
Abstract
Background:
Hypertriglyceridemia is a risk factor for cardiovascular disease and several pregnancy complications. Lipoprotein lipase (LPL) genetic variation modulates nonpregnancy plasma triglycerides, but its effects during pregnancy are unknown. The G allele of the LPL -93T/G promoter polymorphism is 16–23 times more prevalent in Blacks than in Whites, contributing to lower triglycerides in nonpregnant African Americans by increasing LPL expression.
Purpose:
This study investigated whether the triglyceride-lowering effect of -93G is observed in African Americans during pregnancy.
Methods:
Genotyping was performed on 124 African American women with uncomplicated pregnancies for common functional LPL polymorphisms/mutations (-93T/G, D9N, N291S, and S447X). Third-trimester plasma triglyceride, high- and low-density lipoprotein cholesterol, apolipoprotein B, and free fatty acid concentrations were measured with colorimetric assays. Clinical characteristics and lipid values were compared across the -93T/G genotypes.
Results:
Triglycerides were significantly lower in women with the -93GG compared to the -93TT genotype, both with (n = 124, p = .02) and without (n = 108, p = .03) inclusion of participants with other LPL variant alleles. Triglyceride differences persisted after adjustment for prepregnancy body mass index, gestational age at delivery, and smoking. There were no significant differences in the other lipids or apolipoprotein B by -93T/G genotype.
Conclusions:
Despite the considerable metabolic changes accompanying pregnancy, the triglyceride-lowering effect associated with the -93GG LPL genotype in African Americans persists during late pregnancy. The -93GG genotype might protect against pregnancy complications stemming from hypertriglyceridemia, but the overall increased risk of pregnancy complications in African American women points to complex, multifactorial relationships among risk factors, race, and adverse pregnancy outcomes.
Maternal lipid metabolism undergoes a striking transformation during pregnancy, resulting in elevations in plasma triglycerides, cholesterol, and nonesterified fatty acids, with reversal postpartum (Herrera, 2002). Several factors related to estrogen and insulin resistance account for the 50–300% rise in maternal triglycerides over prepregnancy levels during the second half of pregnancy; these factors include enhanced production of very low-density lipoprotein (VLDL) cholesterol by the liver and reduced lipoprotein lipase (LPL) activity (Herrera, 2002; Knopp, Bonet, Lasuncion, Montelongo, & Herrera, 1992). Maternal hypertriglyceridemia plays a key role in the availability of essential fatty acids to the placenta and fetus (Herrera, 2002), and there is a direct relationship between maternal triglycerides and newborn weight in humans (Knopp, Magee, Walden, Bonet, & Benedetti, 1992). On the other hand, increased triglyceride-rich lipoprotein levels are independently associated with cardiovascular disease (Hokanson & Austin, 1996). Evidence also suggests that excessive gestational hypertriglyceridemia contributes to the development of preeclampsia, a cardiovascular disease of pregnancy, and other pregnancy complications (Hubel, Lyall, Weissfeld, Gandley, & Roberts, 1998; Mudd, Holzman, Catov, Senagore, & Evans, 2012; Vrijkotte et al., 2012). Lipid disturbances during pregnancy might also lead to early onset of metabolic diseases in the offspring (Napoli et al., 1999).
Healthy Black women of reproductive age have lower fasting plasma triglyceride concentrations, on average, compared to similarly aged healthy White women (Donahue et al., 1989; Proudler, Godsland, Bruce, Seed, & Wynn, 1996). This profile might contribute to the reduced prevalence of, but not to the increased mortality of, coronary heart disease in Black, compared to White, women (Bransford & Ofili, 2000). Researchers have also observed lower LDL cholesterol levels in African American women as compared with White women, although less consistently than they have noted lower triglyceride levels (Donahue et al., 1989; Metcalf et al., 1998; Morrison et al., 1981; Proudler et al., 1996; Tyroler, Glueck, Christensen, & Kwiterovich, 1980). Few studies have examined ethnic differences in lipid concentrations during pregnancy. However, available data indicate that both fasting and nonfasting triglyceride and cholesterol levels are lower, on average, in women of African descent compared to Caucasian women during early and late gestation (Koukkou, Watts, Mazurkiewicz, & Lowy, 1994; Patrick, Hubel, & Roberts, 2004; Schreuder et al., 2011).
Abnormalities in lipid metabolism have, at least in part, a genetic basis. LPL is the rate-limiting enzyme in the clearance of triglyceride-rich lipoproteins from the circulation. LPL breaks down triglycerides into free (nonesterified) fatty acids and glycerol, which are subsequently taken up by the surrounding tissue to meet local energy demands (Ehrenborg et al., 1997; Hall et al., 1997; Herrera, 2002). A genetic variation within the LPL gene promoter (-93T/G) (rs1800590, dbSNP Human Build 142) increases LPL promoter function (Hall et al., 1997) and is associated with lower circulating triglyceride levels and enhanced postprandial triglyceride clearance in African Americans (Talmud et al., 1998). This triglyceride-lowering effect is not seen in Caucasians due to the strong linkage disequilibrium between -93T/G and the D9N polymorphism (exon 2 of the LPL gene; Hall et al., 1997). The D9N variant (N9) produces a secretion-defective LPL protein (Hall et al., 1997; Talmud et al., 1998), and coinheritance of the -93G and N9 variants thus results in an amplification of the secretion-defective LPL protein (Hall et al., 1997). In contrast, the frequency of the -93G allele is 16- to 23-fold higher in Blacks compared to Whites (Hall et al., 1997; Hall et al., 2000), and its allelic association with the N9 variant is greatly reduced (Ehrenborg et al., 1997; Hall et al., 1997; Talmud et al., 1998).
The influence of the -93G promoter variant on circulating lipids in African American women during pregnancy has not been examined and cannot be assumed from nonpregnancy data, given the major lipid changes that accompany pregnancy. The high frequency of the -93G variant in the absence of the N9 variant in African Americans allowed us, in this study, to directly examine the phenotypic effects of this promoter variant on lipid profile. We hypothesized that the influence of -93T/G would persist during late pregnancy, contributing to lower circulating triglycerides among African Americans with the -93G allele.
Method
Study Population
The Prenatal Exposures and Preeclampsia Prevention (PEPP) Study was a prospective study of women enrolled at the time of presentation to the labor and delivery suite or at ≤20 weeks’ gestation. The University of Pittsburgh Institutional Review Board approved the study, and all participants provided written informed consent. A total of 242 African American women with uncomplicated pregnancy outcomes who provided blood samples during the period 1995–2004 were genotyped for four common LPL polymorphisms/mutations [-93T/G (rs1800590), D9N (rs1801177), N291S (rs268), and S447X (rs328)] for the present and related PEPP studies. Sufficient third-trimester plasma samples for lipid analyses were available from 124 of the 242 women who had been genotyped. We specifically aimed to study the effects of -93T/G. Therefore, subjects with the D9N variant (n = 3 N9 heterozygotes), which reduces LPL activity (Hall et al., 1997), and S447X variant (n = 13 X447 heterozygotes), which is associated with lower plasma triglycerides (i.e., increase in LPL activity; Talmud et al., 1998), were identified and excluded from the primary analysis. The N291S variant (S291) also reduces LPL activity (Hall et al., 1997); however, there were no S291 heterozygotes or homozygotes in our sample of 124 women. Thus, the primary study group for comparison between third-trimester lipid and apolipoprotein B values according to -93T/G genotype consisted of 108 individuals with only wild-type genotypes for the D9N, N291S, and S447X mutations. We conducted secondary analyses that included all 124 subjects with available blood samples.
We classified study subjects as having an uncomplicated pregnancy if they were normotensive and without proteinuria throughout gestation and if they delivered healthy babies. All women had singleton pregnancies; did not use illicit drugs; and had no clinical history of diabetes or renal, hypertensive, or vascular disease; and 95% were nulliparous.
We used self-reported prepregnancy weight and measured height to calculate prepregnancy body mass index (BMI; weight [kg]/(height [m])2). Gestational age–specific birth weight percentiles, adjusted for infant sex and race, were based upon data from Magee-Womens Hospital (Pittsburgh, PA). We collected maternal race and tobacco smoking history (y/n, ≥ 100 cigarettes in lifetime) data via self-report.
Lipid Measurements
Maternal nonfasting (random with respect to last meal) blood samples were collected during the third trimester (n = 124; 34 1/7th weeks to 42 0/7th weeks’ gestation) and centrifuged at 2000 × g for 20 min; the serum and plasma were stored at −80 °C. The gestational age at the time of sampling was determined from the best obstetrical estimate of gestational age (early ultrasound, when available, or last menstrual period consistent with second-trimester ultrasound). Triglyceride, total cholesterol, high-density lipoprotein (HDL) cholesterol, apolipoprotein B, and total free fatty acid concentrations were measured in serum (92.6% of samples) or plasma (7.4%) samples. Triglyceride and total cholesterol were measured using Pointe microplate enzymatic assays (Pointe Scientific, Canton, MI). Free fatty acid concentrations were measured with the enzymatic Wako NEFA C microtiter assay (Wako Chemicals, Richmond, VA). Apolipoprotein B concentrations were determined by immunoturbidimetric assay (Pointe Scientific). HDL cholesterol concentration was determined after selective precipitation by heparin/manganese chloride and removal, by centrifugation, of VLDL and LDL particles (Warnick & Albers, 1978). The cholesterol in the HDL particles was measured as described previously for total cholesterol. The average coefficients of variation between runs for these assays in our laboratories ranged from 3% to 9%. LDL cholesterol was estimated using the Friedewald equation; no triglyceride concentrations were in the range invalid for the equation (>400 mg/dl; Friedewald, Levy, & Fredrickson, 1972).
Genotype Data Collection
DNA was extracted from white blood cell pellets using the salting-out method (Powers et al., 1999). For those samples with no available white blood cell pellet (n = 5), DNA was extracted from serum (Powers et al., 1999). Next, the region flanking the -93T/G locus was amplified via polymerase chain reaction, as described previously (Hubel, Roberts, & Ferrell, 1999). Amplification of the D9N (Exon 2), the N291S (Exon 6), and the S447X (Exon 9) loci has also been described previously (Faustinella et al., 1991; Monsalve et al., 1990). Following loci amplification, -93T/G, D9N, N291S, and S447X genotypes were determined with the oligonucleotide ligation assay (Hubel et al., 1999; Nickerson et al., 1990).
Statistical Analysis
Statistical analyses were conducted with SPSS Statistics, version 21.0 (IBM, Armonk, NY). Because of nonnormal distributions, nonparametric tests (Kruskal–Wallis; Mann–Whitney U) were used to compare clinical variables and lipid and apolipoprotein B concentrations across the three -93T/G genotype groups. Chi-square or Fisher’s exact tests were used for categorical variables. Spearman’s ρ correlations were computed to investigate the bivariate relationships between triglyceride, other lipid and apolipoprotein B levels, and demographic variables. The primary outcome that varied according to -93T/G genotype, triglycerides, was also examined using multiple linear regression models to allow adjustment for potential confounders, including prepregnancy BMI, gestational age at delivery, and cigarette-smoking history. Finally, to investigate whether -93G carriers (GG and GT) were “protected” from the triglyceride-raising effects of high BMI, we conducted a subgroup analysis in which we compared triglyceride concentrations in -93G carriers with BMI above versus below the study median.
Results
Frequency of LPL Variants
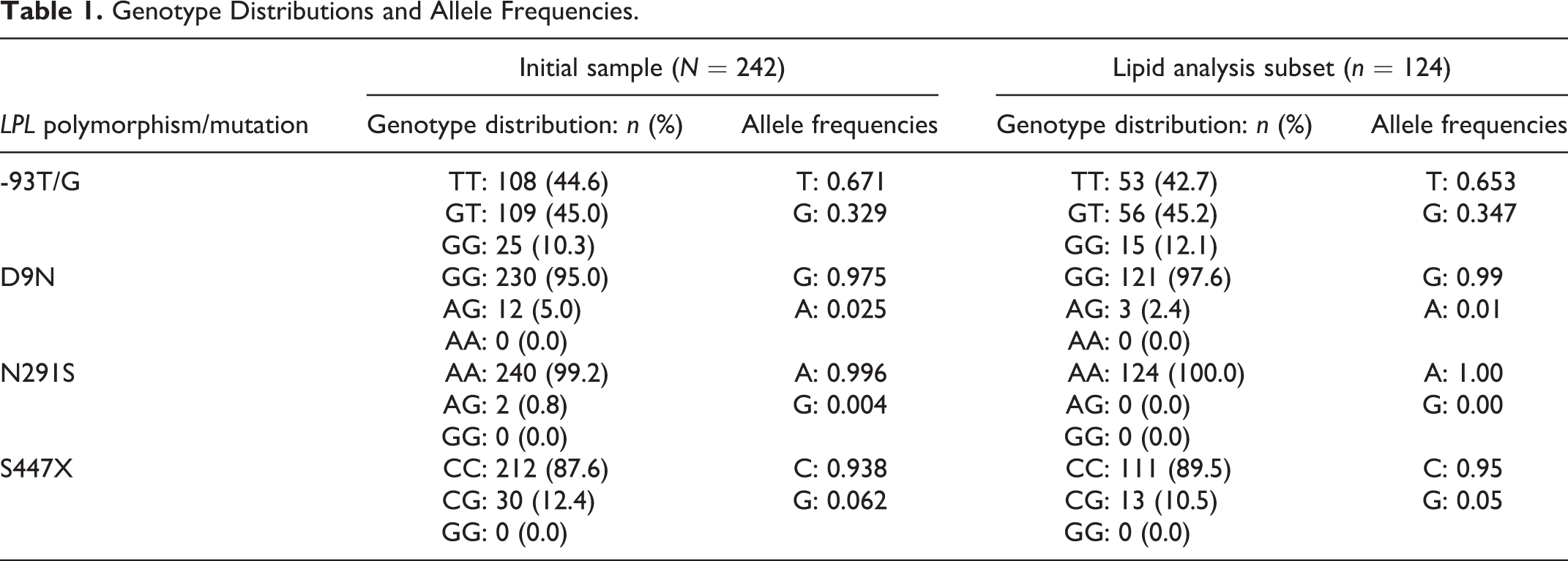
Genotype distributions and allele frequencies of -93T/G, D9N, N291S, and S447X are presented in Table 1. The frequency of the LPL -93T/G promoter variant allele (G) in the 242 healthy pregnant African American women was 33%. Genotype and allele frequencies observed in our lipid analysis subset (n = 124) were similar to values reported for our overall population. The -93T/G polymorphism was in Hardy–Weinberg equilibrium (HWE; N = 242; p > .05). We did not assess HWE for D9N, N291S, or S44X, given their extremely skewed distributions.
Genotype Distributions and Allele Frequencies.
Clinical Characteristics by Genotype
Table 2 presents clinical characteristics, by -93T/G genotype groups, of the 108 women lacking the D9N, N291S, and S447X variants. There were no significant differences in maternal age at delivery, blood pressure measurements before 20 weeks’ gestation or during labor, smoking history, or birth weight centile among the three -93T/G genotype groups. Median gestational age at delivery was slightly, but significantly, different across the three genotype groups, with the TT group delivering about 1 week earlier than the GG group. Median prepregnancy BMI was marginally different among the three -93T/G genotype groups, with median BMI of the GG genotype group higher than that of the TT and GT genotype groups. The proportion of obese subjects (BMI > 30 kg/m2) was significantly higher in the GG genotype group (GG = 53.8%, GT = 22.0%, and TT = 20.0%); comparison across the three groups: χ2(2) = 6.6, p = .037. These relationships remained significant with the inclusion of the 16 women with the D9N (n = 3 N9 heterozygotes) or S447X (n = 13 X447 heterozygotes) variants, except that differences in median prepregnancy BMI and gestational age at delivery became nonsignificant (data not shown).
Demographic and Clinical Characteristics of Subjects With Sufficient Third-Trimester Blood Samples and With Wild-Type D9N, N291S, and S447X Genetic Backgrounds, Stratified by 93T/G Genotype.
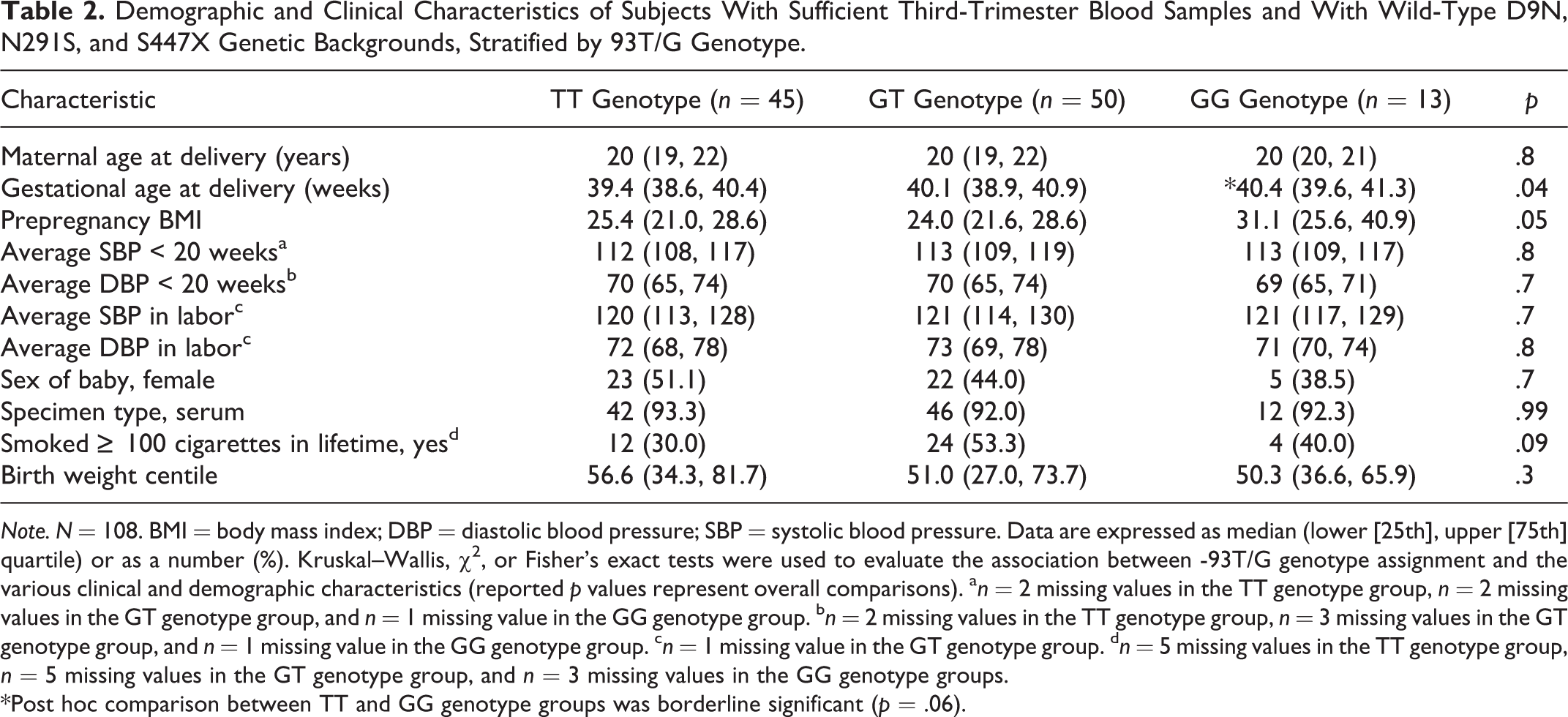
Note. N = 108. BMI = body mass index; DBP = diastolic blood pressure; SBP = systolic blood pressure. Data are expressed as median (lower [25th], upper [75th] quartile) or as a number (%). Kruskal–Wallis, χ2, or Fisher’s exact tests were used to evaluate the association between -93T/G genotype assignment and the various clinical and demographic characteristics (reported p values represent overall comparisons). a n = 2 missing values in the TT genotype group, n = 2 missing values in the GT genotype group, and n = 1 missing value in the GG genotype group. b n = 2 missing values in the TT genotype group, n = 3 missing values in the GT genotype group, and n = 1 missing value in the GG genotype group. c n = 1 missing value in the GT genotype group. d n = 5 missing values in the TT genotype group, n = 5 missing values in the GT genotype group, and n = 3 missing values in the GG genotype groups.
*Post hoc comparison between TT and GG genotype groups was borderline significant (p = .06).
Lipid Levels by -93T/G Genotype
The gestational age at the time of blood sampling for lipid measurements was similar among the three -93G/T genotype groups comprising the 108 women wild-type for D9N, N291S, and S447X (Table 3). Third-trimester triglyceride concentrations were significantly different among the three groups (p = .03), but total cholesterol, LDL cholesterol, HDL cholesterol, apolipoprotein B, and free fatty acids were not. The median triglyceride concentration among women with the GG genotype was significantly lower (by 42 mg/dl) compared to the more common TT genotype group (post hoc p = .026). Median triglyceride concentrations were not significantly different in other pairwise comparisons (GG vs. GT [p = .26]; TT vs. GT [p = .46]). Lipid profiles were not significantly altered by inclusion of subjects who had the D9N or S447X variants (data not shown).
Third-Trimester Lipid and Apolipoprotein B Values, by LPL -93T/G Genotype, in Subjects With Wild-Type D9N, N291S, and S447X Genetic Backgrounds.
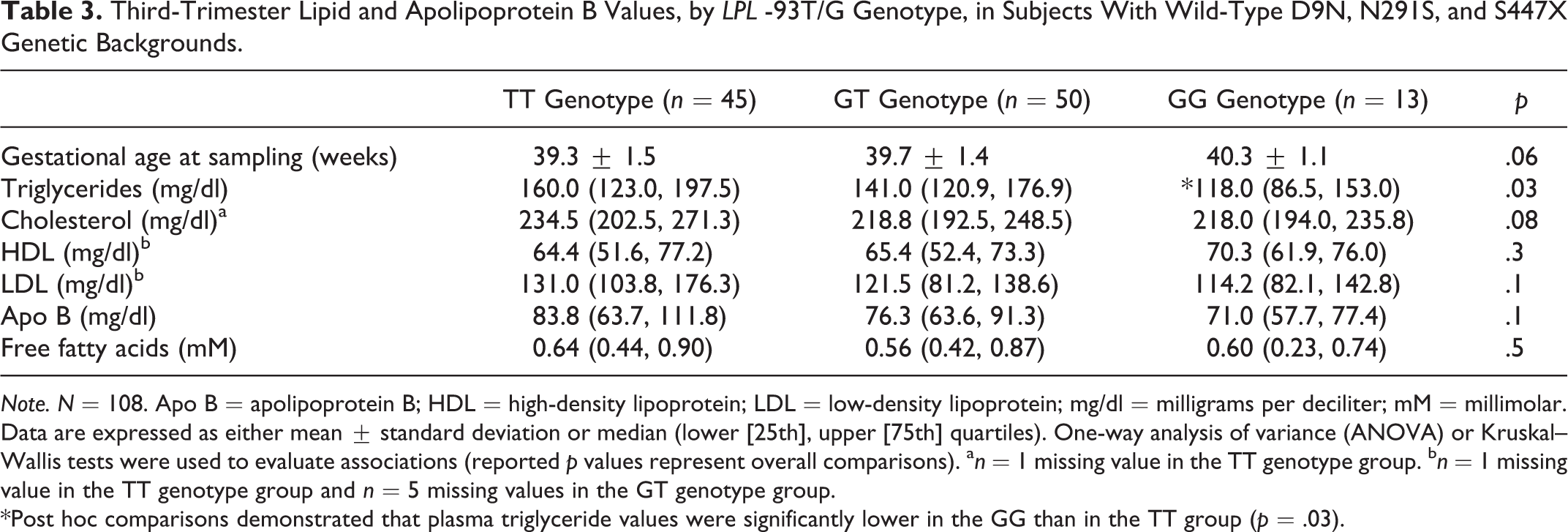
Note. N = 108. Apo B = apolipoprotein B; HDL = high-density lipoprotein; LDL = low-density lipoprotein; mg/dl = milligrams per deciliter; mM = millimolar. Data are expressed as either mean ± standard deviation or median (lower [25th], upper [75th] quartiles). One-way analysis of variance (ANOVA) or Kruskal–Wallis tests were used to evaluate associations (reported p values represent overall comparisons). a n = 1 missing value in the TT genotype group. b n = 1 missing value in the TT genotype group and n = 5 missing values in the GT genotype group.
*Post hoc comparisons demonstrated that plasma triglyceride values were significantly lower in the GG than in the TT group (p = .03).
We used multiple linear regression models to examine the association between -93T/G genotype and triglycerides while adjusting for potential confounders (Table 4). There was a significant difference in triglyceride concentration between the GG genotype and the more common TT genotype in all models, adjusting for prepregnancy BMI, gestational age at delivery, and smoking history.
Effects of -93T/G Genotype on Triglyceride Concentrations, Adjusted for Potential Confounders, in Subjects With Wild-Type D9N, N291S, and S447X Genetic Backgrounds.
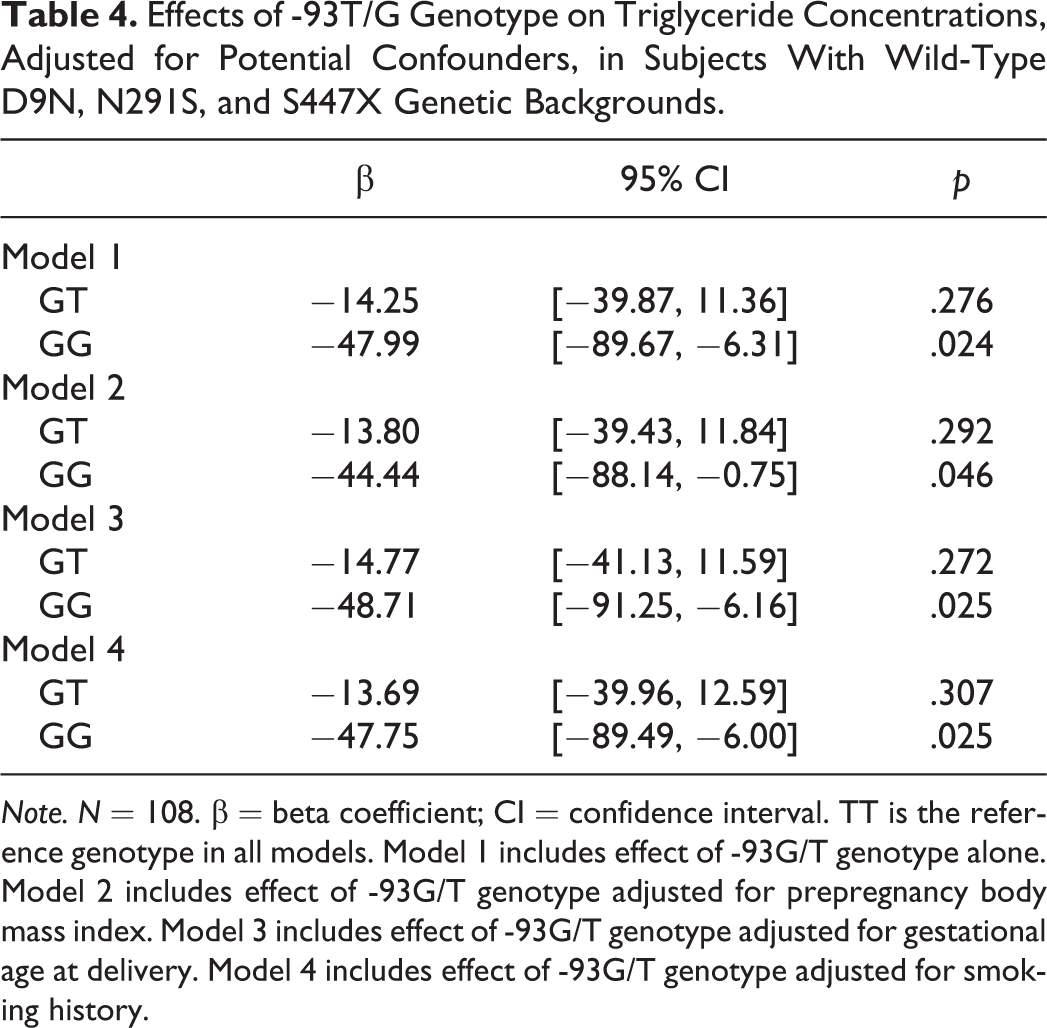
Note. N = 108. β = beta coefficient; CI = confidence interval. TT is the reference genotype in all models. Model 1 includes effect of -93G/T genotype alone. Model 2 includes effect of -93G/T genotype adjusted for prepregnancy body mass index. Model 3 includes effect of -93G/T genotype adjusted for gestational age at delivery. Model 4 includes effect of -93G/T genotype adjusted for smoking history.
Bivariate Correlations
Among the 108 subjects who were wild type for D9N, N291S, and S447X, birth weight centile was positively correlated with triglycerides (rs = 0.2, p = .02). However, we found no associations between triglycerides and prepregnancy BMI, gestational age at sampling, gestational age at delivery, maternal age, blood pressure (p’s > .05), or smoking (U = 1,004.5, p = .5). Triglycerides were positively correlated with apolipoprotein B (rs = .59, p < .001), total cholesterol (rs = .30, p = .001), LDL cholesterol (rs = .22, p = .03), and free fatty acids (rs = .24, p = .01) and negatively correlated with HDL cholesterol (rs = −.31, p = .001). Inclusion of subjects with the D9N and S447X variants did not alter these findings (data not shown).
-93G Carriers (-93GG and -93GT) May Be Protected From the Triglyceride-Raising Effects of High BMI
When restricting our analysis to -93T carriers (-93TT and -93GT; n = 95), we found that women whose BMIs were above the median had significantly higher triglyceride levels (153 mg/dl vs. 141 mg/dl, respectively; p = .04) than those whose BMIs were below the median. In contrast, when we restricted our analysis to -93G carriers (-93GG and -93GT; n = 63), we found that women whose BMIs were above the median did not have significantly higher triglyceride levels (139 mg/dl vs. 135 mg/dl, respectively; p = .49) than those whose BMIs were below the median.
Discussion
In a cohort of 108 African American women with uncomplicated pregnancy outcomes, we found a significant association between -93T/G genotype and third-trimester triglyceride concentrations. Specifically, we found lower triglyceride levels in subjects with the G allele, especially the GG genotype. This relationship was not evident for other lipids (HDL cholesterol, LDL cholesterol, apolipoprotein B, or free fatty acids). A limitation of our analysis was the small sample size, particularly the low number of GGs (n = 13), which potentially limited our statistical power to detect differences in lipids. However, a post hoc power calculation showed that we had 99% power to detect a between-group difference in triglycerides of 20 mg/dl, which is what we observed in our study.
During the second half of normal pregnancy, a state of moderate insulin resistance develops to augment maternal plasma glucose, lipid, and amino acid concentrations for transport across the placenta (Herrera, 2002). Both increased production and decreased hepatic clearance of lipoproteins contribute to gestational increases in postprandial and fasting triglycerides (Alvarez, Montelongo, Iglesias, Lasuncion, & Herrera, 1996). By term, plasma triglycerides increase by 50–300% over nonpregnancy levels, at which time higher triglycerides are found, not only in VLDL, but also in intermediate-density lipoprotein, LDL, and HDL (Hubel et al., 1999; Knopp, Bonet, et al., 1992; Montelongo, Lasuncion, Pallardo, & Herrera, 1992).
Lipid aberrations accompany several pregnancy complications. Hypertriglyceridemia and increased levels of free fatty acids are characteristic of preeclampsia, preceeding its clinical presentation, which may suggest a causal role (Lorentzen, Endresen, Clausen, & Henriksen, 1994). Women with elevated triglycerides (Lorentzen et al., 1994; Vrijkotte et al., 2012) or nonesterified fatty acids (Lorentzen et al., 1994) during the first or second trimester, or with elevated triglycerides years before pregnancy (Magnussen et al., 2007), are at heightened risk of preeclampsia. These findings suggest a predisposing phenotype. A similar dyslipidemic profile may be associated with spontaneous and medically indicated preterm births (Rich-Edwards, Fraser, Lawlor, & Catov, 2013).
On the other hand, a direct relationship between maternal triglyceride concentrations and birth weight has been observed in humans, and severe correction of the physiologic hypertriglyceridemia of pregnancy has negative effects on fetal growth (Herrera, 2002). Although maternal triglyceride-rich lipoproteins do not directly cross the placenta, the fatty acids derived from these lipoproteins provide critical substrate for fetal growth and development via the action of LPL. A higher concentration of triglycerides thus promotes fetal growth. Although the importance of maternal cholesterol for the fetus lessens by late pregnancy, low maternal total cholesterol is associated with fetal growth restriction (Sattar et al., 1999). Such data suggest that both extremely high maternal lipid concentrations (such as hypertriglyceridemia in preeclampsia) and low maternal lipid concentrations (as with low cholesterol and growth restriction) are associated with adverse pregnancy outcomes.
Limited information exists regarding ethnic differences in lipid levels during pregnancy (McGladdery & Frohlich, 2001). Pregnant women of African or Afro-Caribbean origin generally display lower triglyceride, LDL cholesterol, and apolipoprotein B levels and higher HDL cholesterol levels compared with Caucasian women, suggesting that racial differences in circulating lipids persist during pregnancy (Koukkou et al., 1994; Patrick et al., 2004; Schreuder et al., 2011). In addition to environment and lifestyle factors, genetic factors may contribute to the lipid differences observed by race.
Several relatively common genetic variations have been associated with moderate changes in LPL catalytic function. The S447X variant allele X447 has been reported to have an approximate frequency of 8% in Whites and 6% in Blacks (Hall et al., 2000). X447 confers an increased receptor-binding affinity important for LPL-mediated uptake of lipoproteins, offering a likely explanation of the modestly advantageous (lower triglyceride and raised HDL cholesterol levels) lipid profile of carriers (Sagoo et al., 2008). Two variants within the coding region, an aspartic acid-to-asparagine change at amino acid 9 in Exon 2 (D9N) and an asparagine-to-serine change at amino acid 291 in Exon 6 (N291S), are associated with raised plasma triglycerides and risk of coronary artery disease (Fisher, Humphries, & Talmud, 1997). Meta-analysis suggests an effect of D9N or N291S LPL genetic variation to increase the risk of preeclampsia in Caucasian populations (Buurma et al., 2013), although not all studies agree.
The nucleotide transversion of T to G at position -93 from the LPL transcription start site confers an increased expression activity of the LPL promoter (Hall et al., 1997). However, Hall et al. (1997) demonstrated that 83% of a group of Caucasian men with ≥1 copy of the -93T/G promoter variant (GT or GG) were also carriers of ≥1 copy of the D9N variant (DN or NN). This coinheritance of the -93T/G and D9N variants is associated with hypertriglyceridemia due to promoter-mediated increases in expression of the defective N9 enzyme (Hall et al., 1997; Wittrup et al., 1999). In contrast, Hall et al. (1997) also reported that only 17.5% of a group of Afro-Caribbean subjects with ≥1 copy of the -93T/G promoter variant were also carriers of ≥1 copy of the D9N variant. Other studies have identified this relative lack of allelic association in Blacks (Talmud et al., 1998; Wittrup et al., 1999), and this study further supports it (9.0% of African American pregnant women with ≥1 copy of the -93T/G promoter variant were also carriers of one copy of the D9N variant—there were no women who were homozygous for N).
The net result of the low allelic association is that, compared to African Americans with no variants (TT/DD), those with ≥1 copy of the -93T/G variant on a wild-type D9N background (TG/DD and GG/DD) manifest significantly lower triglyceride levels and increased postprandial triglyceride clearance rates (Talmud et al., 1998). The frequency of the LPL -93G promoter variant allele in our sample was 33%, a value intermediate between the variant allele frequencies of 28% reported in a sample of healthy African American men and women (Talmud et al., 1998) and 40.7% reported for Blacks of African origin residing in South London (Hall et al., 2000). This frequency is 16- to 23-fold higher than that observed in White populations (Hall et al., 1997, 2000). Limiting our sample to African American women, therefore, gave us the opportunity to analyze the association of pregnancy lipids with the -93G transition, independent of the D9N variant (N9).
In this study, African American women with the LPL promoter -93GG genotype had significantly lower third-trimester triglyceride concentrations compared to those with the more common TT genotype. This was the case whether or not subjects with the D9N or S447X variants were included in the analysis and despite the fact that triglyceride levels increase with the gestational age of the fetus; the average gestational age at blood sampling was about 1 week later for GG compared to TT individuals. Triglyceride levels remained significantly different in all models, adjusting for prepregnancy BMI, gestational age at delivery, and smoking history. Birth weight centile was positively correlated with triglycerides among the 108 study subjects who were wild type for D9N and S447X.
Consistent with a study by Radha, Vimaleswaran, Ayyappa, and Mohan (2007), the -93GG genotype was associated with higher (prepregnancy) BMI in this study. High prepregnancy BMI, but not gestational weight gain, has been associated with increased maternal triglyceride concentrations across pregnancy (Scifres, Catov, & Simhan, 2014). As expected, the subgroup of -93T carriers (TT and GT) with a prepregnancy BMI above the sample median had significantly higher median third-trimester triglyceride concentrations compared to -93T carriers with BMI below the sample median. In contrast, the triglyceride-raising effect of high prepregnancy BMI appeared to be absent in carriers of -93G (GG and GT). As such, the -93G allele appears to be protective against the gestational triglyceride-raising effects of high BMI. This effect has been noted in nonpregnant subjects (Hall et al., 1997), but we are the first to report it during pregnancy. Given its association with higher BMI yet lower triglycerides, it is biologically plausible that the -93G allele contributes to a “metabolically healthy obese” (Ortega et al., 2013) phenotype.
Our pregnancy plasma and serum samples were predominantly nonfasting. However, differences in the time of last meal relative to venipuncture are unlikely to occur systematically according to genotype in women with uncomplicated pregnancies. Further, measurement of nonfasting triglyceride levels may be more biologically meaningful in Western societies than fasting levels, given that waking hours are predominantly spent in a postprandial state (Miller et al., 2011). Nonfasting levels of fasting-sensitive lipids (especially triglycerides and free fatty acids) may predict future cardiovascular events similarly to, or even better than, fasting levels, as they are more reflective of overall interactions of lipoproteins with the vascular endothelium (Ridker, 2008). Finally, the physiological impact of LPL variants may be most evident postprandially, when the ability of LPL to clear dietary triglycerides is most pronounced (Talmud et al., 1998).
Both increased prepregnancy adiposity, even below the obese range (Hubel & Roberts, 2009), and gestational hypertriglyceridemia (Lorentzen et al., 1994; Mudd et al., 2012) are risk factors for development of preeclampsia or preterm birth. Afro-Caribbean racial origin has been associated with increased risk of preeclampsia and spontaneous preterm birth (Knuist, Bonsel, Zondervan, & Treffers, 1998; Reagan & Salsberry, 2005). In one study, however, Black women did not have significantly greater prevalence of placentation-related pregnancy complications than White women, but their mortality rates were 2–3 times higher (Tucker, Berg, Callaghan, & Hsia, 2007). These increased risk or mortality rates evidently exist despite the reduced propensity for Black women to experience excessive gestational hypertriglyceridemia.
Specific mechanisms contributing to differences in gestational lipid profiles between Black and White women have not been described previously. By lowering gestational circulating triglyceride concentrations, especially in Black women, the -93G allele may influence relationships among adiposity, fetal birth weight, and pregnancy outcome in ways that remain to be further explored. Our findings also highlight the value of considering race separately when evaluating adaptive changes in pregnancy, such as lipid metabolism, that can impact pregnancy outcome.
Footnotes
Acknowledgments
The authors thank the staff of the Preeclampsia Program Project (PEPP) core for pregnancy participant recruitment and blood sample acquisition, Beth A. Hauth of the Heinz Nutrition Laboratory at the University of Pittsburgh Graduate School of Public Health (Rhobert W. Evans, Director) for performing the HDL cholesterol measurements, and Susan DeCroo, Department of Human Genetics, for performing the genotyping.
Author Contributions
MJS contributed to analysis and interpretation, drafted manuscript, critically revised the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy; REF contributed to acquisition and analysis, critically revised the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy; MJG contributed to acquisition, critically revised the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy; DLL contributed to acquisition, critically revised the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy; ADA contributed to analysis and interpretation, critically revised the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy; JMR contributed to interpretation, critically revised the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy; CAH contributed to conception and design, contributed to analysis and interpretation, drafted the manuscript, critically revised the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was supported by National Institutes of Health Grants P01HD030367 (CAH) and UL1RR024153 and UL1TR000005 (University of Pittsburgh Clinical and Translational Science Institute). MJS was supported by the National Institute of Nursing Research T32NR009759-06 and 1F32NR014622-01.