
Abstract
Objectives
A protein called apoptosis inhibitor of macrophages (AIM) helps clear waste from the kidney tubules. Some domestic cats harbor a duplication of exon 3 in the AIM gene, producing a four-domain AIM protein that includes an additional scavenger receptor cysteine-rich subunit. This variant has been associated with increased risk of chronic kidney disease. Pilot studies suggest that homozygosity for this genetic alteration is present in approximately 20% of the cat population. The objective of the current study was to determine the prevalence of the wild-type, heterozygous and homozygous variant AIM exon 3 genotypes in a large population of domestic cats.
Methods
Genomic DNA from 1000 client-owned cats archived in the Program in Individualized Medicine DNA Bank at Washington State University was analyzed by droplet digital PCR to determine AIM exon 3 copy number variation. Samples represented cats of any age, sex, neuter status or clinical condition. Genotype frequencies were summarized with 95% Wilson confidence intervals (CIs) and genotype frequencies were tested for deviation from Hardy–Weinberg equilibrium using χ2 analysis.
Results
Of the 1000 cats, 340 (34.0%, 95% CI 31.1–36.9) were homozygous wild-type (CNV2), 467 (46.7%, 95% CI 43.6–50.0) were heterozygous (CNV3) and 193 (19.3%, 95% CI 17.0–21.9) were homozygous for the variant (CNV4). Genotype frequencies deviate from Hardy–Weinberg equilibrium (χ2 = 4.49, degrees of freedom = 1; P = 0.034).
Conclusions and relevance
Approximately 1/5 cats were homozygous for the AIM exon 3 duplication, and nearly half harbored one copy of the variant allele. This genetic alteration is not restricted to a single breed or type. Given the potential role of this variant in feline kidney disease, the results of this study justify further investigations using genetically confirmed and geographically diverse populations to better define global allele distributions and potential breed predispositions.
Introduction
Apoptosis inhibitor of macrophages (AIM), also known as CD5-like antigen, is a protein synthesized primarily by tissue macrophages that circulates in plasma bound to immunoglobulins.1,2
The wild-type feline AIM protein comprises three scavenger receptor cysteine-rich (SRCR) domains (SRCR1–3). 2 Some domestic cats exhibit a structural variant arising from duplication of exon 3 in one or both alleles of the feline AIM gene. This duplication produces a four-domain AIM protein with an additional SRCR1 subunit and yields three possible genotypes: homozygous wild-type, heterozygous and homozygous variants. 3
The exact phenotype associated with the variant has yet to be elucidated. However, a recent study reported that cats homozygous for the exon 3 variant allele had 17× greater odds of advancing to a higher International Renal Interest Society chronic kidney disease (CKD) stage compared with wild-type cats. 3 These results imply that AIM genotypes likely carry significant clinical relevance with respect to CKD. 3
Although the variant appears common and clinically relevant, the practical utility of AIM genotyping depends on its allele frequency in the general feline population and among breeds. Preliminary data suggested that a high proportion of cats carry the four-domain variant of AIM: 37.5% were heterozygous and 22% homozygous in one pilot study. 2 Consistent results were observed in another cohort of 100 cats, where 40% were heterozygous and 20% homozygous for the variant. 3 However, these earlier studies were limited by small sample sizes. Therefore, the objective of the present study was to determine the prevalence of wild-type, heterozygous and homozygous variant AIM exon 3 genotypes in a larger population of 1000 cats.
Materials and methods
Study design
Copy-number frequencies for exon 3 of AIM (corresponding to two, three or four copies) were determined from 1000 archived DNA samples obtained from the Program in Individualized Medicine DNA Bank (IACUC protocol #6493). These samples originated from client-owned cats admitted to the Washington State University (WSU) Veterinary Teaching Hospital. Cats of any age, sex, and neuter or clinical status were eligible for inclusion. Samples were randomly selected from a repository of more than 1500 banked feline DNA specimens based on storage availability. Genotyping for AIM exon 3 copy number variation (CNV) was performed using the droplet digital PCR protocol described below. 3
Genomic DNA isolation
Genomic DNA was isolated from feline cheek-swab samples using the Quick-DNA Miniprep Kit (Zymo Research) according to the manufacturer’s protocol. DNA concentrations were quantified fluorometrically using a Qubit double-stranded DNA assay (Invitrogen) following the manufacturer’s instructions. Extracted DNA was stored at −80°C until analysis.
Owner-reported breed or type was recorded when available (n = 996/1000); no pedigree verification was performed. Cats reported as domestic shorthair, domestic mediumhair or domestic longhair were classified as random-bred. All other labels (eg, Bengal, Birman, Ragdoll) were grouped as purebred for descriptive purposes.
Determination of feline AIM exon 3 copy number
The analytical methodology followed a previously validated approach. 3
Statistical analysis
Descriptive statistics were used to summarize genotype frequencies. Genotype proportions were expressed with 95% confidence intervals (CIs) calculated using the Wilson score method,4 –6 which provides accurate coverage for binomial proportions, particularly for large or moderate sample sizes. Allele frequencies were calculated from observed genotype counts and expected genotype frequencies under Hardy–Weinberg equilibrium (HWE) using p 2 , 2pq and q2.7,8 Departure from HWE was assessed using a χ2 goodness-of-fit test with one degree of freedom.7,8 The significance level was set at P <0.05. All analyses were performed in R (v4.3) and Excel (Microsoft). Because mixing genetically distinct subpopulations can artificially produce heterozygote deficits (Wahlund effect), we assessed the influence of breed structure by identifying purebred groups that contributed disproportionately to the overall χ2 statistic. High-contributing purebred populations were defined a priori as breeds in which five or more individuals were available and genotype distributions showed an excess of homozygous genotypes (CNV2 or CNV4) relative to random-bred cats. HWE was then re-evaluated in the remaining cohort to determine whether population substructure explained the initial deviation.
Results
Of the 1000 cats analyzed, 340 (34.0%, 95% CI 31.1–36.9) were homozygous wild-type (CNV2), 467 (46.7%, 95% CI 43.6–50.0) were heterozygous (CNV3) and 193 (19.3%, 95% CI 17.0–21.9) were homozygous for the duplicated exon (CNV4). The duplicated allele had an overall frequency of 0.43. The variant was detected across 34 owner-reported breeds or types (Figure 1). Genotype frequencies in the full cohort deviated from Hardy–Weinberg equilibrium (χ2 = 4.49; P = 0.034), reflecting an excess of homozygous genotypes. Inspection of breed-stratified data showed that this deviation was driven by several purebred groups with skewed genotype distributions. Some breeds had high duplication allele frequency (Table S1 in the supplementary material). No individual breed demonstrated significant deviation from Hardy–Weinberg equilibrium when analyzed separately. After removing these high-contributing purebreds (n = 54 cats), the remaining cats (n = 946) no longer deviated from equilibrium (χ2 = 1.43; P = 0.23). In this filtered cohort (n = 946), genotype proportions were 34.6% CNV2 (95% CI 31.6–37.7), 46.8% CNV3 (95% CI 43.7–50.0) and 18.6% CNV4 (95% CI 16.3–21.2).
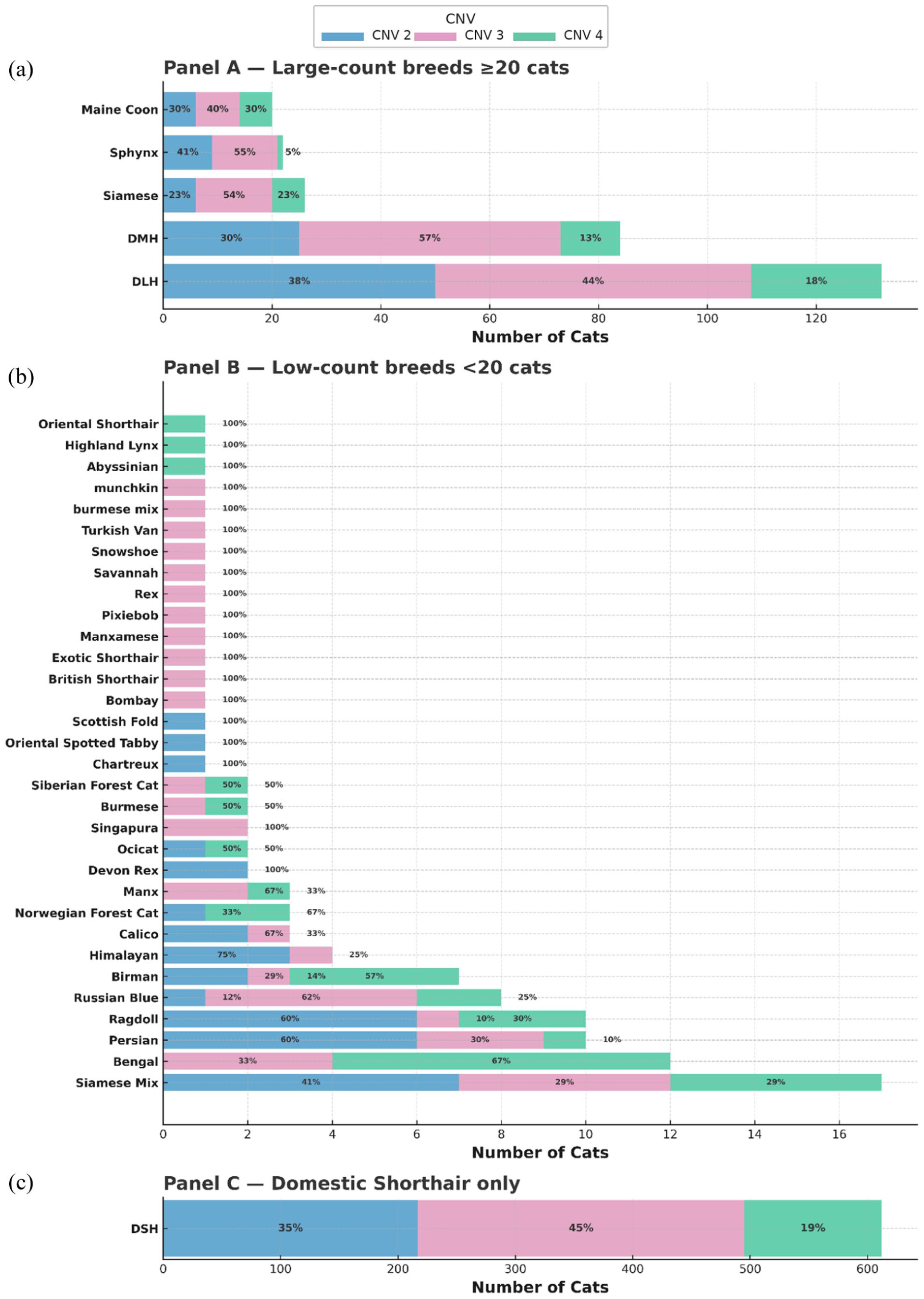
Distribution of feline apoptosis inhibitor of macrophages exon 3 genotypes (CNV2, CNV3, CNV4) across owner-reported breeds/type (n = 996). No breed/type was available for 4/1000 cat samples, and therefore these are not represented in this figure. (a) Breeds with ⩾20 cats. (b) Breeds with <20 cats. (c) Domestic shorthair (DSH) only. Percentages represent within-breed/type genotype proportions. CNV: copy number variation; DMH = domestic mediumhair; DLH = domestic longhair
Discussion
This large-scale population study revealed that approximately 1/5 domestic cats were homozygous for the AIM exon 3 duplication, a variant predicted to introduce an additional SRCR domain through duplication of the SRCR1 subunit. 3 Nearly half of all cats carried a single copy of the duplication, confirming that this allele is common in the general population and consistent with earlier pilot observations. 3 The duplication was identified across 34 owner-reported breeds/types. Although breed information was based on client reports rather than verified pedigrees, and the study was not designed to estimate breed-specific prevalence, the widespread distribution suggests that breeding practices may influence allele frequencies within certain lineages. Genotype frequencies deviated from Hardy–Weinberg equilibrium. Duplication allele frequencies varied substantially among breeds, with higher frequencies observed in Bengal and Birman cats and lower frequencies in Persian and Sphynx cats, while random-bred cats exhibited intermediate values (Table S1 in the supplementary material). Pooling these genetically structured subpopulations resulted in an apparent deficit of heterozygotes consistent with a Wahlund effect.7 –9
It is important to acknowledge that although the overall data set was large, samples came from a single teaching hospital DNA bank, the number of genetically confirmed purebred cats was limited and not all breeds were represented. Therefore, the absence of HWE deviation among the breeds should not be interpreted as evidence that those populations are in true equilibrium or that selection acting on the AIM duplication is absent. Differences in allele frequency – or even selection for or against the duplicated exon – may exist in other purebred lineages or geographic regions not captured here. Larger, breed-specific cohorts will be required to evaluate these possibilities fully.
The AIM exon 3 homozygous variant has been associated with a markedly increased risk of progressive CKD (adjusted odds ratio 9.6, 95% CI 1.38–67). 3 The high prevalence observed here underscores the potential clinical and translational relevance of this variant. No phenotype data (CKD status, creatinine, symmetric dimethylarginine or age) were available for this cohort; therefore, the present study reports only allele and genotype frequencies, not CKD risk within this population. The results from this study, in combination with prior CKD risk estimates, highlight the need for further work to determine how AIM genotyping might inform breeding strategies and clinical monitoring. Recent evidence supports the potential of AIM-based therapies for advanced CKD in cats. 10 The effect of these therapies on disease progression in cats harboring the AIM variant remains unknown and warrants further investigation, as it may represent a significant advance in precision therapeutics for feline CKD.
This study has several important limitations. First, the cohort represents a hospital-based, convenience sample from a single veterinary teaching institution and therefore may not reflect allele frequencies in the general domestic cat population. In addition, no clinical phenotype data (CKD status, renal biomarkers, age, comorbidities) were available for this cohort, and therefore no genotype–phenotype associations can be inferred from these results; all disease associations are based on previously published work.
Conclusions
This study establishes the AIM exon 3 duplication as a common genetic variant in domestic cats. Future investigations involving genetically confirmed purebred populations and broader geographic sampling will be essential to characterize global allele distributions and potential breed-specific predispositions.
Supplemental Material
Table S1
Complete, transparent list of duplication allele frequency (q) and sample size (N) for every breed or owner reported type included in the dataset.
Footnotes
Acknowledgements
Figure 1 was created with ChatGPT.
Supplementary material
The following file is available as supplementary material:
Table S1: Complete, transparent list of duplication allele frequency (q) and sample size (N) for every breed or owner reported type included in the dataset.
Conflict of interest
Dr Villarino may establish a service center at WSU that offers genetic testing.
Funding
Dr Villarino’s kidney research program is supported by the Washington State University College of Veterinary Medicine Kay Yarborough Nelson Distinguished Professor in Feline Health Endowment, a generous gift from Mary Kay Fowler, Washington State University intramural grants program and Washington Research Foundation.
Ethical approval
The work described in this manuscript involved the use of non-experimental (owned or unowned) animals and procedures that differed from established internationally recognized high standards (‘best practice’) of veterinary clinical care for the individual patient. The study therefore had prior ethical approval from an established (or ad hoc) committee as stated in the manuscript.
Informed consent
Informed consent (verbal or written) was obtained from the owner or legal custodian of all animal(s) described in this work (experimental or non-experimental animals, including cadavers, tissues and samples) for all procedure(s) undertaken (prospective or retrospective studies). No animals or people are identifiable within this publication, and therefore additional informed consent for publication was not required.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.