
Abstract
Melphalan, a widely used alkylating chemotherapeutic agent, is known to cause gonadotoxicity, particularly affecting spermatogenesis and steroidogenesis. This study investigated the protective effects of chrysin, a flavonoid with antioxidant and antiapoptotic properties, on melphalan-induced testicular damage in immature male rats. Forty-eight prepubertal Wistar rats were divided into six groups receiving melphalan (0.5 mg/kg/day, intraperitoneally), chrysin (50 or 75 mg/kg/day, orally), or combinations thereof. Treatment spanned 30 days, covering the prepubertal to pubertal transition. Melphalan exposure significantly reduced testosterone levels, disrupted spermatogenesis, and increased oxidative stress, as indicated by elevated malondialdehyde and decreased glutathione peroxidase levels. Gene expression analysis revealed marked upregulation of autophagy (LC3β, Beclin-1, Atg5, Atg7, Atg12) and apoptosis (BAX, caspase-3) markers, alongside downregulation of the antiapoptotic gene BCL2. Histological analysis showed severe damage to seminiferous tubules and reduced Johnsen scores. Chrysin coadministration, particularly at 75 mg/kg, significantly ameliorated these effects by restoring antioxidant capacity, reducing expression of cell death-related genes, and improving testicular structure and hormone levels. These findings demonstrate that chrysin mitigates melphalan-induced gonadotoxicity through modulation of oxidative stress, apoptosis, and autophagy pathways. Chrysin’s dose-dependent protective effects highlight its potential as a natural therapeutic agent to preserve reproductive function in individuals undergoing chemotherapy during prepuberty.
INTRODUCTION
Melphalan, an alkylating agent, is a crucial chemotherapy drug widely used in treating various cancers, including pediatric cancers, where it significantly improves survival rates.1,2 Despite its effectiveness, melphalan treatment in adults can cause considerable gonadotoxicity, resulting in impaired spermatogenesis and hormonal imbalance. 3 However, the exact molecular mechanisms behind melphalan-induced gonadal damage, especially in the prepubertal testis, are not fully understood. The prepubertal testis is characterized by a developmental stage marked by the presence of primarily mitotically active spermatogonia and immature Leydig cells that produce testosterone at much lower levels than in the adult testis. 4
Alkylating agents like melphalan cause their cytotoxic effects mainly by damaging DNA through alkylation, which leads to cell cycle arrest and cell death, primarily via apoptosis and autophagy. 5 Because oxidative stress and cell death are involved in melphalan-induced testicular damage, antioxidant therapies are being investigated for their protective effects. Flavonoids, a group of natural compounds, offer a promising therapeutic approach. Chrysin (5,7-dihydroxyflavone), a major flavonoid found in honey, has shown potential to reduce testicular damage, enhance spermatogenesis, and support steroidogenesis in compromised testes, acting as an antioxidant and antiapoptotic agent, often alongside other chemotherapies.6,7 Chrysin’s ability to protect against oxidative stress-induced damage and its capacity to counteract various forms of cell death, including apoptosis and autophagy, are well established. 8 Therefore, this study aimed to examine how chrysin administration affects the initiation of spermatogenesis and steroidogenesis in the immature testis after exposure to melphalan.
MATERIALS AND METHODS
Experimental design
A total of 48 male Wistar rats (aged 21 days, weighing 150 ± 25 g) were utilized in this experimental study. The animals were obtained and housed at the animal facility of Gonabad University of Medical Sciences under standardized laboratory conditions: a 12-h light/dark cycle, ambient temperature maintained at 22 ± 2°C, and appropriate ventilation. Rats had unrestricted access to standard chow and water throughout the experiment.
Experimental groups
The rats were randomly allocated into six experimental groups (n = 8 per group) based on melphalan exposure and chrysin treatment as follows:
Control group: Received intraperitoneal (IP) injections of normal saline and corn oil (oral gavage) daily for 30 days. Melphalan (Mel) group: Administered melphalan (0.5 mg/kg/day, IP) for 30 days. Melphalan + chrysin 50 mg/kg (Mel + Chr50) group: Treated with melphalan (0.5 mg/kg/day, IP), along with oral chrysin (50 mg/kg/day) by gavage for 30 days. Melphalan + chrysin 75 mg/kg (Mel + Chr75) group: Administered melphalan (0.5 mg/kg/day, IP), in combination with oral chrysin (75 mg/kg/day) for 30 days. Chrysin 50 mg/kg (Chr50) group: Received chrysin alone (50 mg/kg/day, oral gavage) for 30 days. Chrysin 75 mg/kg (Chr75) group: Received chrysin alone (75 mg/kg/day, oral gavage) for 30 days.
Twenty-four hours after the final treatment, rats were anesthetized with ketamine (50 mg/kg) and xylazine (10 mg/kg) administered IP. 9 Bilateral orchiectomy was performed: The right testis was fixed in formalin for histological analysis, while the left testis was homogenized for gene expression analysis via real-time PCR. Blood samples were also collected for biochemical assessments. In this study, melphalan was administered as an IP injection of 0.5 mg/kg daily. 10 Simultaneously, treatment with chrysin was conducted at two doses of 50 and 75 mg 11 daily for 30 days.
Histological processing and tissue preparation
The right testes were fixed in 10% formalin for 48 h. Subsequently, tissues were dehydrated in ascending concentrations of ethanol, cleared in xylene, and embedded in paraffin. Sections of 5 µm thickness were prepared and stained with hematoxylin and eosin. Histological evaluation was performed using a light microscope at 400× magnification. 12
Histomorphometric evaluation of seminiferous tubules
Histological parameters, including seminiferous tubular diameter and epithelial height, were assessed during stages VII and VIII of the spermatogenic cycle. Round cross-sectional tubules were selected randomly, and measurements were taken using ImageJ software. Tubular diameter was measured between two points of the basement membrane, and epithelial height was recorded from the basal lamina to the apical surface of the epithelium. Spermatogenesis was evaluated using the Johnsen scoring system, which rates 30 seminiferous tubules per animal on a scale of 1 to 10 based on the presence and maturation of germinal cells:
Score 10: Full spermatogenesis and intact tubular structure; Score 1: Complete absence of germ and Sertoli cells.
13
(Intermediate scores represent progressive degeneration or absence of specific cell types as per Johnsen’s criteria).
Gene expression analysis via real-time PCR
To investigate the expression of autophagy and apoptosis-related genes, the left testis was homogenized immediately postextraction. Total RNA was isolated using the Yekta Tajhiz Azma RNA extraction kit, Iran, following the manufacturer’s protocol. Complementary DNA (cDNA) was synthesized using a reverse transcriptase enzyme.
Real-time quantitative PCR (qPCR) was carried out using the YTA SYBR Green qPCR MasterMix (2×) and the StepOne Real-Time PCR System (Applied Biosystems). The reaction mix (10 µL) contained 1 µL cDNA, 0.2 µM forward primer, 0.2 µM reverse primer, 3.6 µL nuclease-free water, and 5 µL SYBR Green mix. The housekeeper’s gene was a GAPDH. Relative gene expression was analyzed using the 2−ΔΔCT method.
Thermal cycling conditions: Initial denaturation: 95°C for 10 min; amplification (40 cycles): 95°C for 15 sec, 58°C for 30 sec, 72°C for 30 sec; final extension: 95°C for 15 sec, 55°C for 60 min. 14
Target genes: Autophagy-related: LC3β, Beclin-1, Atg5, Atg7, and Atg12. Apoptosis-related: Caspase-3, BAX, and Bcl-2.
The oligonucleotide primers (Oligogen, South Korea) used in RT-PCR were as follows:
Bax (F primer: TTG CTA CAG GGT TTC ATC CA/R primer: GAG TAC CTG AAC CGG CAT CT),
Bcl2 (F primer: GCTACCGTCGTGACTTCGC/R primer: CCCCACCGAACTCAAAGAAGG),
Caspase3 (F primer: TGTGCTCCAGGCTTCCTTAATC/R primer: AGGCTTATGGGAAATGCTGGAC),
Beclin-1 (F primer: CGAAAGGTGGTGGCAGAAAAC/R primer: ACTATATTCTCGCTGGTACTGAGC),
Lc3β (F primer: TCAGTGAGAGCTGCCTCTGTC/R primer: AGCAGTGGGGATTTACACAGTG),
Atg5 (F primer: AGATCACAGTTCTGGGATGC/R primer: TCAGGCGGTAGAGATCGTAG),
Atg7 (F primer: TGTCTTGCAGCATCCTGAG/R primer: TCAAGAACTTTGGATGAACAGG),
Atg12 (F primer: GCACTCATCGACTTCATCAG/R primer: ACTGCCAAAACACTCATATAGAG),
GAPDH (F primer: GGTCTACATGTTCCAGTATGACTC/R primer: CATTTGATGTTAGCGGGATCTCG).
Biochemical assays
Malondialdehyde assay
Serum malondialdehyde (MDA) levels were measured based on thiobarbituric acid (TBA) reactivity. Briefly, 0.2 mL serum was mixed with 3.0 mL glacial acetic acid and 1% TBA in 2% NaOH. Samples were incubated in boiling water for 15 min, cooled, and the absorbance was recorded at 532 nm using a spectrophotometer.
Superoxide dismutase and glutathione peroxidase assays
Superoxide dismutase (SOD) and glutathione peroxidase (GPx) levels were determined using commercial enzyme-linked immunosorbent assay (ELISA) kits (Randox and Ransod, UK) and analyzed with an ELISA reader (Model: ABER-2, China). GPx activity was measured using the Paglia and Valentine method, which quantifies the oxidation of glutathione by cumene hydroperoxide. SOD activity was assessed based on the inhibition of formazan dye formation via the xanthine–xanthine oxidase reaction.15,16
Testosterone hormone level assays
The serum testosterone level was measured using the DEMEDITEC Testosterone rat/mouse ELISA kit, which relies on competitive binding in an ELISA. The procedure involved adding standards and samples in duplicate to the wells, followed by sequential addition of incubation buffer, enzyme, and substrate with specific incubation times and conditions, including shaking and dark incubation. After washing the wells multiple times, a stop solution was added to terminate the reaction, and absorbance was measured at 450 nm. A standard curve was then created by plotting the average absorbance values against known standard concentrations to determine the testosterone concentration in each sample. 17
STATISTICAL ANALYSIS
Data were analyzed using SPSS version 22. Normality of data distribution was assessed initially. For normally distributed variables, one-way ANOVA followed by Tukey’s post hoc test was used for group comparisons. Results are presented as mean ± standard error of the mean (SEM), with statistical significance set at P < .05.
RESULTS
The spermatogenesis index of seminiferous tubular, tubular diameter, and epithelial height
Histomorphometric Analysis of Spermatogenesis Index, Seminiferous Tubule Diameter, and Epithelial Height.
The Johnson score, seminiferous tubule diameter, and epithelial height were assessed in the testicular tissue of rats across experimental groups, with the results presented in Table 1.
Comparison of the Seminiferous Histological Marker and Testosterone Level in All Experimental Groups

Evaluation of testicular modified Johnsen score, seminiferous tubule diameter, epithelium height, and testosterone level following the administered melphalan and chrysin treatment. Values are mean ± SE. P < .05 are significant,*P < .05 versus control, **P < .05 versus Mel group. (n = 8 per group, one-way ANOVA, Tukey’s post hoc test).
Chr50, Chrysin50 group; Chr75, Chrysin75 group; mel + Chr50, melphalan + Chrysin50 group; mel + Chr75, melphalan + Chrysin75 group; mel, melphalan group.
The analysis revealed a significant reduction in all evaluated parameters (Johnson score, tubular diameter, and epithelial height) in the melphalan and melphalan + chrysin (50 mg/kg) groups compared with the control group. These findings suggest that melphalan induces substantial histological damage to testicular structure. Furthermore, coadministration of chrysin at 75 mg/kg with melphalan significantly improved all measured parameters relative to the melphalan-only group, indicating a protective effect of chrysin at this higher dose. Importantly, administration of chrysin alone (at both 50 and 75 mg/kg) did not result in significant changes in any of the assessed parameters when compared with the control group (Fig. 1).

H&E staining images of adult rat testicular tissue following melphalan administration and chrysin treatment in all study groups.
Serum testosterone level
By evaluating laboratory data and examining serum testosterone levels in the six groups, it was determined that serum testosterone levels in the group receiving melphalan were significantly reduced compared with the control group. Also, the groups receiving melphalan + chrysin at a dose of 50 and melphalan + chrysin at a dose of 75 showed a significant increase in serum testosterone levels compared with the group receiving melphalan alone. In the studies, the groups receiving chrysin at doses of 50 and 75 did not have a significant difference compared with the control group (Table 1).
Oxidative stress markers (GPx, SOD, MDA)
Analysis of oxidative stress biomarkers revealed the following (Fig. 2):
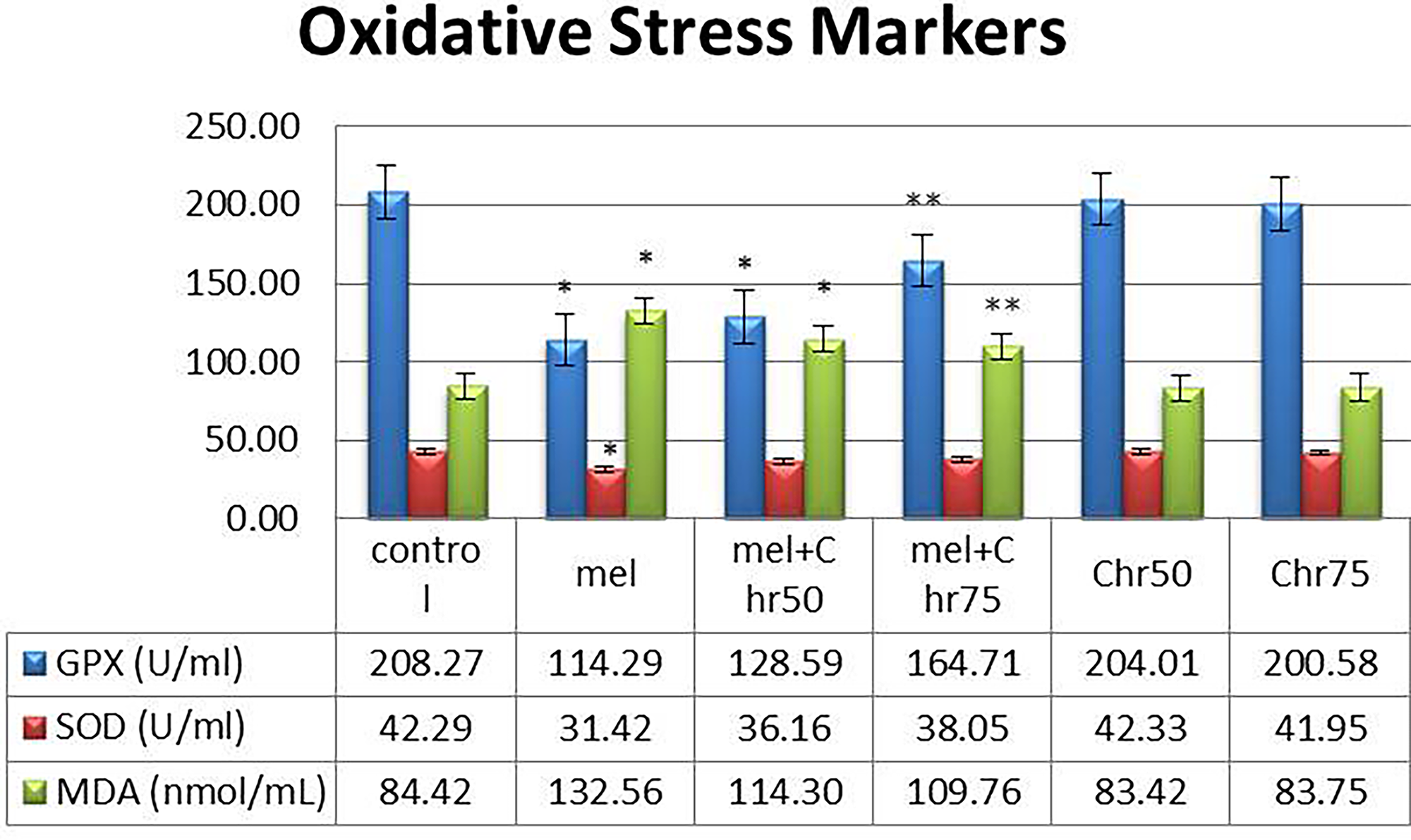
Evaluation of the results of serum levels of oxidative stress markers (MDA, SOD, GPx) following melphalan administration and chrysin treatment in all study groups. Values are mean ± SE. P < .05 are significant,*P < .05 versus control, **P < .05 versus Mel group. (n = 8 per group, one-way ANOVA, Tukey’s post hoc test). GPx, glutathione peroxidase; MDA, malondialdehyde; SOD, superoxide dismutase.
GPx:
A significant reduction in GPx levels was observed in the melphalan and melphalan + chrysin (50 mg/kg) groups compared with the control. However, coadministration of melphalan with chrysin at 75 mg/kg significantly increased GPx levels relative to melphalan alone. Chrysin alone (50 and 75 mg/kg) did not significantly alter GPx levels compared with the control.
SOD:
Melphalan treatment led to a significant decrease in SOD levels. Neither cotreatment with chrysin (50 or 75 mg/kg) nor chrysin alone produced significant changes in SOD levels compared with melphalan or control groups, respectively.
MDA:
MDA levels significantly increased in the melphalan and melphalan + chrysin (50 mg/kg) groups relative to the control. However, the melphalan + chrysin (75 mg/kg) group showed a significant reduction in MDA compared with the melphalan-only group. Chrysin alone did not significantly affect MDA levels.
Apoptosis-related gene expression (caspase-3, bax, BCL-2)
Gene expression analysis indicated (Fig. 3):
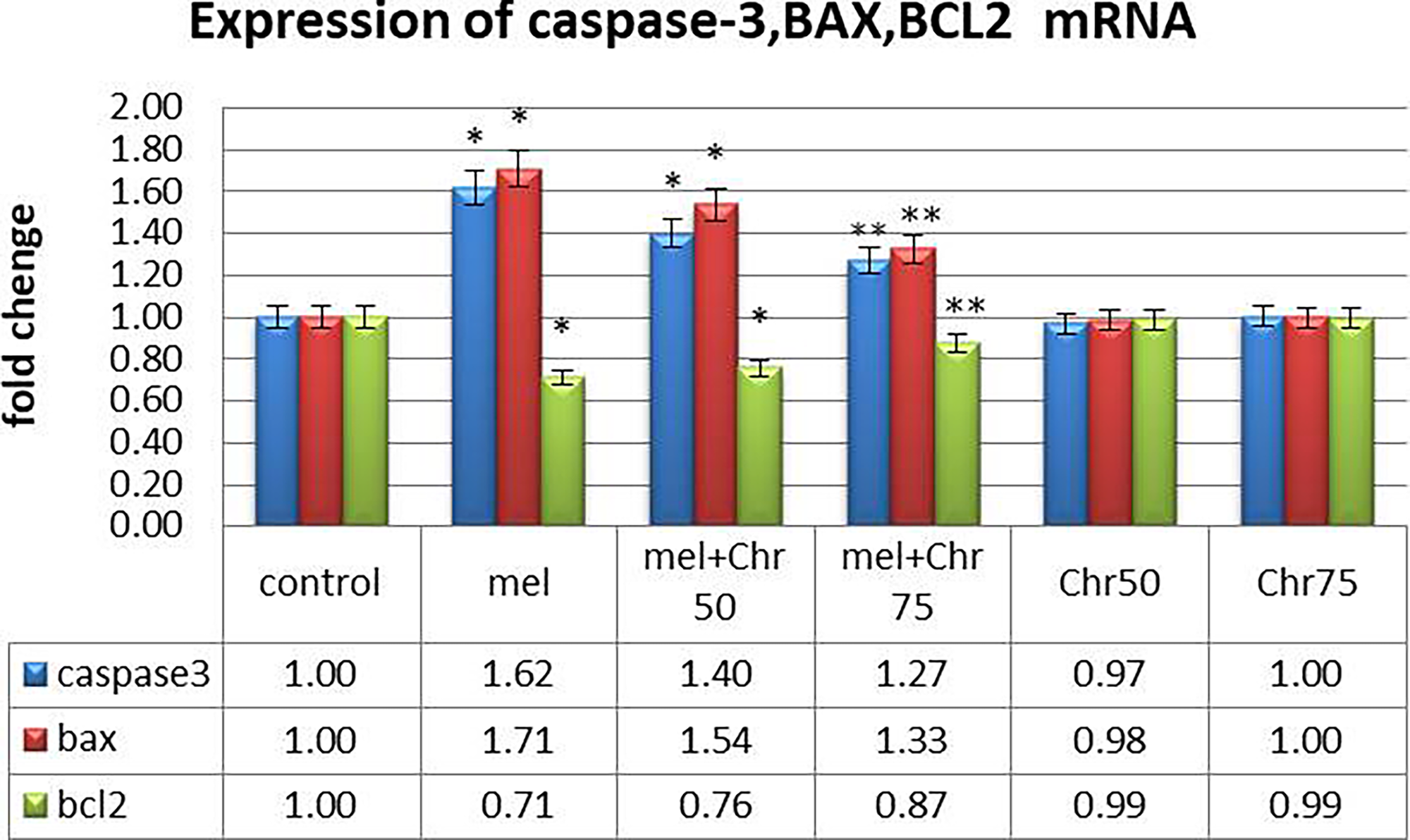
Real-time analysis of caspase-3, BAX, and BCL2 genes as apoptosis markers following melphalan administration and chrysin treatment in all study groups. Values are mean ± SE. P < .05 are significant,*P < .05 versus control, **P < .05 versus Mel group. (n = 8 per group, one-way ANOVA, Tukey’s post hoc test).
Caspase-3 and Bax (proapoptotic genes):
Expression levels of both genes were significantly elevated in the melphalan and melphalan + chrysin (50 mg/kg) groups compared with the control. Cotreatment with chrysin at 75 mg/kg significantly downregulated these genes relative to melphalan alone. Chrysin alone had no significant effect.
BCL-2 (antiapoptotic gene):
BCL-2 expression was significantly reduced in the melphalan and melphalan + chrysin (50 mg/kg) groups. However, chrysin coadministration at 75 mg/kg significantly restored BCL-2 expression compared with melphalan alone. No significant changes were observed in the chrysin-only groups.
Autophagy-related gene expression (beclin-1, LC3-II, ATG5, ATG7, ATG12)
Real-time PCR analysis of autophagy-related genes demonstrated (Fig. 4):
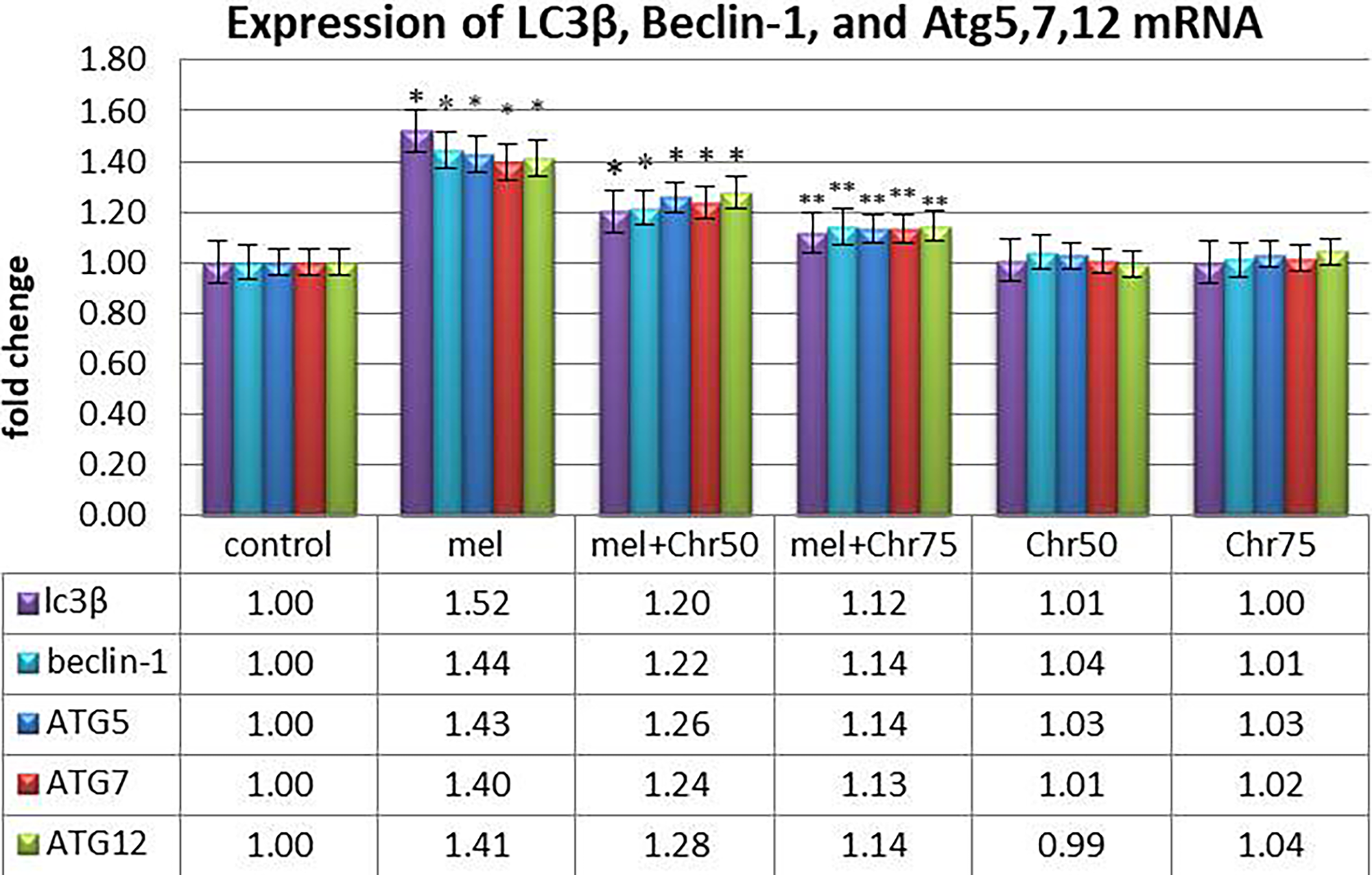
Real-time analysis of LC3β, Beclin-1, and Atg5, 7, and 12 genes as markers of autophagy following melphalan administration and chrysin treatment in all study groups. Values are mean ± SE. P < .05 are significant,*P < .05 versus control, **P < .05 versus Mel group. (n = 8 per group, one-way ANOVA, Tukey’s post hoc test).
Beclin-1, LC3-II, ATG5, ATG7, and ATG12:
All five genes were significantly upregulated in the melphalan and melphalan + chrysin (50 mg/kg) groups compared with the control group. Administration of chrysin at 75 mg/kg along with melphalan significantly reduced the expression of these genes compared with melphalan alone. Chrysin alone (50 or 75 mg/kg) did not significantly impact the expression of these autophagy markers.
DISCUSSION
This study explored how chrysin helps protect spermatogenesis, testosterone production, and testicular cell survival in immature rats exposed to melphalan toxicity. Importantly, the research spanned the transition from prepuberty to puberty in rats, allowing assessment of spermatogenic and steroidogenic activity at the end of treatment. The findings revealed that melphalan significantly damaged reproductive function, evidenced by disrupted spermatogenesis, reduced testosterone levels, and altered antioxidant enzyme markers (elevated MDA and decreased GPx). This increase in oxidative stress was associated with changes in gene expression related to cell death, including autophagy (LC3II, Beclin-1, Atg5, Atg7, Atg12) and apoptosis (BCL2, BAX, caspase-3), indicating heightened cell death in the testes.
Histological and biochemical examinations supported these results, showing structural damage to the testes and hormonal imbalance. Prior studies, such as those by Marvin Meistrich, have demonstrated the long-term reproductive effects of chemotherapy drugs like melphalan, particularly when administered during the prepubertal phase. 18 Other research indicates that melphalan exposure in adolescent male rats at doses of 1.5 mg/kg causes early and significant oxidative and tissue damage in the testes. 19
The results imply that melphalan harms the testes by promoting oxidative stress and activating both autophagy and apoptosis. Since immature testes are especially sensitive to alkylating agents 20 and melphalan can impair spermatogenesis and hormone production, effective protective strategies are crucial.19,21 Consequently, this study assessed chrysin’s ability to mitigate melphalan’s toxicity. Chrysin, a naturally occurring flavonoid known for its antioxidant and anti-inflammatory properties, was observed to support testicular health by lowering oxidative stress and inhibiting inflammatory and cell death pathways.22,23
Flavonoids like chrysin are noted for helping maintain cellular homeostasis by modulating key signaling pathways involved in apoptosis, autophagy, and inflammation.24,25 In groups treated with chrysin, there was a significant reduction in the expression of genes related to apoptosis, along with improvements in markers of spermatogenesis and steroidogenesis. These positive effects are likely due to chrysin’s ability to counteract oxidative stress (decreased MDA and elevated GPx) and decrease cell death. Supporting studies by Mohammadian et al. and Jafari et al. reinforce chrysin’s regulatory role on genes associated with cell death.26,27 In addition, chrysin treatment decreased autophagy gene expression, though levels remained higher than in controls, indicating a modulating effect that lessens stress without fully stopping autophagy. Similar results were shown by Varışlı et al., who demonstrated chrysin’s ability to suppress autophagy markers during drug-induced toxicity. 28
Collectively, the data demonstrate that higher doses of chrysin more effectively reduce apoptosis and autophagy, decreasing cell death. Previous research has shown dose-dependent effects of chrysin on both cancerous and normal cells, with higher doses providing greater suppression of apoptosis and autophagy-related genes. 29 Conversely, Zhao and Robbins highlight the importance of dose optimization, as lower doses may be insufficient to combat oxidative stress, whereas higher doses more strongly activate protective cellular responses. 30 These findings support the effectiveness of the 75 mg/kg dose of chrysin used in this study, emphasizing its superior protective capacity. However, the present findings are limited by the reliance on gene-level analyses without corresponding protein-level validation and by the absence of immunohistochemical assessments, which may restrict functional and spatial interpretation of the molecular changes observed. Future studies incorporating protein expression and histological localization analyses are warranted to further substantiate these results.
CONCLUSION
This study demonstrated that melphalan induces oxidative stress and upregulates genes associated with cell death while impairing spermatogenesis and testosterone synthesis in immature rats. Chrysin administration mitigated these adverse effects by enhancing antioxidant defenses and suppressing apoptotic and autophagic gene expression. These outcomes underscore the potential of chrysin as a naturally derived therapeutic agent capable of alleviating chemotherapy-induced gonadotoxicity and improving overall reproductive health and quality of life in affected individuals.
ETHICAL APPROVAL
All experimental procedures were approved by the Ethics Committee of the Ministry of Health and Medical Education of the Islamic Republic of Iran (Approval No: IR.GMU.REC.1401.176; Date: 2023-02-21). All efforts were made to minimize animal suffering throughout the study. Also, this study adheres to internationally accepted standards for animal research, following the 3Rs principle. The ARRIVE guidelines were employed for reporting experiments involving live animals and promoting ethical research practices.
AUTHORS’ CONTRIBUTIONS
A.-H.E.: Conceptualization and methodology. S.-H.H.: Software. M.M.: Data curation and writing—original draft preparation. S.-H.A.-E.: Visualization and investigation. M.S.: Supervision and writing—reviewing and editing. Z.S.: Software and validation.
Footnotes
ACKNOWLEDGMENT
The authors are grateful to Gonabad University of Medical Sciences for cooperation in all aspects of the present study.
AUTHOR DISCLOSURE STATEMENT
The authors have no relevant financial or nonfinancial interests to disclose.
FUNDING INFORMATION
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.