
Abstract
Schizophrenia is a highly heritable disorder, the genetic etiology of which has been well established. Yet despite significant advances in genetics research, the pathophysiological mechanisms of this disorder largely remain unknown. This gap has been attributed to the complexity of the polygenic disorder, which has a heterogeneous clinical profile. Examining the genetic basis of schizophrenia subphenotypes, such as those based on particular symptoms, is thus a useful strategy for decoding the underlying mechanisms. This review of literature examines the recent advances (from 2011) in genetic exploration of positive and negative symptoms in schizophrenia. We searched electronic databases PubMed, Web of Science, and Cumulative Index to Nursing and Allied Health Literature using key words schizophrenia, symptoms, positive symptoms, negative symptoms, cognition, genetics, genes, genetic predisposition, and genotype in various combinations. We identified 115 articles, which are included in the review. Evidence from these studies, most of which are genetic association studies, identifies shared and unique gene associations for the symptom domains. Genes associated with neurotransmitter systems and neuronal development/maintenance primarily constitute the shared associations. Needed are studies that examine the genetic basis of specific symptoms within the broader domains in addition to functional mechanisms. Such investigations are critical to developing precision treatment and care for individuals afflicted with schizophrenia.
Schizophrenia is a brain disorder that affects roughly 1% of the general population (McGrath, Saha, Chant, & Welham, 2008). It is characterized by severe and debilitating symptoms broadly classified into three domains: positive, negative, and cognitive. Positive symptoms are an excess of “normal” functions, can be quite dramatic, and arise suddenly during a psychotic break. Hallucinations and delusions are the most common of the positive symptoms, but aberrant processes in thought, speech, and behavior may also occur. Negative symptoms are a deficit of “normal” functions and include problems with communication, specifically fluency of speech, problems with thought production, reduced emotional expression (affective blunting), social withdrawal or decreased social drive (asociality), difficulty experiencing pleasure (anhedonia), and restricted desire or motivation (avolition). Cognitive symptoms of schizophrenia generally include deficits in neurocognition (such as impaired attention, memory, information processing, learning, and executive functioning) and social cognition (such as impaired social and emotional processing and attributional bias).
The heritability of schizophrenia (h 2 ∼ 80%) has been definitively established through several family and twin studies (Sullivan, Kendler, & Neale, 2003). Recent advances in research using genomic technologies such as whole-genome sequencing have also confirmed that genetics is a major cause of the illness and that it has a polygenic architecture with risk alleles in several genes contributing to schizophrenia susceptibility. The two leading hypotheses (Escudero & Johnstone, 2014) regarding the etiopathogenesis of schizophrenia are (1) common variant–common disease model and (2) common disease–rare variant model. In support of the first hypothesis, in the largest study of its kind, researchers found 128 linkage disequilibrium independent genome-wide associations (with odds ratio < 1.2) associated with schizophrenia (Schizophrenia Working Group of the Psychiatric Genomics Consortium, 2014), while in support of the second, researchers have also found strong associations between rare variations such as copy number variants (CNVs; 11 CNVs with genome-wide significance levels, odds ratio of 2–60) and schizophrenia (Rees, O’Donovan, & Owen, 2015). But the pathogenic mechanisms of these genetic associations remain largely unknown. This gap in knowledge has been attributed to several factors including the complexity of schizophrenia, which has both genetic and environmental contributors. Also, in genetic association studies, researchers have examined associations with schizophrenia diagnosis as the primary phenotype. Many of the associations as such could be due to an indirect relationship. For example, studies have shown a strong association of schizophrenia risk with the CHRNA5-A3-A4 gene cluster (Freedman et al., 2001), which is also strongly associated with heavy smoking (Ware, van den Bree, & Munafò, 2011). Smoking is highly prevalent in schizophrenia (∼80%; Hughes, Hatsukami, Mitchell, & Dahlgren, 1986), and the demonstrated associations could either be a reflection of this confound (Flint & Munafò, 2014) or the shared effect of the allelic variant (Hong et al., 2011).
Another factor adding to the challenges of delineating the functional mechanisms involved in the pathogenesis of schizophrenia is that the disease is both clinically and genetically heterogeneous (Arnedo et al., 2015). Authors have suggested addressing these challenges by refining the phenotype of schizophrenia. For example, researchers could identify intermediate phenotypes (i.e., heritable characteristics that are on the disease pathway and are more likely to be closely related to the genotype than the phenotype; Rasetti & Weinberger, 2011; e.g., brain activations identified in neuroimaging) or characterize subphenotypes by severity of illness, age of onset, or symptoms (Morris et al., 2010).
Symptoms of schizophrenia make good subphenotypes for several reasons: (1) evidence suggests there is a genetic basis for differences in clinical profiles of schizophrenia (Arnedo et al., 2015); (2) evidence also supports the heritability of symptoms (Kendler et al., 1997; Malaspina et al., 2000); (3) symptoms have been defined and operationalized well; and (4) validated, reliable measures (e.g., the Positive and Negative Syndrome Scale) are available to assess symptoms. In this review of literature, we summarize recent advances in the study of the genetic basis of positive and negative symptoms in schizophrenia. Cognition/cognitive symptoms in schizophrenia are complex, involve an array of cognitive functions, and are the most studied in schizophrenia; hence, they are beyond the scope of this review.
Method
To capture all potentially relevant articles, we searched PubMed, Web of Science, and the Cumulative Index to Nursing and Allied Health Literature using key words schizophrenia, symptoms, positive symptoms, negative symptoms, cognition, genetics, genes, genetic predisposition, and genotype in various combinations and limited the search to English-language articles. Since the field of psychiatric genetics has grown exponentially in recent years, we also limited our search to the last 5 years (since 2011). We reviewed titles and abstracts of retrieved articles to identify articles that focused on the genetic basis of positive and negative symptoms in schizophrenia as a primary or secondary aim. We excluded foreign-language full-text articles and articles that focused on symptoms other than positive or negative symptoms from the review. We found 115 full-text articles that met these criteria and included them in this review. Figure 1 summarizes the review process in a Preferred Reporting Items for Systematic Reviews and Meta-Analyses chart. We extracted evidence from these articles for a genetic basis for positive and negative symptoms in schizophrenia and present it subsequently. We also present a conceptual model for the genetic contributions to these symptom domains based on our synthesis of this evidence.
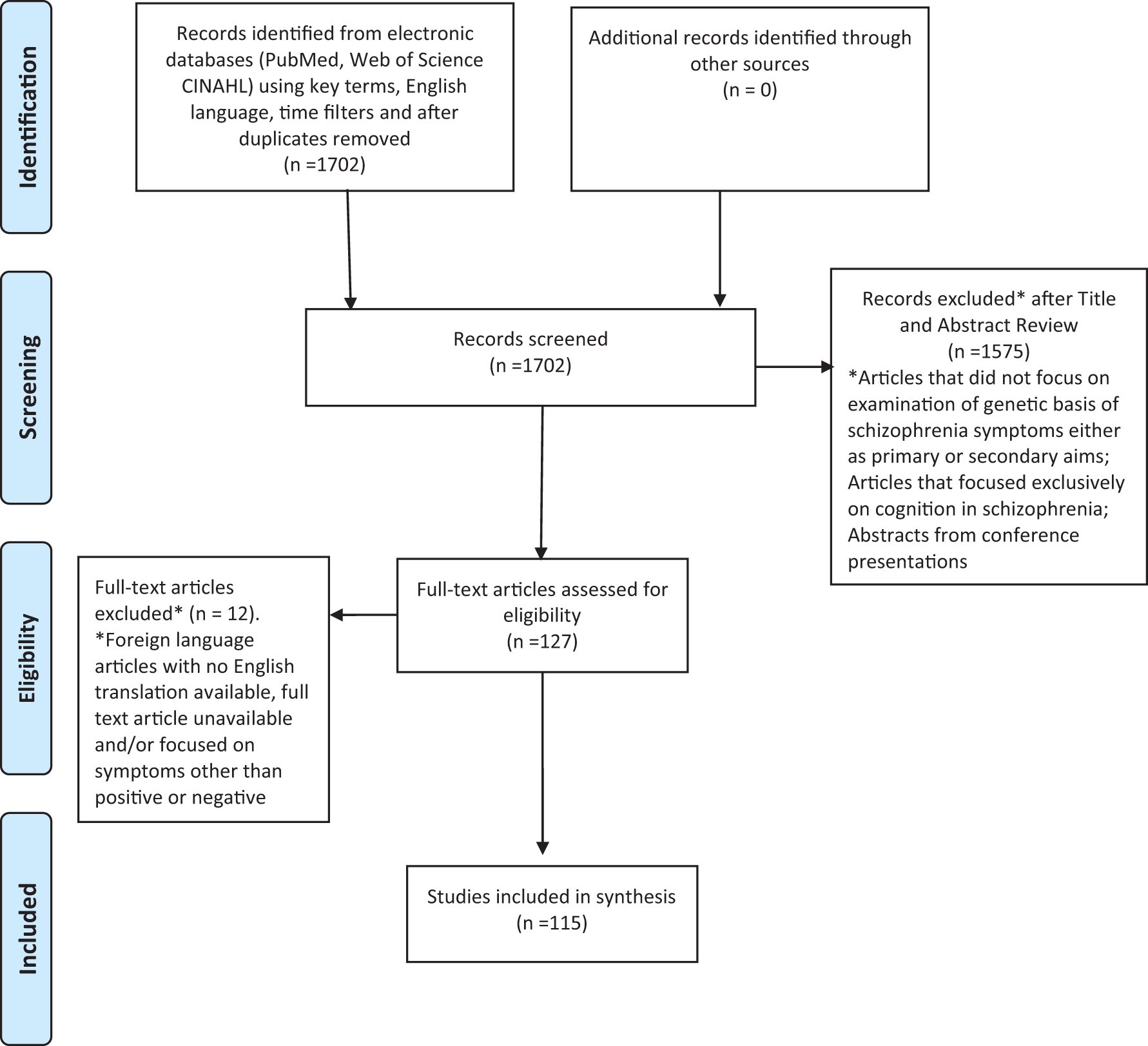
Search strategy and article selection process. Preferred Reporting Items for Systematic Reviews and Meta-Analyses flow diagram from http://www.prisma-statement.org/PRISMAStatement/FlowDiagram.aspx.
Results
Our review revealed that many of the known genetic contributions and potential physiological mechanisms for the positive and negative symptoms of schizophrenia are overlapping, while a few are unique to negative or positive symptoms. We illustrate this overall finding in Figure 2, a model of the genetic associations with positive and negative symptoms of schizophrenia.
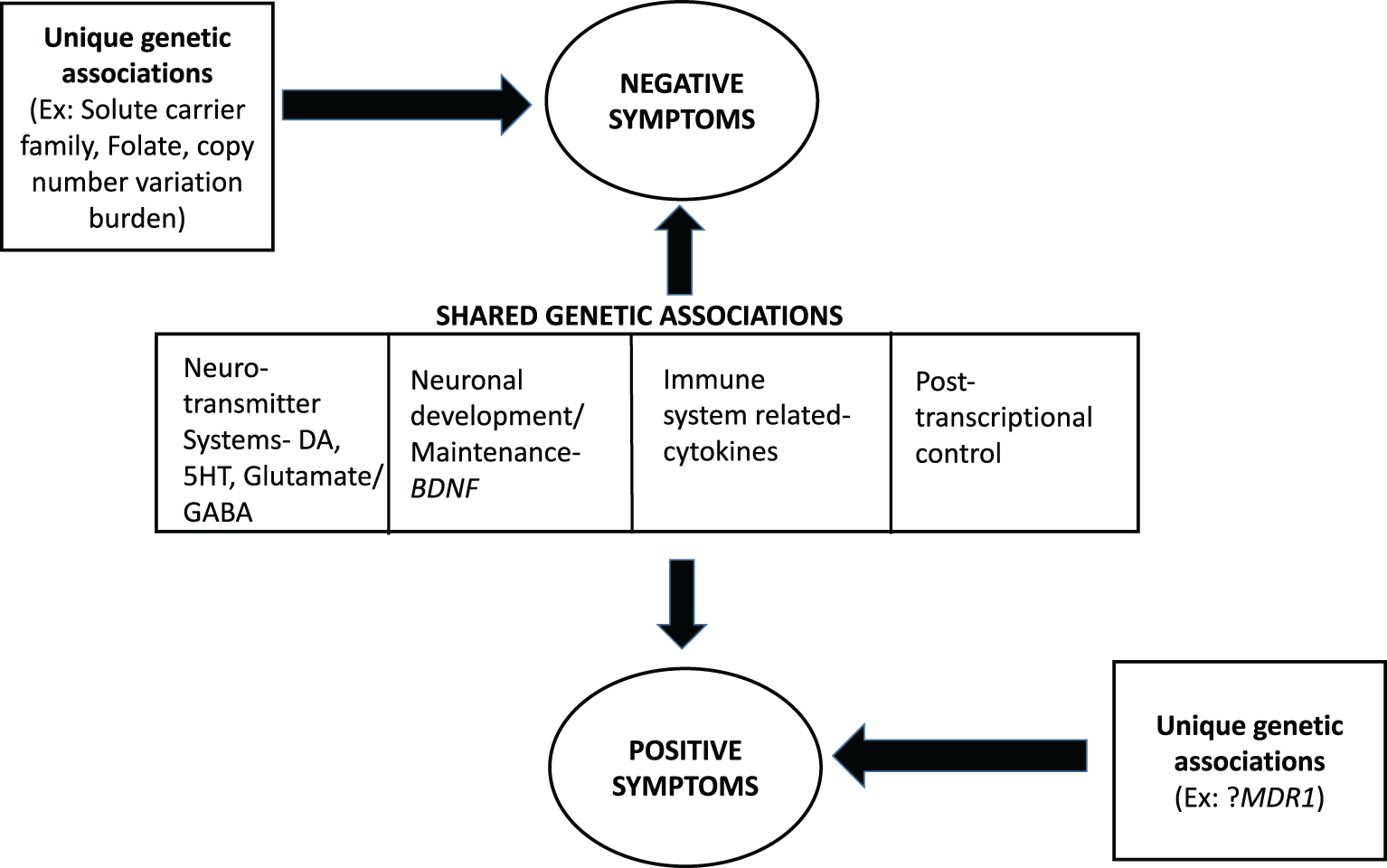
A model for the genetic basis of positive and negative symptoms in schizophrenia. Evidence suggests shared and unique pathways for the two domains with genes involving the neurotransmitter systems: dopamine (DA), which appears to play a significant role; glutamate/gamma amino butyric acid; and serotonin (5HT) signaling. In addition, several other genes (primarily brain-derived neurotrophic factor) influence both domains. There is evidence for unique genetic influences for negative symptoms, but such influences are as yet unclear for positive symptoms.
Genetic Contributions Common to Positive and Negative Symptoms in Schizophrenia
Dopaminergic signaling
Disruption in the dopaminergic system is the oldest and, until recently, the most strongly held hypothesis regarding the pathogenesis of schizophrenia, especially of positive symptoms. Evidence supporting this hypothesis comes primarily from pharmacological studies of neurotransmitter systems, particularly studies on D2 receptor blocking antipsychotic medications. These medications, though very effective for managing positive symptoms, have little to no effect on other symptom categories such as negative and cognitive. Dopamine is a monoamine catecholamine neurotransmitter that acts through its receptors in target cells (Missale, Nash, Robinson, Jaber, & Caron, 1998). There are two classes of dopamine receptors, which are important elements of the dopaminergic system, D1 and D2. D1 receptors increase intracellular cyclic adenosine monophosphate (cAMP) when activated, whereas D2 receptors inhibit cAMP when activated (Missale et al., 1998). cAMP is important for intracellular signal transduction. Dopamine reuptake at the synapse, which is critical for maintaining the presynaptic levels of dopamine, is dependent on dopamine transporters (DATs). Dopamine is degraded by the enzyme catechol-O-methyltransferase (COMT). There are five dopaminergic pathways in the brain: (1) mesolimbic, which arises from the ventral tegmental area (VTA) of the mid brain, projects to the limbic system, and is believed to be involved in positive symptoms; (2) mesocortical, which arises from the VTA, projects to the cortical regions, and is believed to be associated with negative symptoms; (3) nigrostriatal, which extends from the substantia nigra, projects to the striatum, and is involved with movement; (4) tuberoinfundibular, which extends from the hypothalamus to the pituitary gland; and (5) dopamine pathways that arise from several regions and project to the thalamus, with unknown functions (Stahl, 2013).
Dopamine receptors and transporters
Dopamine receptor genes implicated in positive symptoms of schizophrenia include DRD2 and DRD3. Dopamine D2 receptor (DRD2) gene, located on chromosome 11.q22-q23, encodes for the dopamine 2 receptor, which belongs to the G protein-coupled receptor superfamily and is the primary binding target for all currently available antipsychotic medications, both typical and atypical (Kapur & Mamo, 2003). Gene association, pharmacogenetic, and gene expression studies provide converging evidence for an association of DRD2 with positive symptoms. The top-hit single-nucleotide polymorphism (SNP) for this loci, as reported by the Psychiatric Genomics Consortium (Schizophrenia Working Group of the Psychiatric Genomics Consortium, 2014), is rs2514218 which is 47 kb upstream from DRD2 and is associated with risk of schizophrenia. J. P. Zhang et al. (2015) found that homozygotes for the risk C allele at this locus had greater positive symptoms at baseline than T-allele carriers, but after 12 weeks of treatment with aripiprazole or risperidone (both second-generation antipsychotics that reduce striatal dopamine signaling), they had greater improvement in positive symptoms than T-allele carriers. This finding strengthens the hypothesis that positive symptoms are associated with excessive dopaminergic action in the mesolimbic pathway (Davis & Kahn, 1991). Kurian et al. (2011) found that DRD2 expression was decreased in delusional states, and L. Liu et al. (2013) replicated this finding in the Han Chinese population, observing that DRD2 was associated with positive symptoms in acute schizophrenia but not in chronic schizophrenia. In mouse models, atypical antipsychotic medications (irrespective of their type) with a significant interaction with DRD2 elicited temporal- and region-specific gene expression patterns in the nucleus accumbens and striatum, which are involved in dopaminergic signaling (Sakuma et al., 2015).
Evidence for the association of DRD2 variants with negative symptoms comes from genetic association studies (Chien et al., 2013; Zahari, Teh, Ismail, & Razali, 2011), pharmacogenetic studies (S. G. Kang et al., 2015), proteomics and transcriptomics (Pinacho et al., 2013), and animal models (Simpson et al., 2011; Ward et al., 2012). Motivational deficits appear to be the primary negative symptom that is mediated via the striatal dopaminergic system (Ward et al., 2012). Cerebellar dopamine receptor D2, in association with NMDA receptor subunit 1 genes, is associated with negative symptoms, though the functional mechanism for this is unknown (Pinacho et al., 2013).
The dopamine D3 receptor (DRD3) gene encodes for the D3 receptors. Vehof et al. (2012) found that the C allele of the Ser9Gly polymorphism of DRD3, which is an amino acid substitution in the N-terminal extracellular region of the D3 receptor, is associated with improvement in positive symptoms. DAT1, the dopamine transporter gene, encodes for DATs and is expressed primarily in the midbrain dopaminergic neurons projecting to the substantia nigra and ventral tegmental regions (Bannon, Michelhaugh, Wang, & Sacchetti, 2001). Research has shown that the variant rs2455391 of DAT1 is associated with positive symptoms (Zheng, Shen, & Xu, 2012).
Dopamine catabolism
The COMT gene encodes for the COMT enzyme, which is critical in the degradation of dopamine. The variant Val158Met (rs4680) causes differences in enzymatic activity and subsequent variation in the levels of dopamine in the prefrontal cortex (Lachman et al., 1996). COMT Val carriers have more positive symptoms than individuals with schizophrenia who do not carry the variant (Ira et al., 2014). Although there have been conflicting findings in pharmacogenetic studies (Vehof et al., 2012), in a recent meta-analysis, Huang et al. (2016) confirmed that COMT is associated with changes in positive symptoms during treatment with atypical antipsychotic medications. In addition, research has shown that the relationship between environmental factors such as low maternal care and a high-stress environment and positive symptoms is moderated by the Val158Met genotype (Ira et al., 2014).
Researchers have also studied COMT extensively in relation to negative symptoms, and a number of variants have shown strong associations (see Table 1) in several ethnic populations including Caucasian (Roffman et al., 2013), Chinese (C. Y. Chen et al., 2016; W. J. Li et al., 2012), Korean (W. S. Kang et al., 2012), Spanish (Pelayo-Teran et al., 2011), Italian (Bosia et al., 2015), Polish Caucasian (Pelka-Wysiecka et al., 2013), and Xhosa African (Wright et al., 2012). The COMT enzyme is hypothesized to exert its effect on negative symptoms primarily through the mesocortical dopaminergic pathways (Winterer & Weinberger, 2004).
Reviewed Studies That Contribute Evidence for Genetic Contributions to Negative Schizophrenia Symptoms.

Note. GABA = gamma amino butyric acid; miRNA = microRNA.
aAlso an imaging genetics study.
Other dopamine-related genes
Dopamine beta hydroxylase (DBH) gene, encodes for the DBH enzyme, which converts dopamine to norepinephrine. Research has identified associations between DBH polymorphisms in the 5′ area and positive schizophrenia symptoms. The del allele results in reduced promoter activity and subsequently lower levels of DBH and enzyme activity in plasma and cerebrospinal fluid (Cubells et al., 1997). In two studies in the Han Chinese population, researchers found that the del/del genotype was associated with more severe positive symptoms than the ins/del and ins/ins genotypes (Hui et al., 2012; Zhou et al., 2013). This evidence also strengthens the hypothesis that positive symptoms are associated with excessive dopaminergic action.
Cholecystokinin type A receptor (CCKAR) gene encodes for a G-protein-coupled receptor that regulates release of beta endorphins and dopamine in the central nervous system (NCBI gene ID 886). Haplotype analysis of five risk variants (see Table 2) has revealed associations with positive symptoms in schizophrenia (Zheng, Fu, Shen, & Xu, 2012). Although the functional mechanism of the association between CCKAR and positive symptoms is not known, CCKAR coexists in the same dopaminergic neurons (X. Y. Zhang, Zhou, Zhang, & Wei, 2000).
Reviewed Studies That Contribute Evidence for Genetic Contributions to Positive Symptoms of Schizophrenia.
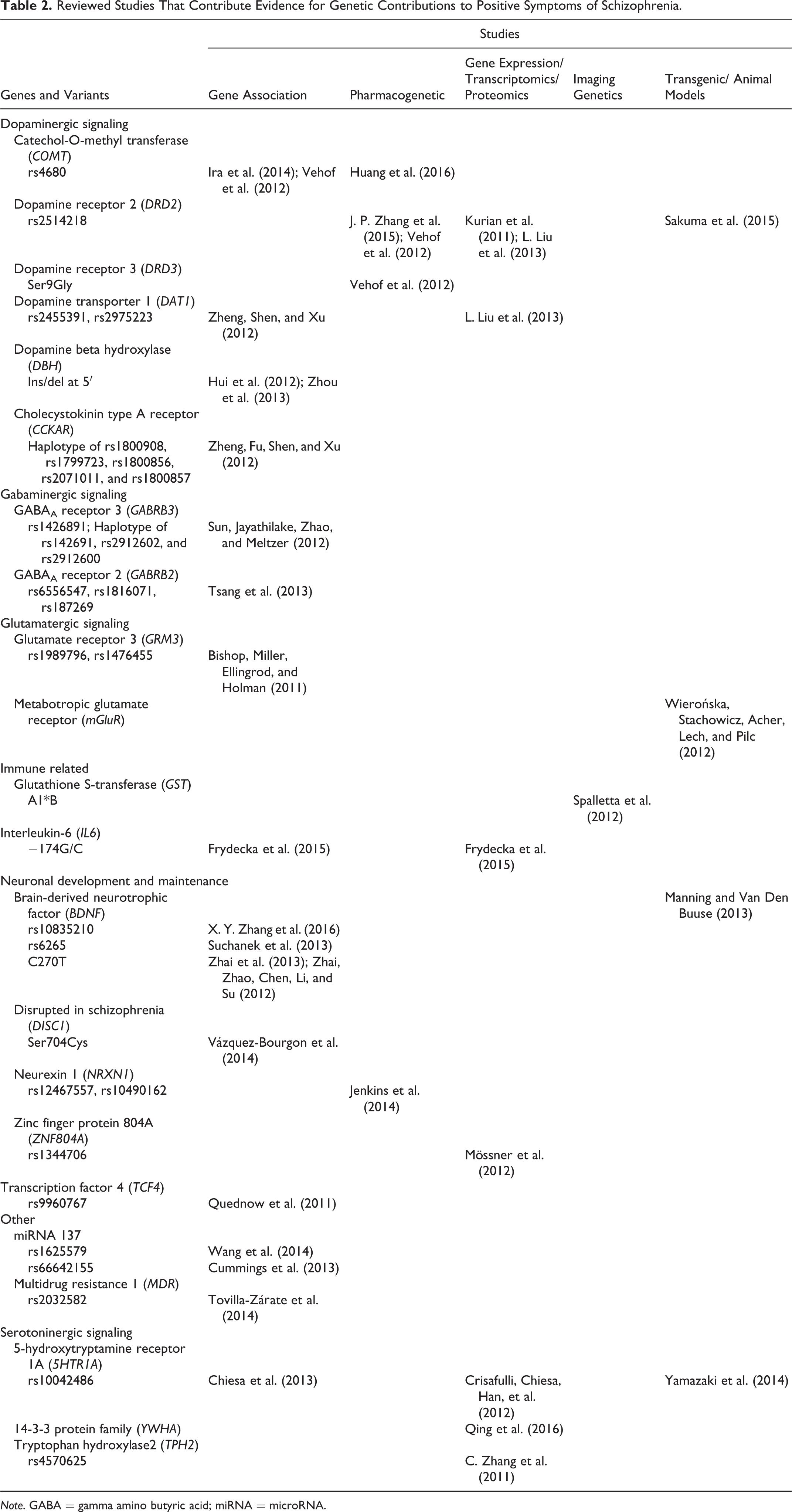
Note. GABA = gamma amino butyric acid; miRNA = microRNA.
ALDH2, though primarily associated with alcoholism and alcohol dependence (S.-Y. Lee et al., 2010), is also associated with negative symptoms of schizophrenia (S. Y. Lee et al., 2015). ALDH2 encodes for the ALDH isoenzyme, which catalyzes dopamine oxidation (Lamensdorf et al., 2000), and the risk variant is associated with improvement in negative symptom scores in patients treated with a combination of risperidone and dextromethorphan (which has a protective effect on monoamine neurons through its NMDA receptor antagonistic action; S. Y. Lee et al., 2015). Monoamine oxidase A and B are also involved in dopamine metabolism and influence negative symptoms, specifically affective flattening (Camarena et al., 2012).
Serotoninergic signaling
Serotonin, or 5-hydroxytryptamine, is another neurotransmitter that has been associated with positive symptoms, primarily through its effect on the dopaminergic system, though the exact mechanism is not known (Stahl, 2013). The 5-hydroxytryptamine receptor (5HTR) gene encodes for the serotonin receptors, which are broadly distributed across the brain, particularly in the corticolimbic areas (Lesch & Gutknecht, 2004). Investigators believe that one of the mechanisms of action of second-generation antipsychotics is through the action of the serotonin receptor 5HTR1A (Jordan et al., 2002). Positive (Crisafulli, Chiesa, Han, et al., 2012) and negative (Chiesa et al., 2011) associations of the 5HTR1A variant rs10042486 with positive symptoms of schizophrenia have been reported in a Korean sample. Animal models provide some evidence for associations of a different serotonin receptor, 5HT5A, with positive-like symptoms such as hyperactivity (Yamazaki et al., 2014).
5HTR1A has also been associated with negative symptoms. Converging evidence from pharmacogenetic (Bosia et al., 2015; Crisafulli, Chiesa, Han, et al., 2012) and gene expression studies (Takekita et al., 2015; Tang, Dalton, Srisawat, Zhang, & Reynolds, 2014) supports the idea that serotoninergic receptors have an influence on negative symptoms of schizophrenia. Researchers found that the G allele of 5HTR1A influenced severity of negative symptoms, possibly due to its inhibitory effect on pyramidal glutamatergic cells, which subsequently inhibits dopamine in the striatum (Diaz-Mataix, Artigas, & Celada, 2006). The presumption is that negative symptoms such as motivational deficits are associated with lower dopamine levels in the mesolimbic dopaminergic pathway.
The YWHA gene (also involved in serotonin metabolism) family encodes for the 14-3-3 proteins that are critical for a wide variety of cellular processes and has been associated with schizophrenia (Bell et al., 2000). Proteins in the 14-3-3 family activate tryptophan hydroxylase, which is involved in the synthesis of serotonin and also has been implicated in important immune cell function in schizophrenia (Qing et al., 2016). In one study, researchers found that dysregulated expression of this gene family in the peripheral white blood cells was associated with positive symptoms of schizophrenia in a Chinese sample (Qing et al., 2016). The tryptophan hydroxylase 2 (TPH2) gene encodes for the TPH enzyme, which regulates the biosynthesis of serotonin. The risk variant rs4570625 of TPH2 is associated with positive symptoms, further strengthening the evidence for the relationship between serotonin and positive symptoms in schizophrenia (Zhang et al., 2011).
Glutamatergic signaling
Glutamate is an excitatory neurotransmitter, and glutamatergic signaling problems with N-methyl-D-aspartate (NMDA) receptor hypofunction is another leading hypothesis regarding the pathophysiology of schizophrenia. This hypothesis originated from data demonstrating the psychotic effects of NMDA receptor antagonists such as phencyclidine and ketamine (Javitt, 1986). In addition to NMDA receptors, there are three other glutamate receptors in the central nervous system, metabotropic glutamate receptors (mGluR), α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors, and kainite receptors (Stahl, 2013), the first two of which have been associated with schizophrenia pathology. mGluR also influences the dopaminergic system (Pin & Duvoisin, 1995). A study of transgenic mGluR knockout mice with pharmacological manipulation showed some association of mGluR4 with positive-like symptoms such as hyperactivity and head twitches (Wierońska, Stachowicz, Acher, Lech, & Pilc, 2012). Glutamate receptor 3 (GRM3) encodes the mGluR3 protein, which is a G-protein-coupled receptor and has been associated with refractory global psychotic symptoms (Bishop, Miller, Ellingrod, & Holman, 2011).
Evidence for associations between genes related to glutamatergic signaling and negative symptoms of schizophrenia comes from gene association, pharmacogenetic, gene expression, and animal studies. The glutamate ionotropic receptor NMDA type subunit 1 (GRIN1) gene, is a protein-coding gene. Transgenic GRIN1 hypomorph (mutation causing partial loss of gene function) mice have severe social deficits (Hanks, Dlugolenski, Hughes, Seymour, & Majchrzak, 2013). The glutamate ionotropic receptor AMPA type subunit 2 (GRIA2) gene, encodes for AMPA subunit 2 and is associated with improvements in negative symptoms in patients treated with antipsychotics (Crisafulli, Chiesa, De Ronchi, et al., 2012). GRM3, a gene that codes for a Type 3 mGluR, is also associated with negative symptom improvement in patients treated with risperidone (Bishop et al., 2015). Converging evidence also comes from studies of glutamate AMPA receptor 1 knockout (GluA1-KO) mice, which demonstrate significantly reduced social behaviors (Barkus et al., 2012). Specific proteins that are believed to be associated with glutamate and dopaminergic pathways in the cerebellum are also associated with negative symptoms of schizophrenia, further strengthening evidence of the association of the glutamatergic system with the negative symptom domain (Pinacho et al., 2013).
Gabaminergic signaling
Gamma amino butyric acid (GABA) is the primary inhibitory central nervous system neurotransmitter. Genes in the GABRB family encode for GABAA receptor beta subunits. Polymorphisms of these genes, specifically of GABRB2 and GABRB3, are associated with positive symptoms of schizophrenia (Tsang et al., 2013) and hallucinations, respectively (Sun, Jayathilake, Zhao, & Meltzer, 2012). A study of platelet-derived growth factor receptor knockout (PDGFR-β-KO) mice, which have deficits in social behaviors (equivalent of negative symptoms in humans) and prepulse inhibition (a physiological measure that is associated with sensory gating and positive symptoms), found associations with phase-locked gamma oscillations, which are believed to be from GABAergic parvalbumin-immunoreactive neurons (Nakamura et al., 2015), further strengthening the evidence for an association between gabaminergic signaling and positive and negative symptoms of schizophrenia.
Genes involved in neurodevelopment, maintenance, and neuroplasticity
Brain-derived neurotrophic factor (BDNF) is a member of the neurotrophin family of growth factors with a critical role in neuronal development, survival, maturation, and maintenance. The BDNF gene, which encodes for BDNF, is highly expressed in the adult brain (Adachi, Numakawa, Richards, Nakajima, & Kunugi, 2014), and several risk variants in this gene have been associated with positive symptoms in schizophrenia. A allele carriers of the variant rs10835210 have more severe positive symptoms than noncarriers (X. Y. Zhang et al., 2016). Another risk variant in this gene, Val66Met (rs6265), has been associated with positive symptoms (hallucinations) in Polish (Suchanek et al., 2013) and Han Chinese samples (Zhai et al., 2013). For variant C270 T, the CT genotype was associated with more severe positive symptoms than the CC or TT genotypes (Zhai, Zhao, Chen, Li, & Su, 2012). Transgenic animal models have demonstrated an association between BDNF and positive symptoms that has a sex effect, with female mice being less sensitive to the genotype effect (Manning & Van Den Buuse, 2013). BDNF is also correlated with negative symptoms (W. Li, Zhou, et al., 2013; Martinho et al., 2012). Low levels of plasma BDNF are associated with more severe negative symptoms, and research has suggested that they are an indicator of poorer neuroplasticity (S. L. Chen et al., 2014). The association between low BDNF levels and negative symptoms has not, however, been consistently reported in the literature (Zhai et al., 2012).
Transcription factor 4 (TCF4), which encodes for basic helix-turn-helix transcription factor, is expressed in both the immune system and the neuronal cells and is required for neuronal differentiation in the developing brain (Ayalew et al., 2012). TCF4 has several splice variants that encode for different proteins, has been associated with delusional symptoms (decrease in delusional states), and is considered a candidate blood marker for delusions (Ayalew et al., 2012). Research has provided further evidence of the association between TCF4 and positive symptoms with the finding that the presence of the risk allele rs9960767 is associated with p50 suppression, which is a physiological marker for sensory gating abnormalities seen in schizophrenia (Quednow et al., 2011). Authors have speculated that impaired sensory gating results in information overload, resulting in psychotic and other cognitive symptoms in schizophrenia (Roussos, 2012). Studies have also shown an association between positive symptoms and ZNF804A, a zinc finger protein gene hypothesized to exert a role in neurodevelopment (Mössner et al., 2012). Genome-wide microRNA (miR) expression studies have also shown associations between positive symptoms and enrichment of genes associated with neuronal maintenance and neurogenesis in the anterior prefrontal regions of the brain (Barnes et al., 2011).
The disrupted in schizophrenia (DISC1) gene encodes for the DISC1 protein, which is involved in neurodevelopmental processes and neural cell signaling through a complex interaction with the DISC1 interactome (Hennah & Porteous, 2009). Presence of the Ser704Cys genotype for this gene is associated with hallucinations (Vázquez-Bourgon et al., 2014), and an earlier study found an association between presence of this genotype and lifetime severity of delusional symptoms in schizophrenia, hypothesized to be mediated by the hippocampus (Ser704Cys is associated with the structure and function of the hippocampus; DeRosse et al., 2007). The neurexin 1 (NRXN1) gene encodes for neurexin 1, a transsynaptic cell-adhesion membrane protein that is involved in neural development, maturation, and maintenance. The risk variants (rs12467557 and rs10490162) are associated with changes in positive symptoms and thought disturbance in patients treated with antipsychotics (Jenkins et al., 2014). Similarly, NRXN1 is also associated with changes in negative symptoms in patients treated with antipsychotics (Jenkins et al., 2014; Lett et al., 2011).
Genome-wide association studies have added to the knowledge regarding genetic associations with negative symptoms, revealing associations with BCL9, C9orf5, ST3-related genes, RNF144, and CTNNA3 (Xu et al., 2013). BCL9 is associated with the Wnt signaling pathway along with genes such as DVL2 and GSK3B (Bousman et al., 2012), which are involved in neuroplasticity, cell survival, and adult neurogenesis. C9orf5 has been linked to information processing, RNF144 is associated with antipsychotic metabolism, and CTNNA3 has been associated with epigenetic regulation, neuronal migration, and synaptogenesis (Xu et al., 2013). NCAM (Piras et al., 2015), AHI1 (Porcelli et al., 2015), and TCF4 (Wirgenes et al., 2012) are other genes critical for neuronal development that are also associated with negative symptoms of schizophrenia.
Immune system genes
Dysregulation of the immune system has been associated with schizophrenia risk, but there is also growing evidence that it is associated with positive and negative symptoms, although the evidence is less compelling at this point for negative symptoms. Interleukin 6 (IL-6) encodes for interleukin-6, a cytokine involved in immune-mediated processes that promotes the survival of catecholaminergic neurons that activate both dopaminergic and serotoninergic neurons in the prefrontal cortex and hippocampus (Zalcman et al., 1994). The IL6-147G/C genotype has been associated with positive symptoms of schizophrenia (Frydecka et al., 2015). Research has also revealed a role for interleukins in the pathogenesis of negative symptoms, specifically an association of IL28B with negative symptoms (C. Chen et al., 2012).
Carrying the *B allele of the glutathione S-transferase (GSTA1) gene was associated with positive symptoms, specifically severe global hallucinations, in an imaging genetic study (Spalletta et al., 2012). GSTA1*B allele was also associated with higher mean diffusivity (in a diffusor tensor imaging study) in the thalamic region, suggestive of microstructural damage in this area. The thalamus is the critical sensory relay station, and these associations suggest a plausible mechanism by which this gene may affect the pathogenesis of positive symptoms.
In their study of methylation patterns in whole blood, J. Liu et al. (2014) demonstrated that 11 cytosine–phosphate–guanine sites played a protective role in decreasing hallucinations and delusions in patients with schizophrenia. The researchers found that CD244, LAX1, TXK, PRF1, CD7, MPG, and MPO, which are primarily involved in immune response, were differentially methylated and concluded that methylation changes could modulate immune system responses that may have a protective effect on positive symptoms. SHP-1 expression, which is an indicator of inflammatory processes, has been associated with negative symptoms (Pesce et al., 2014).
Posttranscriptional control
There is also growing evidence for an association between altered expression of miRs and schizophrenia risk as well as both positive and negative symptoms. miR-137 variants have been associated with positive symptoms (Cummings et al., 2013; Wang et al., 2014), whereas miR137 (Kuswanto et al., 2015), miR181b (Song et al., 2014), and miR449a (Lai et al., 2011) have been associated with negative symptoms, the mechanism of which is thought to be the silencing of gene expression at the posttranscriptional level (Ding, Weiler, & Großhans, 2009).
Unique Gene Associations With Positive Symptoms
Genes associated with positive, but not negative, symptoms of schizophrenia include multidrug resistance 1 (MDR1), which encodes for P-gp, a transmembrane drug transporter in the blood–brain barrier. Polymorphisms of this gene were associated with hallucinations and bizarre behavior in a Mexican sample, although the functional mechanisms of this association are unknown (Tovilla-Zárate et al., 2014). Rare duplication burden has been negatively correlated with positive symptoms (Martin, Robinson, Reutens, & Mowry, 2015a), whereas common genetic variant burden calculated as a polygenic risk profile score was positively associated with positive symptoms (Martin, Robinson, Reutens, & Mowry, 2015b).
Unique Gene Associations With Negative Symptoms
Folate metabolic pathway
Research has shown that a number of genes in the folate metabolic pathway, such as MTHFR, MTR, and FOLH1, are associated with negative, but not positive, symptoms of schizophrenia (Hill et al., 2011; Roffman et al., 2013). Folate is an important substrate involved in intracellular methylation, and alteration in folate metabolism can affect normal brain development and function (Frankenburg, 2007). This association between negative symptoms and folate genes could point to the putative role of neurodevelopmental aberrations in the pathogenesis of negative symptoms.
Solute carrier family (SLC) genes
SLC genes, which encode for membrane transporters such as serotonin and GABA transporters, are associated with the severity of negative symptoms. Specifically, findings have shown that novel missense mutations in SLC39A13 (Kranz et al., 2016) as well as common variants in SLC6A4 (W. Li, Yang, et al., 2013) and SLC6A12 (Park et al., 2011) are associated with negative symptoms, though evidence for these associations has been inconsistent across studies (Carlström et al., 2012; Sun et al., 2012).
Other genes
Research has also found that a number of other genes are associated with negative symptoms, including CPLX1, suggesting involvement of mitochondrial dysfunction, and AVPR1A, which encodes for arginine vasopressin, has a putative role in social and emotional functioning, and is regulated by dopamine and serotonin (Golimbet, Alfimova, Abramova, Kaleda, & Gritsenko, 2015). Also, Fanous et al. (2012) demonstrated a polygenic association with negative/disorganized symptoms by utilizing polygenic scores in the Molecular Genetic Study of schizophrenia.
Discussion
There is a growing body of evidence from different approaches (genetic studies in humans to animal models) to support the genetic basis of positive and negative symptom domains in schizophrenia (see Tables 1 and 2). Although this evidence is preliminary and comes primarily from genetic association studies, using a symptom-based subphenotype approach in genetic studies seems to be a useful strategy to deconstruct pathophysiological mechanisms in a complex condition like schizophrenia. As with schizophrenia risk, the genetic architecture of these symptom domains appears to be polygenic, with several gene associations exerting small effects. The evidence in this review points to shared and unique gene associations in the biologic basis of these symptom domains (see Figure 2).
For positive symptoms, evidence of genetic associations points primarily to dopaminergic signaling, specifically in the mesolimbic regions (Sakuma et al., 2015; J. P. Zhang et al., 2015). Theoretically, this evidence aligns with the hypothesis that excess mesolimbic dopamine influences positive symptoms (Stahl, 2013). The mesolimbic pathway is associated with both positive and some negative symptoms such as motivational deficits, though it is dopamine excess that plays a role in positive symptoms and a deficit in dopamine that plays a role in negative symptoms. It is not surprising that negative symptoms are also associated with major neurotransmitter genes, such as those involved in the dopaminergic, serotoninergic, and glutamatergic systems, as these genes are primarily responsible for chemical neurotransmission in the central nervous system. Authors have hypothesized that these neurotransmitter systems interact to create a “neurochemical mobile,” which is a complicated network that maintains the dynamic balance important for normal function (Qi et al., 2016). Any alteration of this interaction in the form of excess or deficit in neurotransmitter levels or faulty receptors or transporters in one system can lead to a change in another, resulting in systemic disarray. Evidence is lacking as to exactly where and how this disarray leads to positive and negative symptoms of schizophrenia.
Several genes involved in neuronal growth, maturation, and maintenance are also associated with both positive and negative symptoms. Schizophrenia is a neurodevelopmental disorder, but the role these genes play in the pathophysiology of the symptom domains is not clear. There is also evidence to support epigenetic influence (methylation and posttranscriptional control by miRs) in addition to gene effects on these symptoms. There were several genetic associations unique to the negative symptom domain such as the folate metabolic pathway genes, SLC genes, and increased CNV burden, but the exact nature of these relationships is also not known.
The studies we included in this review addressed the issue of genetic contributions to these symptom domains in different ways: some chose the broader symptom domain as a single construct while others examined specific symptoms in each domain (such as hallucinations, delusions, motivational deficit, etc.). This discrepancy could be one explanation (in addition to confounds such as gender and ancestry) of the equivocal findings for some gene associations. Clinically, positive and negative symptom domains have demonstrated small-to-modest correlations with each other (and with the cognitive domain; Keefe et al., 2006), which the shared genetic associations highlighted in this review might explain. Similarly, significant genetic overlap may exist between cognitive domains and positive and negative domains, but such questions were beyond the scope of this review.
The evidence presented in this review suggests that there is merit in pursuing a genomic symptom science approach as such an approach can help delineate pathophysiological processes in complex conditions such as schizophrenia. This approach will be particularly useful for developing precision and personalized treatment and care approaches for patients with specific and challenging clinical profiles. For example, treatment-resistant patients with severe positive and negative symptoms, numerous hospitalizations, and poor treatment response rates who have a genome-wide total duplication burden (Martin & Mowry, 2016) may benefit from one particular treatment approach. Meanwhile, patients with deficit syndrome characterized by a prominence of negative symptoms who do not respond to conventional treatments or who do not have the insight to commit to and follow through with treatment (pharmacotherapy and psychosocial interventions) may require a different approach to therapy. First-line treatments such as pharmacotherapy primarily target positive symptoms and have poor to limited efficacy in the management of negative symptoms and cognitive impairment, resulting in minimal improvement in social and occupational functioning. As negative and cognitive symptom domains significantly contribute to functional impairment, there is a dire need for treatments that target them. A genomic symptom science approach can help elucidate the functional genetic mechanisms for individual symptoms as well as symptom domains and is thus critical for the development of precision treatment and care for patients afflicted with schizophrenia.
Footnotes
Author Contribution
R. M. Xavier contributed to conception, design, data acquisition, data analysis, and interpretation; drafted the manuscript; critically revised the manuscript; gave final approval; and agrees to be held accountable for all aspects of work, ensuring integrity and accuracy. A. Vorderstrasse contributed to conception, design, and interpretation; critically revised the manuscript; gave final approval; and agrees to be held accountable for all aspects of work, ensuring integrity and accuracy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.