
Abstract
Although the effect of gene–gene interaction on nicotine–dopamine metabolism for smoking behavior has been reported, polymorphisms of dopamine D2 receptor (DRD2) and monoamine oxidase A (MAOA) have not been simultaneously examined among smokers. In this study, 481 young Taiwanese men completed a self-report questionnaire on smoking status, and data were obtained on polymorphisms of DRD2 rs1800497, DRD2 rs1079597, MAOA rs309850, and MAOA rs1137070, urinary nicotine, and urinary cotinine. In a comparison of 261 current smokers and 220 never smokers, odds ratios (ORs) for the development of smoking in all genotypes were not statistically significant. Among smokers with DRD2 rs1079597 GG//MAOA rs309850 3-repeat, the OR of heavier smoking was 2.67 times higher (95% confidence interval [CI]: [1.08, 6.59], p = .031) and the score on the Fagerstrom test for nicotine dependence was higher (4.26 vs. 2.83) than in those with DRD2 rs1079597 AA//MAOA rs309850 3-repeat. Adjusted urinary cotinine concentration was significantly different between those two groups (median value: 95.83 ng/μl vs. 133.24 ng/μl, respectively, p = .045). These findings suggest that the interaction of DRD2 rs1079597 and MAOA rs309850 3-repeat affects smoking intensity in young Taiwanese men.
Tobacco smoking is a global health problem and Taiwan is no exception (Health Promotion Administration, 2013). Nicotine is the major psychoactive ingredient in tobacco (Henningfield, Cohen, & Pickworth, 1993). The addictive nature of nicotine is multifactorial, involving both environmental and genetic factors (Agrawal et al., 2012; Caron, Karkazis, Raffin, Swan, & Koenig, 2005). The interaction between nicotine and dopamine should be considered when exploring these genetic factors. Nicotine activates dopaminergic neurons and enhances dopamine release, which leads to feelings of pleasure and reward (Fowler & Kenny, 2014).
Of the genetic variants associated with smoking, one of the most widely studied is the dopamine D2 receptor (DRD2) polymorphism. In particular, researchers have increasingly studied DRD2 rs1800497 (DRD2 TaqI A) and DRD2 rs1079597 (DRD2 TaqI B) in this context (Ohmoto et al., 2013; Spitz et al., 1998). DRD2 is localized in chromosome 11 q22-q23. The C-to-T transition of the DRD2 rs1800497 polymorphism causes an amino acid change (Glu713Lys), which seems to significantly reduce the specificity of receptor binding (Vereczkei et al., 2013). The A-to-G substitution of DRD2 rs1079597 may alter DRD2 density in the striatum (Suriyaprom, Tungtrongchitr, & Harnroongroj, 2013). Investigators have been studying the monoamine oxidase A (MAOA) gene, located in the short arm of the X chromosome in relation to smoking behavior since 2005 (S. Huang et al., 2005). In particular, researchers have typically focused on two polymorphisms of MAOA: MAOA rs309850, a variable number tandem repeat polymorphism, and MAOA rs1137070 (MAOA-LPR), an EcoRV polymorphism (Jin et al., 2006). Researchers have reported that MAOA rs309850 3- and 4-repeat polymorphisms have low and high activity, respectively, in the metabolism of monoamines (Hu et al., 2013). The MAOA rs1137070 C > T allele reduces MAOA enzyme activity (Shiels et al., 2008).
Prior research has reported the effects of gene–gene interaction in relation to nicotine–dopamine metabolism in smoking behavior: for example, interaction between DRD2 and monoamine oxidase B (MAOB; Costa-Mallen, Costa, & Checkoway, 2005) and between MAOA and cytochrome P450 2A6 (Tang et al., 2009). However, no previous research has examined DRD2 and MAOA simultaneously in smokers. Researchers have been investigating environmental factors among Taiwanese smokers since 2005, and reliable and valid Chinese-language instruments to measure smoking behaviors are well established (C. L. Huang, Cheng, & Huang, 2013; C. L. Huang, Cheng, & Wang, 2014; C. L. Huang, Lin, & Wang, 2010). Research on genetic factors for Taiwanese smokers began in 2013 and only one (μ-opioid receptor) gene has been investigated (Chen et al., 2013).
The purpose of the present study was to collect genetic risk information and increase our understanding of smoking status in Taiwanese subjects. Since Hu et al. (2013) observed a possible interaction between DRD2 and MAOA in association with alcoholism in Taiwanese subjects, we hypothesized that single nucleotide polymorphism as well as the interaction between those two genes may be related to smoking status. We recruited only male smokers as participants for the following reasons: Nicotine and cotinine clearances are 13% and 24% higher, respectively, in women not using oral contraceptives than in men (Benowitz, Hukkanen, & Jacob, 2009). There are gender differences in relation to craving, affect, and preference for immediate smoking after cue exposure (Doran, 2014). Taiwanese men report smoking significantly more cigarettes than Taiwanese women (C. L. Huang, Lin, & Yang, 2008), and the proportion of female smokers in the overall female population in Taiwan is low (4.1%; Health Promotion Administration, 2013).
Materials and Methods
Participants and Procedures
We advertised this cross-sectional study to all classes in both Chang Jung Christian University (CJCU) and Central Taiwan University of Science and Technology (CTUST). Volunteers contacted us, and we screened the convenience samples for eligibility at the health centers of the two universities, ultimately enrolling 481 male university students aged 20–25 years. All study subjects gave written, informed consent prior to data collection, and the review boards of CJCU (99004) and CTUST (99016) approved the study. Participants completed a self-report questionnaire for demographic data and smoking background and status as well as the Chinese version of the Fagerstrom Test for Nicotine Dependence (FTND; comprises 6 items, with higher scores indicating greater nicotine dependence; C. L. Huang, Lin, & Wang, 2006).
Current smoking was defined as having smoked at least one cigarette on 1 or more days in the past 30 days. We divided current smokers into two groups according to the intensity of their cigarette smoking: light smokers (<5 cigarettes per day) and heavier smokers (>6 cigarettes per day; Nasim, Khader, Blank, Cobb, & Eissenberg, 2012). Never smokers were defined as persons who had never smoked, even a few puffs, in their lifetime. Exclusion criteria included (1) a history of diagnosed mental health disease or cancer, (2) alcohol or drug abuse (having a history or current status), (3) a severe communication problem, or (4) a terminal illness. When participants completed the survey, trained research assistants who were licensed nurses or licensed medical technologists obtained blood and urine samples at a university health center.
Determination of DRD2 and MAOA Polymorphisms
For determination of genes, we isolated total genomic DNA from the blood cells using a blood DNA isolation kit (Favorgen, Pingtung, Taiwan). This procedure required 5 ml of whole blood with EDTA as anticoagulant. The DRD2 polymorphisms, DRD2 rs1800497 and DRD2 rs1079597, were determined using polymerase chain reaction (PCR)-restriction fragment length polymorphism (PCR-RFLP) methods, as reported previously (Spitz et al., 1998), with minor modification for DRD2 rs1800497. For DRD2 rs1800497, we designed forward primer 5’-ACC TTC CTG AGT GTC ATC AACC-3’ and reverse primer 5’-CTA AAT TTC CAT CTC GGC TCC T-3’ to amplify a 288-base pair (bp) fragment. Utilizing Taq α1 as the restriction enzyme, we observed one (288-bp), two (163 and 125-bp), and three (288, 163, and 125-bp) fragments for the CC, CT, and TT genotypes, respectively. For DRD2 rs1079597, forward primer 5’-GAT ACC CAC TTC AGG AAG TC-3’ and reverse primer 5’-GAT GTG TAG GAA TTA GCC AGG-3’ were used to amplify a 459-bp fragment. Using Taq α1 as the restriction enzyme, we observed one (459-bp), two (267 and 192-bp), and three (459, 267, and 192-bp) fragments for the AA, AG, and GG genotypes, respectively.
The MAOA polymorphisms, MAOA rs309850 and MAOA rs1137070, were determined by the methods of PCR and PCR-RFLP, respectively (Jin et al., 2006). For MAOA rs309850 polymorphisms, the forward primer 5’-ACA GCC TGA CCG TGG AGA AG-3’ and reverse primer 5’-GAA CGG ACG CTC CAT TCG GA-3’ were utilized to yield 321 bp and 351 bp fragments corresponding to the 3- and 4-repeat alleles, respectively. For MAOA rs1137070 polymorphisms, we used forward primer 5’-GGA GAA CGA GCA GCT AGG GAG GTA A-3’ and reverse primer 5’-GAT CTT CAG CAG GCC AGA AAC AGA G-3’. Restriction enzyme EcoRV yielded a 557-bp band for subjects carrying 1460 C/O and two fragments (260-bp and 297-bp) for those carrying 1460 T/O, respectively. DRD2 rs1800497 CC, CT, and TT; DRD2 rs1079597 AA, AG, and GG; MAOA rs309850 3- and 4-repeat and MAOA rs1137070 1460 C/O and 1460 T/O, which had been found in Taiwanese by DNA sequencing method, were run as positive controls in each performance of the genotyping assays. We did not find any error in the approximate 30 performances for each genotype.
Determination of Urinary Nicotine and Cotinine
Urine samples from smokers were obtained around 1 hr after they had smoked the first cigarette of the morning. We measured urinary nicotine and cotinine of smokers using a gas chromatography-mass spectrometry method (GC/MS), as previously described (Dempsey et al., 2004), with the following difference: GC/MS analyses were performed on a ThermoElectron DSQII quadrupole mass spectrometer connected directly to a ThermoElectron Focus gas chromatograph and to an autosampler AS 3000 (Thermo Electron Corporation, Dreieich, Germany). Urine samples were obtained from smokers and were stored at −20°C until analysis. We divided nicotine and cotinine concentrations in the urine of each subject by the number of cigarettes smoked daily as the adjusted nicotine and cotinine concentrations. The tobacco currently used in Taiwanese products contains on average .57–.64 mg of nicotine per cigarette (Health Promotion Administration, 2013).
Statistical Analysis
Calculations were performed in six models, respectively, DRD2 rs1800497, DRD2 rs1079597, MAOA rs309850, MAOA rs1137070, and their interactions. All subjects were divided into two genotypes in Model 2 (MAOA rs309850 3-repeat and MAOA rs309850 4-repeat; and MAOA rs1137070 1460 C/O and MAOA rs1137070 1460 T/O, respectively) and divided into three genotypes in other models (e.g., DRD2 rs1079597 AA//MAOA rs309850 3-repeat, DRD2 rs1079597 AG//MAOA rs309850 3-repeat, and DRD2 rs1079597 GG//MAOA rs309850 3-repeat). Mantel-Haenszel χ2 test was used to calculate the odds ratios (ORs) and 95% CIs of the ORs to evaluate the risk of genetic variants for smoking status and intensity of smoking, by assigning OR as 1.0 for the subjects carrying the appropriate genotype. We applied Student’s t-test, analysis of variance (ANOVA), or the Mann-Whitney U test, as appropriate, to compare quantitative data. We defined a p value of <.05 as statistically significant for each analysis. When the result of an ANOVA was statistically significant, we followed multiple comparisons with the application of Scheffé post hoc test. All data were analyzed using SPSS (version 18.0 software for Windows, SPSS Inc., Chicago, IL).
Results
Participants included 261 current smokers and 220 never smokers. Demographic data for the heavier smokers, light smokers, and never smokers are shown in online supplementary Table 1. Mean age was not statistically different among them (p = .155), while FTND score and adjusted urinary nicotine and cotinine concentrations were significantly different between heavier smokers and light smokers.
ORs for Smoking Status
The online supplementary Table 2 shows the distribution of genotypes for DRD2 rs1800497, DRD2 rs1079597, MAOA rs309850, and MAOA rs1137070. The distributions of genotypes for DRD2 rs1800497 and DRD2 rs1079597 were in line with the Hardy-Weinberg principle. There were no significantly different genotypes between current smokers and never smokers.
With never smokers as the reference group, the 22 ORs for the association with smoking status were not statistically significant (p = .082–.873, data shown in online supplementary Table 3). We divided the 261 current smokers into heavier smokers (N = 165) and light smokers (N = 96). Among the 173 subjects carrying MAOA rs309850 3-repeat, there were 106 heavier smokers and 67 light smokers. The OR of heavier smoking (light smokers as the reference group) was 2.67 times higher in the subjects with DRD2 rs1079597 GG//MAOA rs309850 3-repeat than in those with DRD2 rs1079597 AA//MAOA rs309850 3-repeat (95% CI = [1.08, −6.59], p = .031), as shown in Table 1. The other 21 ORs were not statistically significant (p = .098–1.000, data shown in online supplementary Table 4).
Adjusted Odds Ratios (95% Confidence Intervals) for Smoking Status and Heavier Smoking in Different Genotypes.
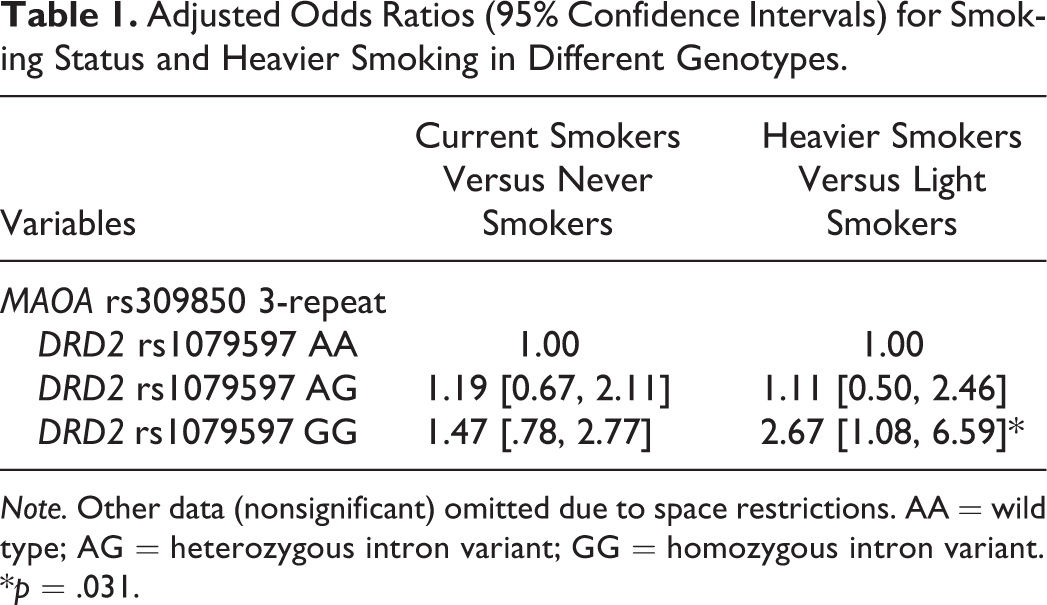
Note. Other data (nonsignificant) omitted due to space restrictions. AA = wild type; AG = heterozygous intron variant; GG = homozygous intron variant.
*p = .031.
FTND
We have FTND results for 182 of the 261 current smokers because of missing data. There were 12 comparison results. Table 2 shows that, for the 119 subjects with MAOA rs309850 3-repeat, FTND was significantly different among the smokers carrying DRD2 rs1079597 AA//MAOA rs309850 3-repeat, DRD2 rs1079597 AG//MAOA rs309850 3-repeat, and DRD2 rs1079597 GG//MAOA rs309850 3-repeat [F (2, 116) = 3.721, p = .027]. Multiple comparisons with the Scheffé post hoc test indicated that FTND were higher in those with DRD2 rs1079597 GG//MAOA rs309850 3-repeat than in those with DRD2 rs1079597 AA//MAOA rs309850 3-repeat, 4.26 vs. 2.83, F (2, 116) = 6.946, p = .033 for F (2, 116). The other 11 comparisons of FTND were not significantly different (p = .101–.976, data shown in online supplementary Table 5).
Comparisons of FTND Results Among Different Genotypes.
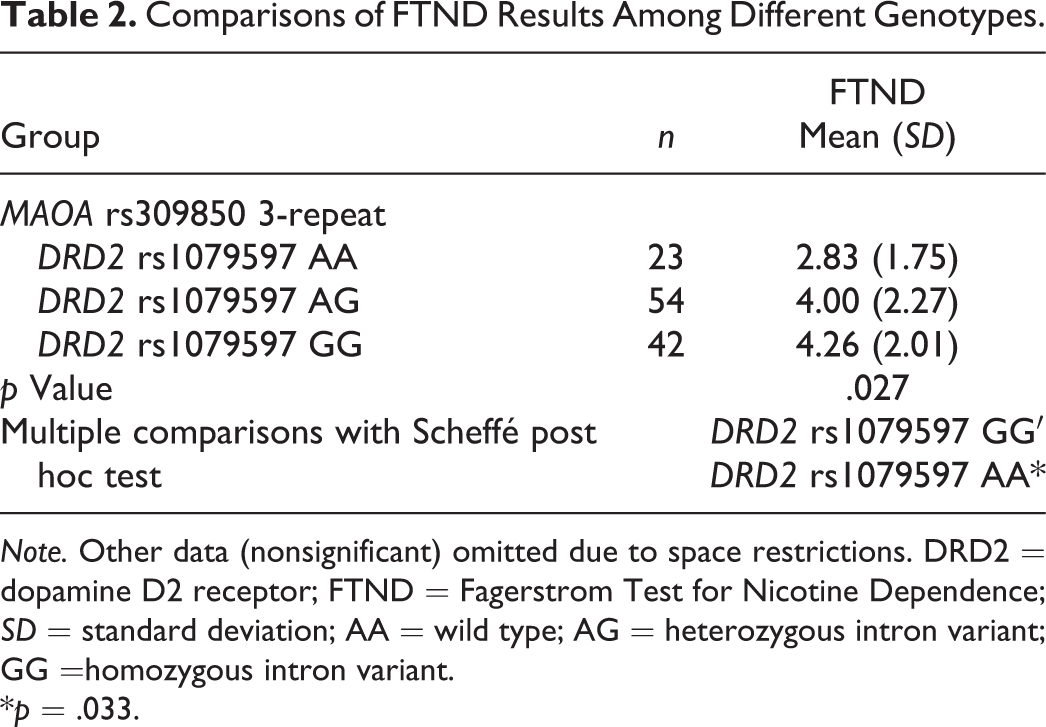
Note. Other data (nonsignificant) omitted due to space restrictions. DRD2 = dopamine D2 receptor; FTND = Fagerstrom Test for Nicotine Dependence; SD = standard deviation; AA = wild type; AG = heterozygous intron variant; GG =homozygous intron variant.
*p = .033.
Adjusted Urinary Nicotine and Cotinine Concentrations
Of the 261 current smokers, 122 provided urine samples. Since all values of both adjusted urinary nicotine and cotinine concentrations were not normally distributed in each group, we used the Mann-Whitney U-test to perform the comparisons. For both adjusted urinary nicotine and cotinine, there were 12 comparison results. As shown in Table 3, among smokers carrying MAOA rs309850 3-repeat polymorphism, adjusted nicotine concentration was significantly different between subjects with DRD2 rs1800497 CC and DRD2 rs1800497 CT (z = 2.362, p = .018) and between those with DRD2 rs1800497 CT and DRD2 rs1800497 TT (z = 2.203, p =.028). Among the smokers with MAOA rs1137070 1460 T/O, adjusted nicotine concentration was significantly different between subjects with DRD2 rs1800497 CC and DRD2 rs1800497 CT (z = 2.002, p = .045) and between those with DRD2 rs1800497 CT and DRD2 rs1800497 TT (z = 2.565, p = .010). For adjusted cotinine concentration, there were statistically significant differences between genotypes of DRD2 rs1079597 AA and DRD2 rs1079597 GG (z = 2.301, p = .021), between genotypes of DRD2 rs1079597 AA//MAOA rs309850 3-repeat and DRD2 rs1079597 GG//MAOA rs309850 3-repeat (z = 2.002, p = .045), between genotypes of DRD2 rs1800497 CT//MAOA rs1137070 1460 T/O and DRD2 rs1800497 TT//MAOA rs1137070 1460 T/O (z = 2.810, p = .005), between genotypes of DRD2 rs1079597 AG//MAOA rs1137070 1460 T/O and DRD2 rs1079597 GG//MAOA rs1137070 1460 T/O (z = 2.015, p = .044), and between genotypes of DRD2 rs1079597 AA//MAOA rs1137070 1460 T/O and DRD2 rs1079597 GG//MAOA rs1137070 1460 T/O (z = 2.726, p = .006).
Comparisons of Urinary Nicotine and Cotinine Concentrations Among Different Genotypes.
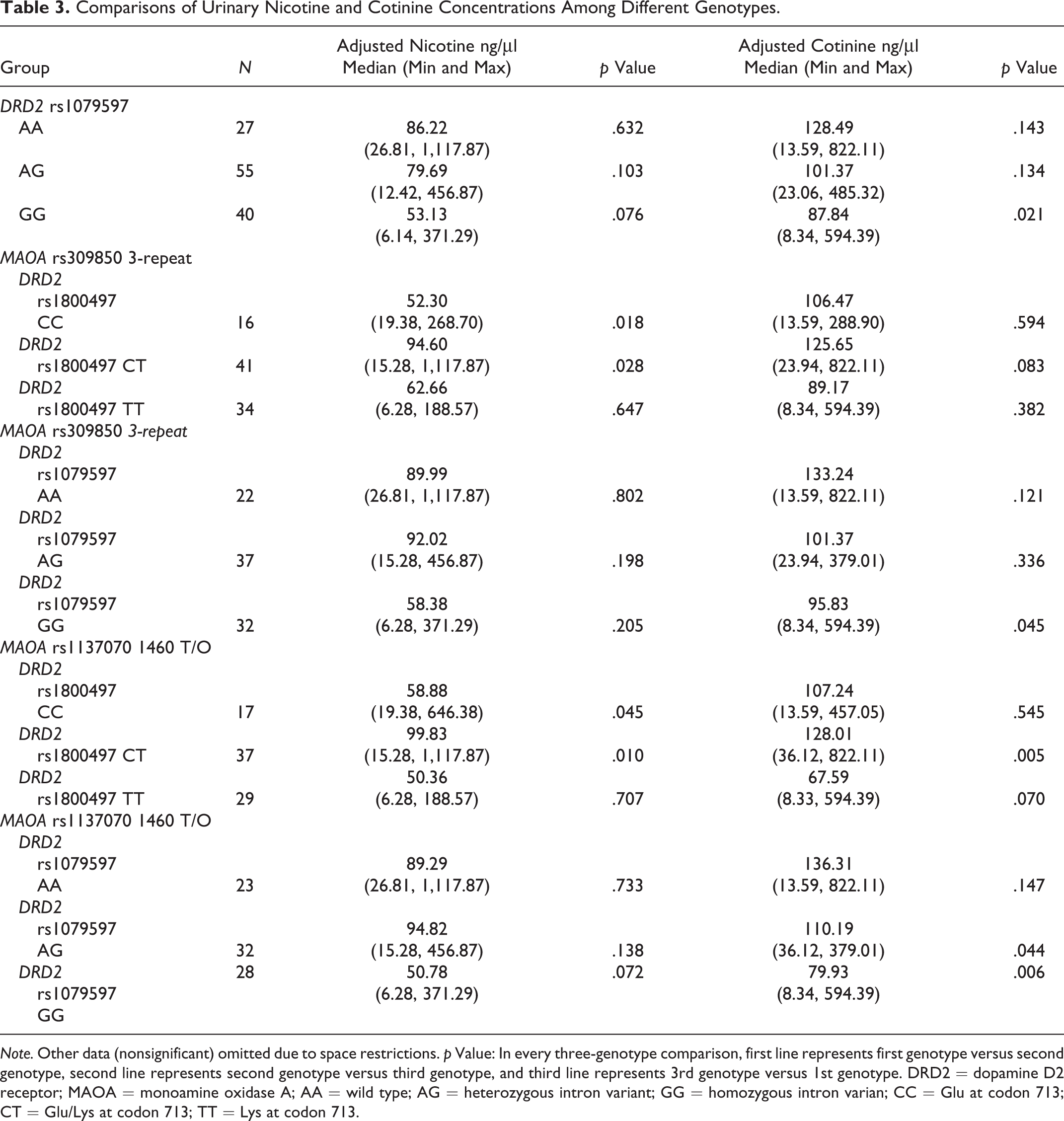
Note. Other data (nonsignificant) omitted due to space restrictions. p Value: In every three-genotype comparison, first line represents first genotype versus second genotype, second line represents second genotype versus third genotype, and third line represents 3rd genotype versus 1st genotype. DRD2 = dopamine D2 receptor; MAOA = monoamine oxidase A; AA = wild type; AG = heterozygous intron variant; GG = homozygous intron varian; CC = Glu at codon 713; CT = Glu/Lys at codon 713; TT = Lys at codon 713.
Discussion
For this study, we enrolled adult male university students who did not abuse alcohol or drugs. Therefore, the confounding factors on smoking behavior, such as age, educational status, alcohol interaction and drug interaction, were absent. Our results demonstrate that polymorphism of a single gene for DRD2 rs1800497, DRD2 rs1079597, MAOA rs309850, and MAOA rs1137070 is not related to smoking status in Taiwanese men, which is consistent with findings from previous studies for other ethnic groups for DRD2 rs1800497 (Hiemstra, Kleinjan, van Schayck, Engels, & Otten, 2014; Ohmoto et al., 2013), DRD2 rs1079597 (Costa-Mallen et al., 2005; Suriyaprom et al., 2013), MAOA rs309850 (S. Huang et al., 2005), and MAOA rs1137070 (Tochigi et al., 2007). However, we found that, among smokers carrying MAOA rs309850 3-repeat, DRD2 rs1079597 GG genotype is a higher risk factor for smoking intensity than DRD2 rs1079597 AA genotype. This finding supports the suggestion that the association between smoking risk and DRD2 gene, if it exists, is related to ethnicity: rs1800497 C (or rs1079597 A) polymorphism seems to be specific to Whites (Ohmoto et al., 2013; Spitz et al., 1998) and Mexican Americans (Robinson et al., 2007), while rs1800497 T polymorphism seems to be specific to Asians (Ohmoto et al., 2013), Poles (Sieminska, Buczkowski, Jassem, Niedoszytko, & Tkacz, 2009), and Russians (Gordiev et al., 2013).
Gordiev et al. (2013) reported that DRD2 rs1079597 AA carriers as well as DRD2 rs1800497 CC carriers had a lower density of DRD2 receptors. Sieminska, Buczkowski, Jassem, Niedoszytko, and Tkacz (2009) hypothesized that lower dopamine receptor density was the cause of higher dopamine availability. Thus, subjects carrying DRD2 rs1079597 GG may have more need for repeated dopaminergic stimuli than those carrying DRD2 rs1079597 AA.
Monoamine oxidase catalyzes the oxidative deamination of biogenic amines such as dopamine, serotonin, noradrenaline, and histamine (Jin et al., 2006). Jin et al. (2006) estimated that MAOA accounts for 70% of neuronal MAO. Therefore, MAOA seems to be more important for smoking status than MAOB, which is why we investigated the MAOA gene rather than the MAOB gene. We hypothesized that individuals with lower MAOA enzyme activity may have enhanced dopamine release (Jin et al., 2006). Thus, for smokers carrying DRD2 rs1079597 GG polymorphism combined with MAOA rs309850 3-repeat (low activity) polymorphism, more nicotine from smoking may be needed in order to release enough dopamine in the brain to produce feelings of pleasure and reward. This might explain, at least partially, why smokers with DRD2 rs1079597 GG//MAOA rs309850 3-repeat polymorphism have an increased risk of nicotine dependence and heavier smoking than those with DRD2 rs1079597 AA//MAOA rs309850 3-repeat polymorphism, as we found in the present study.
Urinary cotinine concentration is a reliable, easy-to-use marker for plasma levels of cotinine and the sum of nicotine metabolites in smokers (Nagano et al., 2010). In the present study, the significant difference in adjusted urinary cotinine concentration between heavier smokers and light smokers (median values: 84.21 ng/μl vs. 131.69 ng/μl, respectively) may indicate that heavier smokers consume cotinine and the sum of nicotine metabolites more rapidly than light smokers. In the smokers in the present study, urinary cotinine concentration after smoking one cigarette differed significantly between DRD2 rs1079597 GG//MAOA rs309850 3-repeat carriers (median value: 95.83 ng/μl) and DRD2 rs1079597 AA//MAOA rs309850 3-repeat carriers (133.24 ng/μl). Cotinine and the sum of nicotine metabolites may be consumed more rapidly in smokers carrying DRD2 rs1079597 GG//MAOA rs309850 3-repeat than in those possessing DRD2 rs1079597 AA//MAOA rs309850 3-repeat. These findings are consistent with the increased risk of nicotine dependence that we observed in smokers carrying DRD2 rs1079597 GG//MAOA rs309850 3-repeat in the present study.
In the present study, adjusted nicotine and cotinine concentrations differed significantly different between two particular genotypes when DRD2 rs1800497 or MAOA rs1137070 1460 T/O was considered. However, those differences were not associated with smoking status. There were statistically significant correlations between DRD2 rs1800497 and DRD2 rs1079597 alleles (r = .76 for current smokers and r = .83 for never smokers; p < .001, data not shown). However, interactions of DRD2 rs1800497 and MAOA genes did not affect smoking status. Spitz et al. (1998) reported that the DRD2 rs1079597 allele seems more predictive than the DRD2 rs1800497 allele for smoking status, and both Costa-Mallen, Costa, and Checkoway (2005) and Robinson et al. (2007) found that polymorphism of DRD2 rs1079597 was a better marker of smoking than the DRD2 rs1800497 polymorphism. Those results may explain, at least partially, the differences we report here between the effects of DRD2 rs1800497 and DRD2 rs1079597 polymorphisms on smoking status. The stronger relationship between smoking status and DRD2 rs1079597 polymorphism also may explain why, among Taiwanese individuals, interaction between DRD2 rs1800497 and MAOA genes is associated with alcoholism (Hu et al., 2013), while interaction between DRD2 rs1079597 and MAOA genes is related to smoking status.
Our study had several limitations. First, study participation was restricted to young male Taiwanese smokers. Because the percentage of female smokers in the Taiwanese population is low, at only 4.4%, female smokers are difficult to recruit as participants. These results may thus not be generalizable to female smokers or to other ethnic groups. Second, the associations we examined involved a small set of biological candidate genes; this was not a genome-wide association study. We based our approach on the premise that the susceptibility alleles for alcohol dependence would be worthy of study. Third, sample size was a potential limitation. Genetic variants usually have small effects compared to nongenetic risk factors. Our sample size could have been underpowered for detecting the effects of the interaction of DRD2 and MAOA on smoking status and cigarette dependence. When we conducted the study, no comparable sample was available. In spite of these limitations, the approach we employed illustrates the effects of the interaction of DRD2 and MAOA, thereby constructing a more complete picture of the genetic architecture of the complex status of cigarette smoking and cigarette dependence.
In conclusion, although not all genotypes of DRD2 rs1800497, DRD2 rs1079597, MAOA rs309850, and MAOA rs1137070 are involved in the development of smoking, the interaction of DRD2 rs1079597 and MAOA rs309850 3-repeat affects smoking intensity in young Taiwanese men. Such understanding provides a useful foundation for nurse professionals to conduct further biological studies of smoking and for nurse practitioners to educate their patients about the genetic risks for developing cigarette dependence. Although smoking and cigarette dependence are complex health problems, understanding the effects of the interaction between DRD2 and MAOA genes on smoking is an important step toward increasing our ability to assess individual and population genetic factors and design appropriate interventions to reduce cigarette smoking.
Footnotes
Author Contribution
CLH contributed to acquisition, analysis, and interpretation of data; drafted the manuscript; gave final approval; and agrees to be accountable for all aspects of work ensuring integrity and accuracy. WCO and PLC contributed to acquisition of data, drafted the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy. CNL, MCC, and CCL contributed to interpretation of data, drafted the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy. YCC and MHL Lin contributed to analysis of data, drafted the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy. CSH Huang contributed to conception and design. drafted the manuscript, critically revised the manuscript, gave final approval, and agrees to be accountable for all aspects of work ensuring integrity and accuracy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was financially supported by National Science Council, Taiwan (contract number: NSC–100–2320–B–371–001), Changhua Christian Hospital, Taiwan (contract number: CCH–ICO10008), and Central Taiwan University of Science and Technology (contract number: CTU102–P–21).