, Salmonella Typhimurium, and Listeria monocytogenes was determined. Cell viability was determined by the pour plate technique, and motility was determined by phase contrast microscopy. Acridine orange/ethidium bromide fluorescent staining was performed using a fluorescence microscope to observe live and dead cells. Bacterial cell damage was analyzed by scanning electron microscopy. Finally, nucleotide leakage (DNA and RNA) was evaluated by measuring absorbance at 260 nm using a UV-VIS spectrophotometer. Hibiscus acid treatments (minimum inhibitory concentration [MIC], minimum bactericidal concentration [MBC], 10× minimum inhibitory concentration [10× MIC]) significantly decreased the viable cell counts of the three pathogenic bacterial strains in a time-dependent manner. Hibiscus acid at concentrations of MIC, MBC, and 10× MIC inhibited the motility of bacteria and caused lysis of the bacterial cell membrane. The damage to the membrane was greater as the concentration of hibiscus acid increased. High red fluorescence was found in the cells treated with hibiscus acid at the MIC and 10× MIC, and the release of intracellular material was observed after 2 h of treatment with the three pathogenic bacteria studied. The antimicrobial effect of hibiscus acid was due to the alteration of the membrane permeability of the three pathogenic bacteria studied. Hibiscus acid may represent an alternative to antibiotics for the treatment or prevention of infectious bacterial diseases.
Keywords
INTRODUCTION
Antibiotics have saved millions of lives and have also represented a revolution in medicine. They have contributed significantly to progress in fields such as solid organ transplants and the survival of premature and immunosuppressed babies, where infections are especially prevalent. However, for some years now, a growing threat has been deteriorating the effectiveness of these drugs: bacterial resistance to antibiotics.1,2 Bacteria resistant to multiple antibiotics represent a major concern worldwide, causing diseases that kill hundreds of thousands of people each year. The possibility of developing resistant bacterial populations when using antibiotics as growth promoters in animal production has led several countries to prohibit the use of antibiotics in the breeding and development of food animals. 3 In response to the significant increase in this type of bacteria, legal measures have been taken to limit or eliminate the use of antibiotics.4–7
In recent decades, it has been necessary to find new alternatives to prevent infectious diseases caused by pathogenic bacteria. For this reason, special attention has been paid to the antimicrobial activity derived from extracts and compounds of plant origin, of which there is a great deal of information from in vitro studies on their bactericidal or bacteriostatic effects against various pathogenic microorganisms, including multidrug-resistant strains.8–11 The exploration of plant-based antimicrobials not only reveals their potent antimicrobial activity but also highlights their potential as sustainable solutions to the escalating crisis of antibiotic resistance. By leveraging the diverse mechanisms of action exhibited by these natural compounds, researchers are paving the way for innovative treatments that could effectively combat drug-resistant infections. 12 One of the potential plants for use against antibiotic-resistant bacteria is Hibiscus sabdariffa. It has been used as an additive to foods and beverages and in traditional medicine for the treatment of some diseases.13–15 Its extracts and/or components have shown antimicrobial activity in vitro and in vivo against multiresistant and resistant pathogenic bacteria to antibiotics.14,16–20 In addition, these extracts and components present other beneficial biological activities; for example, hibiscus acid has been shown to have a vasodilatory effect in the aortas of rats. 21 Additionally, this compound has been shown to inhibit the activity of angiotensin-converting enzymes in vitro with values similar to captopril. 22 In addition, the H. sabdariffa extracts, as well as the hibiscus acid, have been shown to reverse foot ageing. 23
In previous studies, we have shown that hibiscus acid obtained from an acetonic extract of H. sabdariffa calyxes is among the compounds responsible for antimicrobial activity against multiantibiotic-resistant pathogenic bacteria 17 ; however, the antimicrobial mechanism of action of hibiscus acid has not been fully characterized. The objective of this study was to determine the effect of hibiscus acid on the motility, viability, and cell membrane of multiantibiotic resistant enterohemorrhagic E. coli (EHEC), S. Typhimurium, and L. monocytogenes.
MATERIALS AND METHODS
Bacterial strains
Three pathogenic strains were used: EHEC (O157:H7) isolated in our laboratory from raw beef, S. Typhimurium isolated from coriander 24 and L. monocytogenes ATCC 19115. All strains were resistant to multiple antibiotics (kanamycin, neomycin, streptomycin, amikacin, tetracycline, erythromycin, chloramphenicol, ceftriaxone, nalidixic acid, and trimethoprim/sulfamethoxazole) according to the criteria of the Clinical and Laboratory Standards Institute. 25
Extraction of hibiscus acid
Hibiscus acid was obtained as previously reported. 17 Briefly, hibiscus acid was successfully obtained in crystalline form from the dehydrated acetone extract of H. sabdariffa calyces. The purity of the crystals was assessed using thin-layer chromatography, followed by recrystallization. Identification of the crystals was performed through Nuclear Magnetic Resonance Spectroscopy, X-ray Crystallography, and Differential Scanning Calorimetry. Subsequently, the crystals were stored at a temperature between 20°C and 25°C for future use.
Determination of the death curves
The bactericidal activity of hibiscus acid against EHEC, S. Typhimurium, and L. monocytogenes was evaluated by measuring the reduction in the number of colony-forming units (CFUs) over time. 26 S. Typhimurium, EHEC, and L. monocytogenes cells were used to inoculate trypticase soy broth (TSB) containing hibiscus acid at different concentrations, 17 as shown in Table 1. All tubes were incubated at 35 ± 2°C. At different times, each tube was seeded into culture plates containing standard method agar using the pour plate technique and incubated at 35 ± 2°C for 24 h. TSB tubes without hibiscus acid were included as a negative control. All studies were performed in triplicate. An aqueous solution ethylenediaminetetraacetic acid (EDTA) was used as a positive control at a concentration of 0.25 M in all assays conducted in this study. 27 EDTA was added to the TSB tubes containing the pathogen under the same conditions as hibiscus acid. In gram-negative bacteria, EDTA induces permeabilization of the outer membrane and cell lysis. 28 Chelators, such as EDTA, disintegrate the outer membrane, eliminating Mg2+ and Ca2+, and are considered effective and valuable permeabilisers. 29
Concentrations of Hibiscus Acid Used per Pathogen in the Experiments
10× MIC: 10× minimum inhibitory concentration; MBC, minimum bactericidal concentration; MSIC, minimum subinhibitory concentration.
Changes in cell motility
For the motility experiments, the technique described by Gill and Holley 30 was used with some modifications. Instead of incubating at 20°C, EHEC, S. Typhimurium, or L. monocytogenes were incubated in TSB enriched with Yeast Extract (TSB+YE) at room temperature (22–25°C) for 18 h. Bacterial suspensions were centrifuged at 9780 g for 20 min. The supernatant was discarded, and the cell pellets were washed twice with 10 mL of potassium phosphate buffer solution (PBS, 0.5 mM, pH 7.4). The bacterial cell suspension was prepared by resuspending the cell pellet in a tube containing 10 mL of PBS. An aliquot of 350 µL of the bacterial cells was taken in the buffer solution and placed in Eppendorf tubes with 350 µL of TSB + YE (2×). After 5 min of incubation at room temperature, hibiscus acid was added at different concentrations (MSIC, minimum inhibitory concentration [MIC], minimum bactericidal concentration [MBC], 10× minimum inhibitory concentration [10× MIC]) to the medium containing the pathogenic bacteria. Cells without treatment and added EDTA were used as controls. A 10-µL sample of bacterial cells was immediately plated onto a glass slide with coverslip, and the sample was examined by phase contrast microscopy at 100× magnification for 10 min.
Fluorescence microscopy staining
The methodology described by Hameed et al. 31 and Khan et al. 32 was followed by some modifications. Live and dead cells were evaluated by acridine orange and ethidium bromide (AO/EB) fluorescent staining. A 10-µL suspension of EHEC was used to inoculate the centrifuge tubes with 3 mL of TSB and incubated at 35 ± 2°C for 24 h. The bacterial suspensions were centrifuged at 10,000 rpm for 20 min. The supernatant was discarded, and the cell package was washed twice with 3 mL of PBS. Washed EHEC cells were treated with hibiscus acid at two concentrations (MIC and 10× MIC) for 60 min at 35 ± 2°C. After incubation, the bacterial cells were centrifuged, washed with PBS, and stained with a 1:1 AO/EB solution at a concentration of 100 µg/mL for 30 min. The cells were washed with PBS by centrifugation and observed under a UV fluorescence microscope (Motic, BA210E, China). For fluorescence excitation, a 460–495 nm excitation filter was used for green fluorescence, and a 530–550 nm excitation filter was used for red fluorescence detection.
Scanning electron microscopy
The effect of hibiscus acid on the cell morphology of L. monocytogenes and EHEC was observed using scanning electron microscopy (SEM). Briefly, 10 µL of EHEC or L. monocytogenes were used to inoculate the centrifuge tubes with 3 mL of TSB and incubated at 35 ± 2°C for 24 h. The bacterial suspensions were centrifuged twice at 5000 rpm for 20 min; on each occasion, the cell package was resuspended with PBS (0.5 mM, pH 7.4). The washed bacterial cells were added to tubes containing hibiscus acid solution at two concentrations, MIC and 10× MIC, and stored at 35 ± 2°C for 3 h. PBS solution was used as a negative control. Bacterial cells were washed three times at 4500 rpm for 25 min using PBS, and the cell package was lyophilized. The lyophilized cells were double-coated with a gold sputter coating for 3 min each, and the samples were observed under a scanning electron microscope (Oxford, Instruments, JEOL X-max N, Japan). 33
Leakage of intracellular material
The experiment was performed following Tang et al. 34 with some modifications. Separately, 10 µL of EHEC, S. Typhimurium, or L. monocytogenes were used to inoculate the centrifuge tubes with 3 mL of TSB and incubated at 35 ± 2°C for 24 h. The bacterial suspensions were washed twice by centrifuging at 5000 rpm for 20 min. On each occasion, the cell package was resuspended with 3 mL PBS. The washed bacterial cells were added to tubes containing hibiscus acid at different concentrations: MSIC, MIC, MBC, and 10× MIC. EDTA solution was used as a positive control, and PBS solution was used as a negative control. The tubes were stored at 35 ± 2°C for different times (0, 2, 4, 6, and 8 h). At each sampling time, tubes of each microorganism and hibiscus acid concentration were taken and centrifuged at 4500 rpm for 25 min. The supernatant was recovered, and the absorbance at 260 nm at room temperature was determined using a UV-VIS spectrophotometer (Thermo Scientific, NanoDrop one, USA). The results were expressed in the form of optical density recorded at each time interval. The experiment was performed in triplicate.
Statistical analysis
The study utilized a statistical multifactorial design with three replicates. Prior to applying analysis of variance (ANOVA) and the Tukey post hoc test (P ≤ .05) using the STATISTICA program (Version 8.0), the assumptions of normality and homogeneity of variances were evaluated using the Shapiro–Wilk and Levene tests, respectively. Since these assumptions were met, significant differences between treatments were determined by ANOVA followed by the Tukey test.
RESULTS AND DISCUSSION
Death curves of EHEC, Salmonella Typhimurium, and Listeria monocytogenes treated with hibiscus acid
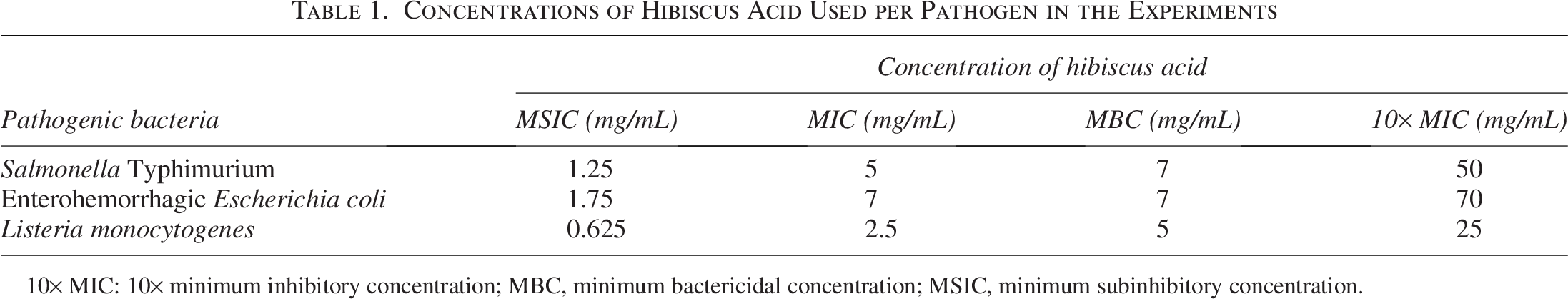
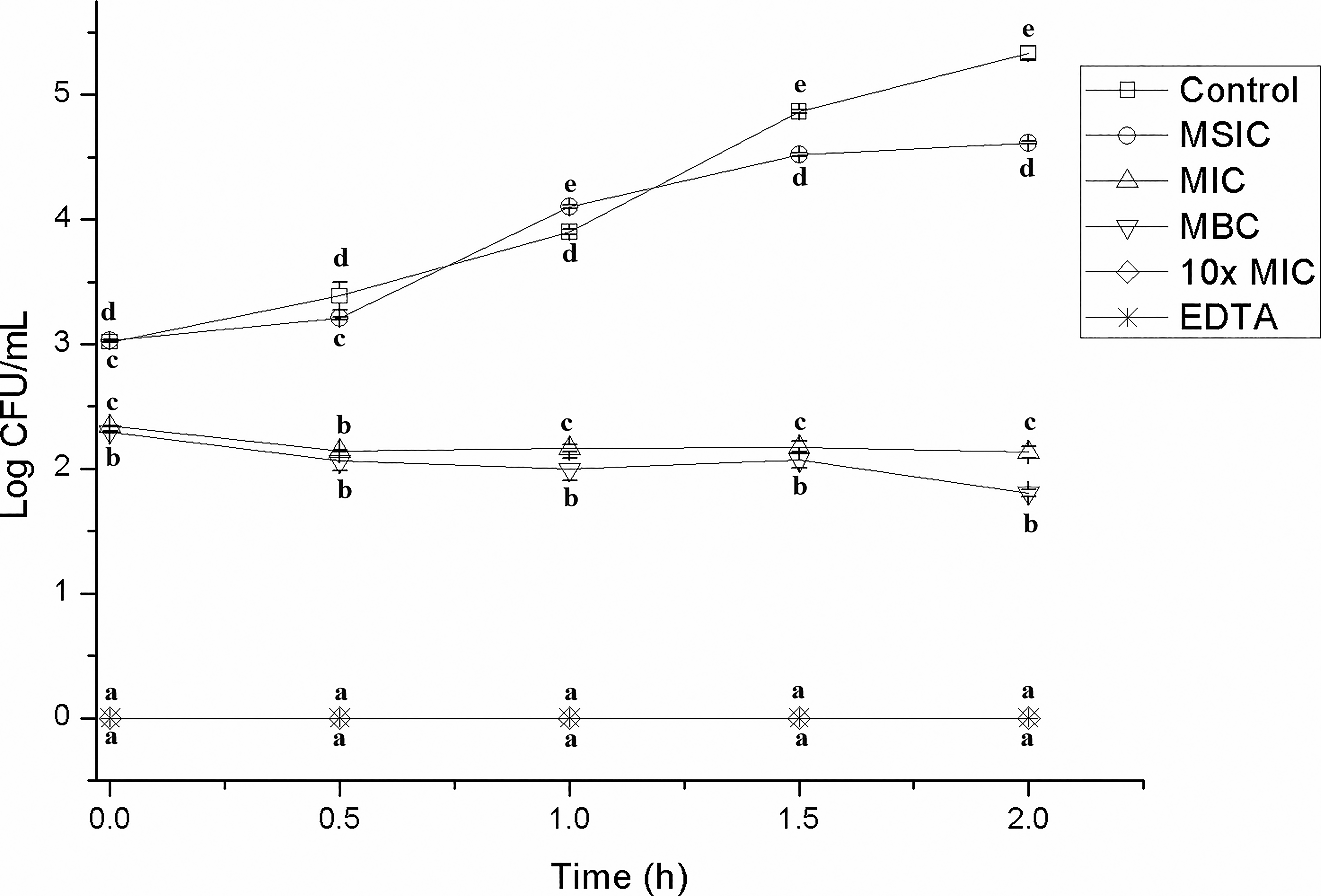
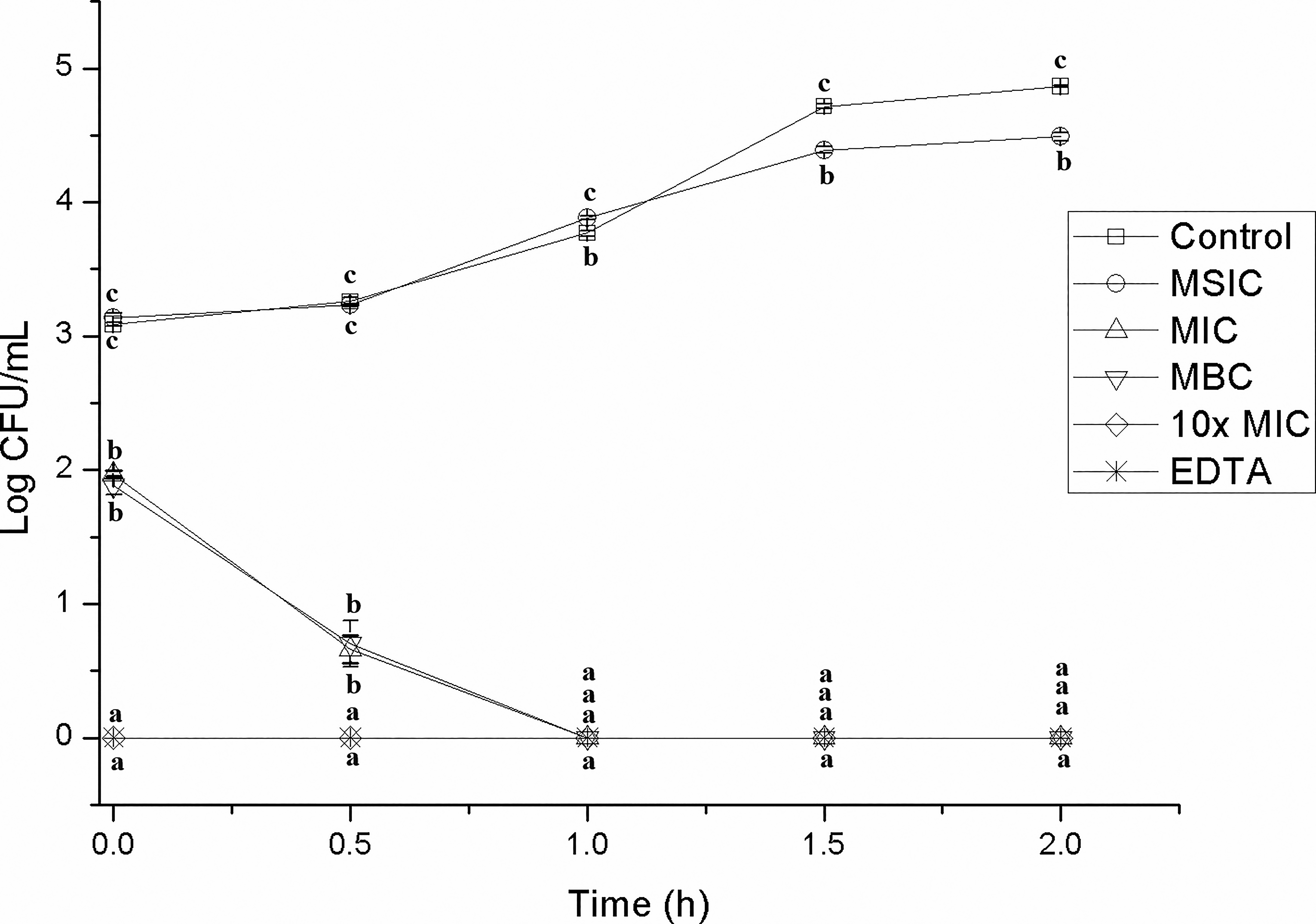
In this assay, hibiscus acid significantly affected the viability of EHEC, S. Typhimurium, and L. monocytogenes (Figs. 1–3, respectively). In all cases, at the hibiscus acid concentration of 10× MIC from the first moment of contact, the three pathogens were no longer detected by the analysis technique (Figs. 1–3). In the case of MIC and MBC concentrations, significant reductions were also obtained from time 0; however, after the first contact time, the effect was different depending on the type of microorganism (Figs. 1–3). The greatest effect of the MIC and MBC concentrations occurred on S. Typhimurium and the smallest on EHEC (Figs. 1–3). The MSIC of hibiscus acid apparently had no effect on the viability or development of the three studied bacteria; they presented a behavior similar to that shown in the control (Figs. 1–3). The results showed the effect of hibiscus acid on the viability and development of EHEC and S. Typhimurium. The bacteriostatic action of hibiscus acid was confirmed through death curve studies, which revealed that both the MIC and MBC exhibited a bacteriostatic effect over a 2-h study period. In contrast, a concentration of 10× MIC demonstrated a bactericidal effect. Bacteriostatic antimicrobial agents primarily inhibit the growth or reproduction of pathogenic microorganisms, leading to the arrest of cell growth, while bactericidal agents induce cell death. 35
Reduction in CFUs of enterohemorrhagic Escherichia coli over time after treatment with varying concentrations of hibiscus acid (1.25–50 mg/mL). Data are represented as the mean ± standard deviation of three independent experiments. Values with different letters expressed significant differences over time at α = 0.05 using the Tukey test. CFUs, colony-forming units; EDTA, ethylenediaminetetraacetic acid; MBC, minimum bactericidal concentration; MIC, minimum inhibitory concentration; MSIC, minimum subinhibitory concentration.
Reduction in CFUs of Salmonella Typhimurium over time after treatment with varying concentrations of hibiscus acid (1.75–70 mg/mL). Data are represented as the mean ± standard deviation of three independent experiments. Values with different letters express significant differences over time at α = 0.05 by the Tukey test. 10× MIC, 10× minimum inhibitory concentration.
Reduction in CFUs of Listeria monocytogenes over time after treatment with varying concentrations of hibiscus acid (0.625–25 mg/mL). Data are represented as the mean ± standard deviation of three independent experiments. Values with different letters express significant differences over time at α = 0.05 by the Tukey test.
The effect of compounds obtained from plants on the viability and development of Salmonella and/or E. coli strains has been reported. For example, Khan et al. 32 reported that carvacrol (MIC: 450 mg/mL) obtained commercially decreased E. coli growth significantly independent of the time after exposure, which decreased dramatically to less than 2 log10 CFU/mL after 60 min. Compared with this study, hibiscus acid managed to reduce growth in less time and at a lower concentration than that used with carvacrol.
Chen et al. 36 evaluated the bactericidal kinetics of myristic acid in pasteurized milk containing ¼× MIC (16 µg/mL), ½× MIC (32 µg/mL), and 1× MIC (64 µg/mL) of the acid, with an initial bacterial inoculum of 1 × 105 CFU/mL. The killing curves indicate that myristic acid significantly inhibited the growth of L. monocytogenes in pasteurized milk. The bacteria grew rapidly from 3.5 to 5.6 log10 in milk during storage under refrigeration without myristic acid, and when the bacteria were exposed to myristic acid with ¼× MIC (16 µg/mL), the growth of L. monocytogenes decreased.
Changes in cell motility
The cell motility of EHEC, S. Typhimurium, and L. monocytogenes, was observed at different hibiscus acid concentrations (0, MSIC, MIC, MBC, and 10× MIC) in a phase contrast microscope. Hibiscus acid at 10× MIC, MIC, and MBC concentrations inhibited the motility of EHEC, S. Typhimurium, and L. monocytogenes in TSB + YE. Hibiscus acid at MIC, MBC, and 10× MIC concentrations resulted in an immediate reduction in the motility of the three pathogenic bacteria examined. Treatment with hibiscus acid at MSIC did not have a notable effect on the motility of any of the microorganisms. The experiments were repeated three times. EDTA, used as a positive control, caused an immediate reduction in the motility of the three pathogenic bacteria.
Gill and Holley 37 observed changes in the motility of E. coli and L. monocytogenes when treated with eugenol, carvacrol, or cinnamaldehyde. Eugenol and carvacrol (10 mM) resulted in an immediate decrease in motile behavior.
Recently, Sedillo-Torres et al. 19 reported that hibiscus acid isolated and purified from H. sabdariffa inhibited the growth of S. Typhimurium when concentrations of 0.5 and 3 mg/disc were tested. These authors concluded that hibiscus acid is responsible for inhibiting both bacterial growth and motility. In our study, the inhibition of the motility of the three pathogenic bacteria in the presence of hibiscus acid (at MIC, MBC, and 10× MIC concentrations) was probably due to the direct inhibition of the flagellar motor by the dissipation of the proton gradient of the membrane, which is closely related to the functionality of the flagellum. 18 Hibiscus acid may influence cell motility by targeting various cellular processes, such as the expression of motility-related proteins, the regulation of signaling pathways, the inhibition of biofilm formation, and the downregulation of genes associated with motility. 38
Scanning electron microscopy
SEM offers valuable information regarding how antibacterial agents influence the external morphology and surface features of bacterial cells. 39 The effect of hibiscus acid on EHEC and L. monocytogenes cells at concentrations of MIC and 10× MIC, and in the control without hibiscus acid, can be observed in Figure 4. The bacterial cells that were not treated with hibiscus acid were intact (Fig. 4a1 and 4b1 for EHEC and L. monocytogenes, respectively). However, when the cells were treated with hibiscus acid at both MIC concentrations (Fig. 4a2 and 4b2) and 10× MIC (Fig. 4a3 and 4b3), cells with irregular morphology and forming conglomerates were observed, which could be related to membrane damage and cell membrane lysis (white arrows), which was not observed in cells treated with the negative control (Fig. 4). According to the SEM analysis, hibiscus acid may interact with the lipid bilayer of bacterial cells and induce the rupture of the cell membrane.

Micrographs of enterohemorrhagic Escherichia coli and Listeria monocytogenes obtained with SEM.
Devi et al. 27 observed that eugenol (1%, v/v) caused deformations of the surface of Salmonella Typhi cells, as well as rupture of the cell membrane, causing a complete loss of cellular integrity.
In our study, SEM examination showed that cells treated with hibiscus acid presented a rough surface and obvious cell shrinkage compared with untreated cells. The loss of membrane integrity and damaged cell surface show that one of the mechanisms of action of hibiscus acid against EHEC and L. monocytogenes is through damage and/or alteration of the cell membrane. Notable external ultrastructural changes were evident in the SEM images of treated cells compared with controls, with many cells appearing rough, twisted, wrinkled, and misshapen, some exhibiting invaginations and others showing protrusions on their surfaces. 39
Fluorescence microscopy staining
The presence of live and dead bacterial cells after hibiscus acid treatment was evaluated with AO/EB staining. Using this stain, live cells appear fluorescent green (AO), and dead cells or those that have lost the integrity of the membrane appear orange (EB). Most of the E. coli cells not treated with hibiscus acid (control) showed green fluorescence related to viable or living cells, which was related to an intact cell wall structure (Supplementary Fig. S1a and Supplementary Data). In contrast, high red fluorescence was found in cells treated with hibiscus acid at both MIC and 10× MIC (Supplementary Fig. S1b, c and Supplementary Data); however, at 10× MIC, a greater number of red or dead E. coli cells were observed. Supplementary Fig. S1b (Supplementary Data) was evaluated with ImageJ, a software tool that quantifies live and dead cells, 40 revealing a total of 165 live cells (green) and 829 dead cells (red). This indicates that CMI can eliminate at least 83% of the total cell population. These results suggest that the treatment of EHEC with hibiscus acid leads to cell death and/or bacteriostatic effects, which is consistent with the results determined by MIC. This correlates with the findings from the SEM analysis, as the cytoplasmic membrane plays a crucial role in several cellular functions such as osmoregulation, respiration, transport, biosynthesis, and lipid synthesis. It is vital to preserve its integrity, as any damage can result in metabolic dysfunction and ultimately lead to bacterial death. 41
Hameed et al. 31 studied the effect of Nd-doped ZnO nanoparticles on the viability of E. coli strains using confocal laser microscopy in the presence of AO/BE staining. The dual staining results suggest that Nd-doped ZnO NP particles cause the death of E. coli and K. pneumoniae cells compared with untreated cells.
Leakage of intracellular material
In the present study, the presence of genetic material outside the cells of EHEC, S. Typhimurium, and L. monocytogenes was quantified after having been treated with different concentrations of hibiscus acid, relating it to an increase in absorbance at 260 nm. Hibiscus acid at all study concentrations (MSIC, MIC, MBC, 10× MIC) affected the integrity of the cell membrane of EHEC, S. Typhimurium, and L. monocytogenes, which led to the release of intracellular material and an increase in absorbance at 260 nm, but this was not observed in the controls (Figs. 5–7). The increase in absorbance at 260 nm depended on the hibiscus acid concentration. Thus, the highest absorbance values were recorded at the hibiscus acid concentration of 10× MIC and with EDTA for the three pathogenic strains studied. The results suggest that hibiscus acid at different concentrations causes alteration of the cytoplasmic membrane after 2 h of contact, since the release of nucleotides began to increase after this time. An increase in absorbance at 260 nm is indicative of leakage of intracellular nucleic acids (DNA and RNA) and consequently reflects a loss of membrane integrity.42,43
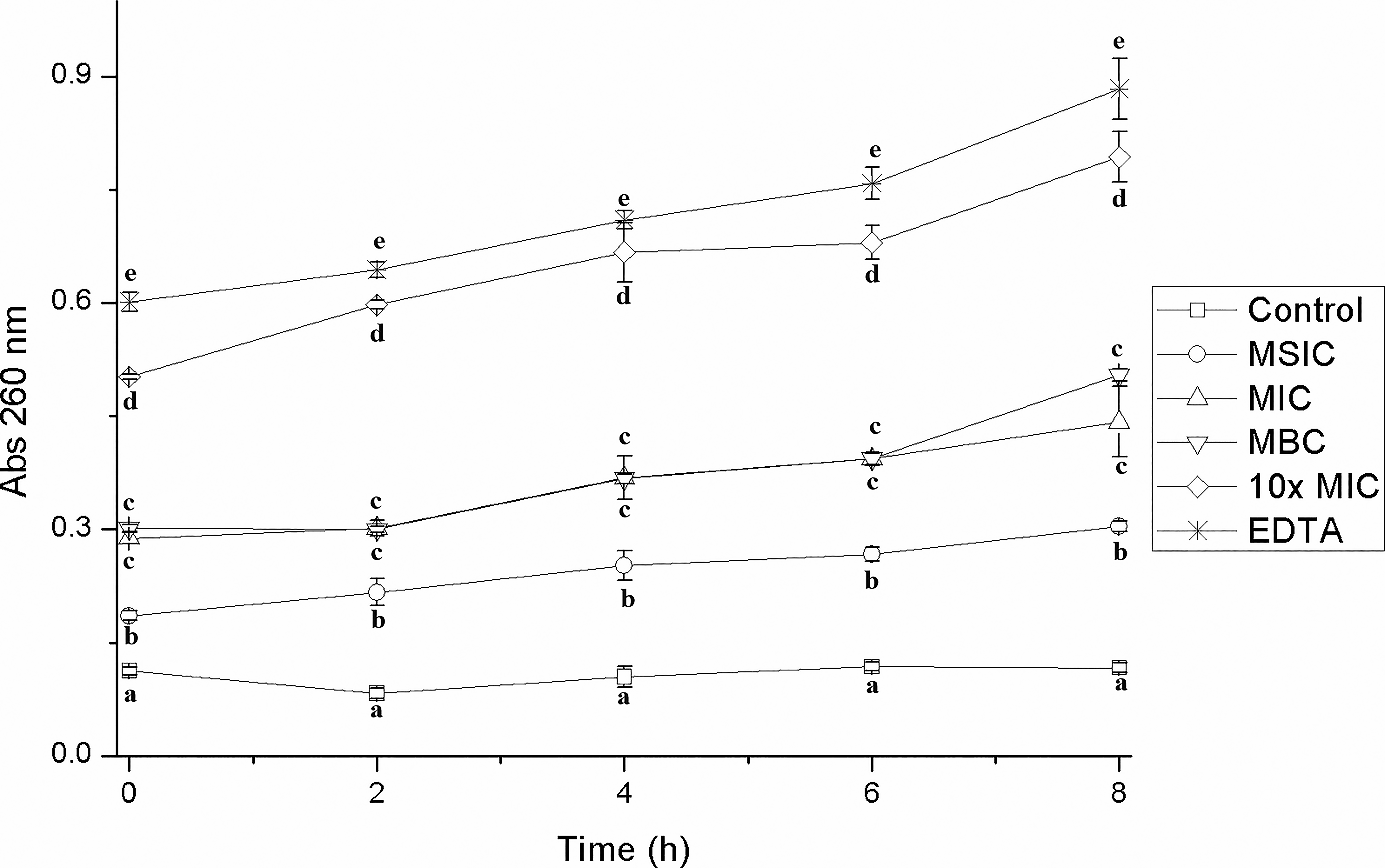
Leakage of intracellular material from enterohemorrhagic Escherichia coli cells previously treated with different concentrations of hibiscus acid (1.25–50 mg/mL) and with different contact times. Abs 260 nm: absorbance at 260 nm. Values with different letters for times express significant differences at α = 0.05 by the Tukey test.
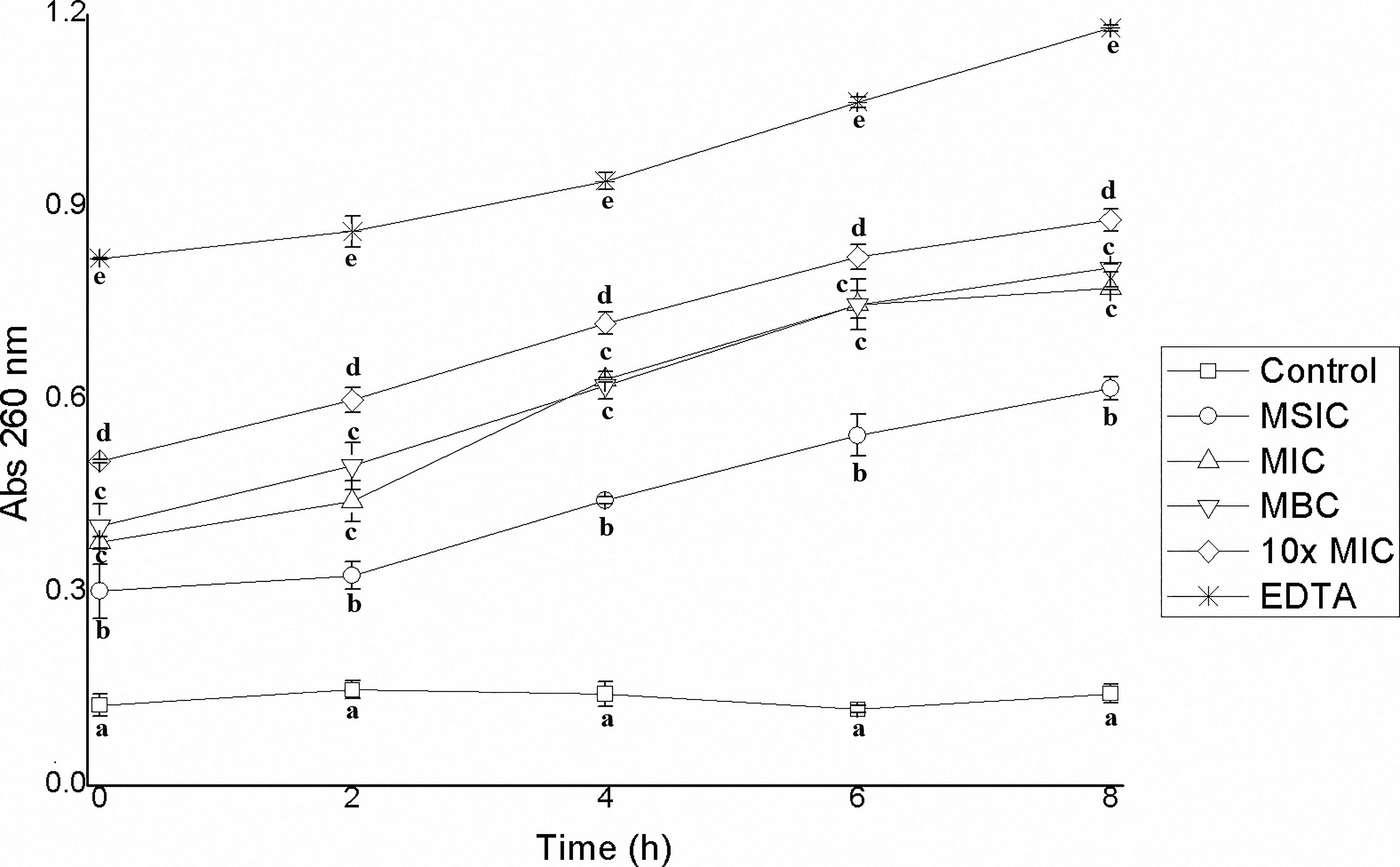
Leakage of intracellular material from Salmonella Typhimurium cells previously treated with different concentrations of hibiscus acid (1.75–70 mg/mL) and with different contact times. Abs 260 nm: absorbance at 260 nm. Values with different letters for times express significant differences at α = 0.05 by the Tukey test.
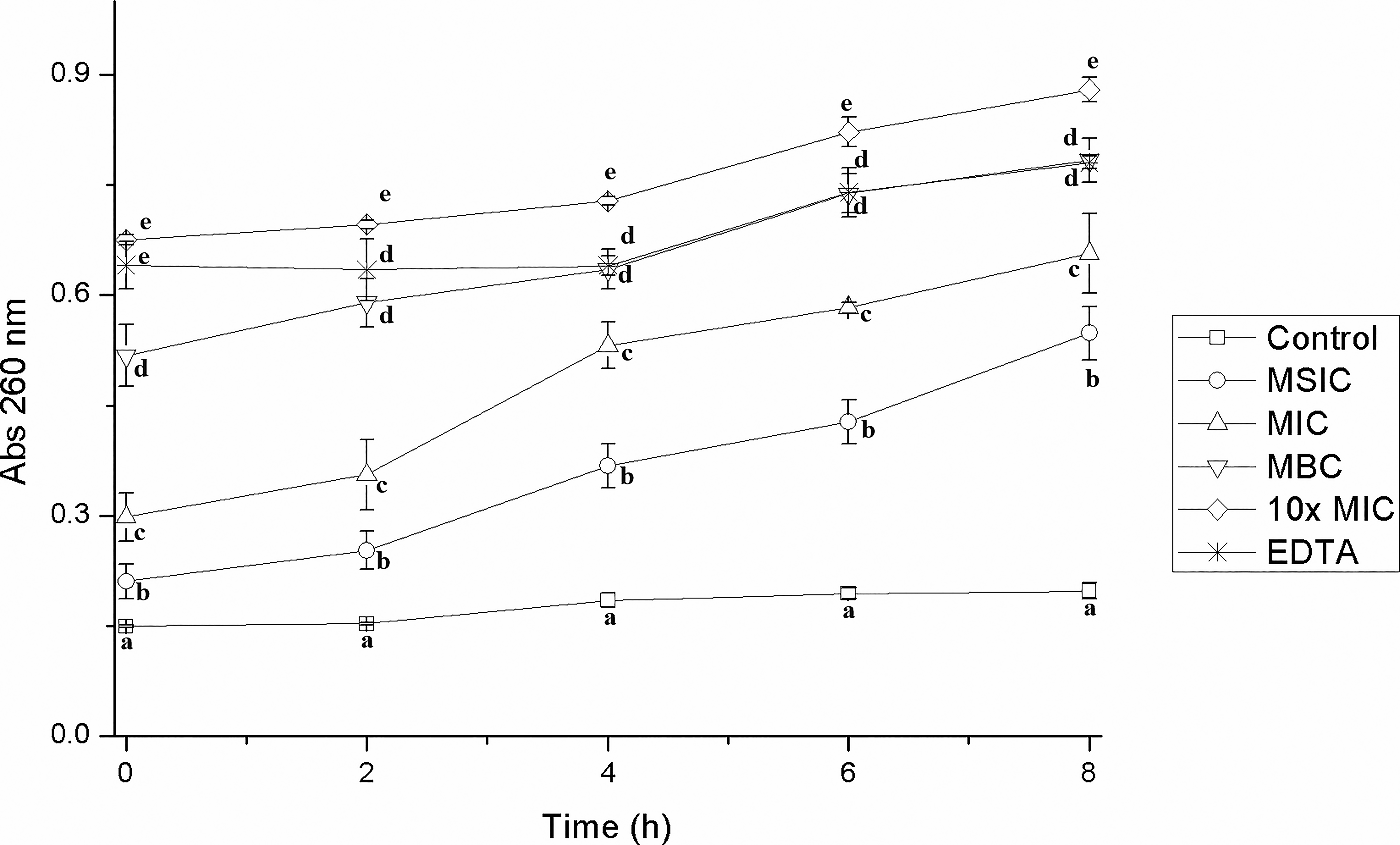
Leakage of intracellular material from Listeria monocytogenes cells previously treated with different concentrations of hibiscus acid (0.625–25 mg/mL) and with different contact times. Abs 260 nm: absorbance at 260 nm. Values with different letters express significant differences over time at α = 0.05 by the Tukey test.
Any effective antimicrobial compound must penetrate or disrupt the bacterial plasma membrane. In the present study, hibiscus acid modified the cell membrane of the three pathogenic bacteria studied, leading to the release of cellular contents, as observed based on absorbance values measured at 260 nm (Figs. 5–7). Hibiscus acid can alter the membrane, allowing for the leakage of intracellular constituents, while other compounds cause only a structural alteration of the external envelope.
The results from this study demonstrate that hibiscus acid exerts a dual mechanism of antimicrobial action against EHEC, S. Typhimurium, and L. monocytogenes, impacting both the external and internal components of bacterial cells.
External effects on the bacterial membrane
The SEM analysis revealed significant morphological changes in bacteria treated with hibiscus acid at MIC and 10× MIC concentrations, including membrane lysis, irregular surface features, and cellular shrinkage. These structural alterations point to direct damage to the bacterial cell membrane, a common target for plant-derived antimicrobial agents. 27 Membrane disruption was further evidenced by the leakage of intracellular contents, as observed through increased absorbance at 260 nm. This suggests that the integrity of the lipid bilayer was compromised, which likely resulted in a loss of cellular osmoregulatory functions, leading to cell death. 27
Internal effects: Leakage of Genetic Material
The increase in nucleotide leakage (DNA and RNA) in the bacterial cells treated with hibiscus acid indicates that the compound not only damages the membrane but also induces internal cellular disruption. Fluorescence microscopy staining confirmed the death of bacterial cells, as indicated by the predominance of red fluorescence in treated cells, especially at 10× MIC concentrations. This suggests a loss of membrane integrity and a subsequent bactericidal effect, consistent with other plant-derived antimicrobials that disrupt both cell walls and internal cellular processes. 27 Furthermore, the significant reduction in viable cells within the first few hours of treatment underscores hibiscus acid’s effectiveness in causing rapid intracellular damage.
The modification of bacterial membranes significantly influences DNA synthesis, particularly in the context of antimicrobial activity. Various compounds can compromise the integrity of the bacterial membrane, which in turn affects DNA synthesis. Certain plant-derived antimicrobial agents can disrupt the membrane, leading to alterations in cellular metabolism and inhibiting protein synthesis—an essential process for DNA replication and repair. 44 Additionally, compounds such as allicin can hinder RNA formation and partially inhibit DNA synthesis, highlighting the critical relationship between membrane integrity and nucleic acid synthesis in bacterial cells. 45
On the contrary, changes to the membrane can affect its permeability and fluidity, subsequently influencing the function of membrane-associated enzymes. For example, when antimicrobial compounds compromise membrane integrity, they can inhibit critical enzymatic processes, leading to metabolic dysfunction and potentially resulting in bacterial cell death. Additionally, the interaction of antimicrobial agents with the membrane can alter its lipid composition, further affecting the activity of enzymes involved in various metabolic pathways. 46
Finally, changes in membrane permeability and fluidity can alter the signaling mechanisms that regulate gene expression. Antimicrobial compounds compromise membrane integrity, which can lead to changes in the expression of virulence factors and other essential genes for bacterial survival and adaptation. Moreover, alterations in membrane properties can affect quorum-sensing mechanisms, which are vital for bacterial communication and biofilm formation. This can inhibit biofilm growth and reduce the expression of genes linked to pathogenicity. Additionally, interactions between phytochemicals and bacterial membranes can induce time-dependent changes in membrane morphology, suggesting an influence on the signaling pathways that regulate cellular responses. 47
Dual mechanism of action
These findings align with previous studies that suggest plant-derived compounds often operate via a combination of membrane disruption and intracellular interference. In the case of hibiscus acid, the primary action appears to be membrane damage, which leads to the release of internal components and eventual cell death. In the case of hibiscus acid, the primary action appears to be membrane damage, which leads to the release of internal components and eventual cell death. In our study, hibiscus acid demonstrated broad-spectrum bactericidal activity against EHEC, S. Typhimurium, and L. monocytogenes. Its mechanism of action involves significant disruption of the bacterial cell membrane, resulting in rapid reductions in cell viability, impaired motility, and leakage of intracellular contents. Furthermore, the internal effects observed—such as nucleotide leakage and loss of cellular viability—indicate that hibiscus acid not only damages the membrane but also interferes with intracellular processes essential for bacterial survival.32,36
When compared with other well-known natural antimicrobials, such as carvacrol, eugenol, and myristic acid, several important points emerge. Carvacrol and eugenol are often effective at lower concentrations,27,32,37 with MIC values typically lower than those observed for hibiscus acid. However, hibiscus acid exhibits a notably rapid bactericidal effect, leading to faster bacterial reduction times—a significant advantage in applications requiring swift microbial inactivation. In contrast, although myristic acid also possesses antimicrobial properties, 36 it generally requires even higher concentrations than hibiscus acid to achieve similar bactericidal effects, thereby limiting its practical use.
Overall, the dual antimicrobial mechanism of hibiscus acid—disrupting both the bacterial membrane and intracellular components—positions it as a promising alternative to conventional antibiotics for controlling pathogenic bacteria. Its robust performance across multiple pathogens, combined with its rapid bactericidal action, supports further exploration and optimization of hibiscus acid-based formulations. Future research should aim to assess its toxicity profile, explore potential synergistic effects with other antimicrobials, and identify its specific intracellular targets to fully understand its spectrum of activity.
CONCLUSIONS
This study demonstrates that hibiscus acid exhibits a potent antimicrobial effect against EHEC, S. Typhimurium, and L. monocytogenes by disrupting bacterial membrane integrity and inhibiting motility. The compound exerts bacteriostatic activity at MIC and MBC but achieves a bactericidal effect at higher concentrations (10× MIC). The antimicrobial mechanism is primarily associated with membrane permeability disruption, leading to the release of intracellular content, as observed in nucleotide leakage assays. SEM revealed notable damage to the bacterial cell membrane, while fluorescence microscopy confirmed cell death through membrane lysis.
The findings suggest that hibiscus acid can target multiple cellular processes, such as motility and membrane integrity, disrupting essential bacterial functions. This action makes it a promising natural antimicrobial agent with the potential to control multidrug-resistant foodborne pathogens. Future research should explore the intracellular effects of hibiscus acid to further elucidate its mechanism of action and validate its application in food safety and infectious disease control.
Additionally, in vivo studies or synergy experiments with sublethal antibiotic doses should be conducted to assess hibiscus acid’s potential in combination therapy against multidrug-resistant pathogens. Such investigations could significantly contribute to global antibiotic resistance management strategies by providing alternative approaches to enhance the efficacy of existing antibiotics and reduce reliance on synthetic antimicrobial agents.
AUTHORS’ CONTRIBUTIONS
L.A.P.-T.: Review and editing, literature search, writing—original draft preparation, and methodology. A.B.-N.: Conceptualization and formal analysis. C.A.G.-A.: Software, literature search, and conceptualization. L.G.-C.: Literature search and review and editing. M.R.T.-V.: Conceptualization, literature search, and review and editing. J.C.-R.: Review and editing, literature search, writing—original draft preparation, and methodology.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This work was supported by Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCyT) for financial support to the project number A1-S-8288 “Antibacterials from jamaica flower calyxes alone and in combination with antibiotics: determination of the mechanisms of action on resistant and non-resistant pathogenic bacteria to antibiotics, the antibacterial effect in vivo and adverse reactions in animals.”
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.