
Abstract
Pain can have negative, physiological and psychological impacts on pregnancy. Pregnant women are fearful of using pain medication because of teratogenic effects. In this study, we evaluated whether exercise could lower pain sensitivity in pregnant mice with neuropathic pain and reduce the negative effects of maternal pain on newborns. We randomly assigned 32 female mice to one of four groups (eight mice/group): Sham surgery with standard environment (SE) or enriched environment (EE) or spare nerve injury (SNI) with SE or EE. Mice in EE groups had access to an exercise wheel. Mothers were evaluated for mechanical sensitivity with Von Frey filaments and for exercise performance with computerized running wheels. Mice were impregnated 2 weeks after the initiation of EE. Pups were weighed and measured for length at birth and evaluated for negative geotaxis, righting, forelimb grasping, rooting, and crawling at 3 days postpartum and for crawling at 6 days postpartum. Following euthanasia, mothers’ frontal cortexes were analyzed for selected neuropeptides. After exercise exposure, only SNI-SE females remained neuropathic. Exercise levels were similar between EE groups. Some brain neuropeptides (endorphins, enkephalins, and oxytocin) from SNI females showed significant differences with exercise. Number of pups was significantly smaller in the SNI-SE group. Significantly more pups died at birth in the SNI-SE group, but pup behavior tests (except righting) were similar across groups. Exercise can reduce neuropathic pain in pregnant mice. Neuropathic pain does not impact motor neurodevelopment of mice pups but does appear to affect litter size and neonatal mortality.
In the general population, approximately 20% of women (Meana, Cho, & Desmeules, 2004) and 10% of individuals under the age of 60 years (Crook, Rideout, & Browne, 1984) experience chronic pain. During pregnancy, women suffering from pain are fearful of using pain medications because of teratogenic effects (Nordeng, Ystrom, & Einarson, 2010). However, untreated chronic pain can lead to physiological changes such as the development of hypertension and psychological changes such as the development of anxiety and depression, all of which can negatively impact pregnancy (Bruehl, Chung, Jirjis, & Biridepalli, 2005). Use of pain medication can also lead to clinical problems. Aspirin and nonsteroidal anti-inflammatory drugs can prematurely close the fetal ductus arteriosus (Koren, Florescu, Costei, Boskovic, & Moretti, 2006). Regular use of opioids can lead to neonatal withdrawal syndrome, and opioids can enter the mother’s milk (Briggs, Freeman, & Yaffe, 2002). Even with small doses of codeine, there is a risk of neonatal abstinence syndrome (Khan & Chang, 1997; Reynolds, Riel-Romero, & Bada, 2007). Thus, a nonpharmacological approach for chronic pain management might improve maternal and fetal outcomes.
Environmental enrichment (EE) has become more widely recognized as a means to alleviate chronic pain (Tai, Yeung, & Cheung, 2018). Exercise is an accessible intervention nurses can propose to patients (Pearce, Evenson, Symons Downs, & Steckler, 2013). It is also a recommended behavior during pregnancy in general to improve outcomes (Davenport et al., 2019). It thus makes sense to explore the role of exercise during pregnancy in women with chronic pain.
EE reduced neuropathic pain by increasing the mechanical sensitivity threshold in sensitized mice (Vachon et al., 2013) and rats (Parent-Vachon & Vachon, 2018). In CD-1 mice, demethylated DNA in the prefrontal cortex, which was associated with neuropathic pain, was reduced with EE (Tajerian et al., 2013). EE reduces allodynia and hyperalgesia and is associated with reductions in the levels of biomarkers of central pain sensitivity (Grace et al., 2016; Ye, Du, & Hua, 2017). EE is often composed of many different elements including social and psychological elements, as well as object novelty, nutrition, and exercise. Exercise alone can induce analgesia by modulating inflammatory cytokines and by causing the release of endogenous opioids in the central and peripheral nervous systems (Kami, Taguchi, Tajima, & Senba, 2016; Kami, Tajima, & Senba, 2017). Exercise can also induce pain in the contexts of a musculoskeletal condition or sedentary lifestyle (Lima, Abner, & Sluka, 2017). However, performing regular exercise typically induces analgesia through activation of endogenous opioids and serotonin (Sluka, Frey-Law, & Bement, 2018). EE, and more specifically, exercise, might therefore be an effective alternative to pharmacological treatments of chronic pain.
Neuropathic pain is a type of chronic pain described as a lesion or dysfunction of the peripheral or central nervous system that causes an increase in the sensitivity of neurons (Mumion, 2018). There is only one published paper on the effects of neuropathic pain during gestation in mice (Rosen et al., 2017). In that study, researchers performed spare nerve injury (SNI) surgery in early gestation, late gestation, and postweaning, which thus did not allow for exploration of the effects of a neuropathic pain condition prior to conception. In another study, authors described the effects of chronic inflammatory pain prior to gestation (Zhou et al., 2018). In that study, researchers created a model of chronic inflammation with hyperalgesia by injecting complete Freund’s adjuvant (CFA) in the mouse hind paw prior to conception. In CFA-treated mothers, researchers observed a higher rate of prematurity, smaller litter size, lower rate of live birth, lower birth weights, and lower success rate in the pups of crawling and the righting reflex. With EE, the pain thresholds of the mothers were increased and all of the negative outcomes were improved. However, the EE comprised a number of different elements including a house-shaped hole, ladders, sliding rope, soft wood for biting, as well as peanuts and sunflower seeds. The two food elements contain anti-inflammatory constituents as well as antioxidants, which may have contributed to the reduction in chronic inflammation and hyperalgesia following CFA injections. Although these findings are interesting, neuropathy is a much more common cause of moderate-to-severe inflammatory pain and hyperalgesia in women.
The objectives of the present study, therefore, were to use a well-recognized model of neuropathic pain in mice (SNI) to evaluate whether exercise can lower pain sensitivity in female mice with chronic neuropathic pain before and during pregnancy, improve the reproductive performance of these mothers, and diminish the negative effects of maternal neuropathic pain on newborns.
Method
Animals and Environmental Conditions
We used thirty-two 8-week-old female and 16 male CD-1 mice for these experiments (specific pathogen-free [SPF] grade, Charles River Laboratories, Kingston, NY). We selected CD-1 mice because of their reproductive proficiency and good maternal behavior (Giknis & Clifford, 2007). Upon their arrival, we kept mice in a standard animal-facility environment (fresh filtered air, temperature 21 °C ± 2 °C, humidity 40–60%, and light–dark cycle 12:12 hr). Water and a rodent diet (Teklad, 2018, global protein diet, Envigo,) were provided ad libitum. Mice were housed two per ventilated polycarbonate cage (Sealsafe Plus GM500, Tecniplast, Inc., Varese, VA).
We randomly assigned female mice to one of four groups (eight mice/group): (1) sham surgery with standard environment (sham SE), (2) sham surgery with enriched environment (sham EE), (3) spare nerve injury surgery with standard environment (SNI-SE), and (4) spare nerve injury surgery with enriched environment (SNI-EE). Male mice arrived 1 week prior to mating for acclimation. All females were provided with nesting material (Nestlet, Ancare, Bellemore, NY). Mice in the standard environment were provided with a mouse igloo (Mouse House, Tecniplast Inc.), while the mice with an enriched environment also had access to an exercise wheel (Fast-Trac activity wheel, BioServ, Flemington, NJ) starting from the second week following the surgery. Previous studies found these wireless running wheels to be effective for evaluating running performance (Sanchez-Roige et al., 2014). The Animal Ethics Research Committee of the Sainte-Justine University Hospital Research Center approved the protocol.
Research Protocol
Figure 1 presents the study time line.
Week 1: Following their arrival, female mice were acclimated to the facilities. Week 2: We took weights for female mice and assessed their mechanical sensitivity using von Frey filaments. Week 3: Female mice underwent surgery on the left leg under isoflurane anesthesia to produce a SNI model (sham surgery as control) according to previous published methods (Decosterd & Woolf, 2000). Briefly, the tibial and common peroneal nerves were tied with 6-0 suture and cut; the sural nerve was left intact. For sham surgery, the nerves were visualized but not cut. Subcutaneous ampicillin 25 mg/kg (ampicillin 250 mg, Pfizer, Lake Forest, IL) was administered immediately following the surgery. The mice were then returned to their home cages. Week 5: We weighed female mice and assessed their mechanical sensitivity with von Frey filaments. Fast-Trac activity wheels were introduced following the von Frey assessments. EE mice had access to these wheels for the duration of the study except for during the computerized running performance evaluations. Running wheels were removed approximately 3 days prior to parturition. Week 6: Exercise was quantified over a 24-hr period by replacing the Fast-Trac activity wheel with a low-profile running wheel (Med Associates Inc., St Albans, VT). The wheel recorded the number of wheel turns per 24 hr. We divided that number by 2 to estimate the activity of each of the two mice per cage. Week 7: All the females were mated with male CD-1 mice with a ratio of one male for two females. Both females were introduced into the male’s cage. Female mice were assessed every day for the presence of a vaginal plug. If a vaginal plug was present, it was considered a sign of successful copulation and was defined as Day 0 of gestation. When both females had been copulated (all within 5 days), they were removed from the male’s cage and placed singly into new cages. The female mice were weighed 10 days later to establish the presence of gestation. Week 8: Female were weighed and locomotor activity was assessed with the low-profile running wheels. At birth: Pups were weighed and length was measured. Day 3, postdelivery: Pups were assessed for motor neurodevelopment, including negative geotaxis, righting, forelimb grasping, and rooting. Day 6, postdelivery: Pups were assessed for crawling.
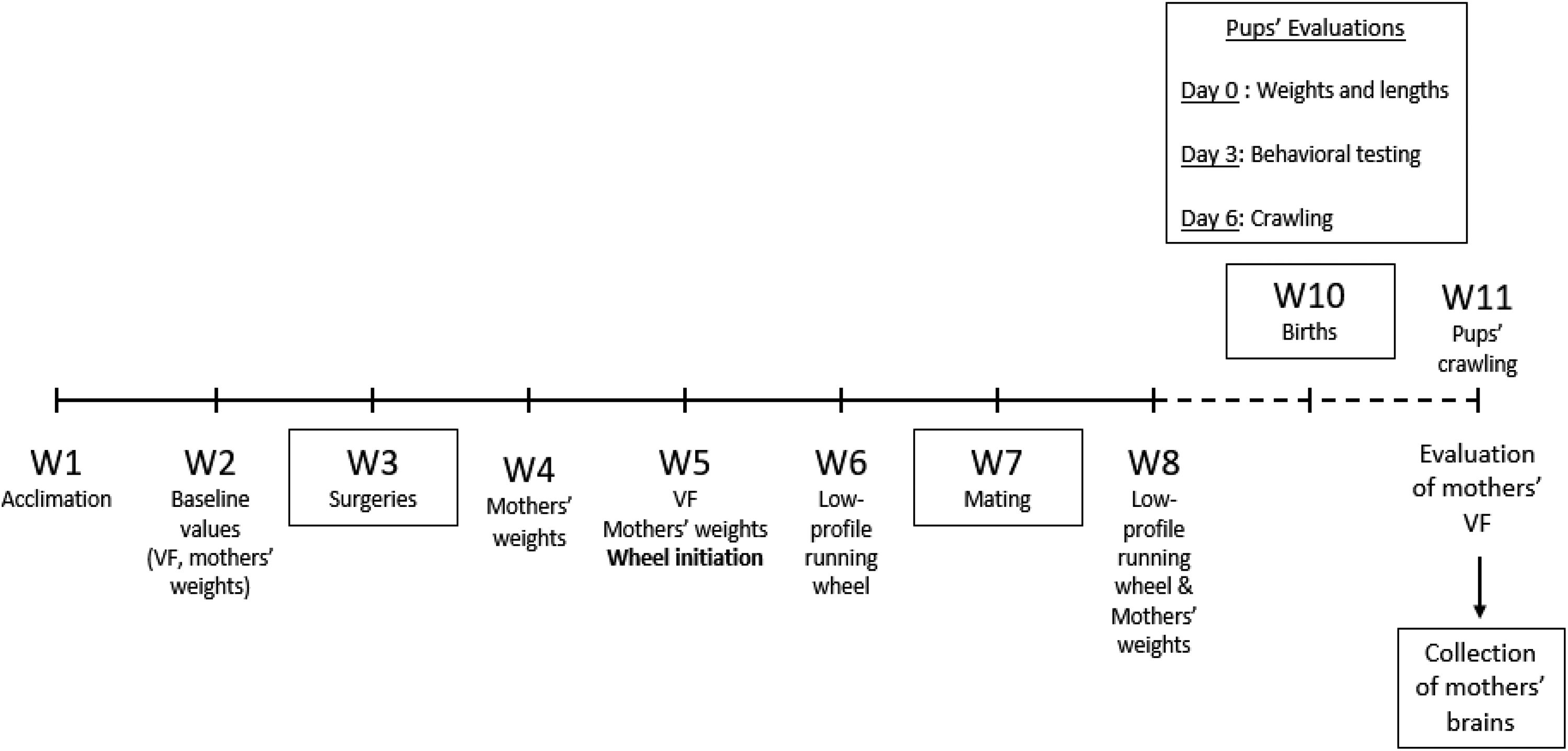
Time line of the research protocol. VF = von Frey filaments mechanical sensitivity evaluations; W = study week.
Evaluations of Female Mice
To assess mechanical sensitivity in the female mice, we followed previously published methods using von Frey filaments (Vachon et al., 2013). We used the lateral plantar surface of both hind paws and calculated a 50% threshold to withdraw (g), as previously described (Chaplan, Bach, Pogrel, Chung, & Yaksh, 1994). Prior to testing, mice were acclimated to the setup for 60 min. We applied calibrated von Frey filaments (Stoelting Co., Wood Dale, IL) to the point of bending for 4 s or until withdrawal. The stimulus intensity ranged from 0.02 to 2.0 g, corresponding to filament numbers 2.36–4.31. We applied an up-and-down method to determine the threshold of mechanical sensitivity.
Total gestation length was calculated from the date of the first presence of the vaginal plug to the birth of pups. The number of live and dead pups was noted on Days 0 and 3 postdelivery.
Following all evaluations, we euthanized females with CO2, rapidly collected their cerebral hemispheres and placed them on dry ice, and then stored them at −80 °C pending mass spectrometry analysis of peptides.
Pup Evaluations
We weighed the pups at birth and measured them from rostral nose to the beginning of the tail using a Vernier caliper. On Day 3 postdelivery, we assessed the motor neurodevelopment of the pups by testing negative geotaxis, righting, forelimb grasping, rooting, and crawling (Glynn, Sizemore, & Morton, 2007; Nguyen, Armstrong, & Yager, 2007).
We tested for negative geotaxis by placing pups face down on a flat, smooth, plastic surface at an angle of 45°. Pups successfully completed the test if they rolled 180° (face up) within 15 s. We recorded a semicompletion and a noncompletion as rotations of at least 90° and less than 90°, respectively.
We assessed righting ability by placing the pup in a supine position with all four paws pointing up. Pups successfully completed the test when they assumed a ventral position with all four limbs upright in a standing position within 15 s. We recorded a semicompletion when pups achieved a ventral position but with some limbs not upright and a noncompletion when pups failed to achieve a ventral position.
We assessed forelimb grasping ability by placing a toothpick in the middle of the paws of the pup. Successful completion of the task was achieved when pups grasped the toothpick. We measured rooting by stroking a toothpick on the side of the face of a pup. The pup completed the test successfully if it turned its head to the side being stroked.
We assessed crawling by placing the pup on a flat surface in the middle of a circle 5 cm in diameter. The pup completed the task successfully if it reached the distal limit of the circle in less than 30 s.
On Day 6 postdelivery, we again assessed the pups for crawling by putting them in the center of a circle 20 cm in diameter in a cage with softwood bedding. Each pup had a maximal time of 2 min to reach the distal limit of the circle. For pups who did not complete the task in 2 min, we recorded a time of 2 min. For crawling evaluations on Days 3 and 6, we recorded both a qualitative (success) and a quantitative (time to perform the task) variable.
We filmed all crawling evaluations (using Logitech Webcam C210, Logitech, Newark, CA). A single experienced evaluator blinded to group assessed all videos. We performed all behavioral tests in pups and mothers in a quiet, dimly lit room.
Evaluation of Brain Peptides
We weighed frontal brains (rostral third) of female mice (from Sham ES, SNI-SE, and SNI-EE) and aliquoted them to reinforced 1.5-ml homogenizer tubes containing 500 µm glass beads. We chose selected brain peptides from our previous pain studies (Ferland, Pailleux, Vachon, & Beaudry, 2011; Pailleux, Vachon, Lemoine, & Beaudry, 2013). A solution of 0.25% trifluoroacetic acid (TFA) was added at a ratio of 1:5 (w:v) and a Fisher Bead Mill was used at 5 m/s for 3 × 60 s to homogenize the sample. The homogenates were centrifuged at 12,000g for 10 min. The protein concentration for each homogenate was determined using a Bradford assay, and all samples were normalized to avoid any bias. To precipitate large and medium proteins, 100 ml of sample was mixed with 100 µl of methanol containing 0.1% TFA. Samples were centrifuged at 12,000g for 10 min. The supernatant was dried using an Eppendorf vacuum concentrator, resuspended in 50 µl of 5% acetonitrile in water containing 0.1% formic acid and transferred into low-volume high-performance liquid chromatography (HPLC) vials. The HPLC system was a Thermo Scientific Vanquish FLEX UHPLC system (San Jose, CA). The chromatography was performed using a gradient mobile phase along with a microbore column Thermo Biobasic C18 100 × 1 mm, with a particle size of 5 μm. The initial mobile phase condition consisted of acetonitrile and water (both fortified with 0.1% of formic acid) at a ratio of 5:95. From 0 to 2 min, the ratio was maintained at 5:95. From 2 to 62 min, a linear gradient was applied up to a ratio of 50:50 and maintained for 3 min. The mobile phase composition ratio was reverted to the initial conditions, and the column was allowed to equilibrate for 20 min for a total run time of 85 min. The flow rate was fixed at 50 µl/min, and 5 µl of sample was injected. A Thermo Scientific Q-Exactive Plus Orbitrap Mass Spectrometer (MS; San Jose, CA) was interfaced with a Thermo Scientific Vanquish FLEX UHPLC system using a pneumatically assisted heated electrospray ion source. MS detection was performed in positive-ion mode using the TOP-10 data-dependent acquisition method. Nitrogen was used for sheath and auxiliary gases, which were set at 10 and 5 arbitrary units, respectively. Auxiliary gas was heated to 200 °C. The heated electrospray ionization probe was set to 4,000 V and the temperature of the ion transfer tube was set to 300 °C. The scan range was set to m/z 400–1,500. Data were acquired at a resolving power of 70,000 (full width at half maximum [FWHM]) using an automatic gain control target of 1.0 × 106 and maximum ion injection time of 200 ms. Product ion spectra (MS2) were acquired at a resolving power of 17,500 (FWHM) using an automatic gain control target of 1.0 × 105, maximum ion injection time of 50 ms, and an intensity threshold of 5.0 × 104. The normalized collision energy was set to 28 (higher energy C-trap dissociation), and the isolation window was set to 2 Da. Peptide identification and quantification were performed using Proteome Discoverer 2.2 with a customized FASTA database including only specific neuropeptide sequences. Targeted peptide quantification was performed at MS1 level using specific precursor masses based on the monoisotopic masses. Peptide identification was performed by extracting specific precursor ions using a 5-ppm mass window and a label-free approach. The data were normalized based on the peak areas observed of the mean of the sham group for relative quantification. Instrument calibration was performed prior to all analyses, and mass accuracy was notably below 1 ppm using Thermo Pierce calibration solution and automated instrument protocol. Peptide sequences were verified based on typical b and y fragments observed at the MS2 level.
Statistical Analyses
Statistical analyses were performed and graphs created using GraphPad Prism 4.0. We used one-way analysis of variance (ANOVA) with repeated measures to compare mechanical sensitivity and body weight of females among groups. If we found significant differences, we then performed comparisons between the group means using the sequential Benjamini–Hochberg method. We also used a one-way ANOVA to compare time of gestation, numbers of pups per group, and crawling (3 and 6 days postdelivery) among groups. If we found significant differences, we performed post hoc Tukey tests. For all qualitative tests (geotaxis, grasping, rooting, righting, and crawling at 3 days), we used a χ2 analysis. If we found significant differences between groups, we calculated odds ratios. We analyzed 24-hr wheel turn count using Student’s t test. We expressed and plotted all data as mean ± standard error of mean (SEM) and considered p < .05 to indicate statistical significance.
Neuropeptide data were analyzed using a one-way ANOVA followed by Dunnett’s multiple comparison test (sham served as the control group). We set significance a priori to p < .05. These statistical analyses were performed using PRISM (Version 8.0).
Results
Maternal Measures
Figure 2 presents the results of the von Frey filament mechanical sensitivity testing of the female mice. At baseline (presurgery), we observed no significant differences between groups. As expected, 2 weeks following surgery, there were significant differences between groups, F(3, 28) = 40.30, p < .0001. Comparisons using the Benjamini–Hochberg method showed that mice in the SNI group significantly increased hypersensitivity to mechanical stimuli compared to those in the sham group (p < .0001). Following exercise exposure (1-week postdelivery), the SNI-SE animals differed significantly on the von Frey tests compared to the other groups (p < .0001), while the SNI-EE animals did not differ significantly from the sham groups on these tests. Running performance did not differ significantly among groups either prior to or during gestation (Figure 3). Weights of mothers did not differ significantly among groups throughout the experiment.
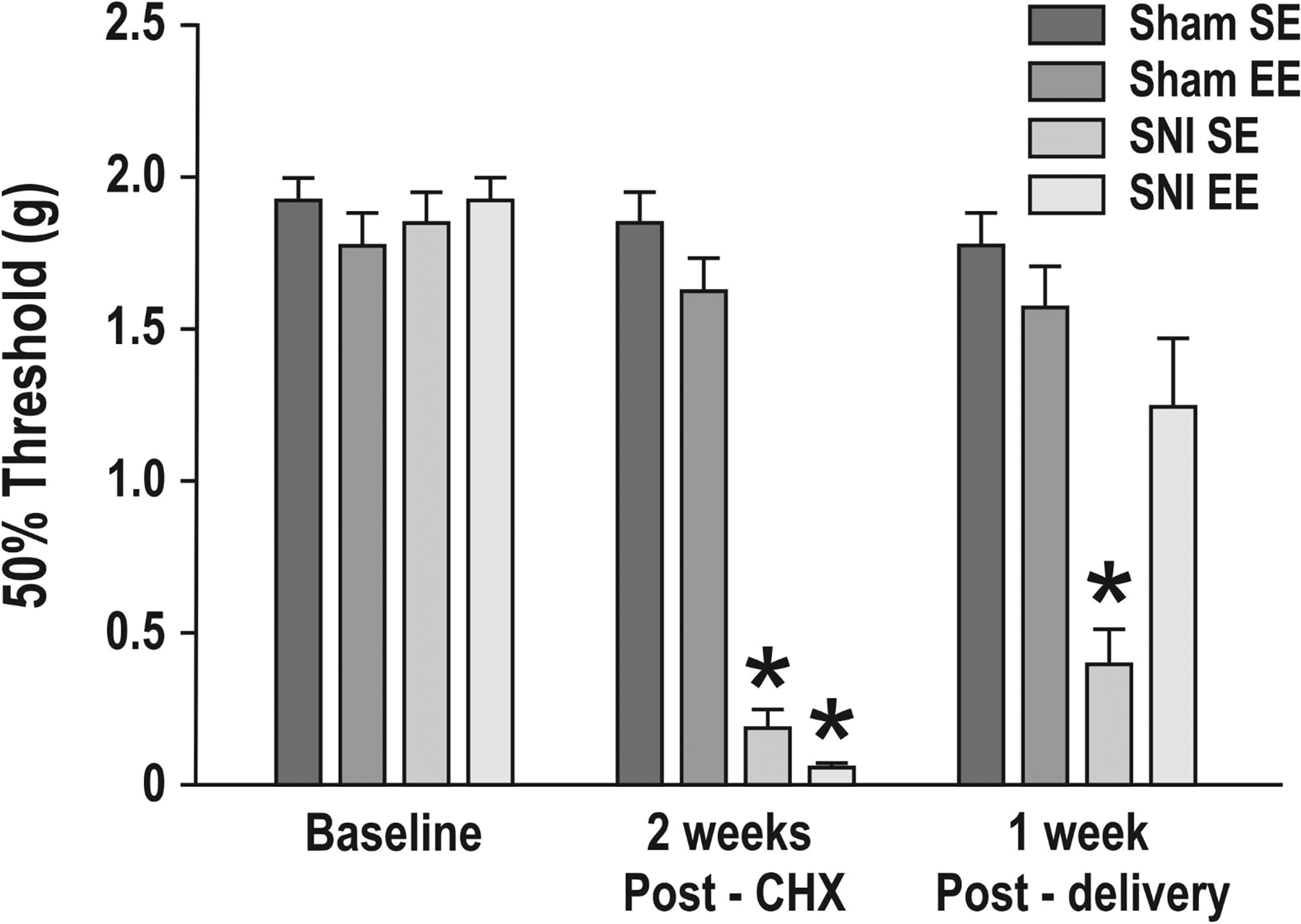
Mechanical sensitivity in female mice in the sham surgery with standard environment (sham SE), sham surgery with enriched environment (sham EE), and spared nerve injury surgery with standard (SNI-SE) and enriched (SNI-EE) environment groups (n = 8/group). Mechanical sensitivity was measured using an up-and-down method (50% threshold) with von Frey filaments at baseline (before surgery), 2 weeks postsurgery (post-surgery (CHX)), and 1 week postdelivery. Data are expressed as mean ± standard error of the mean. *Significant differences between the SNI-SE and SNI-EE groups compared to the sham groups at post-CHX and between the SNI-SE and all other groups at 1week postdelivery (p < .0001).
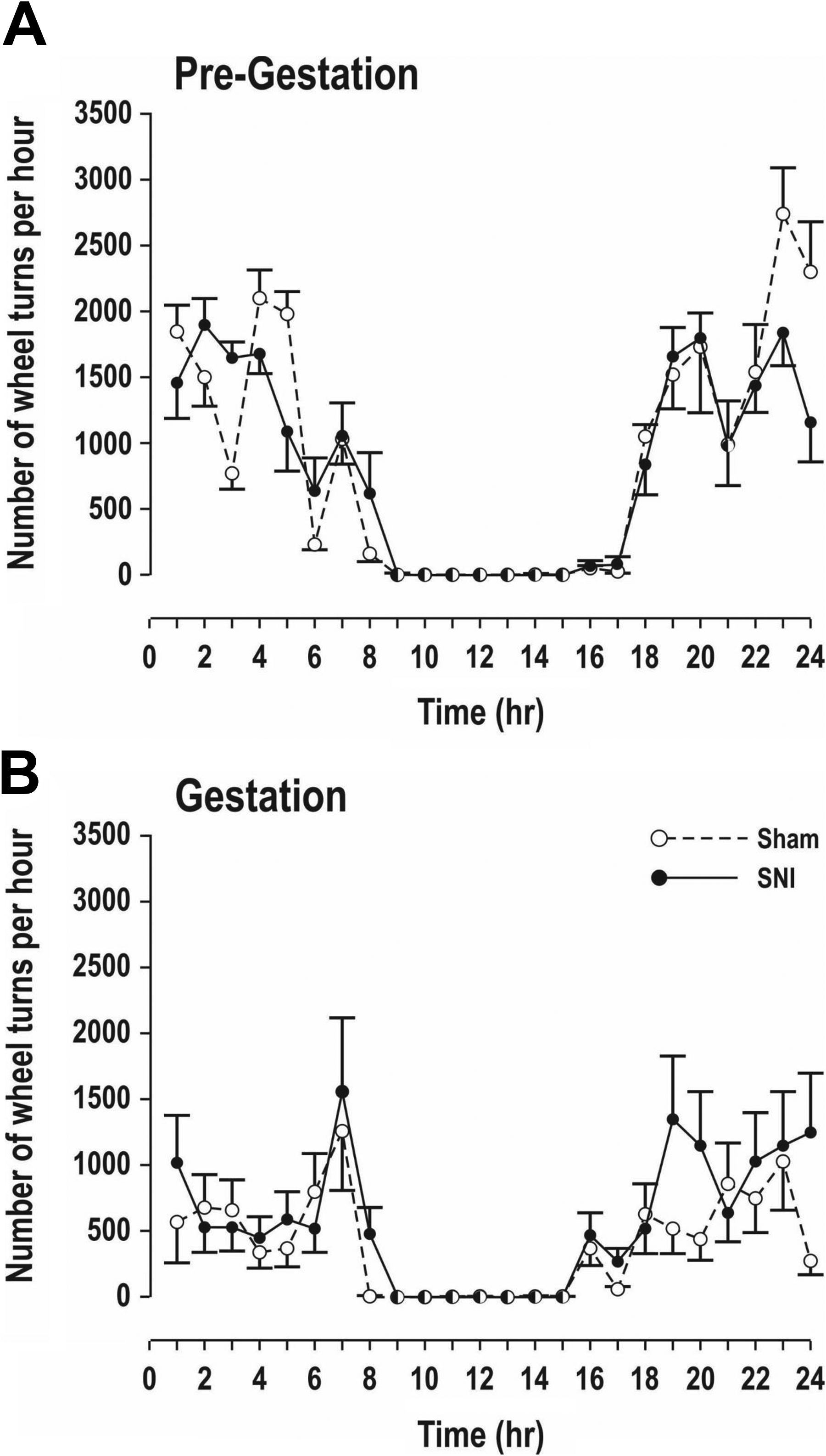
Mean (SEM) number of exercise wheel rotations/hr over 24 hr in the pregestational (A) and gestational (B) stages in sham surgery (sham) and spared nerve injury surgery groups of female mice with environmental enrichment (n = 8/group). There were no differences between groups in either recording period.
Peptide analysis in the frontal cortex of female mice (Figure 4) showed a significant increase for met-enkephalin (p < .05), leu-enkephalin (p < .05), and β-endorphins (p < .001) in the SNI-EE group compared to the sham groups. There was a significant increase of oxytocin (OT) in the SNI-SE group compared to the sham group (p < .05), with levels of OT in the SNI-EE group similar to those of the sham groups. We found nonsignificant differences between the SNI and sham groups for other peptides (dynorphin A, neurokinin A, substance P, calcitonin gene–related peptides [CGRP] 1 and 2).
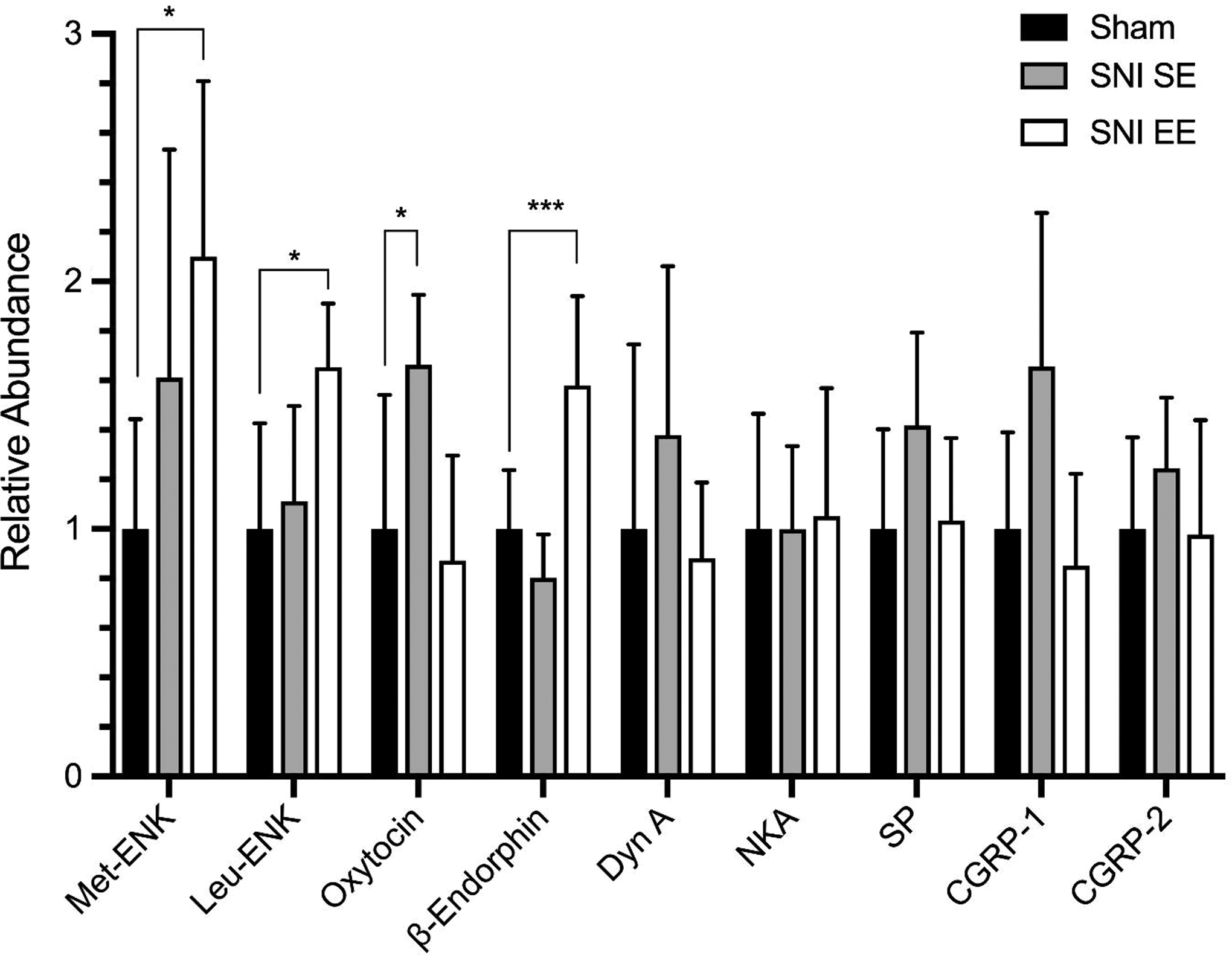
Selected peptide results for female mice presented as normalized data based on the peak mean areas for the sham group for relative quantification. CGRP = calcitonin gene–related peptides; Dyn A = dynorphin A; met-ENK = met-enkephalin; leu-ENK = leu-enkephalin; NKA = neurokinin A; SNI-EE = spared nerve injury surgery + environmental enrichment group; SNI-SE = spared nerve injury surgery + standard environment group; SP = substance P. *p < .05; ***p < .001 when comparing SNI-EE to Sham for β-endorphins. No significant differences were found among sham, SNI-SE, and SNI-EE mice for dynorphin A (Dyn A), neurokinin A (NKA), substance P (SP), calcitonin gene–related peptides (CGRP) 1 and 2.
Gestation Duration and Assessments of Pups at Birth and on Days 3 and 6
The duration of gestation differed significantly among the groups (Table 1), F(3, 28) = 5.24, p = .0054). The post hoc Tukey test revealed that the only significant difference in gestational duration was between sham EE and sham SE groups (p < .01). All gestational durations were within normal expected values.
Gestation Duration, Number of Pups at Birth, and Total Number of Pup Deaths Per Group.

Note. EE = environmental enrichment; SE = standard environment; sham = sham surgery; SNI = spare nerve injury surgery; SEM = standard error of mean.
*p < .05 for comparison between SNI-SE and all other groups. **p < .01 for comparison between sham SE and EE groups.
The number of pups per group differed significantly among groups (Table 1), F(3, 28) = 3.58, p = .023). The post hoc Tukey test revealed that the number of pups in the SNI-SE group was significantly smaller (p < .05) than that in all other groups. There was a significantly higher rate of deaths at birth in the SNI-SE group (n = 6) because no deaths occurred in the other experimental groups. We observed no physical anomalies in pups in any of the groups.
At birth (Day 0), the weight, F(3, 27) = 1.30, p = .29, and length, F(3, 27) = 0.85, p = .48, of the pups did not differ statistically among the groups.
Table 2 presents the results of the behavioral tests we performed on the pups. On Day 3 postdelivery, we found no significant differences among groups for negative geotaxis (χ2 = 2.6276, p = .45), grasping (χ2 = 6.4535, p = .092), rooting (χ2 = 5.9601, p = .11), or crawling (Table 2; χ2 = 1.7338, p = .63). Also, we found no significant difference in the time for crawling between groups, F(3, 27) = 0.44, p = .73. However, the righting reflex differed significantly in the pups from mothers in the SNI-SE group compared to all other groups (χ2 = 13.4131, p = .0038). Odds ratio calculation demonstrated that the chances of observing righting were significantly higher in the SNI-EE group compared to the SNI-SE group by a factor of 3.2. Crawling times also did not differ significantly among groups on Day 6, F(3, 27) = 1.74, p = .18).
Percent of Mouse Pups in Each Group That Successfully Completed the Behavioral Tests on Day 3 Postbirth.
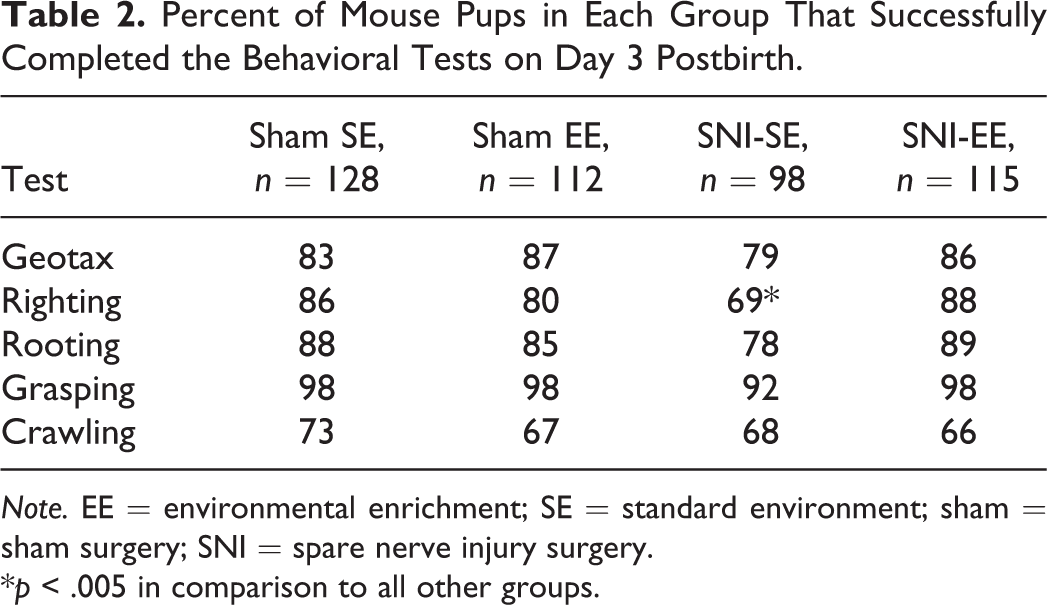
Note. EE = environmental enrichment; SE = standard environment; sham = sham surgery; SNI = spare nerve injury surgery.
*p < .005 in comparison to all other groups.
Discussion
Results of the present study show that exercise with the running wheel significantly alleviated chronic pain induced by SNI surgery in female pregnant mice. Animals that underwent sham surgery showed no significant changes in mechanical sensitivity regardless of exercise condition. Following delivery, females in the SNI-SE group still demonstrated significantly increased pain compared to baseline and the sham groups, whereas females in the SNI-EE group demonstrated no significant difference from baseline or the sham groups. These results suggest that exercise could alleviate chronic neuropathic pain during pregnancy. Importantly, exercise performance did not vary between the sham and SNI females, although the level of exercise, measured by the number of wheel turns per hour over 24 hr, decreased for all groups during pregnancy, which was expected (Eclarinal et al., 2016). The duration of gestation varied significantly only between the two shams, with females in the sham EE group having a mean length of gestation that was 1 day longer than that of the sham SE group. All gestation durations were within the normal range for mice. Weights of mothers did not differ among groups throughout the experiment. These results clearly differ from those of a previous study (Zhou et al., 2018), where pain induced by an inflammatory model of injecting complete Freund adjuvant (CFA) in the posterior hind paw of mice was associated with shorter gestation periods and decreased weight in the mothers. These changes could be attributed to the severity of the model since the CFA injections caused significant swelling and pain for up to 28 days (Zhou et al., 2018), as compared to the standard CFA protocol, which, in previous studies, caused inflammatory swelling that is maximal at 24 hr and persists up to 7 days (Iadarola, Brady, Draisci, & Dubner, 1988; Stein, Millan, & Herz, 1988). The SNI model, apart from a partial motor paralysis, does not cause any significant swelling in the hind paws over time.
Results also show that live pups were similar in many aspects across groups. At birth, they did not differ in weight or length among groups and presented no gross physical anomalies. However, the number of pups was significantly less for the SNI-SE group. There were also mortalities following delivery in this group, while there were none for any of the other groups. These results differ somewhat from those of a previous study (Zhou et al., 2018), where pups were shorter in length and weighed less at birth in the group that received CFA. These results suggest that the implantation and/or development of embryos is altered with chronic or inflammatory pain and that exercise can reverse this.
Behavioral tests in pups (geotaxis, righting, rooting, grasping, and crawling) at 3 and 6 days postbirth showed no significant differences among groups except for the righting reflex, where a significantly smaller proportion of pups in the SNI-SE group were able to perform the test correctly compared to the other groups. We have no explanation at the present time to explain why we found a significant difference for the righting test only. These results differ from the results of Zhou et al. (2018), who concluded that pups showed a deficit in motor development following CFA injection in mothers who were provided with no EE. In the present study, we performed a more extensive battery of tests and found that the pups were much less affected by the neuropathic pain of their pregnant mothers than we expected. Importantly, both the present study and Zhou et al. (2018) tested only motor behaviors and did not evaluate cognitive development at all. Future studies should longitudinally examine brain development and cognitive abilities in adulthood in order to explore possible effects on pups later in life of neuropathic or chronic pain in mothers during pregnancy.
We suggest that cytokines and the endocrine system explain our findings in the present study regarding the reproductive effects of neuropathic pain and exercise on reproduction. We further suggest that central nervous system modifications explain our findings regarding the ability of exercise to alleviate neuropathic pain in pregnant female mice.
Cytokines comprise interleukins (ILs), tumor necrosis factor (TNF), chemokines, and interferons (IFNs). Some are proinflammatory, others anti-inflammatory (Beaulieu, Lussier, Porreca, & Dickenson, 2010). During gestation, a number of cytokines play a key role in normal placental physiology such as the regulation of placental hormones. Production of chorionic gonadotropin is stimulated by IL-1β (Masuhiro et al., 1991; Seki, Zosmer, Elder, & Sullivan, 1997; Yagel, Lala, Powell, & Casper, 1989) and TNFα (Li et al., 1992). Cytokines also participate in the reproductive tissue immune response during gestation (Zdravkovic et al., 1997). IFNs derived from the trophoblasts and TNFα can interfere with viral replication (Franco et al., 1999; Paradowska, Blach-Olszewska, Sender, & Jarosz, 1996). Cytokines also play a role in myometrial contractility, amniotic membrane rupture, and different gestational pathologies such as preeclampsia and intrauterine growth retardation (Bowen, Chamley, Keelan, & Mitchell, 2002). Cytokines can also have deleterious effects on the fetus when they cross the placenta. Following a maternal infection, elevation of cytokines and chemokines can cause neurodevelopmental damage in the fetus, such as cerebral paralysis (Yoon et al., 1990, 2000). However, the contributions of the viral agent and the immune response are not clear. With neuropathic pain, cytokine levels correlate with pain intensity (Koch et al., 2007). We suggest that the SNI model caused inflammation only at the beginning of gestation, which could be related to the decrease in the number of live pups in the SNI-SE group, and because exercise decreases inflammation (Beavers, Brinkley, & Nicklas, 2010), it could also explain the normal reproductive performance of SNI-EE mice.
The endocrine system could also be involved in processes that would explain our reproductive findings (Tennant, 2013). Chronic pain is associated with stress, which stimulates the hypothalamus and pituitary glands, which, in turn, secrete trophic factors that cause the release of adrenal and gonadal hormones (Griep et al., 1998; Moore, Evans, Smith, & Loyd, 1983; Neeck, Federlin, Graef, Rusch, & Schmidt, 1990; Strittmatter et al., 2005). Chronic pain, without immediate relief, may cause elevated serum levels of adrenocorticotropic hormone (ACTH) from the pituitary, cortisol from the adrenals, and testosterone from the gonad glands. If no relief is provided for an extended period of time, various hormones may become depleted (Griep et al., 1998; Neeck et al., 1990; Strittmatter et al., 2005; Tennant, 2013). Serum levels of ACTH from the pituitary, pregnenolone from the adrenals, and progesterone from the gonads may decrease. These hormonal changes can impede embryo implantation (Horne, White, & Lalani, 2000). Decreased levels of estrogen, progesterone, and testosterone may also affect the mothers (Vincent & Tracey, 2008). In adults, estrogen is associated with the availability of opioid receptors and a decrease in anxiety and stress, progesterone with analgesia and sedation, and testosterone with analgesia and stress. All these factors could explain, in part, the increased pain sensitivity in the SNI-SE animals. Previous studies support our results with findings demonstrating that exercise can prevent pregnancy complications and reestablish hormonal balance (Davenport et al., 2019).
In previous studies, we showed that some pain peptides (substance P and CGRP) are upregulated in the spinal cord of CD-1 male mice with neuropathic pain and downregulated with EE (Vachon et al., 2013). The present results with pregnant females clearly show that enkephalins and β-endorphins were increased with exercise in the frontal cortex of SNI-EE compared to SNI-SE females. Interestingly, OT levels in the frontal cortex were increased in the SNI-SE group and decreased in the SNI-EE group. We saw no significant differences for DynA, SP, NKA, or CGRP (1 and 2) peptides between groups.
OT-producing neurons project to different areas of the brain, such as orbitofrontal, cingulate, insular, and medial prefrontal cortices, which all play a role in pain perception (Vargas-Martinez, Uvnäs-Moberg, Petersson, Olausson, & Jimenez-Estrada, 2014). The medial prefrontal cortex, in particular, is involved in chronic pain elicited in a spared nerve injury rat model (Metz, Yau, Centeno, Apkarian, & Martina, 2009). No study has assessed direct OT effects on cortical pain processing; however, OT is an analgesic at the levels of the brain stem and spinal cord (Boll, Almeida de Minas, Raftogianni, Herpertz, & Grinevich, 2017). Our present results suggest that frontal cortex OT levels could be an indicator of pain. Research has shown both endorphins (Gu et al., 2017) and enkephalins (Henry, Gendron, Tremblay, & Drolet, 2017) to play an important role in pain analgesia, and both peptides can be found in the medial prefrontal cortex. Interestingly, research has also shown that enkephalins play a role in resilience in response to chronic stress (Henry et al., 2017), which may accompany chronic pain and would therefore also be important in animal welfare. Pregnant mice in the SNI-EE group in the present study ran extensively on the running wheels. Previous research has shown that opioidergic mechanisms in the cerebrum correlate with exercise-induced mood changes sometimes known as the runner’s high (Boecker et al., 2008). Therefore, our findings on enkephalins, endorphins, and OT correlate well with pain alleviation of neuropathic pain with exercise.
Our results show that exercise can reduce neuropathic pain in pregnant CD-1 female mice. We did not find that neuropathic pain impacts the neurodevelopment of mice pups, but our results suggest that it could impact live births. With replication of this study and exploration of these findings in humans, these findings could have implications for health care. For example, nurses could develop a pain management plan for pregnant women with neuropathic pain that involved regular exercise to reduce hyperalgesia without increasing the risk to the developing fetus.
Footnotes
Acknowledgments
The authors would like to acknowledge Guy Beauchamp (Faculty of Veterinary Medicine, University of Montreal) for statistical analyses on all results (except the peptide results) as well as Marie-Thérèse Parent for the preparation of the figures.
Author Contributions
Madeleine Parent-Vachon contributed to conception, design, acquisition, and interpretation; drafted and critically revised the manuscript; and gave final approval. Francis Beaudry contributed to acquisition, analysis, and interpretation; drafted and critically revised the manuscript; and gave final approval. Denise Carrier contributed to acquisition, critically revised the manuscript, and gave final approval. Graziella di Cristo contributed to conception and design, critically revised the manuscript, and gave final approval. Pascal Vachon contributed to conception, design, acquisition, analysis, and interpretation; drafted and critically revised the manuscript; and gave final approval. All the authors agree to be accountable for all aspects of work ensuring integrity and accuracy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Fond de Développement pour la Médecine des Animaux de Laboratoire (P. Vachon). The mass spectrometry analyses were performed using an infrastructure and operation funded by the Canadian Foundation for Innovation and the Fonds de Recherche du Québec, Government of Quebec (F. Beaudry CFI, John R. Evans Leaders Grant No. 36706).