
Abstract
Objectives
The aim of this study was to investigate the spatial distribution of neutrophil extracellular traps (NETs) in cardiogenic arterial thromboembolism (CATE). Specifically, we aimed to examine the related structural features of NETs in feline arterial thrombi in relation to their arterial locations.
Methods
Paraffin-embedded aortic bifurcations from nine cats with hypertrophic cardiomyopathy (four with CATE and five without) were deparaffinized, and NETs were identified by immunodetection based on colocalization of cell-free DNA, citrullinated histone H3 and neutrophil elastase. The distribution of NETs in thrombi within the aortic bifurcations and common iliac arteries (CIAs) was compared based on their proximity to the descending aorta (proximal, mid, distal). Ten random fields per section were captured at × 10 and × 20 magnification for each section of the clot and analyzed.
Results
The distributions of NETs in thrombi within the aortic bifurcation and CIAs were found to differ in relation to their assigned zones (proximal, mid, distal; P = 0.04); NETs were concentrated mostly in the proximal region in the aortic bifurcations (47.56%, interquartile range [IQR] 14.07–77.95) and CIAs (44.69%, IQR 24.65–85.28), compared with the distal regions (2.69%, IQR 0.10–50.04 [P = 0.027]; 7.08%, IQR 1.27–59.33 [P = 0.02]).
Conclusions and relevance
The variation in NET distribution within arterial thrombi may shed light on the pathogenesis of thrombus growth. This may be due to possible neutrophil entrapment or variations in shear stress.
Keywords
Introduction
Cardiogenic arterial thromboembolism (CATE), a condition associated with high mortality and morbidity in cats, has an incidence of 11.3% in 1008 cats with hypertrophic cardiomyopathy (HCM). 1 Despite the devastating outcome, with a survival rate of <40%, veterinarians have limited tools to identify and treat cats at risk of CATE, as many of these cats are asymptomatic.
The pathophysiology of CATE in cats is complex and poorly understood. Decreased left atrial (LA) function causing blood stasis in the LA and LA appendage has been proposed to facilitate the formation of thrombus, which eventually embolizes to distal arteries, occluding blood flow to tissues. 2 This leads to hemodynamically significant stenosis or occlusion of the distal aorta or arteries subjecting blood cells to pathologic shear stress, which is the frictional force exerted on the endothelium. In other species, extreme variations in shear stress, such as those seen in CATE, have been demonstrated to augment platelet–neutrophil interaction and the formation of neutrophil extracellular traps (NETs). 3 NETs, which are released by activated neutrophils in response to systemic inflammation or pathogen exposure, are composed of cell-free DNA (cfDNA) decorated with histones and neutrophil proteins. 4 In mice, dogs and humans, NETs are known to propagate clot formation by activating the intrinsic pathway of coagulation, facilitating the binding of platelets and blood cells, and inhibiting fibrinolysis.5–7 We recently identified NETs in paraffin-embedded feline arterial thrombi by immunofluorescence microscopy. 8 Similar to ischemic stroke thrombi in humans, feline arterial thrombi consist of NETs within all layers of the thrombus structure, but the cellular composition and distribution of NETs remain unknown. 8 A better understanding of how NETs contribute to CATE would bring us a step closer to expanding our understanding of this devastating disease. If NETs are proven to facilitate thrombus growth in these cats, novel therapies targeting NET formation or dismantling NET structures may be potential therapeutic options.
Herein, we aimed to determine if NET structures in arterial thrombi retrieved from cats with CATE vary in relation to their locations in the aortic bifurcation and iliac arteries.
Materials and methods
This study was approved by the Institutional Animal Care and Use Committee at the University of California, Davis (IACUC 20565). Necropsy samples from four client-owned cats treated at the Veterinary Medical Teaching Hospital of University of California, Davis, for a diagnosis of HCM and CATE were selected for this study. Necropsy samples from research colony cats that were subjects of an unrelated study of HCM, were selected for the control group.Antemortem diagnosis of CATE was made based on at least four of the following clinical signs: (1) sudden-onset of vocalization, paralysis or paresis of one or more limbs; (2) low rectal temperature (<98°F or 37°C); (3) lower motor neuron signs in at least one limb; (4) absent dorsal pedal pulse confirmed by absent Doppler signal; and (5) pale or cyanotic foot pads or nailbed in the affected limb(s). Ante-mortem echocardiography was performed within 72 h of euthanasia and necropsy for all cats. The presence of cardiomyopathy in all cats was confirmed by echocardiogram performed by a board-certified cardiologist. HCM was defined as previously described. 9 Echocardiographic LA enlargement was defined by a LA-to-aortic (LA:Ao) ratio >1.5 in the right parasternal short-axis view, as previously described.10,11 Severe LA enlargement was documented if the LA:Ao exceeded 2.0. The presence or absence of spontaneous echocardiographic contrast (SEC) as a marker of presumed blood stasis, as well as the presence or absence of an observed intracardiac thrombus, was recorded for each cat by the echocardiographic examiner. The aortic bifurcation, including the descending aorta, femoral and common iliac arteries (CIAs), were bluntly dissected and extracted at necropsy within 24 h of euthanasia, as previously described. 8 Tissues were fixed, paraffin-embedded and stored at –20°C until analysis. After deparaffinization, the aortic sections underwent heat-induced antigen retrieval and permeabilization, followed by the immunodetection of NETs, as previously described. 8 In brief, deparaffinized clot sections were stained for citrullinated histone H3 (citH3) and neutrophil elastase (NE) by antibodies, previously shown to cross-react in cats. DNA was stained using 4′,6-diamidino-2-phenylindol (DAPI).
Following immunolabelling, each slide was randomly assigned a number to ensure that the operators were blinded to the ante-mortem diagnosis of the respective necropsy sections. Immunofluorescence microscopy of the aortic bifurcations and CIAs was performed using an EVOS FL Cell Imaging System (ThermoFisher Scientific, Waltham, MA). The location of each thrombus, identified as a discrete intraluminal structure in close proximity to the endothelium, was documented by scanning the entire length of the aortic bifurcation and CIAs using phase contrast microscopy. The longitudinal length of thrombi within the aortic bifurcation and CIAs was then equally divided into three zones (distal, mid and proximal) based on proximity to the descending aorta (Figure 1). Ten random fields in each zone selected by the operator were captured at × 10 and × 20 magnifications. Thrombi that spanned the entire length of the artery were divided into the three zones based on the arterial location. When clots were found in both CIAs, the NET distribution in each zone of both arteries was determined and averaged. To maintain consistency throughout the image-acquisition process, the exposure of each fluorescence channel (DAPI Channel Ex, Texas Red excitation = 357/44 nm, emission = 628/32 nm; green fluorescent protein, excitation = 470/22 nm, emission = 525/50 nm) was kept constant. Combined phase contrast and immunofluorescent images of each zone (Figure 2a,c) were used to further identify cell types and NET structures (Figure 2a,b). NETs were identified based on the colocalization of cfDNA, citH3 and NE by available software (Fiji; National Institutes of Health), as previously described.8,12 The percentage distribution of NETs in each zone of a thrombus was calculated using the following formula:

For the purpose of this study, thrombi that appeared to be separated as a result of artifact were analyzed as a single thrombus. In control sections without visible thrombi, the distribution of perivascular NETs was calculated using the following formula:
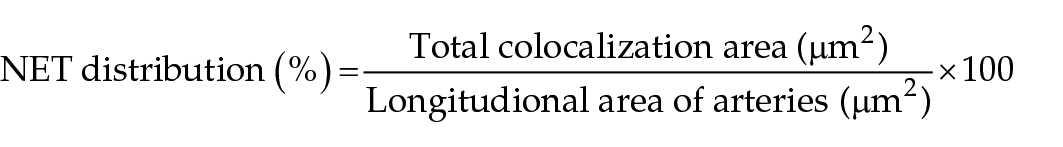

Schematic diagram illustrating how thrombus location was assigned to one of the three zones (A = proximal; B = mid; C = distal), based on proximity to the descending aorta

Representative immunofluorescence images of aortic bifurcations collected from (a,b) a cat with cardiogenic arterial thromboembolism and a cat (c,d) with hypertrophic cardiomyopathy (HCM) without arterial thrombosis. Tissue was fixed, deparaffinized and antigen retrieved by heat induction, blocked and stained for cell-free DNA (DNA, blue), neutrophil elastase (NE, green) and citrullinated histone H3 (citH3, red). (a) Phase contrast microscopy demonstrated the presence of a thrombus composed of erythrocytes (*) surrounding a clump of neutrophils (arrow). Scale bar = 400 μm. (b) Respective immunofluorescence image demonstrating the presence of neutrophils with the expression of intracellular citH3 and NE. Individual panels demonstrating the colocalization of cell-free DNA, extracellular NE and citH3. Original magnification × 10. No thrombus or NETs were found in either (c) phase contrast or (d) immunofluorescence images in a cat without clinical signs of arterial thrombosis. Original magnification × 20. Scale bar = 200 µm
Statistics
The normality of continuous data was tested using visual inspection of histograms and confirmed by Shapiro–Wilk testing. NET distribution (%) in the proximal, mid and distal aspects of thrombi in the aortic bifurcation and CIAs was compared using a Friedman’s test, followed by a Dunn’s post-hoc multiple comparisons test. Unpaired and paired data were compared using the Mann–Whitney test and Wilcoxon signed rank matched-pairs test, respectively. The coefficient of variation (CV) between different thrombus zones was calculated. Data were presented as median and interquartile range (IQR). A P value <0.05 was considered significant.
Results
Cats with CATE had a median age of 8.5 years (range 7–10) and were older than cats in the control group (median age 5 years, range 4–5; P = 0.008). All cats in the CATE group were male neutered. Two of the five cats in the control group were intact females and 3/5 were intact males. Ante-mortem echocardiography revealed that all nine cats in the study met standard echocardiographic criteria for the diagnosis of HCM. 9 All cats with CATE also had severe LA enlargement (LA:Ao >2.0) and SEC within the LA on echocardiography. A single cat with CATE also had an LA appendage thrombus in situ observed by echocardiography. Two of five control cats with HCM had mild LA enlargement; the remaining three had normal LA size by echocardiography. None of the control cats had any evidence of SEC or LA thrombi.
All cats with CATE had thrombi that spanned the aortic bifurcation and CIAs. By including a control group, we demonstrated that post-mortem changes could lead to small amounts of perivascular NETs (4/5 control cats) without the formation of organized thrombi (see Figure 1 in the supplementary material). We also showed that fixation, deparaffinization and immunostaining did not lead to significant autofluorescence (Figure 2d). The median NET distribution in control sections was 0.79% (IQR 0.31–5.49), which was significantly lower than those found in thrombi within the aortic bifurcations (39.76%; IQR 5.73–82.87 [P = 0.032]) and CIAs (30.94%; IQR 25.53–74.20 [P = 0.016]) in cats with CATE (Figure 3a). The overall NET distribution did not differ when comparing thrombi from the aortic bifurcations with the CIAs (P >1.00).
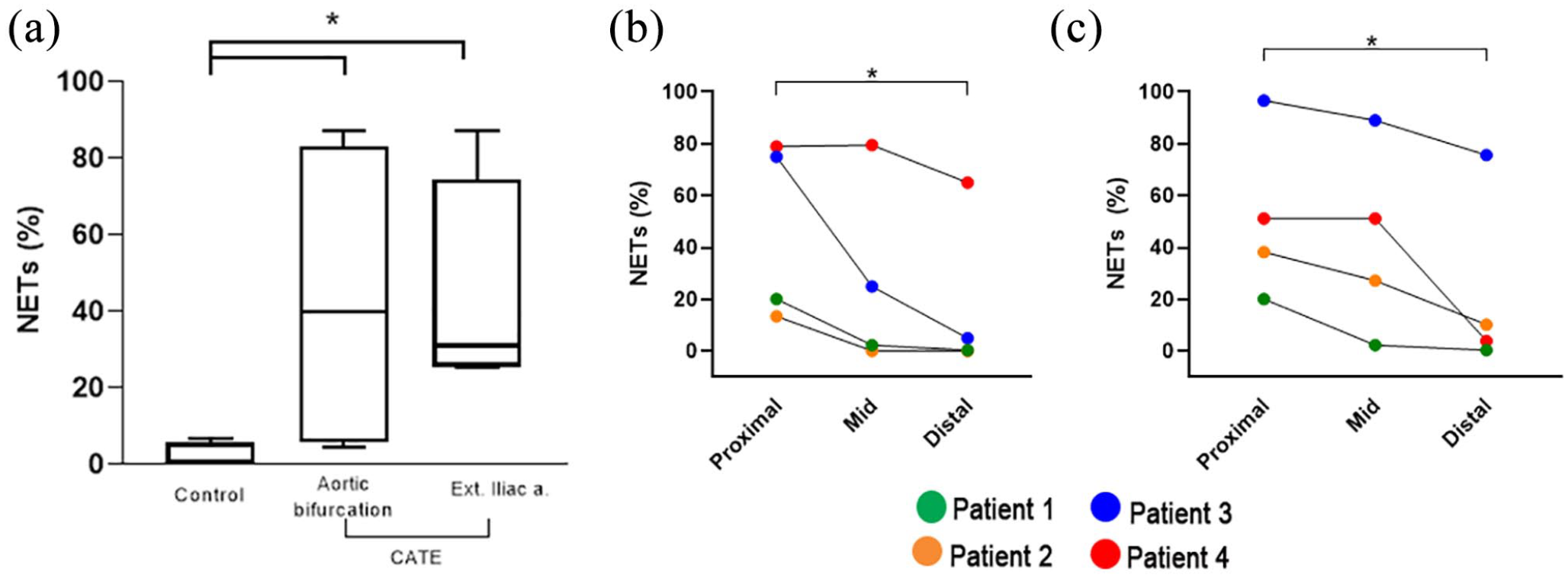
(a) Box-and-whisker plot of neutrophil extracellular trap (NET) composition (%) in thrombi found in the aortic bifurcations and common iliac arteries (CIAs) in 4 cats with cardiogenic arterial thromboembolism (CATE), and of perivascular NETs in the aortic bifurcations of 5 control cats. The middle line represents the median and boxes represent the 25th and 75th percentiles. Whiskers represent ranges. (b,c) Connected dot plots of the median NET distribution in thrombi found in three anatomical locations (proximal, mid and distal) of the (b) aortic bifurcation and (c) CIAs in four cats with CATE. *P <0.05
NET distributions in thrombi within the aortic bifurcation and CIAs were found to differ in relation to their assigned arterial regions (ie, proximal, mid, distal; P = 0.04). We found that the distal sections of thrombi consisted mostly of anucleated cells such as erythrocytes. The proximal zone of thrombi within the aortic bifurcations consisted of significantly more NETs (47.56%; IQR 14.07–77.95) than the distal zone (2.69%; IQR 0.10–50.04 [P = 0.027]) (Figure 3b), although the extent of NETs varied greatly among cats with a CV of 180.1% and 74.5%, respectively. Similarly, NETs were the predominant structural component of thrombi found in the proximal (44.69%; IQR 24.65–85.28) CIAs, compared with the distal CIAs (7.08%; IQR 1.27–59.33 [P = 0.02]), with CVs of 158.1% and 63.37%, respectively (Figure 3c). Thrombi within the mid-sections of the aortic bifurcation and CIAs comprised a mix of anucleated cells and NETs (Figure 4a,b).

Representative immunofluorescence images of a thrombus within the mid-section of the common iliac artery from a cat with cardiogenic arterial thromboembolism (CATE). Tissue was fixed, deparaffinized and antigen retrieved by the heat-induction method, then blocked and stained for cell-free DNA (cfDNA, blue), neutrophil elastase (NE, green) and citrullinated histone H3 (citH3, red). The thrombus within this section was composed of large clusters of neutrophil extracellular traps (NETs; encircled in [a]) within an area of nucleated cells. The thrombus was also composed of anucleated cells (aNc), as shown in the respective phase contrast image (b). Note the thinly interwoven strands of NETs within areas of anucleated cells (arrow heads in [a]). Individual panels demonstrating the colocalization of cfDNA, extracellular NE and citH3 in NETs. Original magnification × 10. Scale bar = 400 µm
Discussion
Using a novel method for the identification and quantification of NETs in paraffin-embedded aortic bifurcations, we demonstrated that NETs were structural components of arterial thrombi in cats with CATE. NETs were predominantly identified in proximal regions of thrombi and this unique distribution decreased as the imaging was performed distally.
CATE occurs when an intracardiac thrombus embolizes to the aortic bifurcation or peripheral arteries, causing decreased flow of oxygenated blood to tissues distal to the mechanical obstruction. 13 Cats with cardiomyopathies are prone to developing intracardiac thrombus in the LA and/or LA appendage. Blood stasis and endothelial dysfunction secondary to LA systolic dysfunction and LA dilatation facilitate blood-to-protein interactions and subsequent thrombogenesis.2,13 The presence of SEC, characterized by dynamic smoke-like echoes in the LA, is found in a majority of cats with CATE. 2 The exact mechanism of SEC formation is unclear in cats. In humans, it is believed to be caused by ultrasonographic scattering secondary to erythrocyte aggregates that are held together by fibrinogen in low-shear environments. 14 We found that thrombi within the distal arteries, where embolism first occurred, were composed predominately of erythrocytes with minimal leukocytes. This further supports the notion that intracardiac thrombi, which form secondary to blood flow stasis, are composed primarily of erythrocytes and fibrinogen. The relative absence of white blood cells, especially neutrophils, likely explains the relative lack of NETs in the distal region of thrombi. We discovered that the distribution of NETs in each of the thrombus zone varied greatly among the four cats with CATE. This high degree of variability could be secondary to the extent of thrombosis, which was found to be most severe in patients 3 and 4. However, given the small number of cats studied, this heterogenicity could also be caused by artefacts introduced during the fixation process. cfDNA within thrombi, which cannot be directly fixed by paraformaldehyde, could be washed away during the dehydration and paraffin embedding process. 15
We also observed that the composition of the arterial thrombi changes in relation to their proximity to the distal aorta, where disruption to laminar blood flow caused by partial or complete occlusion of the arteries could dramatically elevate hydrodynamic forces and shear stress. In addition to the physical entrapment of neutrophils near the site of vascular occlusion, the exposure of neutrophils to extreme variations in shear stress between the heart and aortic bifurcation may also activate neutrophils to release NETs. Yu et al utilized microfluidics to demonstrate that rapid elevation in shear stress and pressure gradient generated by clot-mediated stenosis could trigger neutrophils to release NETs. 3 We therefore hypothesize that while LA dysfunction leads to the formation of a fibrin-rich thrombus decorated mostly with red blood cells in the LA, composition of the thrombus changes drastically following embolism. This transition from a NET-deficient clot to one with high concentrations of NETs within the proximal region of the arteries may have important implications in the prevention and treatment of CATE. NETs may further propagate thrombus growth in cats with CATE as NETs play a direct role in promoting thrombosis. NETs not only possess a polyanionic surface that can readily activate contact phase proteins such as factors XII and XI to initiate the intrinsic cascade, but they also modulate clot remodeling by inhibiting fibrinolysis.16,17 The NET-rich portion of the thrombus is likely resistant to fibrinolysis and may hence explain the failure of fibrinolytic therapy in improving the outcome in cats with CATE. 18 In addition, high concentrations of NETs have been found to induce vascular occlusion independent of fibrin and von Willebrand factors. 19 Therapies such as DNase I that can digest the web-like strands of extracellular nuclear acids can dismantle the structure of NETs and potentially improve the efficacy of fibrinolytic therapies and severity of tissue ischemia secondary to vascular occlusion. As platelet–neutrophil interaction plays a vital role in NETs formation, novel therapies that obliterate the interactions between platelets and neutrophils may reduce thrombus growth in CATE. 20 Further investigations to provide a better understanding of the role of pathologic shear stress and NETs in thrombosis in cats with CATE will bring feline medicine a step closer to expanding the limited therapeutic options for this devastating disease.
The identification of extracellular citH3 in arterial thrombi as part of the structural component of NETs is consistent with the findings in human clinical studies and may have clinical significance in cats. Extracellular histones can further augment NET formation and thrombosis by activating and facilitating circulating platelets to interact with neutrophils.21,22 Citrullinated histones are found to be more abundant in arterial thrombi retrieved from human patients with CATE than those with other thromboembolic origins. 23 Elevated plasma citH3 is also independently associated with atrial fibrillation and mortality in humans with acute ischemic stroke. 24 Although the exact mechanism of this remains unclear, extracellular citH3 may also be elevated in cats with HCM and therefore may have diagnostic utility in predicting clot formation.
This study carries several limitations. First, two-dimensional microscopic analyses of clot sections do not allow for complete quantification of NET structures in thrombi. When blood vessels are completely occluded by thrombus, the pressure gradient across the length of the thrombus drives blood cells and plasma components through the thrombus, producing a gradient of leukocytes and NETs.3,25 Utilization of three-dimensional super-resolution microscopy or additional Z-stacking would have provided more information on the distribution of NETs in relation to the layers of thrombi. Secondly, although NETs were visualized as colocalization of citH3, cfDNA and NE, the proportion of fibrin, von Willebrand factor and neutrophils within thrombi were not assessed in our study. We therefore could not differentiate between in vivo NET formation by circulating neutrophils or artifactual NET formation due to cell lysis, autolysis following death or tissue fixation. However, histone citrullination is an active cellular process that precedes NET formation in neutrophils.26,27 Thus, the colocalization of citH3 in NET-rich thrombi indicates that in vivo NET formation is likely to have occurred in these cats. Third, although a systematic method and blinding were utilized to minimize observer bias, the method described here was semiquantitative, and that direct visualization of a clot might result in biases during the quantification process. Further research is required to standardize methods to assess NET formation in veterinary species. Lastly, the high variability in NETs distributions requires a follow-up study with a larger number of affected cats.
Conclusions
The observed presence and variation in NET-related structures in arterial thrombi shed light on the pathogenesis of thrombus growth during CATE. Results of this study warrant further investigations into the contribution of shear stress induced NETs formation in cats with CATE.
Supplemental Material
Figure 1
Representative immunofluorescence microscopy image of aortic bifurcation from a cat with hypertrophic cardiomyopathy
Footnotes
Acknowledgements
The authors would like to acknowledge Dr Kevin Woolard for his expertise and use of the fluorescence microscope.
Supplementary material
The following file is available online:
Figure 1: Representative immunofluorescence microscopy image of aortic bifurcation from a cat with hypertrophic cardiomyopathy.
Conflict of interest
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The study was supported by funds from the University of California, Davis, Center for Companion Animal Health (CCAH 2018-30-F).
Ethical approval
The work described in this manuscript involved the use of non-experimental (owned or unowned) animals. Established internationally recognized high standards (‘best practice’) of veterinary clinical care for the individual patient were always followed and/or this work involved the use of cadavers. Ethical approval from a committee was therefore not specifically required for publication in JFMS. Although not required, where ethical approval was still obtained, it is stated in the manuscript.
Informed consent
Informed consent (either verbal or written) was obtained from the owner or legal custodian of all animal(s) described in this work (either experimental or non-experimental animals, including cadavers) for all procedure(s) undertaken (either prospective or retrospective studies). No animals or people are identifiable within this publication, and therefore additional informed consent for publication was not required.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.