
Abstract
Mitochondrial dysfunction affects skeletal muscle (SkM) function and is critical in the etiology of age-related sarcopenia. The sirtuin 1-PGC1α pathway is a key regulator of mitochondrial mass, structure, and function. However, pathway activity decreases with aging. Cacao flavanols show promise in their ability to activate mitochondrial pathways. We evaluated the capacity of the flavanol (+)-epicatechin (+Epi) to stimulate such a pathway and favorably impact mitochondrial and oxidative stress (OS)-associated endpoints in aged SkM. Using 23-month-old male Sprague-Dawley rats, an 8-week oral administration of +Epi (1 mg/kg/day) was implemented, and results were compared versus vehicle-treated controls. Assessments included the nicotinamide adenine dinucleotide (NAD)/sirtuin 1/PGC1α pathway, acetylated proteins levels, mitochondrial function and biogenesis, as well as OS-related endpoints in SkM. +Epi increased the NAD/NADH ratio, activation of sirtuin 1, the deacetylation of nuclear protein content, including that of PGC1α. Also, +Epi induced increases of TFAM and NRF1 mRNA levels, deacetylation of mitochondrial complex I and V, increases in complex I activity, sirtuin 3, complexes I and V, mitofilin, and TFAM protein levels. SkM citrate synthase activity and ATP content increased with +Epi. OS markers in proteins and lipids were reduced, while buffering systems (superoxide dismutase 2 and catalase protein and activities) increased. In white blood cells, we documented serial reductions in mitochondrial DNA content and citrate synthase activity with aging, which were either fully or partially reversed with +Epi. Results demonstrate that +Epi treatment yields positive effects on mitochondrial biogenesis and function, leading to decreased OS and improved SkM bioenergetics in aged rats.
Keywords
INTRODUCTION
Aging is associated with a loss of skeletal muscle (SkM) mass (sarcopenia), leading to a progressive reduction in functional capacity. 1 It is well established that with aging, reductions in mitochondrial mass and function, and thus ATP generation underpins in many organs losses in function as seen in SkM and kidney. 2 Aging-associated mitochondrial dysfunction is also linked with the production of excess free oxygen radicals (i.e., oxidative stress; OS) and inflammation. 3 It is proposed that the sustained presence of such processes likely facilitates and/or promotes a progressive loss of SkM mass. 1 Conversely, to the extent that mitochondrial dysfunction can be prevented or reversed, a preservation of SkM mass and function is likely to occur. Currently, only exercise has been demonstrated to limit aging-associated loss of SkM, yielding significant reductions in functional decline. 4 However, in older individuals a sustained exercise regimen can prove to be difficult to achieve and/or sustain as per the presence of diseases such as osteoarthritis. Thus, there is great interest in identifying safe and effective alternatives for the targeting of sarcopenia.
The dysregulation of protein acetylation/deacetylation rates in mitochondria appears as an important cause for organelle dysfunction.5,6 With aging, protein acetylation levels increase, leading to perturbations in mitochondrial homeostasis.5,6 In cells, protein deacetylation is in part executed by sirtuins, which are nicotinamide adenine dinucleotide (NAD)+ dependent protein deacetylase enzymes. 6 While sirtuins can modulate mitochondrial function, they also influence the production of free radicals as per their ability to regulate buffering systems. 7 The protein sirtuin 1, which is located in the nucleus, is strongly expressed in SkM and plays a pivotal role in transcription factor deacetylation and of other key proteins. 7 Sirtuin-1 is involved in the modulation of multiple cellular processes, including metabolism, DNA repair, inflammation, and aging, 7 and as such has been identified as an important therapeutic target. Mitochondrial biogenesis is also recognized as being partly regulated in the nucleus by the sirtuin 1-PGC1α axis, primarily because the sirtuin deacetylates PGC1α and increases its transcriptional activity 8 leading to the stimulation of this process. Inside mitochondria, sirtuin 3 deacetylates enzymes and proteins in the citric acid cycle and electron transport chain, thereby promoting oxidative phosphorylation. 9 Another key role of sirtuin 3 is to deacetylate superoxide dismutase 2 (SOD2), enhancing its enzymatic activity. 10 Sirtuin enzymatic activity is dependent on adequate levels of its substrate NAD+.6 It has been documented that the deacetylation capacity of sirtuins decreases with aging due in part, to a diminished cellular NAD+ pool. 11 Several experimental approaches have been employed to sustain cellular NAD+ levels, and therefore, boost sirtuin deacetylation in order to ameliorate mitochondrial dysfunction, including the pharmacological inhibition of CD38 (one of the most NAD+-utilizing enzymes), caloric restriction, fasting, exercise training, and resveratrol treatment. 12 However, there is a limited amount of knowledge on the impact of targeted treatments on NAD+, sirtuins, and relevant protein activity status in aged SkM mitochondria-related endpoints.
We previously reported on the stimulatory effects of the flavanol (-)-epicatechin (Epi) on SkM mitochondrial biogenesis, function, and OS buffering systems in conditions such as aging and obesity or in the setting of diseases such as muscular dystrophy or Gulf War illness.13–16 We also recently reported on the capacity of the epimer +Epi to reverse SkM atrophy and capillary rarefaction in 25 month old male rats.17,18 Using samples from the same cohort of aged rats, we wished to evaluate the capacity of +Epi to improve mitochondrial indicators of organelle density and function as well as of OS status and associated buffer system-related endpoints. We also explored for underlying mechanisms associated with mitochondrial stimulatory effects with an emphasis on the regulation of the sirtuin 1-PGC1α axis, while blood samples were used to explore select relevant endpoints and the effects of treatment.
MATERIALS AND METHODS
Please see the Supplementary Data for detailed materials and methods.
RESULTS
Effects on NAD/NADH and deacetylation activity
Figure 1 denotes the levels of NAD+ (A), NADH (B), as well as the NAD+/NADH ratio (C) in SkM. Animals treated with +Epi demonstrated an ∼80% increase in NAD+, no difference in NADH, and therefore, an elevated (>100%) NAD+/NADH ratio vs. controls. Assessment of nuclear NAD+-dependent deacetylase sirtuin 1 activity (phosphorylation) was evaluated by Westerns after nuclear protein isolation from SkM (D). +Epi-treated animals demonstrated ∼60% higher levels of phosphorylation vs. controls. Nuclear protein acetylation levels were evaluated by Westerns using an anti-acetylated lysin antibody (E). Control animals demonstrated increased levels of total nuclear protein acetylation vs. +Epi-treated rats, which were reduced by ∼50%.
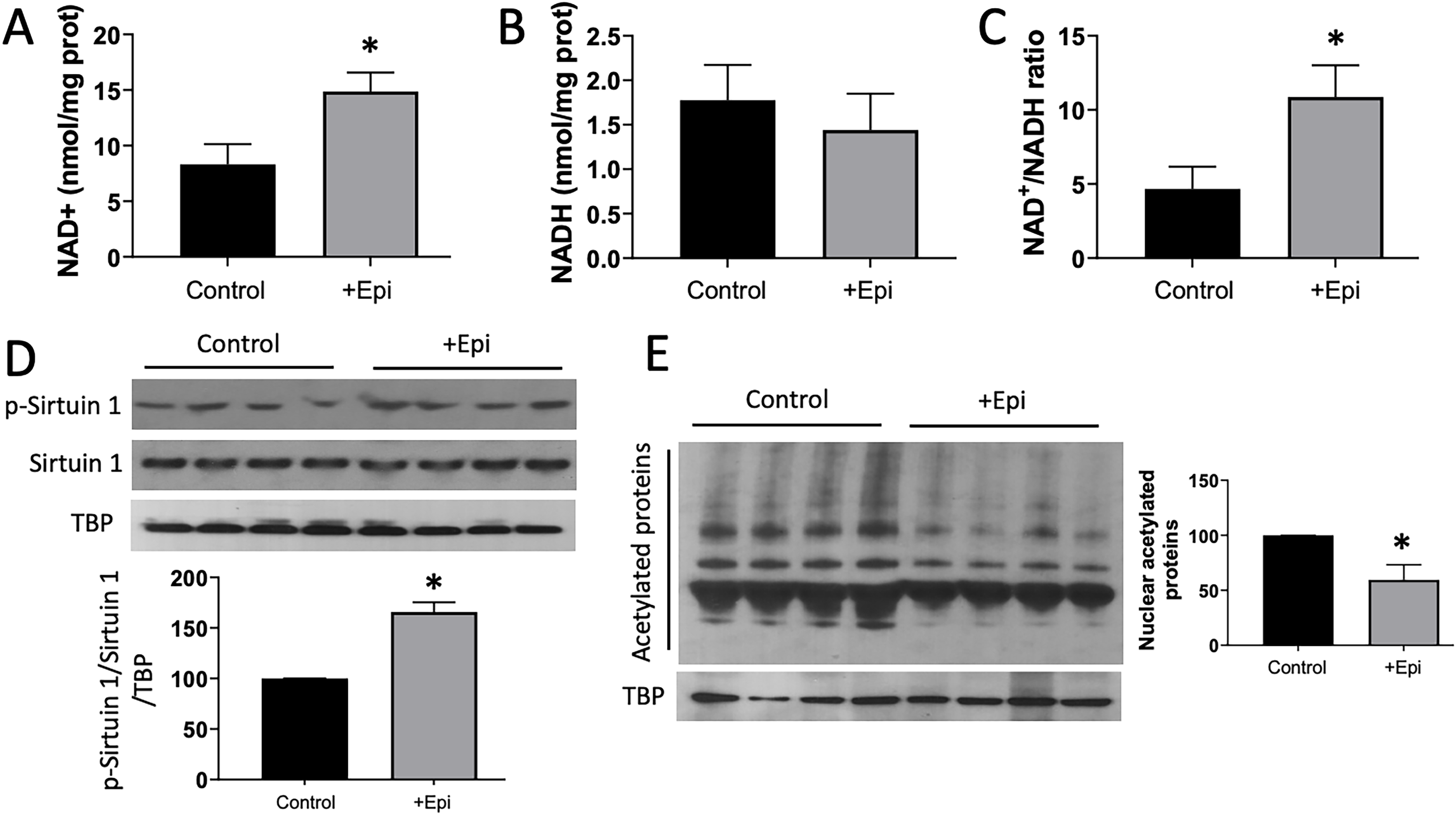
Effects of +Epi on aged gastrocnemius NAD+, nuclear SIRT1 activation and nuclear protein acetylation levels. (
Effects on regulators of mitochondrial biogenesis
Acetylated PGC1α was determined after nuclear protein isolation and lysin immunoprecipitation (Fig. 2A) and was quantified to report relative amounts using nuclear PGC1α and tata-binding protein (TBP) as nuclear and protein loading controls (B). Results demonstrate ∼70% decreased level of acetylated PGC1α in +Epi-treated animals vs. controls. Figures 2C and D report on the relative amount of TFAM and NRF1 mRNA levels. +Epi-treated animals demonstrated ∼50% increased amounts for both mRNAs. mtDNA copy number (E) shows a trend for increased levels in +Epi-treated animals.
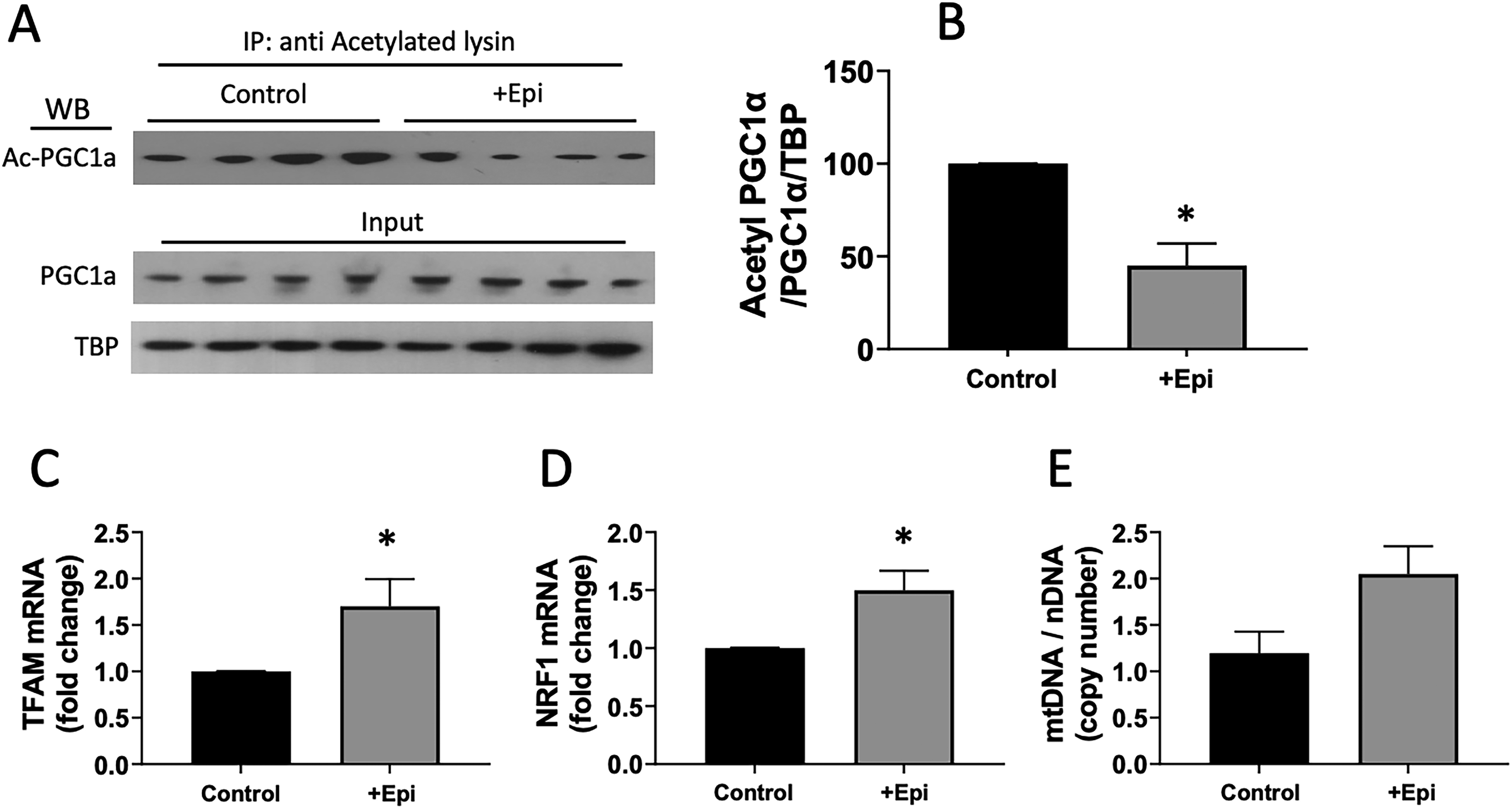
Effects of +Epi on aged on gastrocnemius acetylated PGC1α, TFAM, NRF1 mRNAs and mtDNA levels. (
Effects on mitochondrial structure and function
The NAD+-dependent deacetylase sirtuin 3 was evaluated in SkM by Westerns (Fig. 3). +Epi-treated animals demonstrated ∼35% increased amount of sirtuin 3 versus controls (A). Acetylated mitochondrial complexs I and V were determined after lysin immunoprecipitation (B). +Epi-treated animals demonstrated ∼50% decreased amounts of acetylated complex I and acetylated complex V vs. controls. Mitochondrial complex I activity was evaluated (C) and +Epi increased it by ∼100%. Figure 4 reports on complex I and V, mitofilin, and TFAM relative protein levels as evaluated by Westerns. Results demonstrate ∼20–30% increased content for such proteins in SkM of +Epi treated animals vs. controls (A). Mitochondrial function surrogates citrate synthase activity (B) and ATP content (C) denoted increases of > 200% and ∼90%, respectively, with +Epi treatment.
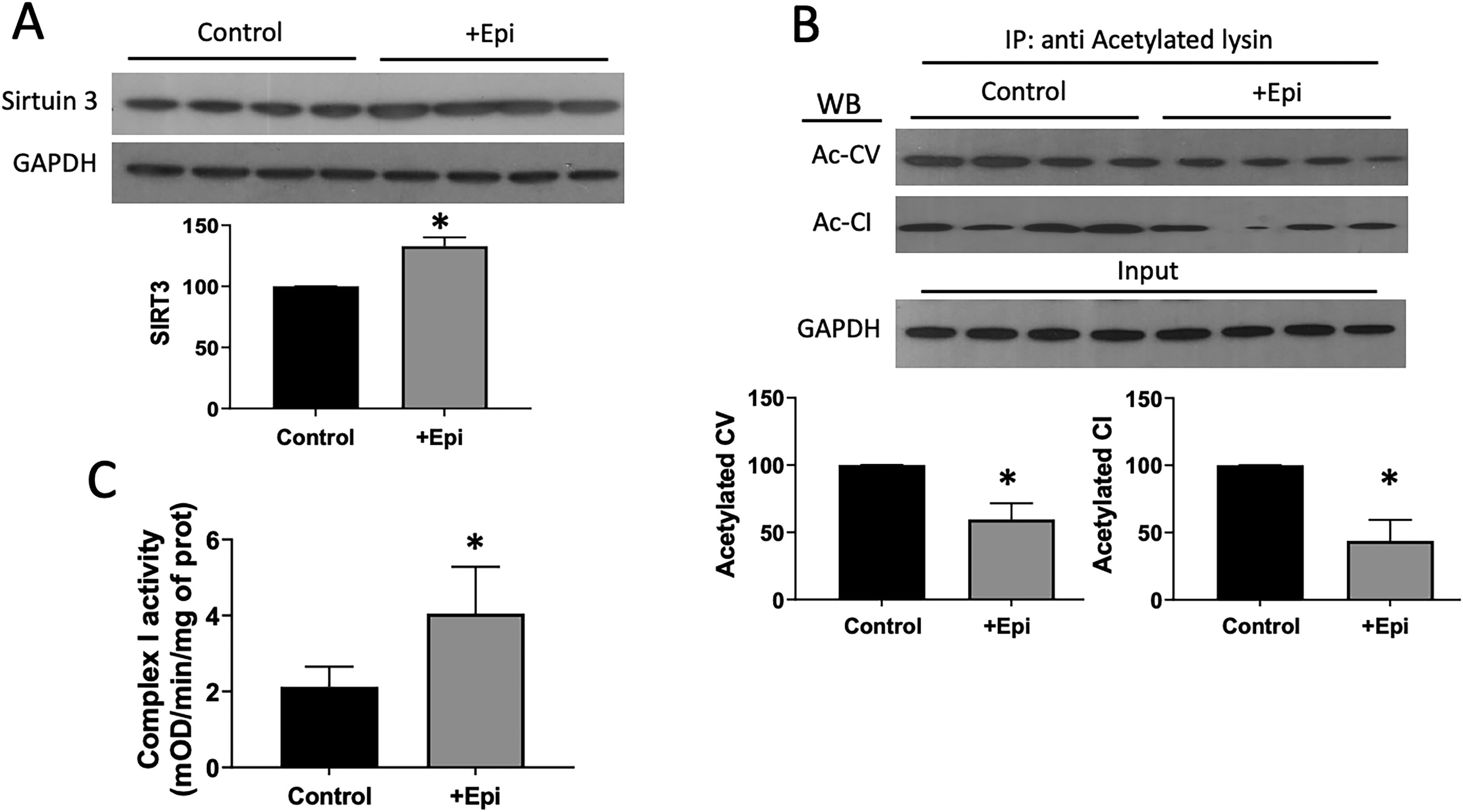
Effects of +Epi on aged gastrocnemius SITR3, acetylated mitochondrial complex I and V and complex I levels and/or activity.
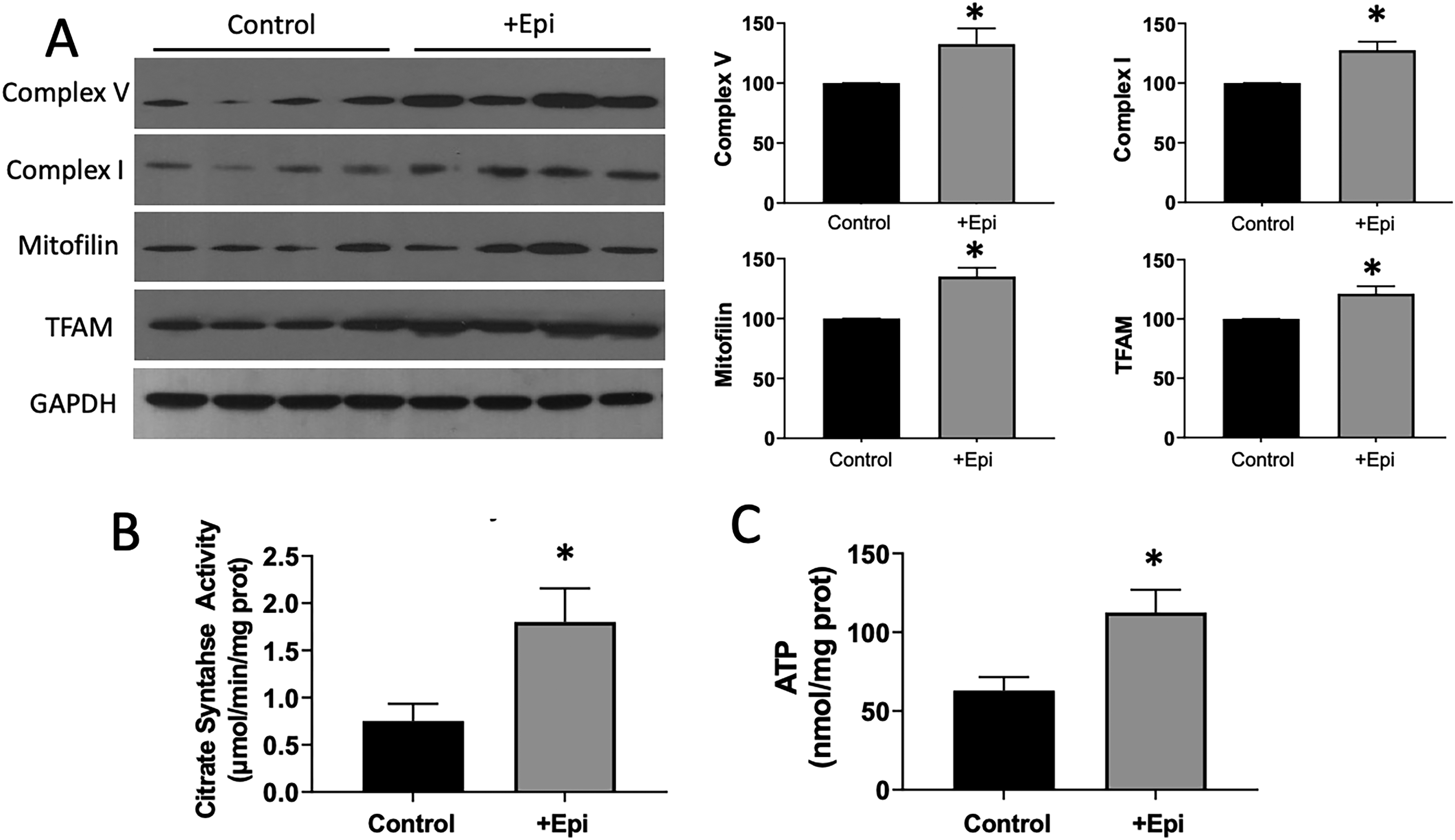
Effects of +Epi on aged gastrocnemius mitochondrial proteins and function.
Effects on OS and buffering systems
Figure 5 denotes the assessment of OS-related endpoints in plasma and gastrocnemius of aged rats. Panel A illustrates a time course analysis of protein carbonylation in plasma of rats at 6, 12, 18, 23, and 25 months of age. From the 6 to 25-month time point, plasma carbonyl protein content was consistently increased to more than double over time. However, 8 weeks of +Epi treatment in rats (from 23 to 25 month) yielded significant decreases of ∼40% for this endpoint at the end of treatment. Panel B illustrates protein carbonyl content, while panel C denotes malondialdehyde levels in gastrocnemius. +Epi-treated animals demonstrated ∼60% decreased content of both OS markers in gastrocnemius vs. controls. Panel D reports on total levels of oxidized proteins analyzed by Westerns. +Epi-treated animals demonstrated ∼50% decreased oxidized protein content in SkM versus controls. Figure 6 denotes antioxidant enzyme assessment in gastrocnemius of aged rats. Panel A illustrates SOD2 and catalase content evaluated by Westerns. +Epi-treated animals demonstrated ∼100% increased content of both antioxidant enzymes in gastrocnemius vs. controls. Panels B and C illustrate SOD2 and catalase enzyme activity in gastrocnemius. +Epi-treated animals demonstrated increased activity of ∼60% and ∼100%, respectively, versus controls.
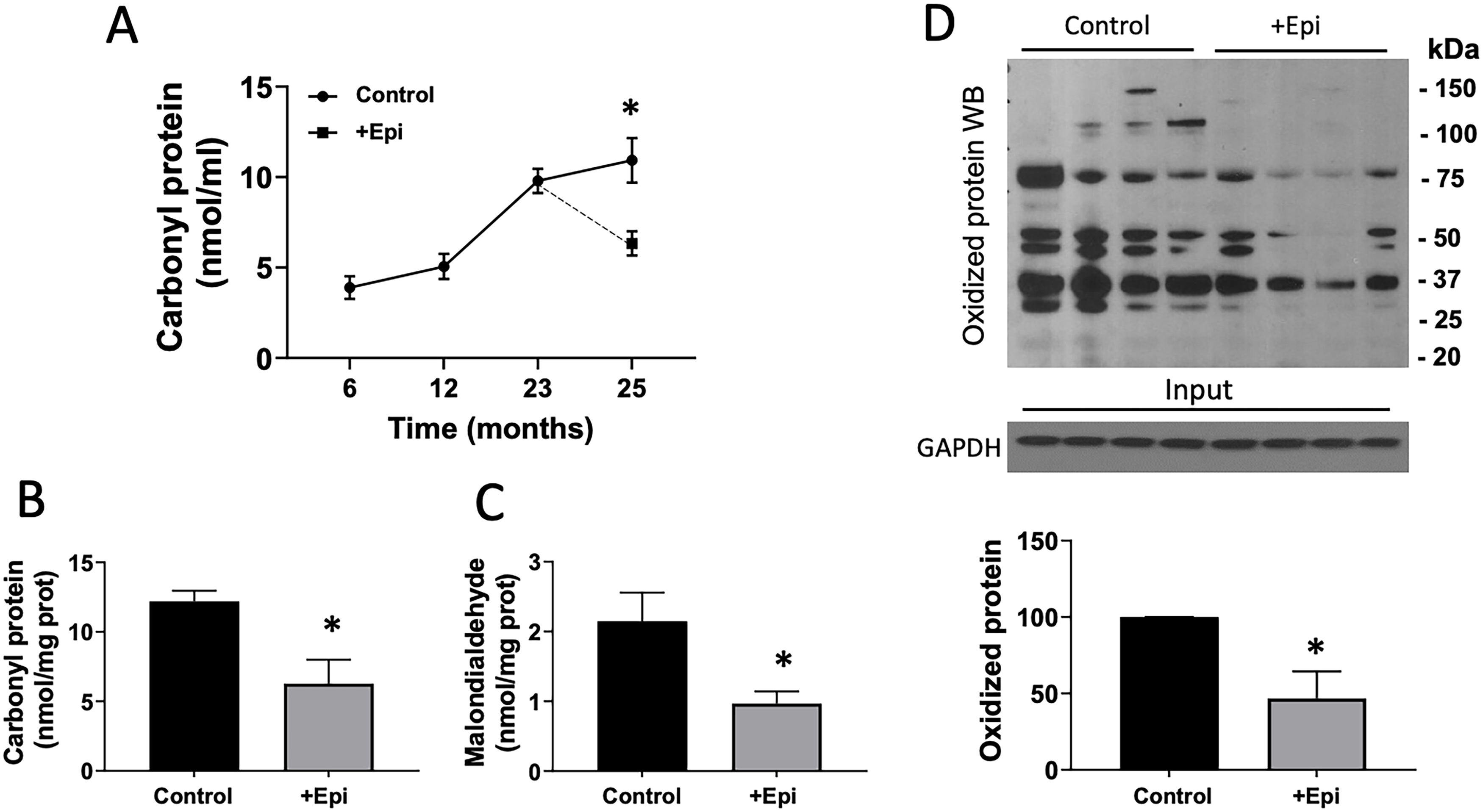
Effects of +Epi on oxidative stress markers in plasma and gastrocnemius of aged rats.
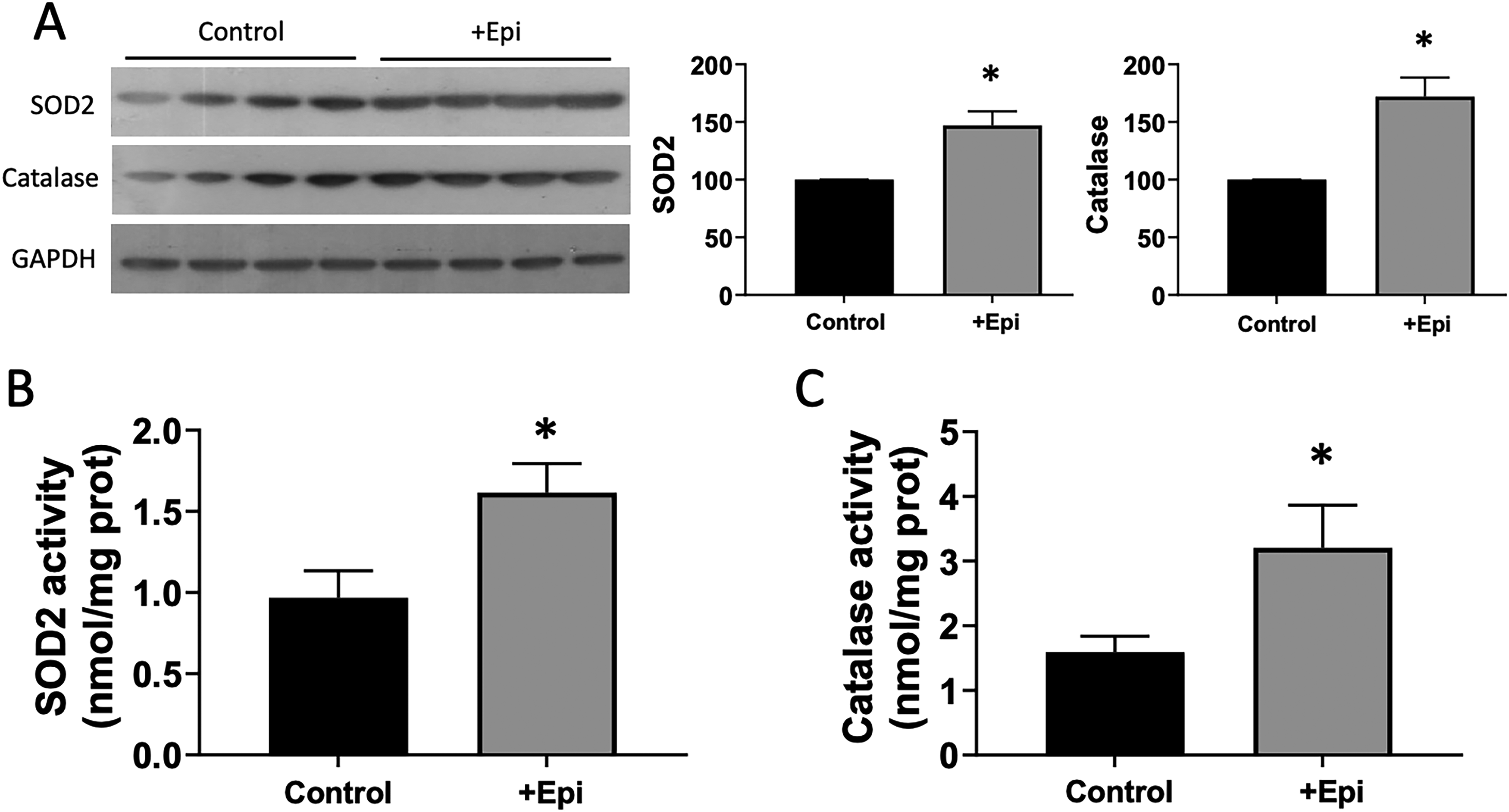
Effects of +Epi on antioxidant buffering systems of aged muscle.
Effects on mitochondria white blood cell endpoints
Figure 7 denotes the serial assessment of mitochondrial DNA copy number (mtDNA) and citrate synthase activity in white blood cells collected at 6, 12, 18, 23, and 25 months of age. From the 6 to 12-month time point, mtDNA content was unchanged (A). However, from 12 to 25 months of age, mtDNA decreased over time (A). The 8 weeks of +Epi intervention in rats at the 23-month time point yielded a trend for increases in mtDNA until the end of treatment (25 months of age). Panel B illustrates a time course analysis of citrate synthase in white blood cells of rats at 6, 12, 18, 23, and 25 months of age. From the 6 to 25-month time point, citrate synthase activity decreased by ∼60% over time. However, 8 weeks of +Epi treatment yielded a significant ∼100% increase in activity by 25 months of age.
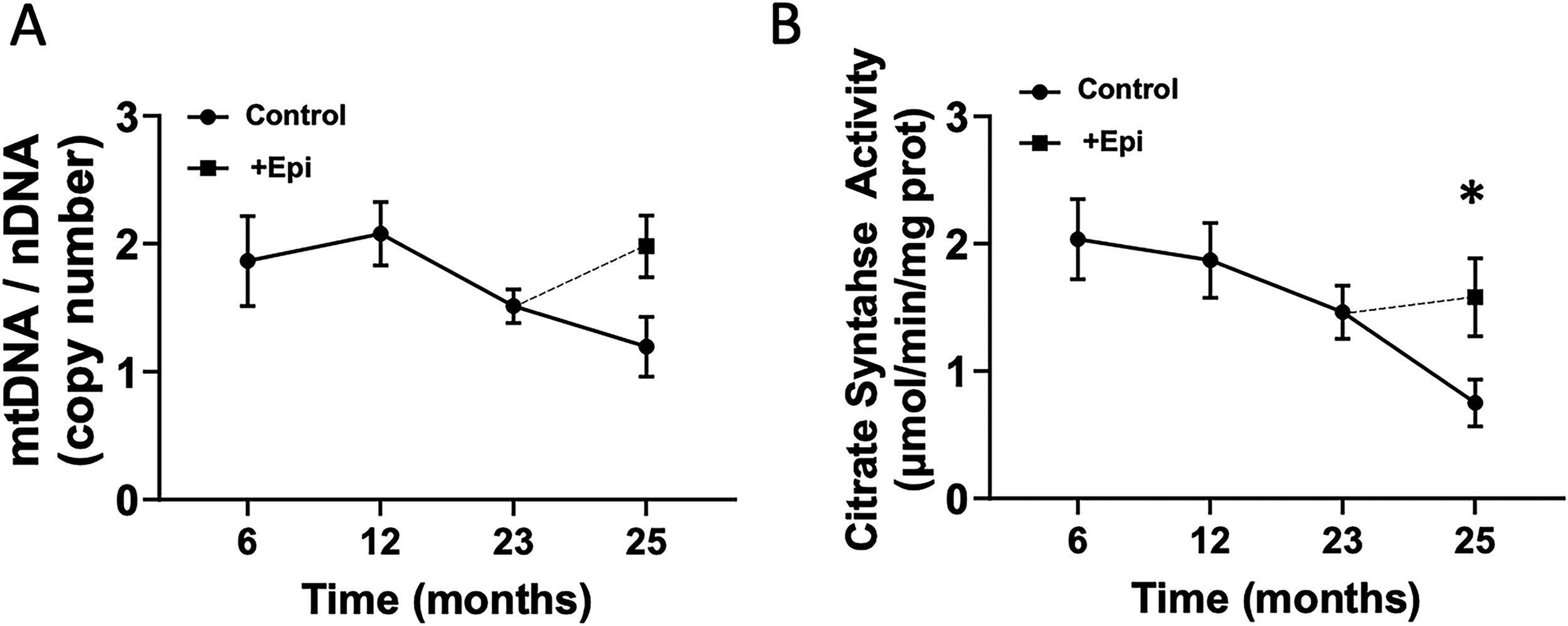
Effects of +Epi on white blood cell mitochondrial DNA and citrate synthase activity of aged rats.
DISCUSSION
Unique results from this study demonstrate that in a rat model of aging-induced sarcopenia, +Epi increases the SkM NAD/NADH ratio, sirtuin 1 activity, and consequently, the deacetylation of nuclear proteins, including PGC1α. Effects are accompanied by increases in TFAM and NRF1 mRNA levels, increases in sirtuin 3, complex I and V, mitofilin and TFAM protein levels, deacetylation of mitochondrial complexes I and V, and increased complex I activity. SkM citrate synthase activity and ATP content increase. OS was reduced while buffering system protein levels and activities increased. In white blood cells, we also documented serial reductions in mtDNA content and citrate synthase activity with aging, which were partially or fully reversed with +Epi. Altogether, the concerted activity of such biological actions suggests that part of the beneficial effects of +Epi on sarcopenia relates to the favorable activation of mitochondria-related axis.
Aging-associated conditions and/or diseases have become a great burden to health care systems, and for many of these entities, therapeutic options remain limited. Sarcopenia in particular, has gained high levels of attention as per its close relationship with frailty, falls, and consequently, higher mortality. 19 While exercise represents a useful tool to counter sarcopenia, its widespread use is limited by conditions such as osteoarthritis and obesity or access to suitable training facilities. 20 Thus, the high level of interest in identifying candidate agents that target sarcopenia. In aging populations, agents that can be provided orally and carry exceptional safety profiles would be preferred. Flavonoids are polyphenolic compounds found ubiquitously in a wide range of plants, fruits, and vegetables. 21 Many of the positive health effects derived from the consumption of such products are attributed to the presence of flavonoids. Evidence is derived from epidemiological studies that associate that regular consumption of products rich in flavonoids with decreased cardiometabolic and other chronic disease risk.22–24 Thus, the interest in identifying and isolating such agents so as to explore their pharmacological properties, therapeutic potential, and mechanisms of action. 21
Over the course of several years, a large body of related literature has been generated whereby specific flavonoid isolates have been examined for their potential to exert positive effects on various systems, conditions, and/or diseases. 21 We recently published a review and meta-analysis on preclinical and clinical studies reporting on the beneficial effects of specific polyphenolic compounds or flavonoids on SkM health. 25 It is readily apparent that compounds such as quercetin and green tea catechins have the capacity to favorably impact SkM health, including mass and function. However, there is a lack of a systematic analysis for specific flavonoids, and there can be large discrepancies on key issues such as the doses used, types of outcomes reported, and clinical applicability.
Cacao contains high amounts of the flavonoid Epi, and the health-promoting effects of the moderate consumption of cocoa products are linked to the actions of this compound. Kuna Indians who regularly consume a cacao-based beverage are protected against age-dependent reductions in vascular and renal function 26 and demonstrate extremely low mortality indices for chronic diseases. 27 In support of these observations, clinical studies have demonstrated reductions in blood and organ inflammation and OS levels with cocoa/dark chocolate consumption. 28 We have previously explored the potential of +Epi to exert stimulatory effects under controlled conditions on cultured SkM cells with promising results. 29 Furthermore, in humans and animal models, we examined the pharmacokinetic and partial pharmacodynamic properties of +Epi, with recent results suggesting a higher efficacy profile vs. the epimer -Epi. 30 Using SkM samples obtained from the same-aged animals used in this study, we previously reported that treatment with +Epi led to significant increases in front paw grip strength, treadmill time, and muscle mass vs. controls. 17 Treatment also led to the activation of key intracellular pathways associated with the regulation of muscle mass. +Epi also significantly reversed adverse changes in plasma and/or SkM TNF-α, IGF-1, atrophy and protein synthesis related endpoints vs. controls. In a separate recent publication, using the same samples we also reported on the angiogenic effect of +Epi on SkM, whereby increases in capillary density were reported vs. controls an action, that was linked to the activation of the VEGF signaling pathway. 18 As a whole, stimulatory effects on angiogenesis and atrophy-related pathways appear to provide a coherent and integrative framework for how +Epi can effectively translate its effects into enhanced structure and function in the setting of aging-induced sarcopenia.
However, strong biological links have been reported between mitochondrial status and changes in muscle mass. In animal models, causal relationships have been reported whereby increased mitochondrial density via the forced expression of the organelles leads to the stimulation of myofiber formation. 31 In a study by Always et al., after the induction of SkM cryoinjury in mice, the transplantation of mitochondria led to an enhanced restoration of muscle mass and function. 32 Conversely, in conditions whereby a loss of mitochondria density, structure, and/or function takes place, muscle atrophy can ensue.33,34 In humans, studies have linked impaired mitochondrial structure, function, or density with sarcopenia, while positive effects on muscle mass have been linked to the beneficial effects of exercise on this organelle.4,35 Thus, mitochondria appear to play critical roles in determining the preservation of muscle mass and function as per it’s capacity to produce ATP and ameliorate OS levels. Despite age-associated mitochondrial dysfunction being extensively investigated, the precise mechanisms that drive such changes remain to be determined. Candidate mechanisms to drive such detrimental changes may include decreased mitochondrial biogenesis, function, and/or diminished mitochondrial DNA levels. 35
Accumulating evidence suggests that sirtuin 1 and PGC-1α play a key role as upstream activators of mitochondrial biogenesis. 8 Sirtuin 1-mediated deacetylation activates PGC-1α through a functional protein-protein interaction, and once PGC-1α is active, it works in concert with several transcription factors to stimulate the expression of a broad set of mitochondrial genes.36,37 These transcriptions factors include NRF-1 and -2 whose activation increases the expression of multiple mitochondrial proteins, including oxidative phosphorylation complexes and TFAM. 38 Reduced levels of PGC-1α have been found in aged animals, and loss of PGC-1α has been suggested to be an important contributor to mitochondrial dysfunction in aging-associated diseases. 39 As per the results reported in this study, +Epi activates the sirtuin 1 and PGC-1α mitochondrial biogenesis-related pathway, leading to enhanced levels of key mitochondrial proteins, which ultimately translate into improved bioenergetics given the noted increases in citrate synthase activity and SkM ATP levels.
Using aged mice, we previously reported that 2 weeks of -Epi treatment of aged (26-month-old) mice reduces SkM OS levels as determined by protein-free carbonyl content while restoring the buffering system levels toward those noted for young animals. 14 Using a high-fat diet mouse model of obesity and insulin resistance, we also previously reported increases in SkM protein carbonylation and nitrotyrosilation, as well as a decrease in SOD2 and catalase activity. 40 -Epi treatment reversed such changes toward those of normal mouse muscle. Thus, Epi has the capacity to effectively modulate ROS buffering systems in aged or diseased SkM that translate into reduced levels of OS. The deacetylase sirtuin 3 which is primarily located in mitochondria, is known to play an important role on the regulation of ROS buffering systems. A major target of sirtuin 3 deacetylation is SOD2. 10 In this study, +Epi treatment increased sirtuin 3, SOD2, and catalase protein levels in aged SkM, documenting the manner by which +Epi led to reduced OS levels.
With the intent of documenting broader effects of +Epi beyond those noted for SkM, we also evaluated changes in PBMC mtDNA copy number and citrate synthase activity. The analysis of samples collected at 6, 12, and 23 months indicates an age-dependent decrease in such endpoints, which were apparently either fully restored (mtDNA) or partially reversed (citrate synthase activity) with 1 month of treatment with +Epi. Thus, as we have documented in our previous reports, Epi has the capacity to positively impact mitochondria-related system in a broad manner and not act in a tissue-selective manner. This property is seen as advantageous as per the adverse impact that aging has on SkM and other systems such as the vasculature.
CONCLUSIONS
In conclusion, +Epi has the capacity to positively impact on pathways that activate key regulatory systems in mitochondria and ROS buffering systems. Such effects translate into improved SkM bioenergetics and reduced levels of OS. As per the well-documented safety profile of Epi and favorable pharmacokinetics, clinical trials are warranted to explore its potential to be used as an agent that favorably impacts either in a preventive or therapeutic manner, sarcopenia.
AUTHORS’ CONTRIBUTIONS
I.R.-S.: Investigation, data curation, writing—original draft preparation, reviewing, editing. R.O.-R.: Investigation, data curation, writing—original draft preparation. V.N.: Investigation, data curation, writing—original draft preparation. G.C.: Supervision, data curation, formal analysis, original draft preparation, reviewing, editing. F.V.: Conceptualization, funding acquisition, project administration, supervision, data curation, writing—original draft preparation, reviewing, editing.
Footnotes
AUTHOR DISCLOSURE STATEMENT
F.V. (co-founder) and G.C. are stockholders of Epirium Bio Inc.
FUNDING INFORMATION
This study was partly supported by DoD GW180044 and VA I01BX3230, I01BX5894 to Dr. Villarreal and IPN 20195988 and CONACYT 283938 to Dr. Ramirez-Sanchez.