
Abstract
Chronic kidney disease (CKD) is a systemic condition associated with inflammation and oxidative stress, affecting organs beyond the kidneys. Although rarely emphasized, the eyes may also be affected but underlying molecular mechanisms remain largely unexplored. The gut–kidney and gut–eye axes are emerging as therapeutic targets with prebiotics like ResistAid®-a Larch Arabinogalactan (LAG) supplement with antioxidant and immunomodulatory effects—showing promise through gut microbiota modulation. This study assessed ResistAid®’s effects on ocular gene expression in a CKD rat model. Twenty four Wistar rats were assigned to Sham (S), Sham + Treatment (ST), Nephrectomized (N), Nephrectomized + Treatment (NT) (n = 6 each). CKD was induced by 5/6 nephrectomy. The treatment was administered via gavage for 30 days at a dose of 5.35 mg/day, adapted from human recommendations. At day 30, blood and tissues were collected. Expression of antioxidant enzymes (Cat, Sod1, Gpx1, Gpx4) and other genes (Nfe2l2, Nfκb1, Timp3, Hif-1α, Kim1, Klotho) was analyzed by qPCR. Biochemical and well-being assessments were also conducted. Nephrectomy, regardless of treatment, increased Sod1 and Hif1ɑ expression in eye and blood; Specific to NT animals, ocular Gpx1, Gpx4 and Nfe2l2 expressions were markedly elevated when compared with N animals and blood Kim1 and ocular Nfκb1 expressions were not elevated, differing from N animals. No significant changes were observed between the S and ST groups. CKD induces systemic oxidative and inflammatory responses. ResistAid® partially mitigated these effects in blood and eye, suggesting systemic and local benefits, possibly via gut microbiota modulation.
INTRODUCTION
Chronic kidney disease (CKD) stands as a multifaceted health challenge with significant global impact, ranking as the 9th leading cause of death in Brazil 1 and the 10th globally.1,2 Its far-reaching implications extend beyond renal dysfunction, with recent research unveiling intriguing connections between CKD and ocular health, underscoring the need for innovative approaches to comprehensive disease management.3,4
Emerging evidence highlights the critical role of gut microbiota in modulating systemic health through, but not limited to, the gut–kidney 5 and gut–eye axes.6–8 Dysbiosis, or an imbalance in gut microbial communities, has been implicated in CKD progression, contributing to systemic inflammation, oxidative stress and metabolic disturbances. 5 Similarly, the gut–eye axis suggests that gut microbiota alterations can influence ocular health by a myriad of mechanisms, including the modulation of inflammatory and oxidative pathways linked to retinal and corneal integrity.6–8 Prebiotic supplements have been shown to modulate immune responses by promoting the growth of beneficial gut bacteria, improving gut–barrier integrity and reducing systemic inflammation. 9
Given the established role of oxidative stress in CKD pathophysiology10–13 it is proposed that a dietary supplement with antioxidative capabilities may have the ability to attenuate the progression of this disease by mitigating oxidative chain reactions, thus leading to improvements in biochemical and molecular parameters.14–17
In this pursuit, the present study scrutinizes the potential impact of ResistAid® (Lonza Ltd., Basel, Switzerland), renowned for its larch arabinogalactan (LAG)—the principal component, which has demonstrated prebiotic properties 18 phenols, and polyphenols (including the flavonoids taxifolin and quercetin), known for their antioxidant and immunomodulatory capacities,19–21 on ocular gene expression within the CKD milieu. As the ocular manifestations of CKD remain an underexplored domain, investigating the influence of this supplement on ocular gene expression promises insights beyond traditional paradigms.
The exploration of molecular intricacies through the analysis of key genes—namely, Sod1, Gpx1 and Gpx4 (antioxidant enzymes), Nfe2l2, Hif1α, and Nfkb1 (transcription factors that regulate antioxidant responses, hypoxia, and inflammation, respectively), as well as Kim1 and Klotho (emerging biomarkers of kidney damage)—aims to provide a deeper understanding of how this supplement may intersect with CKD-related ocular changes. This investigation could help elucidate its potential preventive or therapeutic effects. Not only does this research contribute to the growing body of CKD-related knowledge, but it also holds significant implications for future translational efforts in ocular and systemic health interventions.
MATERIALS AND METHODS
Animals
Twenty-four Wistar Rattus norvegicus (120–180 g) obtained from the Faculdade de Medicina do ABC (FMABC) animal facility matrices were used in this study. The animals were group-housed, maintained under a 12-h light/dark cycle, at room temperature (21 ± 2°C), and provided ad libitum access to water and rodent feed—conditions optimal for species development. All experiments in this study were approved by the Ethics Committee on Animal Use (CEUA) at the FMABC University Center (Process No. 09/2022; Annex 1) and strictly followed guidelines for the use of experimental animals regulated by CONCEA. Animals were randomly assigned to four experimental groups: sham-operated rats (Sham-“S”), sham-operated with treatment (Sham + Treatment-“ST”), animals with induced CKD (Nephrectomized-“N”), and animals with induced CKD and treatment (Nephrectomized + Treatment-“NT”), with a total of six animals per group.
Induction of CKD
All animals underwent a 5-day adaptation period before experimental procedures. CKD was induced by the subtotal nephrectomy method (5/6x), 22 with anesthesia using ketamine 80 mg/kg, IP, and xylazine 10 mg/kg IP. A midline incision in the abdominal wall was made to perform total nephrectomy of the right kidney and occlusion of the posterior and inferior branches of the left renal artery with nonabsorbable suture, causing ischemia in two-thirds of the left kidney. Sham-operated group animals underwent a sham surgery, where only the abdominal cavity was opened, and manipulation of intestinal loops occurred, followed by suture. After surgery, Tramadol 10 mg/kg/BID IM for 3 days, Meloxicam 1 mg/kg 1× daily SC for 3 days, and Cefalotin 15 mg/kg 1× daily IM for 3 days were administered.
Treatment protocol
Animals were treated from the surgical procedure (D0), daily, for 30 days, corresponding to a subchronic period. The supplement was administered via gavage of 1.5 g/day for a 70 kg adult human 18 (21.4 mg/kg/day) to 5.35 mg/day for a reference 250 g rat. The supplement was diluted in saline solution and administered in a maximum volume of 1 mL/day. The supplement was well tolerated by the animals, with no signs of toxicity or adverse effects throughout the treatment period.
We selected a 30-day period to evaluate the subchronic effects of the supplement on disease progression. This timeframe was chosen because it encompasses the critical phases of disease onset and early progression following the surgical procedure. The primary mechanisms involved in the development and establishment of the disease, particularly inflammatory and oxidative pathways, are predominantly activated during this initial postsurgical period in the animal model. Therefore, a 30-day treatment allows us to assess the early intervention potential of the supplement in modulating these key processes.
Tissue collection and RNA extraction
After euthanasia by isoflurane overdose, one eye (left) was collected and weighed on an analytical balance and blood was obtained through caudal vena cava puncture. The tissue was kept at −80°C until processing. The material was macerated using the TissueRuptorII equipment (Qiagen®, DE). For RNA extraction, 50–100 mg of tissue in 1000 μL of TRIzol reagent (TRIzol LS Reagent, Thermo Fisher, United States) were used following the manufacturer’s instructions. The extraction procedure was carried out following a standard protocol.
cDNA synthesis
RNA samples (1 μg initially) were converted into cDNA using the QuantiTect Reverse Transcription kit (Qiagen, cat no 205311), following the manufacturer’s protocol.
Gene expression analysis
The expression of Sod1, Gpx1, Gpx4, Cat, Hif-1α, Nfe2l2, Nfκb1, Kim1, and Klotho genes was evaluated by real-time PCR (qPCR). Specific primers for each selected gene were designed using the Primer3 Input 0.4.0 software, available at http://frodo.wi.mit.edu/primer3/. The primer sequences were checked for specificity using the Primer–BLAST program, available at http://www.ncbi.nlm.nih.gov/tools/primer-blast. For the relative expression normalization of the target genes, expression values of the reference gene Rpl13a will be used.
The primer sequences used for gene expression analysis are listed as follows. For Sod1, the forward primer was 5′-GCGTCATTCACTTCGAGCAG-3′ and the reverse primer was 5′-CCTCTCTTCATCCGCTGGAC-3′ (191 bp). For Gpx1, the forward primer was 5′-CCCGGGACTACACCGAAATG-3′ and the reverse primer was 5′-GTAAAGAGCGGGTGAGCCTT-3′ (223 bp amplicon). For Gpx4, the primer pair 5′-CCTCGCAATGAGGCAAAACC-3′ (forward) and 5′-CCCTTGGGCTGGACTTTCAT-3′ (reverse) (250 bp). For Cat, 5′-GCGGATTCCTGAGAGAGTGG-3′ (forward) and 5′-GAGGGTCACGAACTGTGTCA-3′ (reverse) (188 bp). For Hif-1α, the forward primer was 5′-TGTTGTCTGTGGGGTTTGCT-3′ and the reverse primer was 5′-TGCTGTGATCTGGCATTCGA-3′ (195 bp). The primers for Nfe2l2 were 5′-ATGGAGCAAGTTTGGCAGGA-3′ (forward) and 5′-CAGTGGTAGTCTCAGCCTGC-3′ (reverse) (88 bp). For Nfκb1, the forward primer 5′-AACTATGTGGGGCCTGCAAA-3′ and reverse primer 5′-TTGCAAAGCCAACCACCATG-3′ (151 bp). The Kim1 gene was amplified using the forward primer 5′-GCAGCGGTCGATACAACATA-3′ and reverse primer 5′-GAACCATCCAGGAATCTCCA-3′ (122 bp). For Klotho, the primer sequences were 5′-GTTCGAGCCCAAACCGTCTA-3′ (forward) and 5′-TGCACACGGTGTACTCTTCC-3′ (reverse) (102 bp). Finally, the reference gene Rpl13a was amplified using forward primer 5′-GTGGTCGTACGCTGTGAAG-3′ and reverse primer 5′-ACAGTGCGCCAGAAAATGC-3′ (152 bp).
Real-time PCR amplifications were performed on an Applied Biosystems 7500 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) in a final volume of 15 μL containing: 1X SYBR Green mix (Quantitec SYBR Green PCR kit, QIAGEN cat. no. 204054), 15 pmol of each specific primer, and 1.5 μL cDNA. The cycling parameters include an initial hot start step at 95°C for 10 min, followed by 45 cycles of 95°C for 15 s and 60°C for 25 s. Gene expressions and differences in expression between the analyzed groups were evaluated using the 2−ΔCq method [Pfaffl, 2001, A new mathematical model for relative quantification in real-time RT-PCR]. 23
Evaluation of biochemical parameters
In the assessment of various biochemical parameters, standard methodologies from the Clinical Analysis Laboratory of the Centro Universitário FMABC were followed. Quantifications of serum urea and creatinine were performed to evaluate renal function and determine the onset of CKD.
Serum creatinine measurement
Biochemical analyses of serum creatinine were determined using the modified Jaffé kinetic-colorimetric method.
Serum urea measurement
The ELITech kit (ELITech S.A., France) was used according to the manufacturer’s protocol.
Estimated glomerular filtration rate
The glomerular filtration rate (eGFR-μL/min) was estimated using the method recommended by Besseling and Pieters,
24
which utilizes weight (W-g), serum creatinine (C-μmol/L), and serum urea (U-mmol/L). The blood for biochemical assessments was obtained through caudal vena cava puncture following euthanasia by Isoflurane overdose.
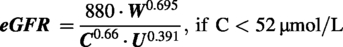
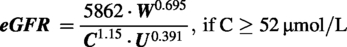
Subsequently, the eGFR values were normalized based on the estimated total body surface area (eTBSA), calculated using the method proposed by Korhonen et al.,
25
which takes into account weight (g) and a predetermined constant (k).
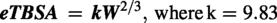
Statistical analysis
Data were expressed as mean ± standard deviation (SD). Prior to statistical comparisons, all datasets were tested for normality using the Shapiro–Wilk test and for homogeneity of variances using Levene’s test. When data met the assumptions of normal distribution and homogeneity, they were analyzed by one-way analysis of variance (ANOVA) followed by Tukey’s or Bonferroni’s post hoc test, as appropriate. For non-normally distributed data, the Kruskal–Wallis test was applied followed by Dunn’s multiple comparison test. A significance level of P < .05 was considered statistically significant. All analyses were performed using GraphPad Prism® 9.0 software.
RESULTS
To confirm the establishment of the experimental model of CKD, the eGFR (mL/min/1.73 m2) was assessed. A one-way ANOVA followed by Tukey’s multiple comparisons test demonstrated a significant reduction in eGFR values in the N group (2.608 ± 0.8663) compared with the S group (4.819 ± 0.7215, P < .0001), as well as in the NT group (2.349 ± 0.3361) compared with S (P < .0001). Furthermore, the ST group (4.106 ± 0.3335) exhibited significantly higher eGFR values than both the N (P = .0021) and NT (P = .0004) groups.
For Sod expression in the eye, a Kruskal–Wallis test followed by Dunn’s multiple comparisons test revealed a significant increase in the N group (29.36 ± 19.50) compared with S (4.567 ± 2.955, P = .0261), as well as in the NT group (46.57 ± 28.59) compared with S (P = .0059) and ST (5.330 ± 1.091, P = .0263). In the blood, Sod expression was significantly higher in the N group (55.09 ± 12.30) compared with both S (9.152 ± 14.97, P = .0061) and ST (11.56 ± 14.34, P = .0184), with no other significant differences among the remaining comparisons.
For Gpx1 expression in the eye, significantly higher levels were observed in the NT group (51.16 ± 6.503) compared with S (4.875 ± 1.744, P = .0034) and ST (4.917 ± 1.922, P = .0025). In the blood, the ST group (3.992 ± 1.874) presented lower levels compared with N (14.00 ± 4.369, P = .0087) and NT (14.69 ± 4.303, P = .0042). Regarding Gpx4 expression in the eye, the NT group (9.354 ± 1.682) exhibited significantly higher levels compared to both S (2.228 ± 1.170, P = .0107) and ST (2.215 ± 0.9164, P = .0122). In the blood, however, Gpx4 expression did not differ significantly among groups (S: 2.748 ± 1.356; N: 5.430 ± 2.928; ST: 2.803 ± 0.8185; NT: 5.855 ± 2.234; Fig. 1).
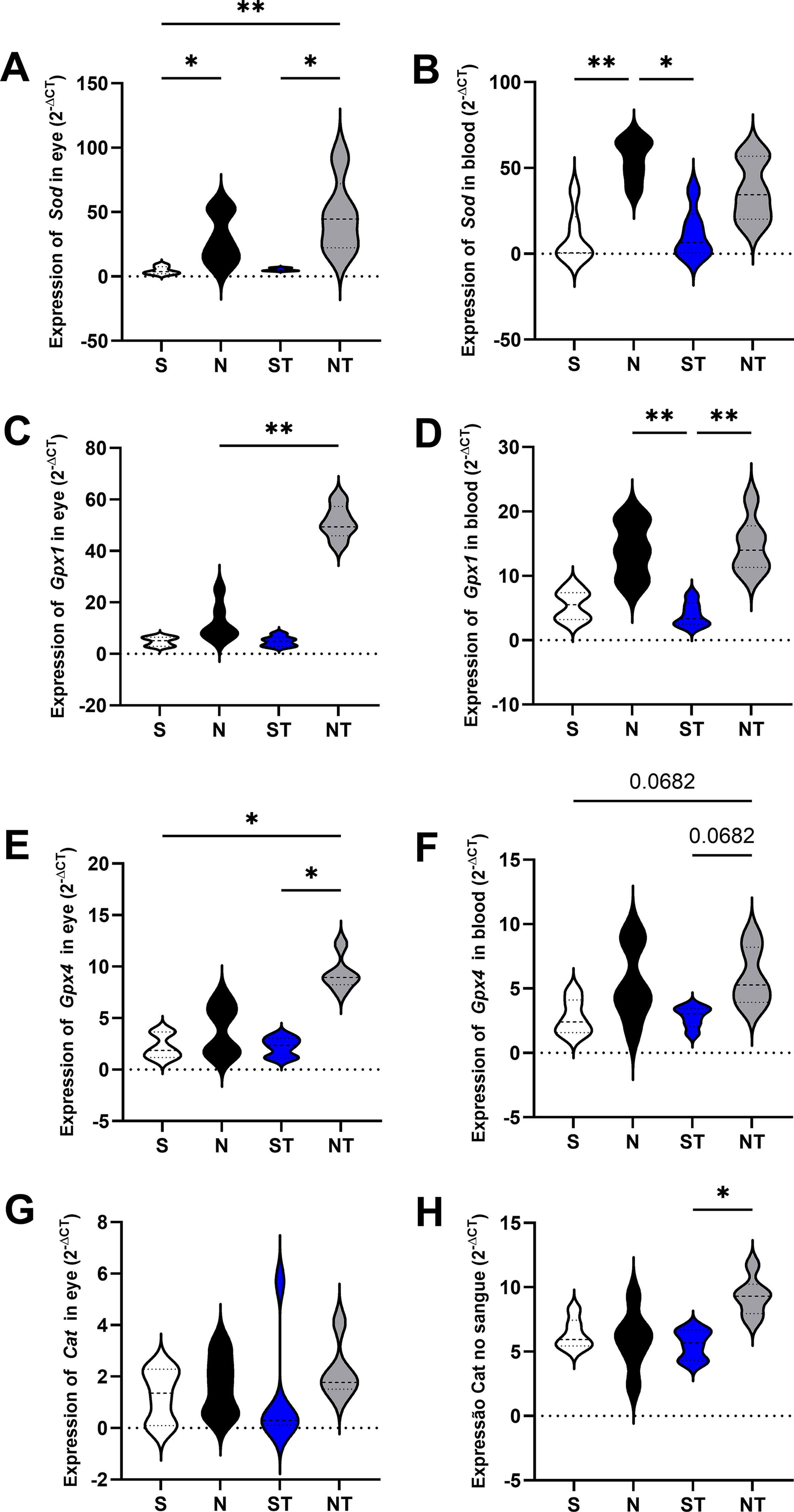
Gene expression of the antioxidant enzymes SOD, GPX1, and CAT in ocular and blood tissues of the experimental groups. The figure shows the expression of SOD in the eye
Hif expression in the eye was significantly higher in the N group (20.94 ± 9.068) compared to S (1.220 ± 0.7431, P = .0318), and in the NT group (34.82 ± 14.07) compared to both S (P = .0032) and ST (1.258 ± 0.6316, P = .0154). In the blood, a similar pattern was observed, with increased expression in N (6.865 ± 1.666) and NT (7.175 ± 3.532) relative to S (0.7317 ± 0.6549, P = .0115 and P = .0049, respectively), and higher levels in NT compared with ST (1.107 ± 0.4139, P = .0330) (Fig. 2).
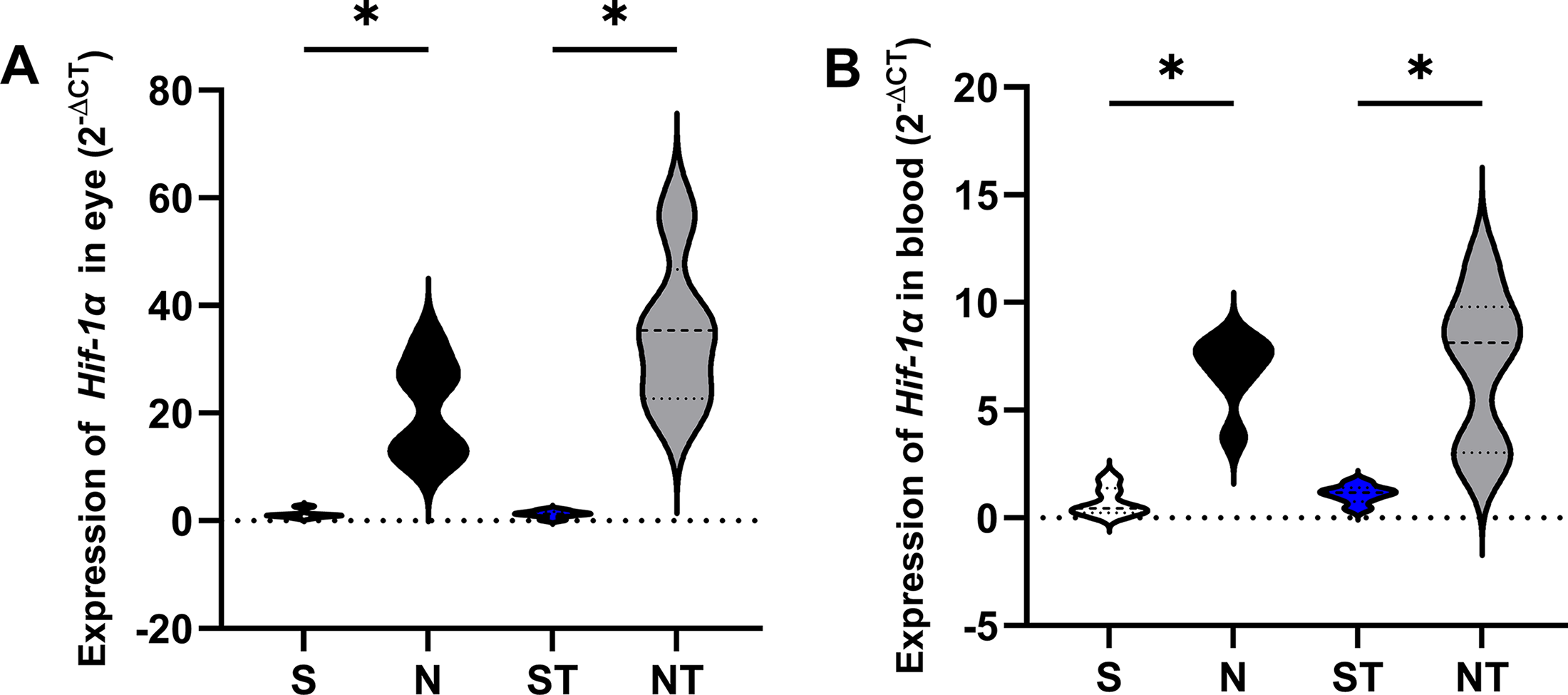
Relative expression of Hif1α in eye and blood tissues across experimental groups. Gene expression levels were analyzed using the 2^-ΔCq method. In the eye
For Kim1 expression in the eye, no significant differences were detected (S: 0.1419 ± 0.2452; N: 28.06 ± 38.27; ST: 3.817 ± 9.141; NT: 5.568 ± 12.19). In the blood, the N group (42.37 ± 56.14) showed significantly higher levels compared to S (0.6265 ± 0.9083, P = .0286) and ST (0.1191 ± 0.1212, P = .0209) (Fig. 3).
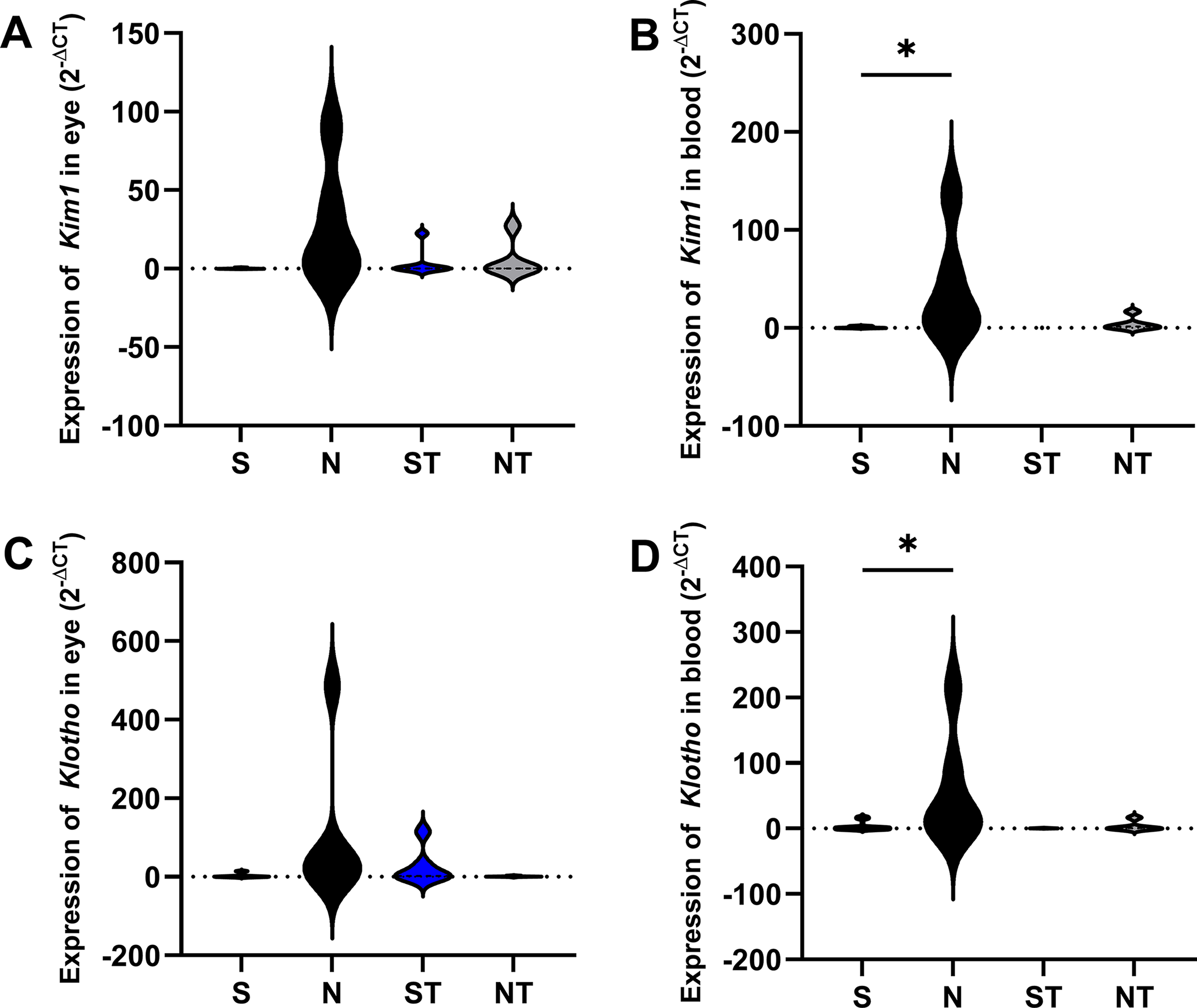
Gene expression of Kim1 and Klotho in ocular and blood tissues of animals from the S (sham), N (nephrectomized), ST (sham treated with Resistaid), and NT (nephrectomized treated with Resistaid) groups. Panel
Nfe2l2 expression was significantly elevated in the eye of NT animals (57.06 ± 65.56) compared with S (0.3287 ± 0.5508, P = .0172), with no other pairwise comparisons reaching statistical significance (S vs. N, P = .1108; S vs. ST, P > .9999; S vs. NT, P = .3259; N vs. ST, P = .2290; N vs. NT, P > .9999; ST vs. NT, P = .6447). In the blood, no significant differences were observed among the groups (S: 0.8432 ± 0.9175; N: 12.19 ± 13.26; ST: 1.825 ± 1.757; NT: 5.842 ± 5.721). The analysis of Nfkb1 expression in the eye revealed significantly higher levels in the N group (50.89 ± 10.44) compared with S (0.1105 ± 15.47), ST (0.5809 ± 10.11), and NT (2.301 ± 6.494), as determined by Kruskal–Wallis (H = 8.1433, P = .0431) followed by Dunn’s test. In the blood, the N group (34.12 ± 40.40) showed significantly higher Nfkb1 expression compared with S (2.108 ± 2.237, P = .0499). No other significant differences were found (S vs. ST, P > .9999; S vs. NT, P = .0664; N vs. ST, P = .4430; N vs. NT, P > .9999; ST vs. NT, P = .5031; Fig. 4).
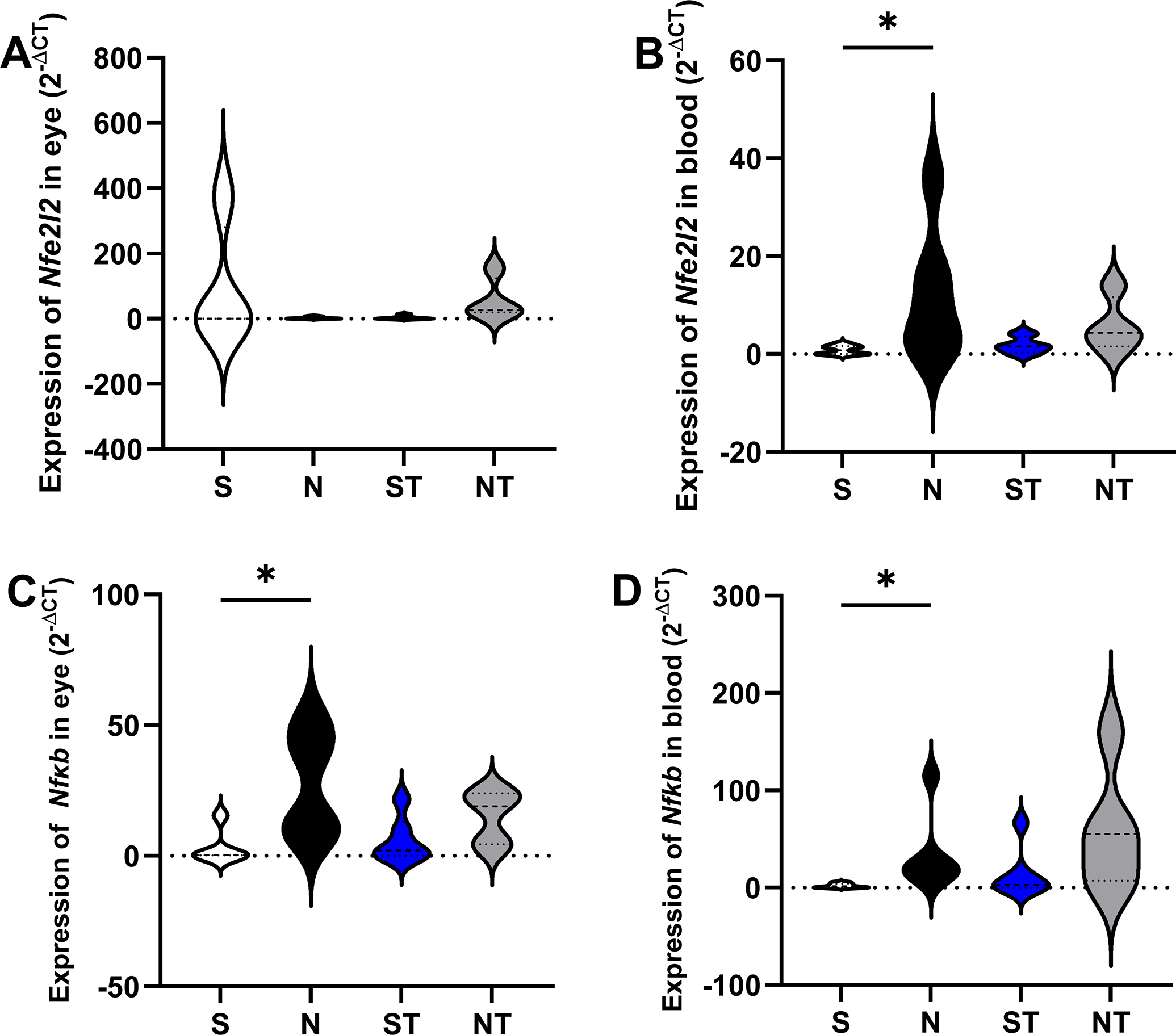
Gene expression of Nfkb1 and Nfe2l2 in ocular and blood tissues of animals from the S (Sham), N (nephrectomized), ST (Sham treated with Resistaid), and NT (nephrectomized treated with Resistaid) groups. Panel
DISCUSSION
The findings of this study provide significant insight into the molecular mechanisms underlying CKD and the potential therapeutic effects of antioxidant and prebiotic supplementation. By analyzing gene expression patterns in both blood and ocular tissues, we have highlighted systemic and tissue-specific changes related to oxidative stress, inflammation and hypoxia. These findings contribute to a better understanding of CKD pathophysiology and its systemic impact, particularly on ocular health.
CKD is well known to trigger systemic inflammation and oxidative stress, in part due to retention of gut-derived uremic toxins. In our model, CKD promoted changes in blood gene expression patterns, such as a pronounced upregulation of Nfe2l2 (encoding Nrf2) and key downstream antioxidant enzymes (namely Sod1, Gpx1, Gpx4, and Cat) in the nephrectomized group (N), consistent with a heightened but functionally active antioxidant response. This pattern suggests that, at least within the observed time frame, the organism mounted an effective transcriptional response to oxidative stress, likely via Nrf2-mediated signaling. These findings align with previous reports linking oxidative stress to CKD progression10,24 and reflect an adaptive, compensatory mechanism to counteract oxidative damage.
Additionally, inflammatory markers, such as Nfκb were elevated in both nephrectomized groups, irrespective of supplementation (N and NT), illustrating a state of systemic stress in which both proinflammatory and antioxidant defenses are upregulated. The role of Nfκb in modulating immune responses and its association with inflammatory diseases further supports its importance in CKD pathology. 25
Moreover, the consistent upregulation of Hif-1ɑ, a transcription factor responsive to reduced oxygen availability, across both nephrectomized groups suggests that CKD triggers systemic hypoxic signaling, confirming other groups’ findings.26–28 Given the relatively short duration of our experimental model (30 days), it’s likely that early proinflammatory states and microvascular/endothelial impairment underlie this hypoxic response.26–28 This interpretation is further substantiated by the elevated Nfκb1 expression, indicating activation of inflammatory pathways, which are well known to exacerbate tissue hypoxia and oxidative stress.
Supplementation with LAG and polyphenol-rich extracts (in the form of ResistAid™) modulated these systemic effects. In the NT group, even though Nfe2l2 expression levels were not significantly affected, antioxidant enzyme expression (Sod1, Gpx1, Gpx4) remained elevated, while Cat (whose expression is not modulated by this transcription factor 29 levels increased, suggesting enhanced oxidative stress management, potentially mitigating systemic oxidative injury. The absence of significant changes in the Treated Sham (ST) in regard to the Sham (S) group suggests that this supplementation does not alter antioxidant activity in the absence of CKD-induced stress.
The expression of markers of kidney damage and repair offered further insight into CKD pathophysiology and the potential benefits of dietary interventions. Kim1, a well-established indicator of renal tubular injury, 30 was, as expected, elevated in the blood of the nonsupplemented group (N) but, surprisingly, not in the supplemented group (NT). This divergence indicates a potential protective effect of the supplementation regimen. Similarly, Klotho, a protein associated with antiaging and repair mechanisms, was upregulated in the CKD environment (N group), pointing to an involvement of this protein in counteracting CKD-related damage.31–33 Intriguingly, the NT group displayed expression levels similar to those of the healthy (sham) animals. This normalization may indicate that the supplement attenuates the degree of renal injury and stress, thereby reducing the physiological need for Klotho-mediated reparative responses.31–33 Together, these findings suggest that dietary intervention not only mitigates renal injury but also contributes to the preservation of homeostatic signaling pathways involved in repair and regeneration.
Gene expression patterns in ocular tissues of CKD rats largely mirrored those observed in blood, indicating that CKD exerts systemic effects that extend to the eyes. Elevated Sod1, Hif-1ɑ, Kim1, and Klotho expression levels in ocular tissue of CKD rats reflect shared stressors such as oxidative stress and hypoxia.3,4 However, there seemed to be lower levels of Nfe2l2 in the eyes, coupled with significant variability; this lower and unstable activation of the oxidative stress response regulator may underlie the observed decrease in Gpx1 and Gpx4 expression in nephrectomized but not treated (N) rats, suggesting tissue-specific blunted antioxidant response.
Notably, the expression of Gpx1 and Gpx4 in the eyes increased when nephrectomized rats were exposed to the supplement (NT group), in accordance with the blood data, and the Cat levels, differently from what was seen in blood, were in line with those of Sham rats. This could possibly be explained by the fact that Cat transcription, unlike Gpx1 and Gpx4, is not linked to Nfe2l2, relying instead on alternative pathways. 29
These patterns illustrate the complexity of the interaction between hypoxia and antioxidant defenses. Hypoxia triggers a cascade of cellular adaptations designed to mitigate the oxidative stress arising from the elevated reactive oxygen species (ROS). Central to this response is Hif1-ɑ, a master regulator of hypoxia-induced gene expression, which promotes both metabolic adaptation and the induction of antioxidant proteins, such as heme oxygenase-1 (HO-1) and metallothioneins.34,35 Experimental models show that hypoxia often leads to increased ROS production, potentially causing damage unless tightly counteracted by a coordinated upregulation of antioxidant enzymes—SOD, CAT, and GPX among them.36,37 Importantly, transcription factors such as Nrf2 integrate these signals, mediating spatial and temporal regulation of antioxidant responses. For instance, hypoxic preconditioning has been shown to activate Nrf2 in skeletal muscle, enhancing antioxidant capacity and physiological resilience. 35
These findings underscore a delicate balance: while moderate ROS levels serve essential signaling functions, excessive ROS can drive oxidative injury and inflammation. 38 Hence, although antioxidant supplementation may reduce oxidative damage, it could also interfere with endogenous adaptive mechanisms if not finely tuned. 39 In this context, the restoration of Gpx1 and Gpx4 expression in the NT group suggest that LAG and polyphenol supplementation may enhance antioxidant defences without disrupting physiological hypoxic signaling, enabling a more adaptive and protective redox state in ocular tissues.
Of particular interest, Nfκb1 expression in ocular tissue was reduced in the NT group, compared to the nontreated CKD group (N), raising the possibility of local modulation of the inflammatory pathways by the LAG supplement. This aligns with previous studies showing that coupons such as Quercetin, 40 Polyunsaturated fatty acids (PUFAs) such as Docosahexaenoic Acid (DHA) and Eicosapentaenoic Acid (EPA), 41 β-Cryptoxanthin, 42 and RS9, 43 can modulate Nfκb1 expression, and thus the inflammatory response, or NFE2L2 expression, enhancing oxidative stress resistance—or both. To our knowledge, this is the first time gene expression pattern modulation has been shown on eye tissue in response to supplementation, be it with antioxidants, prebiotics, or other substances, highlighting a novel avenue for therapeutic intervention aimed at ocular protection.
Emerging research highlights the role of the gut microbiota in CKD pathophysiology, particularly through its interaction with the gut–kidney axis. Dysbiosis, characterized by an imbalance in gut microbial populations, has been associated with increased production of uremic toxins such as p-cresyl sulfate, indoxyl sulfate, and trimethylamine-N-oxide, all of which contribute to systemic inflammation, oxidative stress, and renal dysfunction.5,44 The intestinal barrier is often compromised in CKD, leading to increased gut permeability—an effect commonly referred to as “leaky gut,”-which facilitates the translocation of endotoxins and inflammatory mediators, further exacerbating renal injury.5,44 In this context, our findings dovetail with the growing recognition of the gut–organ axes in CKD.
Prebiotic interventions, such as LAG, have shown promise in modulating gut microbial composition, enhancing populations of beneficial bacteria like Bifidobacterium and Lactobacillus, while reducing opportunistic pathogens. 18 LAG and polyphenols may function synergistically as fermentable fibers and microbial modulators, enriching populations of bacteria that produce short-chain fatty acids (SCFAs), which exert systemic anti-inflammatory effects, potentially mitigating CKD progression and associated complications, and attenuating proteolytic fermentation. Additionally, polyphenol-rich dietary components and probiotic supplementation have been investigated for their ability to attenuate gut-derived inflammation and preserve renal function. 18
Beyond the gut–kidney axis, mounting evidence suggests an interconnection between gut microbiota and ocular health, forming a gut–eye axis. Dysbiosis-driven systemic inflammation and circulating microbial products may reach distal tissues, including the retina, where they can contribute to ocular pathologies, including diabetic retinopathy, age-related macular degeneration, and glaucoma.6–8 SCFAs, particularly butyrate, have been implicated in neuroprotection and may help regulate the inflammatory processes associated with retinal degeneration. 45 Furthermore, gut-derived metabolites, including those influenced by prebiotics such as LAG, can modulate blood–retinal barrier integrity and immune homeostasis, thereby influencing disease progression in both renal and ocular conditions.5–8,44,46,46
Our study provides functional support for this emerging axis: Retinal gene expression analysis revealed reduced inflammation (Nfκb1) signaling alongside enhanced antioxidant responses (Gpx1, Gpx4) in the LAG-treated group (NT). While the exact signaling pathways remain to be elucidated the emerging gut–eye framework provides a plausible mechanistic explanation for the observed ocular benefits.
A limitation of this study is the use of an animal model, which may restrict the direct extrapolation of the results to human physiology. Therefore, clinical trials are necessary to confirm the effects of LAG in patients with CKD. Moreover, additional research is required to elucidate the specific microbial and metabolic pathways involved in the gut–kidney and gut–eye axes, as well as to define the most effective therapeutic strategies. Beyond these limitations, we acknowledge that this study was limited to gene expression analyses, and mRNA levels do not necessarily reflect protein abundance or functional outcomes. Histological and biochemical assessments were not included, as this experiment was conceived as a molecular screening approach aimed at identifying early transcriptional targets involved in oxidative and inflammatory pathways in CKD animals treated with LAG. Future studies are being planned to integrate these molecular findings with histopathological and biochemical analyses to confirm whether the observed gene expression changes translate into functional and morphological improvements.
Nonetheless, the current findings provide a mechanistic rationale for the reported immunomodulatory benefits of ResistAid® and support the gene expression changes observed in this study.
CONCLUSIONS
These findings highlight the systemic and ocular implications of CKD, particularly the interplay between oxidative stress, inflammation, hypoxia, and dysbiosis. CKD-associated ocular changes could contribute to visual impairment, underscoring the importance of understanding systemic-ocular connections in this disease. Further research should also investigate the role of gut–microbiota in mediating these effects, paving the way for integrated therapeutic approaches targeting CKD, eye conditions, and its systemic manifestations. The protective effects of LAG supplementation on oxidative (e.g., Sod1, Gpx1, Gpx4, Cat, Nfel2l) and inflammatory markers (e.g., Nfκb) suggest a potential utility in mitigating CKD-related complications, although translational research is still needed.
AUTHORS’ CONTRIBUTIONS
P.H.A.R.: Conceptualization (equal), writing—original draft (lead), methodology (equal). I.P.D.: Methodology (equal). I.B.L.: Formal analysis (equal). G.P.: Methodology (equal). E.F.S.: methodology (equal). I.D.R.T.: Methodology (equal); B.C.A.A.: Conceptualization (equal) and formal analysis (equal). V.L.L.: Conceptualization (supporting) and writing—original draft (supporting). G.L.V.: Conceptualization (equal) and formal analysis (lead). F.L.A.F.: Conceptualization (lead) and writing—review and editing (equal).
Footnotes
ACKNOWLEDGMENTS
The authors express sincere gratitude to Lonza (Basel, Switzerland) for their generous provision of the ResistAid™ supplement free of charge for experimental use, and to MSc. Andrey Lopes for facilitating access to the compound and supporting the logistics of its acquisition.
AUTHOR DISCLOSURE STATEMENT
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. However, it should be noted that Lonza (Basel, Switzerland) provided the ResistAid™ supplement free of charge for experimental use.
FUNDING INFORMATION
This study was partially funded by a grant from FAPESP (grant no.