
Abstract
Cobalamin (vitamin B12) deficiency is a subtle progressive clinical disorder, affecting nearly 1 in 5 individuals > 60 years old. This deficiency is produced by age-related decreases in nutrient absorption, medications that interfere with vitamin B12 absorption, and other comorbidities. Clinical heterogeneity confounds symptom detection for elderly adults, as deficiency sequelae range from mild fatigue and weakness to debilitating megaloblastic anemia and permanent neuropathic injury. A better understanding of genetic factors that contribute to cobalamin deficiency in the elderly would allow for targeted nursing care and preventive interventions. We tested for associations of common variants in genes involved in cobalamin transport and homeostasis with metabolic indicators of cobalamin deficiency (homocysteine and methylmalonic acid) as well as hematologic, neurologic, and functional performance features of cobalamin deficiency in 789 participants of the Women’s Health and Aging Studies. Although not significant when corrected for multiple testing, eight single nucleotide polymorphisms (SNPs) in two genes, transcobalamin II (TCN2) and the transcobalamin II-receptor (TCblR), were found to influence several clinical traits of cobalamin deficiency. The three most significant findings were the identified associations involving missense coding SNPs, namely, TCblR G220R (rs2336573) with serum cobalamin, TCN2 S348F (rs9621049) with homocysteine, and TCN2 P259R (rs1801198) with red blood cell mean corpuscular volume. These SNPs may modify the phenotype in older adults who are more likely to develop symptoms of vitamin B12 malabsorption.
Keywords
To maintain health, humans must have an adequate intake of cobalamin (vitamin B12) and retain the ability to absorb and assimilate the vitamin. Absorption of cobalamin is dependent upon the coordinated actions of binding proteins that attach to ingested dietary protein in the stomach and intestine and a family of receptors that facilitate entry into the plasma and tissues (Herrmann, Obeid, Schorr, & Geisel, 2003; Rosenblatt & Fenton, 2001). After an ileal enterocyte absorbs dietary cobalamin bound to intrinsic factor from the intestinal lumen, it is paired with the transport protein transcobalamin II (TCN2), released into the portal circulation, and carried to cells in target tissues. Under normal physiologic conditions, TCN2 is 20–30% saturated and represents 10–20% of the total cobalamin pool in plasma (Refsum, Johnston, Guttormsen, & Nexo, 2006). TCN2 bound to cobalamin, also called holotranscobalamin (holoTC), is the biologically active form of cobalamin in the body and has a half-life of approximately 2 hr (Lindgren, Kilander, Bagge, & Nexo, 1999). The biologically active cobalamin/TCN2 complex binds to the transcobalamin II-receptor (TCblR) found in the plasma membrane of most cell types. The receptor/TCN2/cobalamin complex is endocytosed and transported to several subcellular compartments (Christensen & Birn, 2002). Approximately 80% of the circulating cobalamin is bound to transcobalamin I and III (haptocorrin). This complex remains in the circulation with a 10-day half-life (Carmel, 1985). In addition to acting as a storage pool for cobalamin, haptocorrin removes potentially toxic cobalamin analogs from the bloodstream. Haptocorrin binds to the liver-specific asialoglycoprotein receptor. As these receptors are not expressed in other tissues, the role of haptocorrin in transporting the active form of cobalamin is unclear (Burger, Schneider, Mehlman, & Allen, 1975; Hardlei & Nexo, 2009).
Biochemically, cobalamin is a cofactor for two essential enzymatic reactions. In the first, methylcobalamin serves as a cofactor for the cytoplasmic enzyme methionine synthase. This enzyme transfers methyl groups from 5-methyl-tetrahydrofolate to homocysteine, producing tetrahydrofolate for nucleic acid synthesis and methionine for methylation reactions and protein synthesis. The second cobalamin-dependent reaction is catalyzed by the mitochondrial enzyme
The pathophysiology of cobalamin deficiency manifests hematologic and neurologic sequelae. When cobalamin availability is low, nucleic acid synthesis is compromised (reviewed in Shane & Stokstad, 1985). In rapidly dividing hematopoietic precursor cells, abnormal DNA synthesis halts cell division, which affects up to 90% of a patient’s red blood cell precursors (Aster, 2005). Clinical testing shows decreased hemoglobin concentration and elevated mean corpuscular volume (MCV), the joint presence of which is a classic hallmark indicator of megaloblastic anemia (Andres et al., 2006). Because cobalamin metabolic pathways overlap with folate metabolism in the cytoplasm, megaloblastic work-ups must always include evaluation of a cobalamin metabolite profile in addition to serum folate. Well-meaning health care providers aiming to treat megaloblastic anemia with folic acid without fully reviewing metabolic cobalamin parameters can improve the patient’s hematologic status while leaving neurologic pathology untreated and likely to progress (Pfeiffer, Caudill, Gunter, Osterloh, & Sampson, 2005).
Cobalamin deficiency has long been associated with age-related neuropathology, but the molecular mechanisms remain unknown (reviewed in Reynolds, 2006). In older adults, cobalamin deficiency can present with a heterogeneous preclinical spectrum impacting multiple facets of health. For example, older adults with preclinical cobalamin deficiency experience nonspecific symptoms erroneously attributed to aging, including malaise, vertigo, fatigue, mood alterations such as depression, and sleep and gait disturbances (Carmel, 2000). In a number of studies, supplementation with a combination of vitamin B12, vitamin B6, and folic acid has slowed the rate of brain atrophy and cognitive decline, suggesting a potential role for vitamin B12 in these age-related neurological changes (de Jager, Oulhaj, Jacoby, Refsum, & Smith, 2012; Douaud et al., 2013; Smith et al., 2010). Factors such as medications that impair absorption, age-related decreases in gastric function, and comorbidities that drive increased needs result in a high prevalence of cobalamin deficiency for older adults, estimated at 6% to more than 40% (Allen, 2009; Baik & Russell, 1999; Johnson et al., 2003; Lindenbaum, Rosenberg, Wilson, Stabler, & Allen, 1994; Pennypacker et al., 1992). Unrecognized cobalamin deficiency in older adults may result in an exaggerated disability trajectory involving higher frequency of hospital admissions, lengthier and more severe hospitalizations, or greater degrees of chronic disablement significantly affecting mobility and quality of life (Bartali et al., 2006). Recently, the geriatric syndrome of frailty, a chronic and progressive disablement stemming from age-related declines in energy metabolism and the inflammatory, endocrine, skeletal muscle, and neurologic systems, has been linked to the cobalamin metabolite MMA (Matteini et al., 2008).
Disablement and frailty in elderly persons is a significant public health concern because of increased vulnerability to morbidity, mortality, and adverse health outcomes (Fried et al., 2001). Preventing or delaying the disablement and frailty process in older adults is a key area of research, and improved understanding of the genetic factors that contribute to its development may allow the identification of susceptible individuals and suggest personalized strategies for intervention and prevention (Walston et al., 2006). For example, nurses caring for elderly patients experiencing vague, nonspecific symptoms could query their genetic makeup and combine this with laboratory tests to determine if cobalamin could be used to prevent disability or frailty.
Genetic variants affecting proteins that transport cobalamin in blood and deliver it into cells may influence the amount of cobalamin available to cells. The TCN2 carrier molecule and its receptor (TCblR) are an important ligand and receptor in which structural variations can affect vitamin B12 binding. Altered TCN2 structure results in decreased cellular availability for essential metabolic reactions and influences susceptibility to deficiency (Miller, Ramos, Garrod, Flynn, & Green, 2002). To explore these factors, we tested for associations between common genetic variants in the TCN2 and TCblR genes and cobalamin-related biochemical and clinical outcomes in 795 Women’s Health and Aging Studies (WHAS) participants.
Materials and Methods
Study Design and Population
We conducted a candidate gene association study using genetic material from participants in the WHAS. Because allele frequencies and physiologic and metabolic traits can differ between Caucasians and African Americans, we included genetic variants unique to each population and used race and genotype as independent variables in our study design (1000 Genomes Project Consortium et al., 2012; Cheng, Chan, Cembrowski, & van Assendelft, 2004; Stabler et al., 1999).
Previous authors have described the WHAS in detail (Fried, Bandeen-Roche, Chaves, & Johnson, 2000; Guralnik, Fried, Simonsick, Bandeen-Roche, & Kasper, 1995). In brief, WHAS 1 (1992–1995) and WHAS 2 (1994–1996) were prospective, observational research initiatives examining physical-disability trajectories in community-dwelling elderly women in Baltimore, MD. WHAS 1 sampled the one-third most disabled women and WHAS 2 sampled the two-thirds least disabled women; WHAS subject data can be pooled to represent the community-dwelling elderly (Bandeen-Roche et al., 2006; Chaves, Ashar, Guralnik, & Fried, 2002; Chaves et al., 2005; Leng, Xue, Tian, Walston, & Fried, 2007; Semba, Garrett, Johnson, Guralnik, & Fried, 2000; Semba et al., 2005; Walston et al., 2005). The WHAS protocols were approved by the Johns Hopkins Medical Institutions Institutional Review Board. The genotyping portion of this research was approved by the Office of Human Subjects Research at the National Institutes of Health and the Institutional Review Board of the University of Utah.
Outcome Variable Selection
The overarching hypothesis of this study was that genetic variation contributes to the clinical heterogeneity observed for cobalamin deficiency in older adults. The use of a population with well-characterized and measured phenotypes is instrumental in exploring genetic associations. For this reason, we generated genetic data on blood samples from WHAS to take advantage of the extensive and sophisticated clinical trait information available on these study participants.
We selected nine outcome variables from the broader WHAS data set to reflect a cobalamin deficiency phenotype in the older adult; these include biochemical, hematologic, neurologic, and functional performance parameters. For biochemical outcomes, we chose measurements commonly used in the diagnosis and management of cobalamin deficiency, namely, serum cobalamin, homocysteine, and MMA. For hematologic outcomes, we chose hemoglobin and MCV, as low cobalamin produces hallmark changes of megaloblastic anemia. For neurologic outcomes, we chose the following clinical measures known to be associated with age-related cobalamin deficiency: depression, as measured by the Geriatric Depression Scale (GDS) and peripheral neuropathy, as measured by vibration perception testing (VPT). For functional performance outcomes, we used measures of decreased muscular strength and capacity that indicate altered mitochondrial energy/oxidation states, that is, hand grip strength and 4-meter walking speed.
Blood Analyte Measurements
Measurements of serum cobalamin, folate, creatinine, homocysteine, MMA (Penninx et al., 2000; Stabler et al., 1999), hemoglobin, and MCV (Chaves et al., 2006; Williamson et al., 1995) were obtained as previously reported.
Previous studies of elderly individuals have demonstrated that modest elevations in homocysteine and MMA can occur as a result of renal insufficiency (Lindenbaum et al., 1994; Pennypacker et al., 1992; Stabler, Lindenbaum, & Allen, 1996). However, in older adults, singular use of serum creatinine as a kidney function assessment is recognized to significantly underascertain presence of impaired renal status (Giannelli et al., 2007). In this study, we used serum creatinine to estimate creatinine clearance for WHAS 1 and 2 subjects through the Cockcroft–Gault formula: creatinine clearance (ml/s) × weight (kg) × [140 − age (year)]/[72 × serum creatinine (mg/dl) ×0.85] (Cockcroft & Gault, 1976).
Neurologic Clinical Measures
Depression
Depressive symptoms were evaluated with the GDS, a psychometric depression assessment for elderly adults (Feher, Larrabee, & Crook, 1992; Yesavage et al., 1982). The GDS is a validated 30-item survey comprising dichotomous (yes/no) questions and requiring approximately 8 min for administration.
Peripheral neuropathy
VPT was conducted to assess for peripheral neuropathy. As WHAS 1 protocols used a vibrometer and WHAS 2 protocols used a tuning fork, WHAS 2 subject data were excluded to ensure reliability. In a test modeled after a diabetic neuropathy protocol, subjects placed their right toe on the Vibratron II’s (Physitemp Instrument, Inc., Clifton, NJ) platform and reported a vibratory stimulus using a yes/no forced choice procedure (Ferrucci, Kittner, Corti, & Guralnik, 1995; Maser et al., 1989). Measured vibration units were converted to microns and a mean vibratory threshold was identified (Resnick, Vinik, Heimovitz, Brancati, & Guralnik, 2001; Volpato, Leveille, Blaum, Fried, & Guralnik, 2005).
Functional Performance Measures
Hand grip strength
A JAMAR hand dynamometer was used to measure hand grip strength (Model #BK-7498; Fred Sammons Inc., Burr Ridge, IL). Testing was performed with subjects in a seated position and elbow flexure at a 90° angle. Subjects were asked to grasp the dynamometer and squeeze as hard as possible 3 times on each hand; the best measure in the stronger hand was recorded.
4-meter walking speed
Subjects walked a 4-meter course at their usual speed two times and once more as fast as possible. Using the faster of the two usual-pace walks, average walking speed was calculated by dividing the length of the walk (in meters) by the time in seconds required to complete it. Subjects were permitted to use a cane, walker, or walking aid but not assistance from another person.
Genetic Variant Measurement
Single nucleotide polymorphism (SNP) selection
SNPs were selected to cover common variation in TCN2 and TCblR using public databases (HapMap [Phases 1 and 2 full data set], dbSNP [Build 125], and NCBI [Build 35]). For both Caucasian and African American ancestral groups, Haploview (4.0) Tagger (Broad Institute) was used to select tag SNPs in the candidate genes, including 10 kilobases (kb) flanks on both ends of the genes. Because both candidate genes were relatively small (19 kb and 6 kb) and there was high likelihood of dropped assays due to working with whole-genome amplified (WGA) material, we built redundancy into our assays. An r 2 threshold was set at 0.9 and minor allele frequency at 0.05. Additional candidate SNPs included variants with probability of exerting a functional effect, such as those located in or near exons or promoters. A total of 51 SNPs were selected, that is, 29 SNPs in TCN2 and 22 SNPs in TCblR.
SNP genotyping
Genotyping of the WHAS WGA samples was performed using the Sequenom MassArray iPLEX platform (San Diego, CA). Allele-specific primer extension products were resolved using matrix-assisted laser desorption ionization - time of flight (MALDI-TOF) mass spectroscopy. Primer sequences (Supplementary Table 1) were designed and used as described in the published protocol (Gabriel, Ziaugra, & Tabbaa, 2009).
To ensure quality of genotypes obtained with WGA DNA, all samples were genotyped twice and unresolved discordant genotypes dropped. For each SNP, an average of 93.8% (range: 90.6–97.1%) of all samples were successfully typed in duplicate. The average concordance of duplicate genotypes was 99.3% (range: 97.8–100%). The average call rate was 95.2% (range: 92.2–98.4%). SNPs with large departures from Hardy–Weinberg equilibrium (HWE, p < .001) in either racial subset were excluded. The only exception was the retention of TCblR rs2336573 (p = 4.9 × 10−13) because previous work indicates that genotyping error is an unlikely explanation for failure to adhere to HWE for this SNP (Pangilinan et al., 2010).
Statistical Analysis
Descriptive and inferential statistics were analyzed using the SPSS program (v12.0 and v20.0). For the study’s sociodemographic and health characteristics and outcome variables, summary statistics, frequency distributions, and independent t-tests were evaluated. Assessment of the outcome and covariate variables for linearity, homogeneity of variance, homogeneity of regression slopes, and measurement reliability indicated no violations of statistical assumptions. Severe outliers (>3 standard deviations away from the mean) were removed from analysis.
Two-way analysis of variance (ANOVA) and covariance (ANCOVA) tests were conducted to evaluate associations between the biochemical, hematological, neurologic, and physical-performance outcome parameters and the two independent variables, SNP genotype and race. WHAS subjects were placed into one of the three genetic categories (AA, AB, and BB) according to their genotypes and stratified by race. Several outcomes in the two-way ANCOVA models were adjusted for appropriate covariates. Significant (p < .05) SNP main effect F tests were explored further using the Tukey least significant difference procedure. The Bonferroni adjustment was used to correct for α inflation arising from multiple testing.
Results
Demographic and Clinical Characteristics
Demographic and other baseline health characteristics of the 789 WHAS study participants are shown in Table 1. The 591 Caucasian participants represented 75% of the study sample and had more years of total education and lower mean body mass index. Biochemical and clinical outcome summary statistics are shown in Table 2. There were significant differences between Caucasians and African Americans, making race an important independent variable. Because there are several hundred publications devoted to investigation of WHAS subjects, we do not provide a detailed discussion of sample characteristics here (Chaves, Garrett, & Fried, 2000; Ferrucci, Guralnik, et al., 1995; Fried et al., 2000; Kasper & Simonsick, 1995; Penninx et al., 2000; Stabler et al., 1999; Williamson et al., 1995).
Selected Sociodemographic and Health Characteristics for WHAS Subjects.
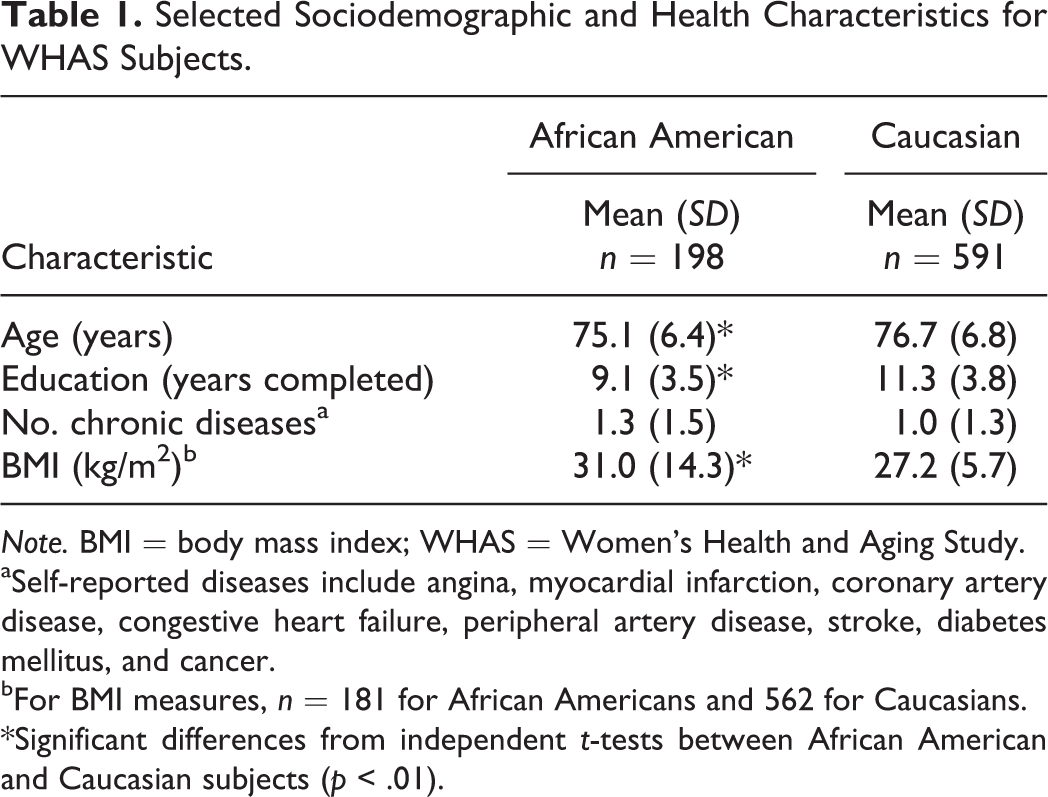
Note. BMI = body mass index; WHAS = Women’s Health and Aging Study.
aSelf-reported diseases include angina, myocardial infarction, coronary artery disease, congestive heart failure, peripheral artery disease, stroke, diabetes mellitus, and cancer.
bFor BMI measures, n = 181 for African Americans and 562 for Caucasians.
*Significant differences from independent t-tests between African American and Caucasian subjects (p < .01).
Outcome Variable Summary Statistics by Race.
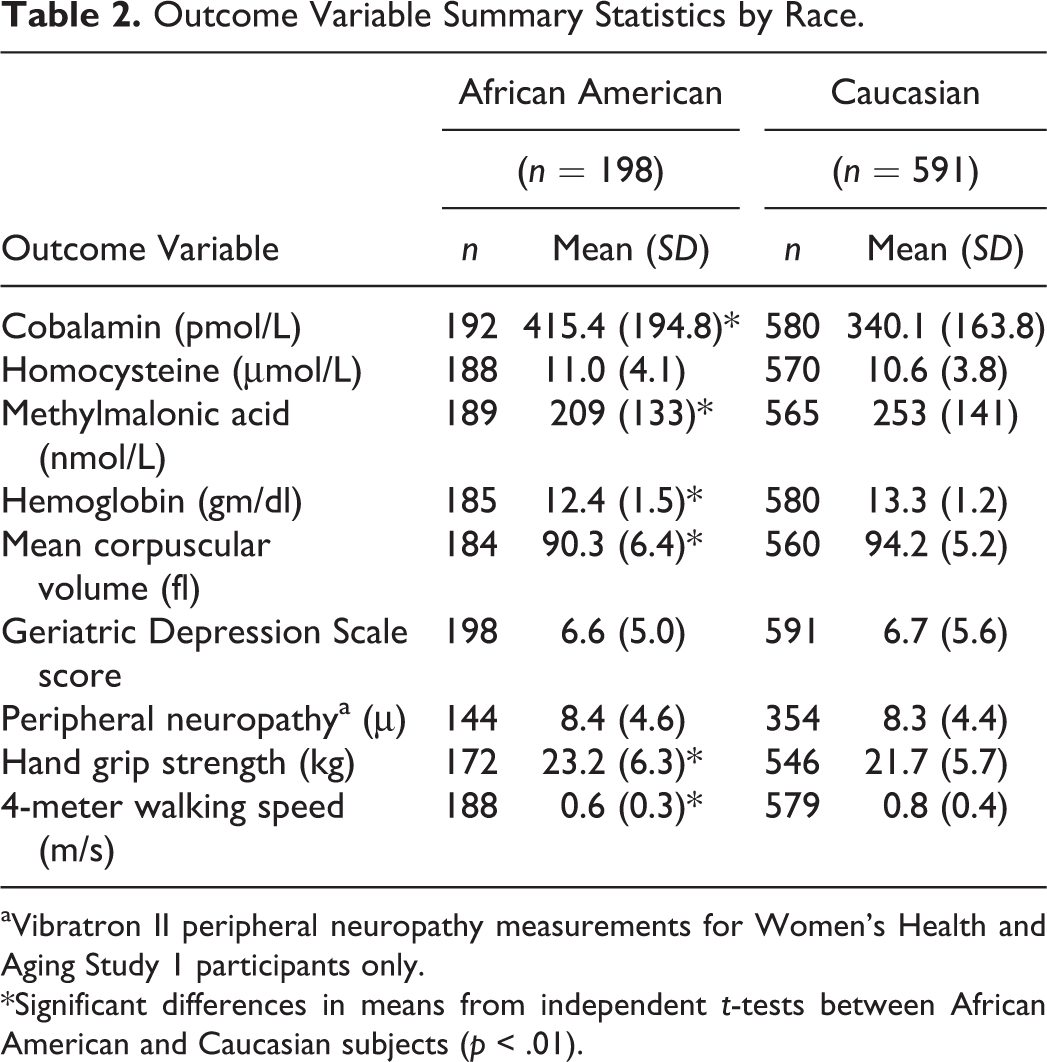
aVibratron II peripheral neuropathy measurements for Women’s Health and Aging Study 1 participants only.
*Significant differences in means from independent t-tests between African American and Caucasian subjects (p < .01).
Genetic Data
Of the total 51 SNPs selected for genotyping in TCN2 and TCblR, 38 SNPs were successfully genotyped. Of these, 6 SNPs demonstrated significant, unexplained HWE departures and were removed from analysis. Genotype and allele frequencies for the remaining genotyped SNPs are presented in Table 3. Of these, 11 SNPs had minor allele frequencies <0.05 in both Caucasian and African American populations and were excluded. A final total of 21 SNPs were entered into the ANOVA and ANCOVA models: 14 SNPs from the TCN2 gene and 7 from the TCblR gene. Bonferroni adjustment for multiple testing of the 21 SNPs across the nine study outcomes yielded a corrected α significance threshold of 0.0003 (0.05/189).
Genotype and Allele Frequencies for TCN2 and TCblR Single Nucleotide Polymorphisms.
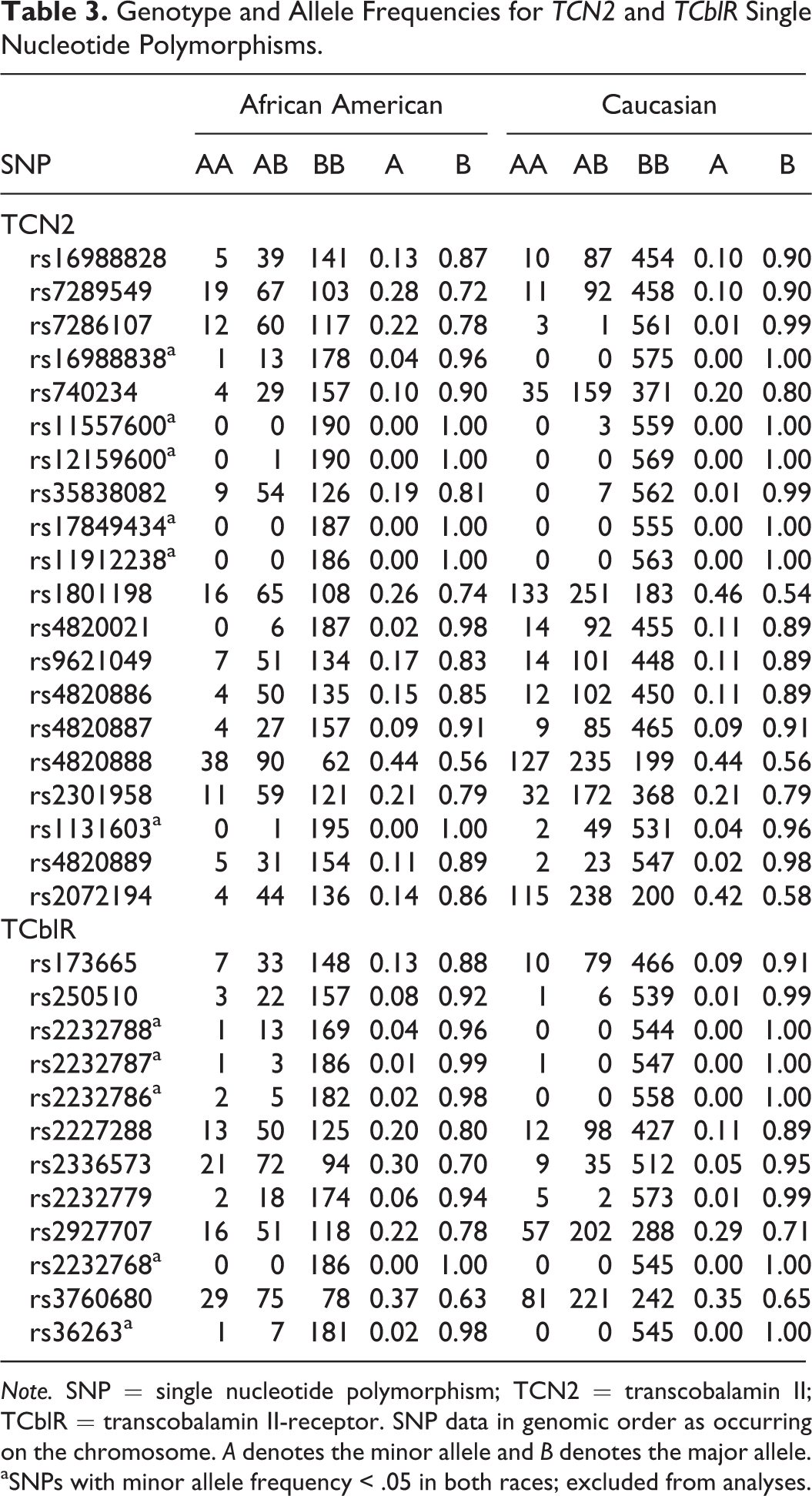
Note. SNP = single nucleotide polymorphism; TCN2 = transcobalamin II; TCblR = transcobalamin II-receptor. SNP data in genomic order as occurring on the chromosome. A denotes the minor allele and B denotes the major allele.
aSNPs with minor allele frequency < .05 in both races; excluded from analyses.
ANOVA and ANCOVA Findings
Table 4 provides a summary of the significant genetic associations we obtained. Due to well-characterized differences between Caucasians and African Americans in the WHAS sample for most study outcomes, we highlight only significant genetic findings.
TCN2 and TCblR Two-Way Analysis of Covariance (ANCOVA) and Variance (ANOVA) Significant SNP Association Results.
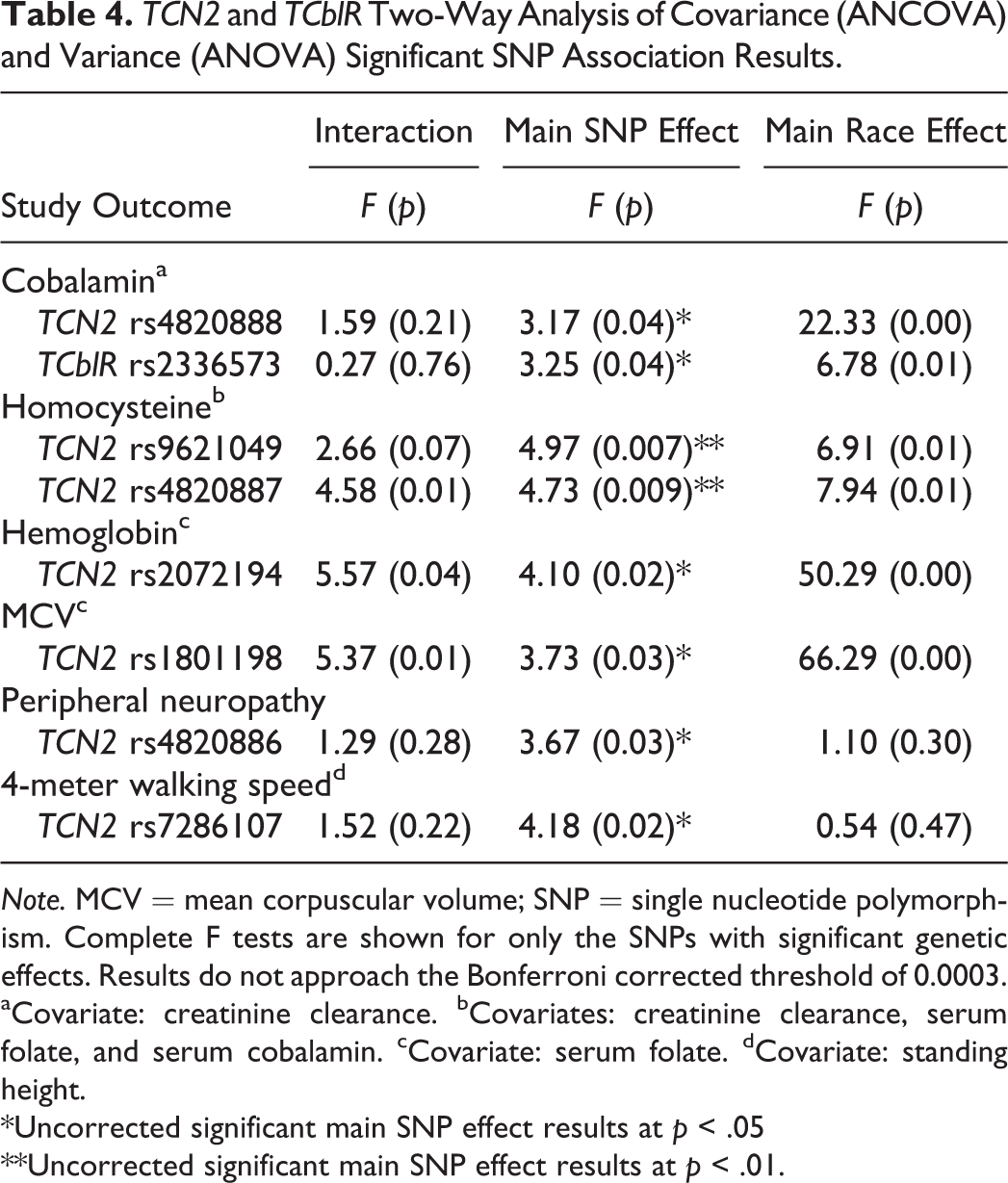
Note. MCV = mean corpuscular volume; SNP = single nucleotide polymorphism. Complete F tests are shown for only the SNPs with significant genetic effects. Results do not approach the Bonferroni corrected threshold of 0.0003.
aCovariate: creatinine clearance. bCovariates: creatinine clearance, serum folate, and serum cobalamin. cCovariate: serum folate. dCovariate: standing height.
*Uncorrected significant main SNP effect results at p < .05
**Uncorrected significant main SNP effect results at p < .01.
No genetic main effect reached the stringent Bonferroni-adjusted threshold level of significance of 0.0003. However, eight SNPs were significant at the p < .01 or p < .05 level. For these eight SNPs, Table 5 summarizes significant post hoc comparison tests at the p < .05 and p < .01 level observed for genotype group comparisons in the overall WHAS sample and within stratified race groups.
Post Hoc Results, Women’s Health and Aging Study (WHAS) Participant Means and Sample Sizes by Race for Significant Single Nucleotide Polymorphism (SNP) F-tests.
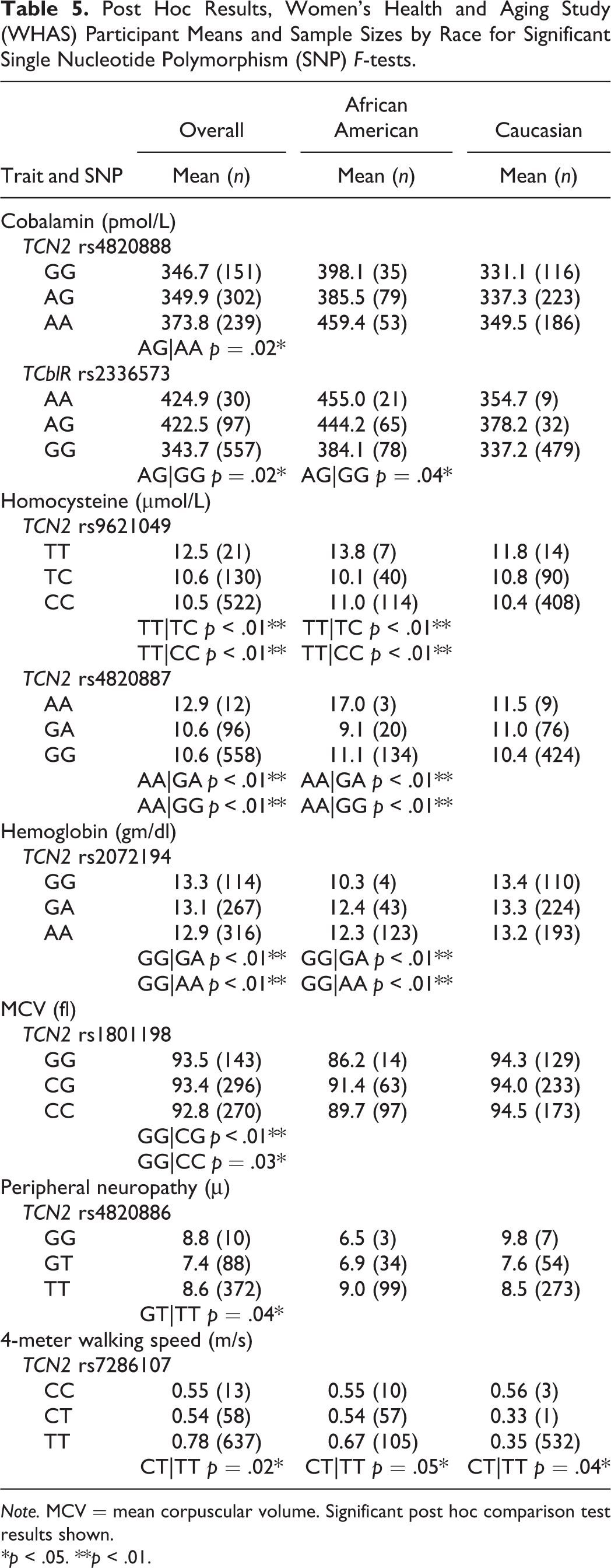
Note. MCV = mean corpuscular volume. Significant post hoc comparison test results shown.
*p < .05. **p < .01.
Cobalamin
We identified two significant SNP associations for cobalamin level, TCN2 rs4820888 and TCblR rs2336573. TCN2 rs4820888 A>G is located in intron 7 and showed a statistically significant main genetic effect for cobalamin, F(2, 692) = 3.17, p = .04. TCblR rs2336573 G>A is a missense polymorphism located in exon 4, resulting in a glycine-to-arginine amino acid change at codon position 220 (G220R). There was a significant association between rs2336573 and serum cobalamin levels, F(2, 684) = 3.25, p = .04.
Homocysteine
We identified two significant SNP associations for homocysteine level, TCN2 rs9621049 and TCN2 rs4820887. TCN2 rs9621049 is a C>T missense polymorphism in exon 7 that results in an amino acid change of a serine to phenylalanine at codon position 348 (S348F). This SNP demonstrated the strongest level of statistical significance for a genetic effect, F(2, 673) = 4.97, p = .007, among all of the SNPs. The second strongest association in this study was between mean homocysteine levels and TCN2 rs4820887, a G>A SNP in intron 7, F(2, 666) = 4.73, p = .009.
Hematologic parameters
We identified two significant SNP associations with hematologic parameters, TCN2 rs2072194 with mean hemoglobin concentration and TCN2 rs1801198 with MCV. TCN2 rs2072194 is an A>G polymorphism in intron 8 of the TCN2 gene and was significantly associated with mean hemoglobin concentrations of WHAS subjects, F(2, 700) = 4.10, p = .02. TCN2 rs1801198 is a missense C>G polymorphism in exon 6 that results in an amino acid change of a proline to an arginine at codon position 259 (P259R). It demonstrated a significant association with MCV, F(2, 709) = 3.73, p = .03. Important to interpreting this result is the difference between the Caucasian subjects’ mean MCV (94.2 fl) and the African American subjects’ mean MCV (90.3 fl). We observed a bimodal peak in the outcome variable’s distribution curve, and Levene’s test for equality of variances was violated (data not shown). SNPs for this outcome not meeting the more stringent Bonferroni-corrected α significance levels should be cautiously interpreted (Tabachnik & Fidell, 2007). For this study, it may be reasonable to view the positive association for rs1801198 with additional caution.
Neurologic parameters
We identified one significant SNP association with a neurologic parameter, TCN2 rs4820886 with peripheral neuropathy mean micron measurements. TCN2 rs4820886 is a G>T polymorphism in intron 7 and demonstrated a significant main genetic effect, F(2, 470) = 3.67, p = .03.
Functional performance parameters
One SNP in TCN2, rs7286107, was associated with 4-meter walking speed. TCN2 rs7286107 is a C>T polymorphism in intron 1 and is close to an area known to impact gene regulation. TCN2 rs7286107 was significantly associated with mean walking speeds, F(2, 708) = 4.18, p = .02.
Discussion
We examined the associations of genetic variants with selected metabolic and clinical outcome parameters in WHAS participants, including serum cobalamin, its two primary metabolites (homocysteine and MMA), and six cobalamin-deficiency clinical phenotype characteristics. Although no result reached the stringent Bonferroni threshold of p = .0003, eight variants were significant at the p = .007 to p = .04 level. Of these, five variants (TCN2 rs9621049, TCN2 rs4820887, TCN2 rs2072194, TCN2 rs4820886, and TCN2 rs7286107) had decreased cell sizes (<10), where genotype group differences in the overall WHAS sample could affect strength of the reported association. Stratifying the sample clinical characteristics produces some groups with small numbers, making adequate comparisons difficult for some SNPs in this study. Population genotype distributions in this study are similar to those reported in HapMap data.
Three associations involved coding SNPs (TCblR rs2336573, TCN2 rs9621049, and TCN2 rs1801198). The first is an association between the TCblR G220R missense coding SNP (rs2336573) and the mean serum cobalamin concentrations. Regardless of race and accounting for estimated creatinine clearance, individuals with a GG (220GG) genotype had significantly lower mean serum cobalamin, demonstrating a difference of 81.2 pmol/L from subjects with an AA (220RR) genotype. This SNP is strongly linked to a neighboring SNP (TCblR rs150384171, D′ = 1, r 2 = 0.39) on the same chromosome, a phenomenon called linkage disequilibrium (LD). The TCblR rs150384171 SNP is a three base-pair deletion in exon 2 that results in a glutamic acid deletion. We previously reported the stronger influence of TCblR rs150384171 on serum cobalamin compared to an SNP in perfect LD with TCblR rs2336573 (rs9426, D′ = 1, r 2 = 1; Stone et al., 2011). Although direct testing of rs150384171 was not possible in our WHAS sample due to genotyping platform incompatibility and low minor allele frequency (<0.01), it remains the best candidate for being the underlying causal variant driving the detected association between serum cobalamin and TCblR rs2336573.
A second key finding was the association between the TCN2 S348F missense SNP (rs9621049) and the serum homocysteine levels. After adjusting for several covariates, TT (348FF) homozygotes have ∼2.0 μmol/L higher mean homocysteine concentrations than CC (348SS) homozygotes or CT (348SF) heterozygotes (p = .003). Although the strength of this association is not definitive, we obtained very similar results for TCN2 rs4820887, in intron 7. TCN2 rs9621049 and rs4820887 share significant LD (D′ = 1, r 2 = 0.678), indicating that they are not independent. As a missense SNP, rs9621049 (S348F) is an attractive candidate for being functional and directly influencing homocysteine levels.
The association between TCN2 S348F (rs9621049) and homocysteine we found in this study is not consistent with findings of a previous study. Lievers et al. (2002) failed to find an association between homocysteine and TCN2 S348F (rs9621049) but did find an association between the TCN2 P259R missense SNP (rs1801198) and homocysteine that was dependent upon total serum cobalamin level. These differences may be due to dissimilarity in patient sampling and demographics, as Lievers et al. (2002) studied an ethnically Dutch population of 190 middle-aged individuals with cardiovascular disease and 601 normal healthy control subjects.
Although TCN2 rs1801198 was not associated with homocysteine in our study, it was associated with MCV—our third key finding. After accounting for serum folate, individuals with a GG (259RR) genotype had significantly higher MCV, demonstrating a 0.7 fl difference from subjects with a CC (259PP) genotype. Interestingly, African Americans with a GG genotype had significantly lower MCV values compared to other genotype groups, which contrasted with the overall P259R GG genetic effect. Based on the distribution of MCV by genotype in each race (Table 5 and data not shown), this result may be driven by a race-specific effect in African Americans. The racial differences in our MCV data are consistent with published population-based hematologic studies indicating that African American individuals’ hemoglobin and MCV are biologically lower than those of Caucasian individuals (Cheng et al., 2004).
Although this research is the first to report an association between TCN2 rs1801198 and the MCV trait, previous research has suggested that this SNP exerts a functional biologic effect. Miller, Ramos, Garrod, Flynn, and Green (2002) reported that the rs1801198 genotype influenced serum holoTC and MMA concentrations in a sample of 128 healthy adults at the University of Missouri. A clinical study identified interactions between total serum cobalamin on homocysteine and serum holoTC on homocysteine, as modified by TCN2 P259R (rs1801198) SNP genotype, in a cohort of 554 community-dwelling elderly Latinos in northern California (Garrod, Allen, Haan, Green, & Miller, 2010). A Norwegian study of 10,601 adult cancer–registry patients identified a positive association between TCN2 rs1801198 genotype and serum MMA (Fredriksen et al., 2007). And finally, an Irish study of 613 adult men demonstrated that homozygous patients with the C allele for rs1801198 had lower serum cobalamin and homocysteine concentrations (Stanislawska-Sachadyn et al., 2010). Although our results should be interpreted with caution, there is precedent for our P259R finding, as this SNP has been reported to be associated with other cobalamin-related traits.
The work outlined in this study extends the previously published WHAS reports on the evaluation of MMA with SNPs in TCN2 (Matteini et al., 2008, 2010). In these previous candidate gene analyses, the authors reported no significant associations between MMA levels and TCN2 SNPs. However, the WHAS sample size of 326 Caucasians in those studies was substantially less compared to the presently reported research (591 Caucasians) and did not include African Americans.
Strengths and Limitations
Strengths of this research include the use of a well-characterized elder adult cohort to ascertain effects of genetic variation on metabolic parameters of cobalamin deficiency. As there was limited quantity of genomic starting material, this study demonstrates that use of whole-genome amplification is a sound technique to augment previous clinical and epidemiological research initiatives on archived samples. Also notable is that this investigation includes examination of genetic variation of two genes coding for protein products that directly interact at the level of the cell.
A key limitation of this study is failure to generate high-quality data for all TCN2 and TCblR SNPs in every study subject. This failure stemmed from implementation of stringent quality control criteria and genotyping challenges associated with using WGA material produced from limited DNA in storage for nearly 20 years. We anticipated this difficulty and designed assays to redundantly cover these genes. Discounting four Caucasian-specific SNPs and nine African American–specific SNPs in these two genes, our coverage was nearly complete at a stringent r 2 threshold of 0.9.
Another limitation to this work is that environmental factors (i.e., medications and comorbidities) known to affect cobalamin deficiency were not included in these analyses. Because many of the SNPs had low allele frequencies in African American and Caucasian subgroups and several covariates were already used, additional covariates would have further reduced power and removed our ability to detect genetic effects.
Finally, this study measured total serum cobalamin as opposed to holoTC concentration; only 20% of the total cobalamin is bound by transcobalamin II, with the remaining 80% bound to haptocorrin. However, previous WHAS investigation of cobalamin metabolites did not include holoTC concentration and there was insufficient participant sample volume to run this test.
Nursing Implications
Despite the limitations of this study, our findings suggest that there are additional biologic factors contributing to an elderly adult’s susceptibility to decreased cobalamin nutritional status. In that the estimated frequency of this condition ranges from 6% to more than 40% and the disability trajectories are more severe for these patients, nursing interventions could be designed for individuals at risk of progressing from subclinical disease to full blown deficiency. For example, older adults presenting with vague, heterogeneous preclinical symptoms (e.g., malaise, vertigo, fatigue, mood alterations, peripheral insensitivity, sleeping, and gait disturbances) and demonstrating a higher risk SNP profile in key cobalamin metabolism genes may be candidates for cobalamin-deficiency testing and benefit from early intervention with regular cobalamin supplementation via intramuscular injections.
Presently, it is not known whether elevated cobalamin metabolites are mediated by genetic factors such as the SNPs in this study and an older adult’s cobalamin intake. Although genome-wide associations studies have identified new cobalamin-related genes (Hazra et al., 2009; Tanaka et al., 2009), this study focused on evaluating all common variants in two candidate genes involved in cobalamin delivery to cells. Additional nursing research in more highly powered and diverse patient cohorts is needed to replicate the TCblR rs2336573 (G220R), TCN2 rs9621049 (S348F), and TCN2 rs1801198 SNP (P259R) findings reported here. However, this work may contribute to identification of valuable cobalamin-related targets for future studies, leading to more accurate diagnostics and supplementation regimens for older adults experiencing preclinical symptoms.
Footnotes
Authors’ Note
This article partially fulfilled doctoral requirements from the University of Utah, College of Nursing.
Author Contributions
E. Kurnat-Thoma contributed to conception, design, data acquisition, data analysis, and interpretation; drafted and critically revised the article; gave final approval; and agrees to be held accountable for all aspects of work ensuring integrity and accuracy. F. Pangilinan contributed to design, data acquisition, data analysis, and interpretation; critically revised the article; gave final approval; and agrees to be held accountable for all aspects of work ensuring integrity and accuracy. A. Matteini contributed to design, data acquisition, data analysis, and interpretation; critically revised the article; gave final approval; and agrees to be held accountable for all aspects of work ensuring integrity and accuracy. B. Wong contributed to data analysis and interpretation; critically revised the article; gave final approval; and agrees to be held accountable for all aspects of work ensuring integrity and accuracy. G. Pepper contributed to data analysis and interpretation; critically revised the article; gave final approval; and agrees to be held accountable for all aspects of work ensuring integrity and accuracy. S. Stabler contributed to data acquisition and interpretation; critically revised the article; gave final approval; and agrees to be held accountable for all aspects of work ensuring integrity and accuracy. J. Guralnik contributed to conception, design, data acquisition, and interpretation; critically revised the article; gave final approval; and agrees to be held accountable for all aspects of work ensuring integrity and accuracy. L. Brody contributed to conception, design, data acquisition, data analysis, and interpretation; critically revised the article; gave final approval; and agrees to be held accountable for all aspects of work ensuring integrity and accuracy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Intramural Research Programs of the National Human Genome Research Institute and the National Institute on Aging (National Institutes of Health). Dr. Kurnat-Thoma was supported by a predoctoral Intramural Research Training Award from the National Institute of Nursing Research.