
Abstract
Osteoarthritis (OA) is characterized by inflammation-driven chondrocyte senescence and extracellular-matrix degradation. However, the molecular mechanisms linking inflammatory stress to chondrocyte aging remain poorly understood. Here, we identify cinnamyl alcohol (CA) as a natural small-molecule compound that attenuates OA progression through polymeric immunoglobulin receptor (PIGR)-mediated signaling in vitro. CA reduced inflammatory cytokine production, suppressed senescence-associated secretory phenotype gene expression, and preserved cartilage homeostasis in lipopolysaccharide- or interleukin-1β-stimulated chondrocytes. In a destabilization-of-the-medial-meniscus mouse model, intra-articular CA administration markedly alleviated cartilage degeneration and matrix loss. Integrating network pharmacology, molecular docking, and mass-spectrometry-based proteomic profiling, we identified PIGR as a convergent target of CA, validated by limited proteolysis (drug affinity responsive target stability) and loss-of-function assays. PIGR silencing abolished CA’s antisenescent and cartilage-protective effects, confirming its essential role. Mechanistically, CA restored PIGR expression to modulate inflammatory signaling and maintain chondrocyte phenotype stability. These findings uncover a previously unrecognized CA–PIGR axis that couples inflammatory stress to cartilage aging and suggest CA as a promising natural therapeutic candidate for OA management.
Keywords
INTRODUCTION
Osteoarthritis (OA) is a degenerative joint disease characterized by progressive cartilage degradation, synovial inflammation, and subchondral bone remodeling. 1 As the most common form of arthritis, OA affects millions of people worldwide, causing pain, disability, and significantly reducing quality of life. 2 The pathogenesis of OA is complex and multifactorial, involving mechanical, inflammatory, and metabolic factors. 3,4 Recent studies have highlighted the role of cellular senescence in OA progression, suggesting that targeting age-related changes in chondrocytes could be a promising therapeutic strategy. 5 –7
Aging chondrocytes have the capacity to produce senescence-associated secretory phenotype (SASP) components autonomously. This results in elevated levels of catabolic cytokines, including interleukin-6 (IL-6), chemokine (C-X-C motif) ligand 2 (CXCL2), and tumor necrosis factor-α (TNF-α). 8 These cytokines, in turn, stimulate the production of matrix-degrading enzymes, with matrix metallopeptidase 13 (MMP13) playing a pivotal role in the permanent breakdown of type II collagen (COL2) networks in osteoarthritic cartilage. 6,9,10 Moreover, SASP factors facilitate intercellular communication between senescent chondrocytes and their healthy counterparts, potentially inducing senescence in the latter. 11,12 Consequently, addressing chondrocyte senescence has emerged as a promising strategy for OA treatment. However, the complete picture of the underlying mechanisms driving chondrocyte senescence in OA remains to be fully elucidated.
Natural products have long been a rich source of bioactive compounds with potential therapeutic applications. 13 –15 Cinnamyl alcohol (CA) is a well-known component of cinnamon bark and cinnamon twigs, and it has anti-inflammatory, antibacterial, and vasodilatory properties. 16 –18 While CA has shown promise in various disease models, its potential for mitigating OA progression and chondrocyte senescence remains largely unexplored.
Inflammatory mediators, such as IL-1β and lipopolysaccharide (LPS), play crucial roles in OA pathogenesis by promoting cartilage degradation and chondrocyte dysfunction. 19 In vitro models using these inflammatory stimuli on chondrocytes have been widely used to investigate potential OA therapeutics. 20 In addition, the destabilization-of-the-medial-meniscus (DMM) model in mice has emerged as a reliable approach for studying OA progression and evaluating therapeutic interventions in vivo. 21
Recent advancements in network pharmacology have provided new tools for predicting drug targets and understanding complex mechanisms of action. 22 This approach has been successfully applied to natural products, offering insights into their molecular targets and potential therapeutic applications. 23 However, the specific molecular targets and mechanisms underlying CA’s potential effects on OA remain to be elucidated.
The polymeric immunoglobulin receptor (PIGR) is a transmembrane protein primarily known for its role in mucosal immunity. 24 PIGR is classically induced by pro-inflammatory cues that converge on Nuclear factor kappa B (NF-κB), including microbial products such as LPS (via TLR4) and cytokines such as TNF-α and IL-1β, through an NF-κB-responsive element in intron 1 of the human PIGR gene. 25 –27 These pathways have been delineated in epithelial systems and, more recently, linked to broader inflammatory circuitry (e.g., STAT3/IL-17). 28 However, whether PIGR modulates inflammation-driven senescence and extracellular matrix (ECM) homeostasis in articular chondrocytes has remained unknown. Here, by integrating network pharmacology and mass spectrometry (MS), docking, target engagement (drug affinity responsive target stability [DARTS]), and loss-of-function studies, we identify PIGR as a mechanistic node that couples LPS/IL-1β stress to chondrocyte senescence and cartilage catabolism, and we demonstrate that CA restores PIGR levels and requires PIGR to exert its antisenescence, pro-homeostatic effects.
MATERIALS AND METHODS
Mouse models and treatments
C57BL/6J mice were maintained in a conventional specific-pathogen-free setting, with a standard 12-h light–dark cycle and unrestricted access to food and water. The DMM model was surgically induced following established protocols. 29 In brief, mice were anesthetized, and the operative area was prepared by removing hair and applying an antiseptic. A medial parapatellar approach was used to expose the right knee joint, laterally dislocating the patella and fully flexing the knee. Using a microsurgical blade, the anterior medial meniscotibial ligament was severed. Successful transection was visually confirmed by gently manipulating the medial meniscus with delicate forceps. The joint space was irrigated with sterile saline, followed by suturing of the articular capsule using 6-0 resorbable polyglycolic acid thread. Skin closure was accomplished with 5-0 sutures. A control group underwent a sham procedure on the right knee, which involved a skin incision, medial capsulotomy, and subsequent closure of both capsule and skin, without ligament transection. All animal experimental protocols were performed in accordance with the Guidelines for the Care and Use of Laboratory Animals and were supervised by the Laboratory Animal Welfare and Ethics Committee of the Army Medical University.
Intra-articular injection of CA in mice
Establishment of the DMM model: Fifteen 12-week-old male C57BL/6 mice were randomly divided into three groups: (1) sham operation group: mice (n = 5) received sham operation; (2) DMM + low-dose group: mice (n = 5) received DMM operation and intra-articular injection of 10 mM CA (HY-Y0078, MedChemExpress, New Jersey, USA) twice a week for 8 weeks; (3) DMM + high-dose group: mice (n = 5) received DMM operation and intra-articular injection of 20 mM CA twice a week for 8 weeks.
Von Frey test
To evaluate mechanical allodynia, we utilized a set of von Frey filaments (NC12775-99, North Coast Medical Inc., CA, USA). The testing apparatus consisted of a wire mesh grid, upon which mice were placed for a 30 min acclimation period prior to the assessment.
The mechanical withdrawal threshold was determined by applying filaments of varying forces to the central plantar surface of the hind paws. A response was considered positive if the animal exhibited any of the following behaviors: rapid paw withdrawal, paw licking, or paw shaking.
To minimize experimental bias, all von Frey filament tests were conducted by a single investigator who was blinded to the treatment groups. This approach ensured consistency in technique and interpretation of responses across all subjects.
Histological examination
After CA treatment was completed, the knee joints underwent decalcification in a 10% w/v EDTA solution for 7 days, followed by paraffin embedding. Sagittal sections, each 5 μm thick, were cut serially and subjected to Safranin O/Fast Green staining for morphological assessment. The analysis focused on three distinct levels of sections through the knee’s medial compartment, with each level separated by 50 μm. To evaluate the intensity of the OA-like characteristics, two independent observers, blinded to the experimental conditions, employed the Osteoarthritis Research Society International (OARSI) scoring system as previously outlined in the literature. 29,30
Immunohistochemistry
For immunohistochemical analysis, tissue sections underwent deparaffinization and hydration, followed by enzymatic treatment. This process involved incubation with 0.1% trypsin (37°C, 15 min), then exposure to a mixture of 0.25 U/mL chondroitinase ABC (C3667, Sigma-Aldrich, Missouri, USA) and 1 U/mL hyaluronidase (H3506, Sigma-Aldrich, Missouri, USA) at 37°C for 60 min. Subsequently, sections were incubated overnight at 4°C with antibodies targeting COL2A1 (28459-1-AP, Proteintech, Wuhan, China), MMP13 (18165-1-AP, Proteintech, Wuhan, China), PIGR (TB7817, Abmart, Shanghai, China), and P53 (10442-1-AP, Proteintech, Wuhan, China). Visualization was achieved using IHC Detect Kit for Rabbit/Mouse Primary Antibody (PK10006, Proteintech, Wuhan, China). Hematoxylin was applied as a counterstain. Quantitative assessment of immunohistochemical staining was performed using ImageJ software, applying global thresholding consistently across comparable sets of images.
Cell culture and treatment
C28/I2 human chondrocytes were maintained in Dulbecco’s Modified Eagle Medium (DMEM) enriched with 10% FBS and supplemented with antibiotics (50 μg/mL penicillin-streptomycin). The culture environment was set at 37°C with 5% CO2.
For inflammatory response studies, C28/I2 cells were seeded in 6-well plates at a density of 2 × 105 cells per well. These cultures were then exposed to either 10 ng/mL IL-1β (HY-P7028, MedChemExpress, New Jersey, USA) or 10 mg/L LPS (HY-D1056, MedChemExpress, New Jersey, USA) for 48 h, with untreated wells serving as controls.
To investigate the effects of CA, C28/I2 cells were treated with varying concentrations of CA (0, 0.05, 0.25, 1.25, and 1.5 μM) prepared in Phosphate-buffered saline (PBS).
For PIGR gene silencing experiments, C28/I2 cells underwent transfection with PIGR-specific shRNA plasmids (sc-42964, Santa Cruz Biotechnology, Texas, USA) using Lipofectamine 3000 (L3000015, Thermo Fisher, Massachusetts, USA) as the transfection agent. The transfected cultures were maintained for 48 h before subsequent analyses.
Cell counting kit-8 assay
For the assay, C28/I2 cells were plated in 96-well format at a concentration of 4000 cells per well, with each well containing 100 μL of culture medium. Following a 48-h treatment period, 10 μL of cell counting kit-8 (CCK8) reagent (HY-K0301, MedChemExpress, New Jersey, USA) was introduced to each well. The plates were then incubated for an additional 2 h, after which absorbance measurements were taken at a wavelength of 450 nm. All experiments were repeated three times to ensure reliability and reproducibility.
Drug affinity responsive target stability
The DARTS assay was employed to evaluate the interaction stability between small molecule drugs and their target proteins. This method was adapted from a previously published protocol. 31 The procedure began with the preparation of cell lysates from either C28/I2 cells. These lysates were then exposed to either CA at 200 μM or PBS (vehicle control) for 1 h at ambient temperature.
Following this incubation, the lysates, containing equivalent amounts of protein, underwent controlled proteolysis using thermolysin. This digestion step was carried out at room temperature for 10 min. The proteolytic reaction was then halted by the addition of Phenylmethylsulfonyl fluoride (PMSF), with the samples being placed on ice for 5 min. To quantify the levels of the target proteins posttreatment, Western blot analysis was performed on the processed samples.
SA-Green staining
To detect senescent cells, we employed the Senescence-Tracker™ Senescent Cell Green Fluorescent Staining Kit (SA-Green, C0607M, Beyotime Biotechnology, Shanghai, China). This assay leverages the detection of senescence-associated β-galactosidase activity.
Following the application of various experimental treatments, the culture medium was aspirated from the cells, which were then gently rinsed with PBS. The first step in the staining process involved the application of an endogenous β-gal inhibitor working solution. The cells were then incubated with this solution at 37°C for 1 h. Subsequently, the SA-Green staining working solution A was carefully introduced to the cell culture. This step was followed by another 1-h incubation at 37°C in darkness to preserve the stain’s fluorescent properties. Upon completion of the staining procedure, the cells were examined using a fluorescence microscope. The proportion of senescent cells was determined by calculating the percentage of cells exhibiting positive staining.
Real-time reverse transcriptase-quantitative PCR analysis
RNA was extracted for real-time reverse transcriptase-quantitative Polymerase chain reaction (PCR) (RT-qPCR) analysis using RNA Extraction Buffer (R0006, Beyotime Biotechnology, Shanghai, China) on cell samples. Complementary DNA was generated from 1 μg of extracted RNA using RT Master Mix for qPCR II (gDNA digester plus) (HY-K0510A, MedChemExpress, New Jersey, USA). The RT-qPCR procedure utilized SYBR Green qPCR Master Mix (HY-K0501A, MedChemExpress, New Jersey, USA) following the manufacturer’s guidelines. Specific primers for target genes were used as detailed in Supplementary Table S1. Gene expression levels were quantified using the 2−ΔΔCT method, with results expressed as fold changes relative to a reference gene.
Western blot analysis
For Western blot analysis, cells were subjected to a series of preparation steps. First, cells were rinsed thoroughly with prechilled PBS, and proteins were extracted using Radioimmunoprecipitation assay (buffer) (RIPA) buffer supplemented with protease inhibitors. The resulting lysate was heat treated (100°C, 10 min) and then centrifuged at high speed. The clear fractions were then separated and further processed by sonication and centrifugation again. Samples were stored at −80°C until required.
Proteins were separated by Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to Polyvinylidene difluoride (PVDF) membranes. The membranes were then blocked with 5% skim milk in Tris-buffered saline with Tween 20 (TBST). Primary antibodies were incubated overnight at 4°C, followed by treatment with appropriate secondary antibodies for 1 h at room temperature. Signal detection was performed using chemiluminescent substrates and visualized using the GelDoc imaging system. Details of all antibodies are provided in Supplementary Table S2.
NETWORK PHARMACOLOGY
The 2D Structure data file (SDF) structure and Simplified molecular input line entry system (SMILES) string of CA were obtained from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/) and imported into the TargetNet (http://targetnet.scbdd.com/) online data platform, and 75 targets were retrieved.
Using “Osteoarthritis (OA)” as the search term, 5207 component targets were obtained on the Genecards (https://www.genecards.org/) online data platform. The gene data obtained by screening disease-related targets and component-related targets were intersected, and a total of 40 intersection targets were obtained. The intersecting targets were imported into the STRING database (Homo sapiens; multiple proteins) to construct a protein–protein interaction (PPI) network, representing the potential in vivo response network of CA in OA.
The results of the mutual mapping were imported into the online tool DAVID (https://david.ncifcrf.gov/). Homo sapiens was selected as the species, and a threshold of P < 0.05 was applied. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were then performed, including GO Biological Processes, GO Molecular Functions, GO Cellular Components, and KEGG pathways. The GO and KEGG pathway enrichment results were plotted according to the P value for visual analysis.
MS ANALYSIS
Primary mouse chondrocytes (P1) were isolated and cultured as described above. After reaching approximately 80% confluence, cells were treated with IL-1β (10 ng/mL) or IL-1β + CA (0.25 μM) for 48 h. At the end of treatment, cells were washed twice with cold PBS and lysed in RIPA buffer (Beyotime, Shanghai, China) supplemented with 1 mM PMSF and protease inhibitor cocktail. The lysates were sonicated on ice and centrifuged at 12,000 g for 10 min at 4°C to remove debris. Total protein concentration was determined using a BCA protein assay kit (Thermo Fisher Scientific, USA).
Equal volumes of protein lysis were sent to Shanghai Baipu Biotechnology Co., Ltd. for MS analysis. Proteins identified by at least one unique peptide were considered valid. Differentially expressed proteins were ranked according to their MaxQuant scores, and the top 10 proteins were selected for further analysis. These candidates were then cross-referenced with the 40 predicted CA–OA targets identified by network pharmacology and molecular docking, yielding three overlapping proteins (PIGR, Peptidylglycine alpha-amidating monooxygenase (PAM), and Heme oxygenase 1 (HMOX1). Among them, PIGR exhibited the highest score and was chosen for subsequent validation experiments.
Molecular docking
The crystal structure of human PIGR (PDB ID: 1XED, resolution 1.90 Å) was obtained from the Protein Data Bank (https://www.rcsb.org/structure/1XED). The 3D structure of CA (CAS: 104-54-1) was downloaded from PubChem. Protein and ligand files were converted to PDBQT format, and docking was performed using AutoDock Vina 1.2.2 within a 40 Å × 40 Å × 40 Å grid box (grid spacing = 0.05 nm). The lowest-energy complex was visualized in PyMOL 2.5.
Statistical analysis
All statistical analyses were performed using GraphPad Prism 10.0 (GraphPad Software Inc., USA). Data are presented as mean ± Standard error of the mean (SEM). Differences among multiple groups were analyzed using one-way Analysis of variance (ANOVA) followed by Tukey’s post hoc multiple-comparison test. For clarity, significance indicators in each figure represent the most biologically relevant pairwise comparisons (e.g., control vs. IL-1β, IL-1β vs. IL-1β + CA) derived from the ANOVA post hoc analysis. For comparisons between two groups, unpaired two-tailed Student’s t-tests were used. The significance threshold was set at P < .05. Detailed statistical parameters and sample sizes are provided in the corresponding figure legends.
RESULTS
CA enhances chondrocyte viability and maintains cartilage homeostasis in vitro
We first evaluated the effect of different concentrations of CA (0.05, 0.25, 1.25, 1.5 μM) on C28/I2 cell viability using the CCK8 assay (Fig. 1A). The results showed that CA at concentrations between 0.05 and 1.25 μM upregulated cell viability, peaking at 0.25 μM, while 1.5 μM CA significantly inhibited C28/I2 cell proliferation.
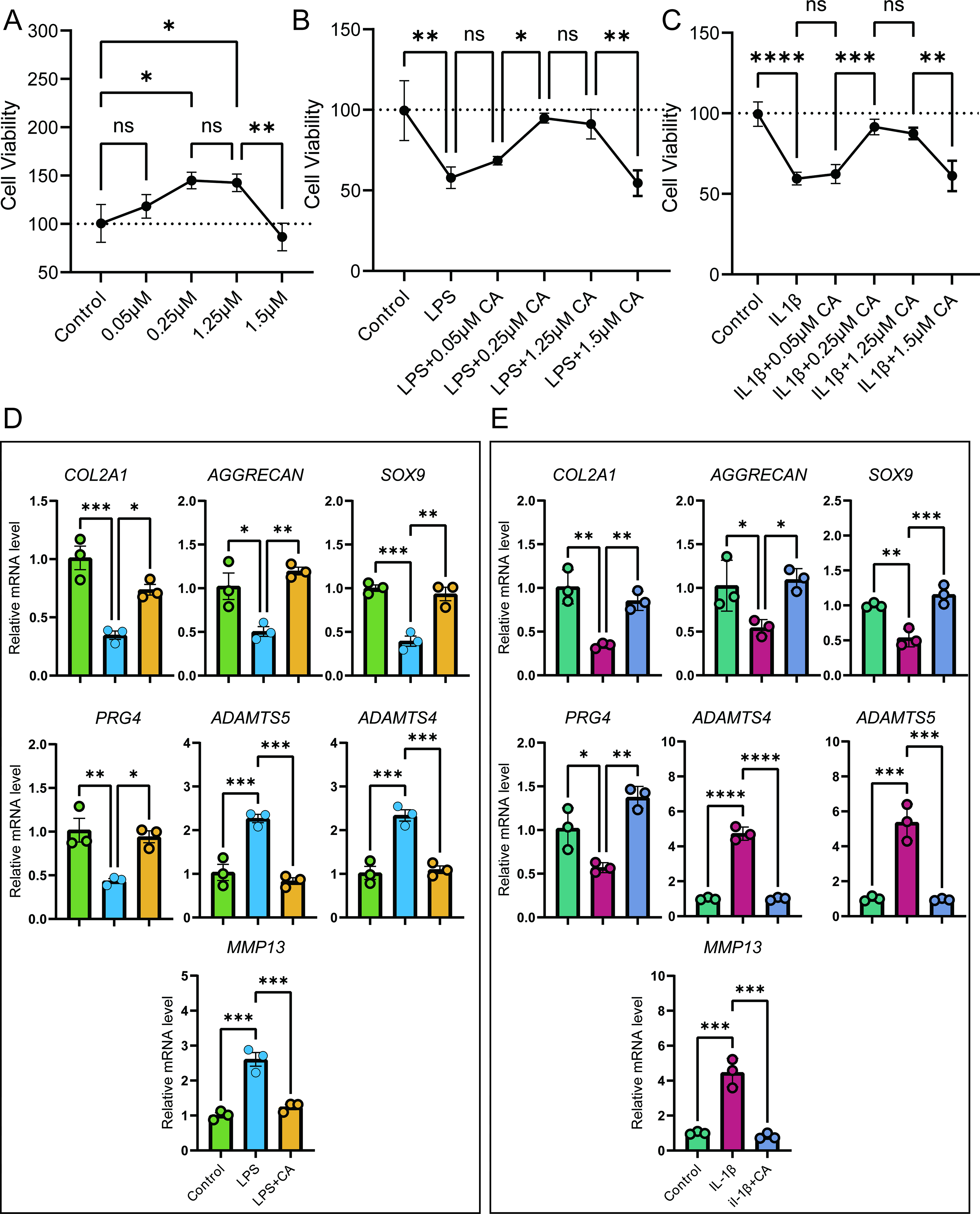
Effects of cinnamyl alcohol (CA) on chondrocyte viability and metabolism under inflammatory conditions.
To mimic the in vivo OA cartilage environment, we treated C28/I2 cells with IL-1β (Fig. 1C) or LPS (Fig. 1B) in vitro and examined the potential alleviating effects of different CA concentrations using the CCK8 assay. Compared with the control group, both LPS and IL-1β groups showed decreased cell viability, indicating that LPS and IL-1β inhibit chondrocyte viability. Treatment with various concentrations of CA (0.05, 0.25, 1.25) on LPS- or IL-1β-treated cells improved chondrocyte viability, with 0.25 μM CA showing the strongest recovery effect. Consequently, 0.25 μM CA was used for subsequent studies.
We then investigated the impact of CA on chondrocyte synthesis and degradation capabilities. Under LPS treatment (Fig. 1D), C28/I2 cells showed significantly decreased levels of cartilage synthesis-related markers (collagen, type II, alpha 1 [COL2A1], AGGRECAN, SRY-box transcription factor 9 [SOX9], and proteoglycan 4 [PRG4]) and increased levels of cartilage degradation markers (ADAM metallopeptidase with thrombospondin type 1 motif [ADAMTS]4, ADAMTS5, and MMP13). Treatment with 0.25 μM CA significantly reversed these changes. Similarly, IL-1β-treated C28/I2 cells exhibited effects comparable to LPS treatment (Fig. 1E), and 0.25 μM CA treatment effectively ameliorated the IL-1β-induced dysregulation of chondrocyte synthesis and degradation. These findings suggest that CA may have a significant protective effect on OA cartilage.
CA mitigates chondrocyte senescence in osteoarthritic conditions
Given that OA is an age-related disease, we investigated the effects of CA on chondrocyte senescence in the context of OA (Fig. 2). We employed the SA-β-Gal assay using SA-Green fluorescent probes to specifically detect senescence-associated β-galactosidase activity in cells (Fig. 2A,B).
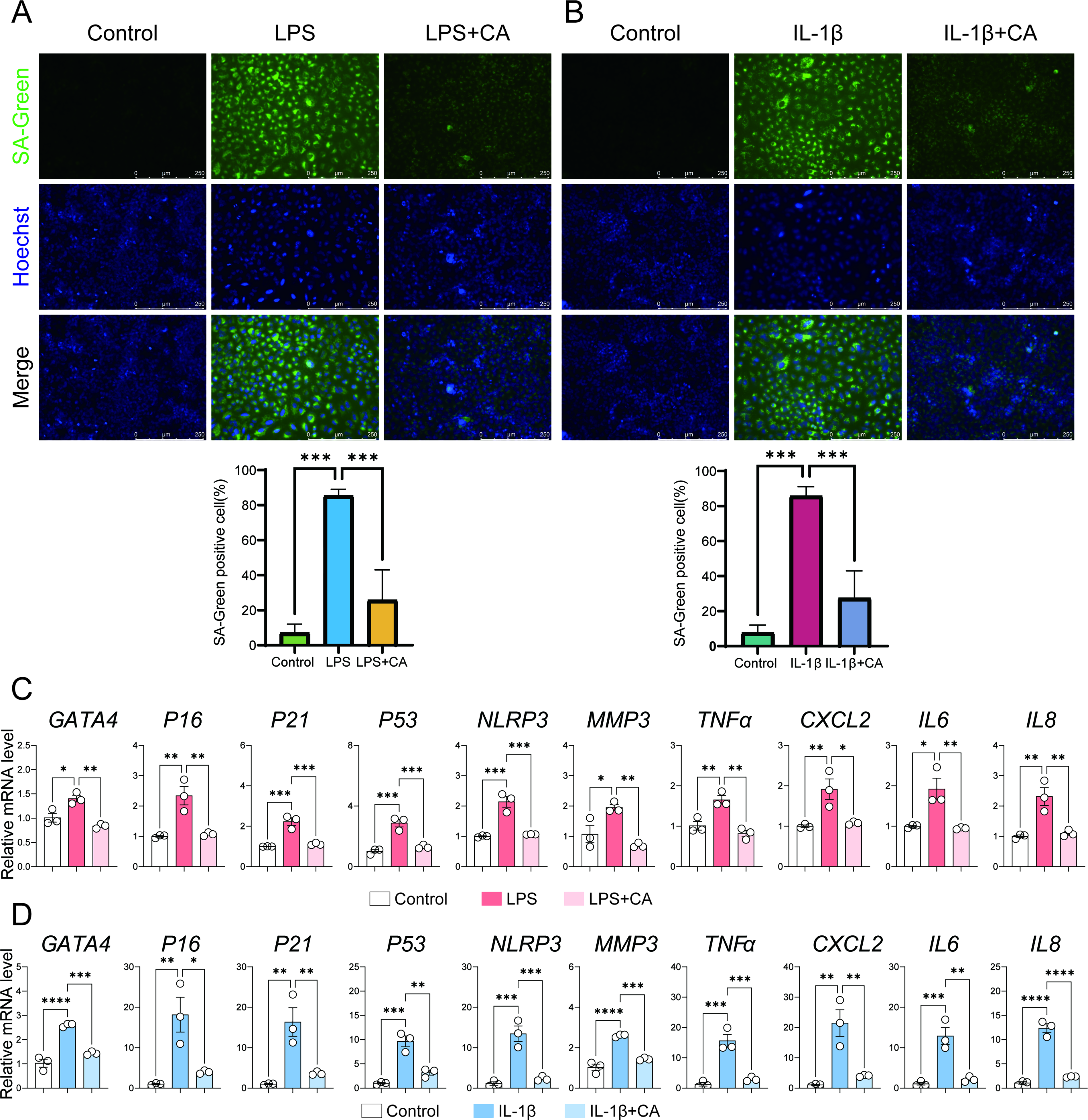
Cinnamyl alcohol attenuates chondrocyte senescence under inflammatory conditions.
LPS treatment significantly increased the proportion of SA-Green positive C28/I2 cells compared with the control group, indicating effective induction of chondrocyte senescence. Notably, CA treatment markedly reduced the proportion of SA-Green positive cells in the LPS-treated group (Fig. 2A). Similar results were observed with IL-1β treatment, where CA also decreased the proportion of SA-Green positive cells induced by IL-1β (Fig. 2B).
To further elucidate the impact of CA on cellular senescence, we examined its effect on the SASP using RT-qPCR. LPS significantly upregulated the expression of SASP-related molecules (GATA binding protein 4 [GATA4], P16, P21, MMP3, TNFα, CXCL2, IL6, IL8, NLR family pyrin domain containing 3 [NLRP3]) in C28/I2 cells compared with the control group. Importantly, CA treatment significantly attenuated the expression of these SASP-related markers (Fig. 2C). Consistent results were observed in the IL-1β model, where CA similarly reduced the expression of SASP-related genes induced by IL-1β (Fig. 2D).
These findings collectively demonstrate that CA effectively mitigates chondrocyte senescence induced by inflammatory stimuli such as LPS and IL-1β, suggesting its potential therapeutic role in age-related OA progression.
Furthermore, Western blot analysis revealed that, compared with the control group, LPS treatment decreased COL2 protein levels while increasing MMP13 and P53 protein levels. Notably, CA supplementation partially restored COL2 levels and significantly reduced MMP13 and P53 levels (Supplementary Fig. 1A). Similar effects were observed in the IL-1β model (Supplementary Fig. S1B). These protein-level changes corroborate our mRNA expression findings and further support the protective role of CA against chondrocyte senescence and matrix degradation in OA-like conditions.
CA ameliorates OA progression in a DMM mouse model
To further investigate the effects of CA on OA in vivo, we employed the destabilization of the DMM mouse model, which exhibits pathological features similar to those observed in patients with OA and is widely used to examine the therapeutic efficacy of various compounds against OA. CA was administered via intra-articular injection following DMM surgery, and mouse knee joints were collected 8 weeks post-DMM (Fig. 3A).

Cinnamyl alcohol ameliorates osteoarthritis progression in a DMM mouse model.
Joint pain, a primary clinical symptom of OA, was assessed using the von Frey test. As expected, DMM surgery resulted in decreased paw withdrawal thresholds. Interestingly, CA injection significantly increased the paw withdrawal threshold, indicating alleviation of OA-associated pain (Fig. 3B).
Cartilage erosion and loss of superficial cartilage are hallmark pathological features of OA. Safranin O staining of knee joint sections, a marker for proteoglycans, revealed that CA treatment substantially improved these pathological changes (Fig. 3C). Quantitative analysis using the OARSI scoring system showed significantly lower scores in the CA-treated group compared with the vehicle group (Fig. 3H).
Immunohistochemical staining further demonstrated that protein levels of anabolic markers Col2 (Fig. 3D,I) and Aggrecan (Fig. 3E,J) were reduced in cartilage tissue following DMM surgery; however, this effect was reversed with CA administration. Moreover, CA treatment decreased the protein levels of the catabolic marker MMP13 (Fig. 3F,K) and the senescence marker P53 (Fig. 3G,L) compared with the vehicle group.
These findings collectively demonstrate that CA effectively ameliorates OA progression in vivo by alleviating pain, preserving cartilage integrity, maintaining cartilage homeostasis, and reducing senescence in the DMM mouse model.
CA demonstrates a favorable safety profile in vivo
To assess the safety of CA administration, we monitored mouse body weight and major organ weights upon sample collection. Compared with the sham surgery group, mice receiving intra-articular CA injections did not show statistically significant differences in body weight gain (Fig. 4A). Furthermore, no significant differences were observed in spleen and liver weights between the CA-treated and sham surgery groups (Fig. 4B, C).

Safety assessment of cinnamyl alcohol administration in vivo.
Histological analysis of the liver revealed that CA did not induce any notable pathological changes compared with the sham surgery group (Fig. 4D). These results collectively suggest that intra-articular injection of CA does not elicit any apparent toxicity, indicating a favorable safety profile for potential therapeutic applications.
Network pharmacology analysis reveals PIGR as a potential target of CA in OA
To further explore the potential targets of CA in alleviating OA cartilage pathology and senescence, we conducted a network pharmacology analysis. By intersecting OA targets with CA action targets (Fig. 5A), we identified 40 shared target proteins. KEGG and GO analyses revealed that these targets were enriched in multiple signaling pathways related to cartilage ECM damage and aging (Supplementary Fig. S2). For instance, GO terms such as cellular response to LPS and oxidative stress response were associated with the LPS-induced model. In contrast, the regulation of TNF, Toll-like receptor, Nucleotide-binding oligomerization domain (NOD)-like, and NF-κB signaling pathways was closely related to inflammation and aging processes, further confirming the therapeutic potential of CA against OA. In the constructed PPI network (Fig. 5B), node size and color intensity reflect the degree value and betweenness centrality of each protein, indicating its overall connectivity within the network. MS analysis of mouse primary chondrocytes (P1) treated with IL-1β or IL-1β + CA yielded 10 top-scoring potential target proteins (Fig. 5C–D). Intersection of these proteins with the 40 predicted docking targets revealed three shared candidates—PIGR, PAM, and HMOX1—with PIGR showing the highest score (Fig. 5E). These combined results highlight PIGR as a convergent target supported by both computational prediction and experimental identification.
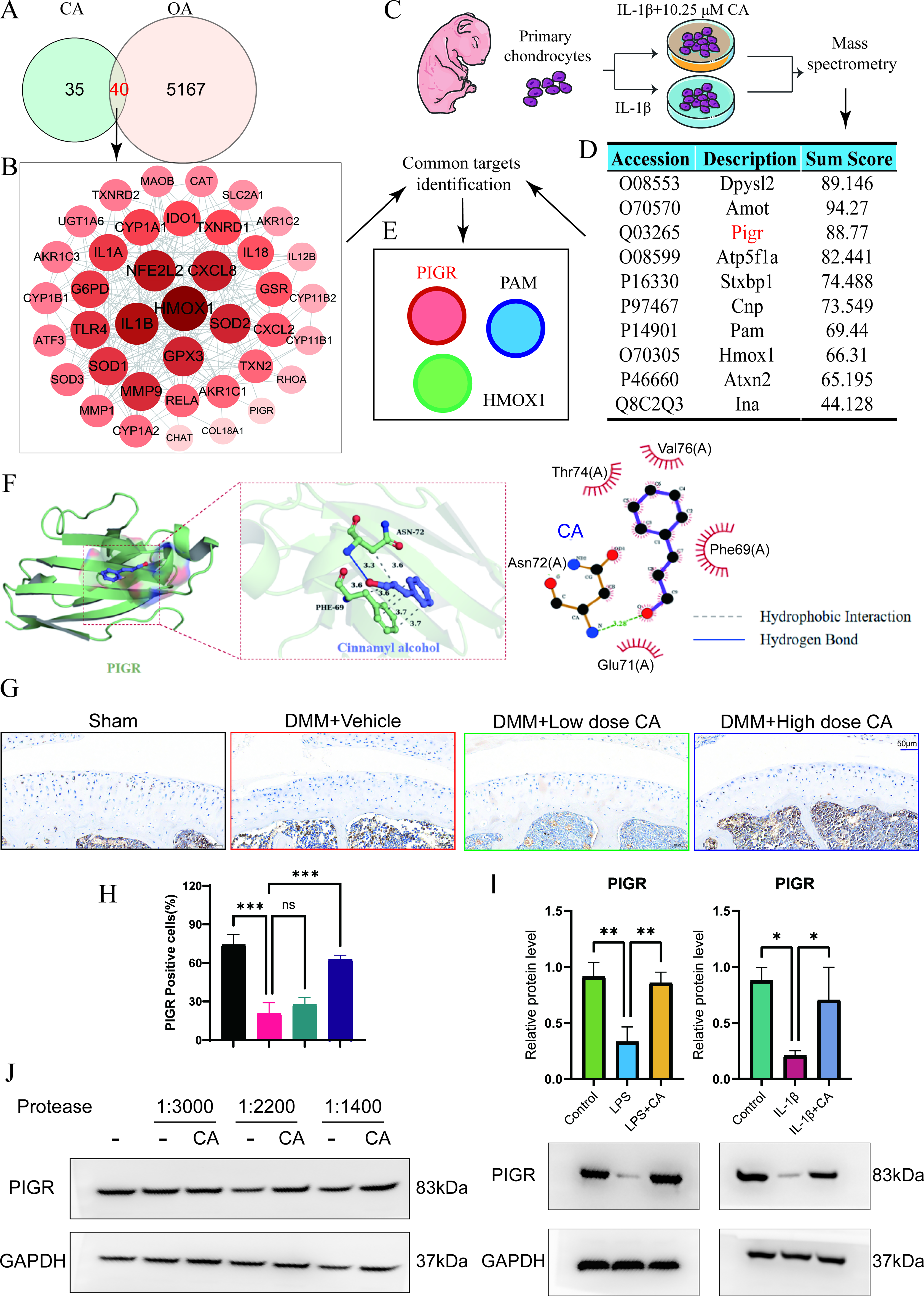
Network pharmacology analysis reveals PIGR as a potential target of cinnamyl alcohol in osteoarthritis.
Molecular docking simulations were used to model the binding capacity of CA to PIGR. The 2D structure of CA was downloaded from PubChem, and the protein structure of PIGR was obtained from the PDB database. After preprocessing both the ligand (CA) and protein receptor (PIGR), AutoDock Vina was used for docking. The binding affinity (kcal/mol) was used to evaluate the binding strength between the small molecule ligand and the receptor. PyMOL (v2.2.0) was used to visualize the conformation with the lowest binding energy. The molecular docking results showed that CA could form a stable molecular conformation with PIGR (Fig. 5F). Purple represents CA, and green represents PIGR. The docking score of CA to PIGR was −4.45 kcal/mol, suggesting a moderate binding affinity compatible with a transient small-molecule–protein interaction. CA formed a stable binding interface with the receptor through hydrogen bonds (blue solid line) and hydrophobic interactions (gray dashed line). A hydrogen bond was observed with Asn72, and multiple hydrophobic contacts were formed with Phe89, which together may enhance the specificity of ligand–receptor recognition.
Immunohistochemistry results showed that PIGR protein expression decreased after DMM surgery in mice, and CA treatment reversed this DMM surgery-induced reduction (Fig. 5G–H). Furthermore, in vitro experiments (Fig. 5I) demonstrated that compared with the control group, LPS or IL-1β treatment reduced PIGR protein expression, while 0.25 μM CA treatment reversed this LPS- or IL-1β-induced decrease in PIGR protein levels.
To explore whether PIGR is a potential CA-binding target that mediates chondrocyte homeostasis, we performed a DARTS assay. Western blot analysis was conducted on DARTS samples (with or without CA) treated with a series of thermolysin proteases. We found that CA protected the PIGR protein from protease digestion (Fig. 5J), further supporting PIGR as a potential target of CA.
PIGR knockdown attenuates the protective effects of CA on chondrocyte senescence and ECM degradation
To investigate the role of PIGR in chondrocyte senescence in vitro, we employed shRNA to suppress PIGR expression. RT-qPCR analysis confirmed high silencing efficiency of shPIGR compared with the negative control (shRNA group) (Supplementary Fig. S3).
We first examined the impact of PIGR silencing on LPS-induced damage (Fig. 6A). The experiment was divided into four groups: LPS, LPS + CA, LPS + CA + shRNA, and LPS + CA + shPIGR. Compared with the LPS group, treatment with CA or CA + shRNA inhibited the increase in cartilage catabolic markers and the decrease in anabolic markers. However, PIGR silencing significantly weakened CA’s regulatory capacity on cartilage damage. Similar effects were observed in the IL-1β-induced in vitro model (Fig. 6B).
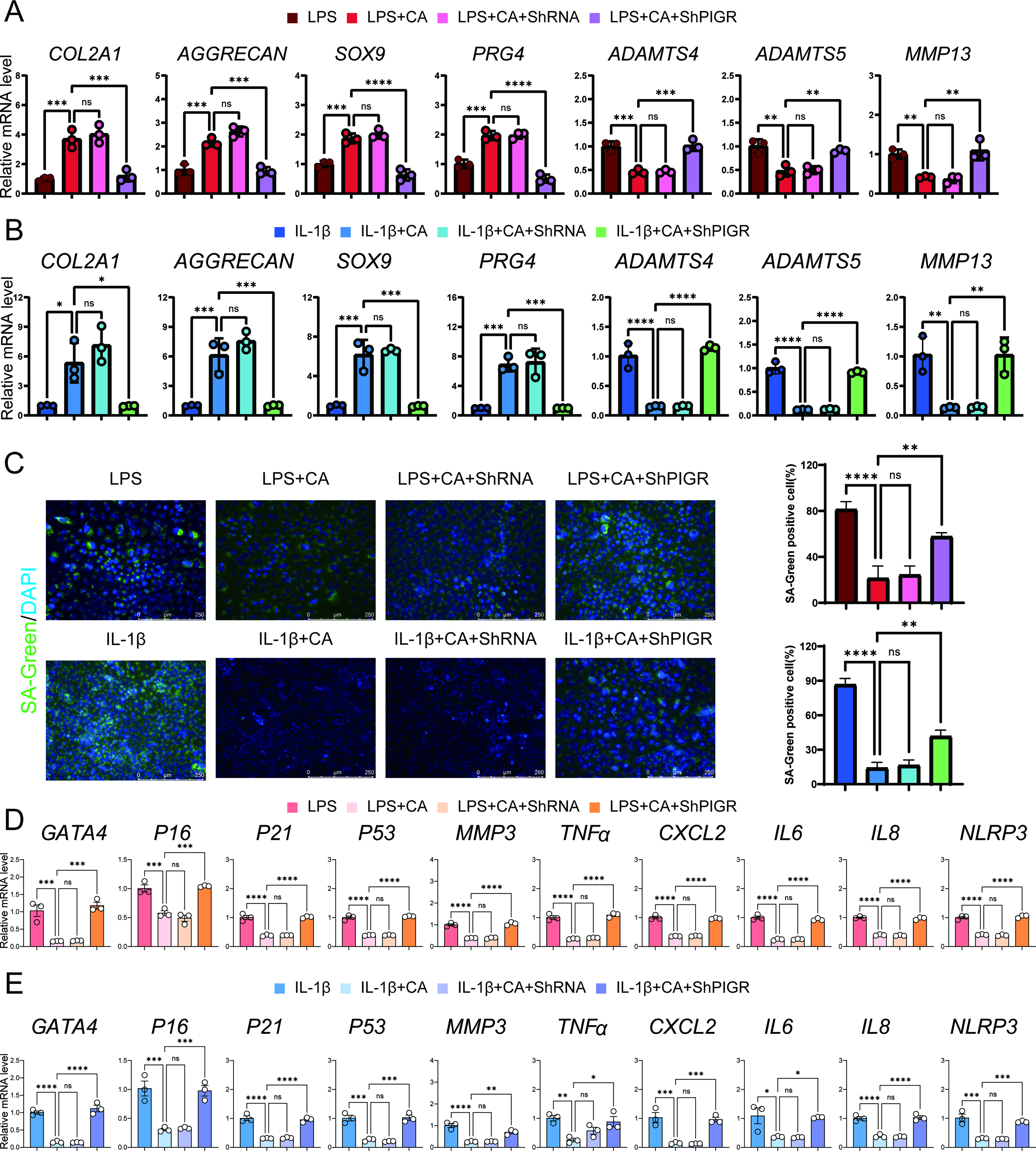
PIGR knockdown attenuates the protective effects of cinnamyl alcohol on chondrocyte senescence and ECM degradation.
SA-β-Gal assay staining revealed that CA’s ability to reduce the proportion of positive cells induced by LPS or IL-1β was attenuated by PIGR silencing (Fig. 6C). This suggests that PIGR may be a key regulator of cartilage ECM and cellular senescence.
We further examined the expression of SASP-related genes by RT-qPCR. As expected, SASP-related genes were significantly upregulated in PIGR-knockdown chondrocytes compared with the negative control group, in both the LPS model (Fig. 6D, Supplementary Table S5) and the IL-1β model (Fig. 6E, Supplementary Table S6).
Collectively, these data demonstrate that PIGR plays a crucial role in restraining chondrocyte senescence and joint ECM degradation, and that its loss accelerates OA-related changes. These findings further support PIGR as an essential mediator of the protective effects of CA against OA.
In this study, we demonstrate that CA exhibits potent antisenescent and cartilage-protective effects that may involve PIGR-associated regulation of inflammatory pathways. IL-1β and LPS are classical activators of the NF-κB and STAT3 signaling cascades that promote chondrocyte senescence and SASP expression. Although these pathways were not directly examined in our experiments, previous studies have shown that PIGR expression is NF-κB-responsive and can mediate cytokine feedback regulation. 25,26 Based on this evidence, we propose that restoration of PIGR by CA may attenuate inflammatory amplification loops, leading to decreased MMP13 and SASP expression while preserving ECM homeostasis.
The protective effects of CA observed in our study are consistent with previous research highlighting its anti-inflammatory effects. 16,32,33 Our results extend these findings to the context of OA, demonstrating CA’s ability to preserve cartilage integrity and function. The amelioration of OA symptoms in the DMM mouse model, coupled with the favorable safety profile, suggests that CA could be a promising therapeutic agent for OA management.
Notably, our study integrated network pharmacology prediction with MS to achieve a more reliable target identification strategy. This dual-filtering approach allowed us to narrow the broad set of predicted CA–OA interaction targets to a small group of experimentally supported proteins. Among the overlapping candidates, PIGR emerged as the top-ranked protein, thereby representing a convergent finding from both computational and experimental datasets. This integrative strategy not only strengthens confidence in PIGR as a bona fide mediator of CA activity but also exemplifies a generalizable framework for discovering mechanistic targets of natural compounds in complex diseases such as OA.
The identification of PIGR as a potential mediator of CA’s effects provides a new perspective on the molecular mechanisms underlying OA pathogenesis and treatment.
The multifaceted effects of CA on chondrocyte senescence and ECM homeostasis likely stem from its interaction with multiple signaling pathways. Our network pharmacology analysis revealed enrichment in pathways related to inflammation, oxidative stress, and cellular senescence. These findings are consistent with recent studies showing the involvement of NF-κB, Mitogen-activated protein kinase (MAPK), and PI3K/AKT pathways in OA progression. 34,35 The novel identification of PIGR as a CA target suggests an unexplored mechanism in OA pathogenesis. PIGR has been primarily studied in the context of mucosal immunity, 36 and our findings indicate a potential role in cartilage homeostasis that warrants further investigation.
The therapeutic potential of CA demonstrated in this study has significant implications for OA management. Current OA treatments are largely symptomatic and do not address the underlying disease progression. 37 CA’s ability to modulate multiple aspects of OA pathogenesis, including inflammation, senescence, and ECM degradation, positions it as a promising candidate for disease-modifying OA drugs. Moreover, the favorable safety profile observed in our study suggests that CA could be well-tolerated in clinical applications, potentially offering a new avenue for long-term OA management with minimal side effects.
A key strength of this study is its comprehensive approach, combining in vitro, in vivo, and in silico methods to elucidate the therapeutic potential and mechanisms of CA in OA. The use of both LPS and IL-1β-induced models in vitro, coupled with the DMM mouse model in vivo, provides robust evidence for CA’s efficacy across different OA-like conditions. Furthermore, the identification of PIGR as a novel target through network pharmacology and subsequent validation experiments demonstrates the power of integrating computational and experimental approaches in drug discovery and mechanistic studies. Moreover, our findings extend the biological relevance of PIGR beyond its classical role in mucosal immunity, suggesting that it may also participate in regulating inflammatory–senescent crosstalk within cartilage. This hypothesis aligns with reports that curcumin and salicin attenuate OA by suppressing NF-κB and MAPK activation. 14,15 In this broader pharmacological context, CA may function as a multitarget anti-inflammatory compound that modulates senescence-associated signaling. Future investigations will determine whether PIGR directly interacts with canonical inflammatory pathways such as NF-κB or STAT3 in chondrocytes.
Despite these promising findings, several limitations should be addressed in future studies. First, while the DMM model is widely used in OA research, it may not fully recapitulate the complexity of human OA, which often involves multiple risk factors. 38 Future studies should explore CA’s efficacy in other OA models and ultimately in clinical trials. Second, the long-term effects of CA administration and potential side effects need further investigation. In addition, the exact mechanism by which PIGR influences cartilage homeostasis remains to be elucidated. Future research should focus on unraveling the downstream signaling pathways of PIGR in chondrocytes and exploring its potential interactions with other known OA-related pathways.
CONCLUSION
Our study provides compelling evidence for the therapeutic potential of CA in OA treatment. By demonstrating CA’s ability to mitigate chondrocyte senescence, maintain ECM homeostasis, and alleviate OA symptoms in vivo, we have laid the groundwork for future investigations into CA-based interventions for OA. The identification of PIGR as a novel target of CA opens new avenues for understanding OA pathogenesis and developing targeted therapies. As the burden of OA continues to grow globally, these findings offer hope for developing more effective, disease-modifying treatments that could significantly improve the quality of life for millions of OA patients.
AUTHORS’ CONTRIBUTIONS
Y.M. contributed to conceptualization, methodology, supervision, project administration, funding acquisition, writing—review and editing, and visualization. Q.Y. carried out investigation, formal analysis, data curation, validation, and writing—original draft. Q.Z. participated in methodology, investigation, and validation and provided resources. J.D. was responsible for investigation, data curation, and resources. H.T. contributed software for network pharmacology and molecular docking, performed formal analysis, and prepared visualizations. Y.H. conducted the in vivo investigations, managed data curation, and performed validation. Y.L. provided supervision, funding acquisition, and writing—review and editing. All authors have read and approved the final article, agree to be accountable for all aspects of the work, and consent to its exclusive submission to the Journal of Medicinal Food.
Footnotes
DATA AVAILABILITY STATEMENT
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
AUTHOR DISCLOSURE STATEMENT
There is no conflict of interest to declare.
FUNDING INFORMATION
No funding was received for this article.
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.