
Abstract
Psychosocial stress is increasingly apparent in children and can affect many areas including immune responses in childhood (Danese & Lewis, 2017). According to Liu and colleagues (2017), “stress is a state of threatened homeostasis provoked by a psychological, environmental, or physiological stressor” (p. 1). The ability to mitigate psychosocial stress in childhood is of utmost importance to avoid negative health outcomes that can extend into adolescence and adulthood (Carlsson et al., 2014; Danese & Lewis, 2017; O’Connor et al., 2020). Specifically, psychosocial stress in childhood has been linked to physiological stress responses, cardiovascular disease, type 2 diabetes, cancer, obesity, metabolic syndrome, inflammation, immune dysregulation, mental illness, and poor cognitive development (Danese & Lewis, 2017; Garner, 2013; G. E. Miller et al., 2021; Nusslock & Miller, 2016; O’Connor et al., 2020).
Early life stress and exposure to ongoing stressors in childhood, such as maltreatment, have also been associated with inflammation in adolescence and adulthood (Danese & Lewis, 2017; O’Connor et al., 2020). Kotas and Medzhitov (2015) define inflammation as “a protective response to extreme challenges to homeostasis, such as infection, tissue stress, and injury” (p. 823). Because of the detrimental effects physiologic stress and inflammation may have on health outcomes in childhood and later in adulthood, clinically relevant markers of chronic physiologic stress and chronic inflammation need to be assessed in studies of children.
Chronic physiologic stress is described as the prolonged release of primary mediators, such as glucocorticoids, catecholamines, and cytokines, in response to any stressor (Condon, 2018). Chronic stress increases the risk for negative secondary outcomes (Condon) including chronic inflammation (Liu et al., 2017). Chronic inflammation is slow, long-term inflammation with continued exposure to social, psychological, environmental and/or biological factors over extended periods of time that may cause prolonged physiologic stress and inflammatory responses and result in negative secondary outcomes (Furman et al., 2019).
Despite the known effects of stress and inflammation in childhood on adult health, there are unique challenges to examining chronic stress and inflammation in children. Clinicians and researchers must have the ability to accurately and reliably measure chronic stress and inflammation in children. This is especially true as children are exposed to ongoing stressors related to the COVID-19 pandemic and long COVID symptoms with chronic inflammation (Katella, 2021; Younger, 2021). Therefore, the purpose of this paper is to briefly review stress and inflammatory pathways, identify biomarkers used to measure chronic stress and inflammation particularly in clinical and community settings, and to discuss methodological considerations when measuring stress and inflammation in children.
Stress Pathways
During times of stress the body must adapt physiologically to cope and maintain homeostasis. The term allostasis has been used to describe this adaptation as the body produces mediators, such as cortisol, to maintain homeostatic functions (McEwen, 2007; 2017). Exposure to an external or perceived stressor primarily activates 2 stress pathways to maintain homeostasis: the sympathetic nervous system (SNS) via the autonomic nervous system (ANS), and the hypothalamic-pituitary-adrenal (HPA) axis. These 2 stress pathways are intertwined and complementary to each other.
The SNS is activated when the hypothalamus triggers the adrenal glands through the ANS and leads to the release of epinephrine. This stress pathway is the sympathetic-adrenal-medullary pathway. Epinephrine increases heart rate, blood pressure, and respiratory rate, and also triggers the release of glucose. Typically, the SNS stress response, also known as the “flight or fight” response, is an acute stress response, and is accompanied by activation of the HPA axis (Evans et al., 2013).
Relatedly, stressors activate the HPA axis through secretion of corticotropin-releasing hormone (CRH) from hypothalamic paraventricular nucleus neurons (Herman et al., 2016). CRH triggers the pituitary gland to release adrenocorticotropic hormone (ACTH), which then causes release of glucocorticoids (GCs) from the adrenal glands. The primary GC hormone released by the HPA axis in humans is cortisol, and activation of the HPA axis is dependent on positive and negative feedback loops with high levels of cortisol inhibiting ACTH secretion and low levels of cortisol stimulating ACTH secretion. Secretion of GCs leads to hepatic glycogenolysis and lipolysis, and results in increased blood glucose levels. GCs released by the HPA axis typically have immunosuppressive and anti-inflammatory effects, and short-term exposure to stressors typically results in protective adaptations. However, long term exposure to stressors with chronic stress and chronic activation of the HPA axis can lead to maladaptation thus resulting in tissue damage and negative health outcomes (Bellavance & Rivest, 2014; McEwen, 2007). Often this maladaptation is termed allostatic load which has been described as a tenuous balance between protective and damaging effects of these physiologic stress mediators (Juster et al., 2010; McEwen, 2017). Maladaptive responses often result in over and under production of stress mediators which may be measured with biomarker analysis (Juster et al., 2010).
Inflammatory Pathways
When there are threats to homeostasis, inflammation is a protective response that causes the release of inflammatory mediators, such as cytokines and chemokines, and results in local and systemic changes to maintain homeostasis (Kotas & Medzhitov, 2015; Netea et al., 2017). These inflammatory mediators, including IL-6 and TNF-α, can either stimulate or stop the various homeostatic systems and are both dominant and antagonistic to signals of homeostasis. Inflammatory responses are successful when they are followed by restoration of homeostasis. However, because inflammation occurs when homeostasis is disrupted and it can override homeostatic processes, inflammation has the potential to become locked and thus becomes chronic when it does not resolve (Kotas & Medzhitov, 2015).
Inflammation is dependent on macrophage and neutrophil infiltration as they are the 2 main sources of inflammatory mediators (Kotas & Medzhitov, 2015). Along with these, several peripheral mediators of the inflammatory response have the ability to fluctuate availability and abundance of immune cells in response to stressful stimuli. One such acute mediator is damage-associated molecular patterns which are released in response to physical, chemical, or metabolic noxious stimuli (Furman et al., 2019). However, chronic stress has the ability to deplete macrophages thus making the body more susceptible to chronic inflammation (Barrett et al., 2021). For example, sub-acute chronic stress boosts humoral immunity (producing antigen-specific antibodies) while simultaneously suppressing cell-mediated immunity (not dependent on antigens). This balance of pro- and anti-inflammatory effects is mediated by glucocorticoid effects on the T-helper 1 and T-helper 2 that stimulate or inhibit the production of cytokines (Elenkov & Chrousos, 1999).
Pathways Between Stress and Inflammation
Evidence to link stress and stressful experiences to inflammation, especially relationships between stress in childhood and inflammation in adolescence and adulthood, is plentiful (Danese & Lewis, 2017; O’Connor et al., 2020). According to the neuroimmune network hypothesis (Miller et al., 2021), early life stress activates a positive feedback loop between networked brain regions and peripheral inflammatory cells. While parallel activation of stress and immune pathways occurs in response to physical or psychological threats (Kautz et al., 2020), neurotransmitters, neuropeptides, and hormones secreted by sympathetic neurons and/or the adrenal medulla also activate receptors found on immune cells (Carlsson et al., 2014). Activation of immune cells is mediated through various multidirectional feedback mechanisms. Acute stressors result in advantageous adaptations by the immune system, whereas chronic stress can be detrimental to the immune system with either suppressed or exaggerated immune responses (Carlsson et al., 2014).
The relationship between stress and inflammation is thought to be bidirectional between the neuroendocrine and immune systems, with the same neuropeptides mediating both stress and inflammation (Black, 2002; Liu et al., 2017). In normal conditions, stress and inflammatory pathways work together to maintain homeostasis. Through the cytokine-glucocorticoid feedback cycle, proinflammatory cytokines increase in response to physical or mental stressors, and in turn these cytokines increase circulating levels of cortisol (Corwin & Ferranti, 2016). This circulating cortisol binds to receptors of inflammatory cells, such as white blood cells, and block production of proinflammatory cytokine transcription factors, thus resulting in decreased cytokine response and maintenance of homeostasis (Corwin & Ferranti, 2016).
Chronic inflammation may be attributed to hormonal changes that characterize the stress response, while the HPA axis may be activated via inflammatory stimuli (Black, 2002). Under normal physiologic conditions, physiological stress responses would not activate chronic inflammatory responses. However, during times of chronic stress response and dysregulation, stress pathways activate the immune response and result in the chronic release of and exposure to inflammatory substances (Liu et al., 2017). In response, prolonged exposure to inflammatory substances may act as a chronic stressor, further perpetuating the relationship between chronic stress and inflammation. Similarly, when the immune system is activated, there may also be increased stress responses through SNS pathways and reduced corticosteroid responsiveness by inflammatory cells (Corwin & Ferranti, 2016; Liu et al., 2017). Without resolution in stress or inflammation, negative health outcomes such as metabolic syndrome, hypertension, dyslipidemia, hyperglycemia, obesity, psychopathologies, cardiovascular disease, cancer and chronic kidney disease may be noted (Danese & Lewis, 2017; Furman et al., 2019).
Resolution of Inflammation related to Stress
Resolution of inflammation and stress pathways are of utmost importance to maintain homeostasis and avoid negative health outcomes associated with prolonged exposure. Resolution of stress is initiated by the removal of the stressor. Conversely, resolution of acute inflammation is an active process with the reprogramming of cells that involves the increased consumption of neutrophils and other apoptotic granulocytes by macrophages to clear tissue debris (Netea et al., 2017). This is coupled with the ‘turning-off’ of chemoattractant signals to decrease leukocyte chemotaxis to the site of injury and increase in the production of ‘immunoresolvants’ that begin the resolution phase (Buckley et al., 2013).
If inflammation is not moderated or resolved, it becomes chronic and is often termed ‘hyperinflammation’ (Manjili et al., 2020). This overactive response results in a blunting of the adaptive immune response and an exponential increase in cytokine production often termed a ‘cytokine storm.’ In severe respiratory diseases such as Severe Acute Respiratory Syndrome, Middle Eastern Respiratory Syndrome, and more recently, COVID-19, this is manifested by much higher levels of inflammatory cytokines, such as IL-6 and TNF-α, which also correlate with disease severity and ICU admission rate (Manjili et al., 2020). This inability to resolve the acute phase of inflammation has been postulated as the underlying mechanism of long COVID (Mehandru & Merad, 2022).
Similarly, autoimmune responses may also be related to the inability to resolve inflammation. Autoimmune disease is a chronic inflammatory condition in which an individual’s immune system attacks their own body. Autoimmune diseases cycle through sequential phases of initiation, propagation, and resolution (Rosenblum et al., 2015). Initiation, and disease severity, is dependent upon genetic predisposition and environmental triggers. While failure of regulatory mechanisms is associated with all stages of autoimmune disease, the propagation phase is characterized by self-perpetuating inflammation. The activation and proliferation of autoreactive T cells, with the inability to suppress and resolve inflammatory responses by regulatory T cells, results in tissue injury and clinical disease (Rosenblum et al., 2015).
Biomarkers of Stress and Inflammation in Children
Given the emphasis on stress in children and the resulting impact of stress on inflammation, biomarkers of stress and inflammation should be considered when examining these phenomena in research. Biomarkers used to measure stress in children may be classified as central or peripheral biomarkers. Central biomarkers of stress are chemical signals directly from the brain while peripheral biomarkers are made in surrounding tissues in response to central signals. When considering these biomarkers, it is important to make the distinction between acute stress and chronic stress since relevant biomarkers may differ.
Biomarkers related to stress may be classified in a number of ways. Alpha amylase (AA) and dehydroepiandrosterone sulfate (DHEA-S) are commonly used to measure stress, but these measures may be more representative of an acute stress response rather than a chronic stress response. For example, AA peaks within 15 minutes after an acute stressor and returns to baseline within 30 minutes (Petrakova et al., 2015). Similarly, DHEA-S also has a short-term increase of 1 hour in response to acute stressors (Dutheil et al., 2021). Therefore, these biomarkers may not be the best choice to measure chronic stress in children. However, there are several biomarkers that are more indicative of chronic stress responses, and these are classified by central and peripheral stress responses.
Central Biomarkers of Chronic Stress
Central biomarkers of stress include hormones and growth factors that are released in response to stressors. While these central biomarkers may better represent the stress response, they may be more difficult to measure in clinical and community settings. These include ACTH and brain derived neurotrophic factor (BDNF).
As previously discussed, ACTH release is stimulated by CRH in a pulsatile fashion which changes over the course of the day. This diurnal variation in ACTH levels makes measuring ACTH difficult in a community setting as levels can be affected by stressful situations (e.g., blood draws) and time (Allen & Sharma, 2022). BDNF is a growth factor that, along with 3 other related neurotrophins, are largely responsible for brain development and plasticity as an adult (Notaras & van den Buuse, 2020). BDNF’s role in the stress response is multifactorial and low levels do not definitively constitute a reduction in stress tolerance or resilience (Notaras & van den Buuse, 2020). Because the role of BDNF is multifaceted, its application in the community setting is not practical as reference ranges and intra-person variation make measurement and interpretation in outpatient cohorts difficult.
Peripheral Biomarkers of Chronic Stress
Peripheral biomarkers of stress are released after primary biomarkers of stress. These biomarkers have a secondary role in the stress response. Measuring peripheral biomarkers of stress, such as cortisol, is usually accomplished through saliva, serum, urine, hair, or nails. Therefore, these biomarkers may be easier to obtain for stress measurement particularly in the clinical and community setting.
Cortisol
When an individual encounters a stressor perceived as threatening, cortisol, the major stress hormone, is released by the adrenal glands in response to a signal from the hypothalamus (McEwen, 2007; McEwen & Wingfield, 2003). Cortisol stimulates the release of glucose, enhances the use of glucose by the brain, and inhibits or curbs nonessential functions of the immune system, the digestive system, the reproductive system, and growth processes (McEwen & Wingfield, 2003). Cortisol follows a diurnal pattern with cortisol levels peaking to the highest level in the morning just after waking and gradually following throughout the day (Miller et al., 2016). In an acute situation, the effects of cortisol diminish once the stressor has passed. However, when the stressor is chronic, the stress response is not turned off resulting in continual disruption of body systems and abnormal changes in cortisol. Eventually, the body may not physiologically respond to the chronic stress resulting in blunted levels of cortisol and increased inflammatory responses with negative health outcomes (McEwen & Gianaros, 2011; Silverman & Sternberg, 2012). Abnormal changes in cortisol can be measured by assessing hyper- and hypo-reactivity to acute stress tests, such as the Trier Social Stress Test for Children, or by collecting cortisol levels throughout the day to evaluate diurnal rhythm (Dickerson & Kemeny, 2004; Gunnar et al., 2009). While hyper- and hypo-reactivity responses to acute stress tests indicate abnormal HPA axis response, dysregulation in cortisol diurnal rhythm, such as blunting (no changed from morning to night) or increases in cortisol from morning to afternoon samples may more accurately represent chronic physiological stress (Gunnar et al., 2009; Miller et al., 2016).
Cortisol can be measured in a number of different specimens including saliva, serum, urine, hair and nails. Condon (2018) and Rodriguez and colleagues (2016), in addressing biomarkers including cortisol, discussed the advantages and disadvantages of various types of specimens to measure cortisol. Hair and nail specimens may more readily measure chronic stress as they reflect levels of cortisol over time (Bates et al., 2017; Liu & Doan, 2019) and can easily be used with infants and young children. Serum, saliva, and urine, on the other hand, can be used to measure both acute and chronic stress with varying degrees of burden related to collection as well as invasiveness (Condon, 2018).
Normal cortisol levels in children are 5–25 mcg/dL for morning specimens and 2 to 14 mcg/dl for afternoon specimens (Petersen, 1981). Values are usually lower in the afternoon based on the diurnal rhythm. However, diurnal patterns seen in adults may not be exhibited in children until the age of 4 due to daytime naps (Wilkinson & Goodyer, 2011), and afternoon levels can be higher than morning values if specimens are collected after the child has been napping (Rice et al., 2018). Values can also be affected by disease (e.g., Cushing Syndrome), by medications (e.g., Ritalin or Growth Hormone), or by food/drink intake (Stachowicz & Lebiedzińska, 2016).
Complementary measures of Chronic Stress
While cortisol appears to be the most useful and feasible measure of chronic stress in children, researchers have attempted to use other biomarkers to measure stress. For example, epinephrine and norepinephrine are commonly used to measure the stress response. Epinephrine and norepinephrine are catecholamines that are released by the adrenal glands in response to acute stressors. The hypothalamus stimulates this release as part of the fight-or-flight response. Because epinephrine and norepinephrine are more representative of the acute stress response of the SNS, they do not represent chronic stress in children (de Kloet et al., 2005; Joëls & de Kloet, 1989; Tank & Lee Wong, 2014). Similarly, secretory IgA (s-Iga) may represent a stress response because it acts as a first line of defense when epithelial tissues are exposed to harmful toxins and microorganisms (Condon, 2018). However, high levels of s-IgA may indicate an immune response to injury or infection rather than a specific response to stress.
Other biomarkers that may not be specific to the stress response but often accompany stress in children include glucose, HbA1c, triglycerides, and cholesterol levels (Noushad et al., 2021). Additionally, oxytocin and prolactin may be biomarkers that are considered indirect measures of the stress response because they are known to have protective factors in response to stress. For example, oxytocin is a major regulator of oxidative stress radicals that has anti-stress capabilities including decreasing blood pressure and cortisol levels (Jankowski et al., 2020). While these biomarkers may not directly represent the stress response in children, collection of these complementary biomarkers may help create a holistic understanding of the stress response in children.
Biomarkers of Chronic Inflammation
Common biomarkers of inflammation, such as C-reactive protein (CRP), TNF-α, and IL-6, are frequently used to measure chronic inflammation in children. While CRP and IL-6 have both anti-inflammatory and pro-inflammatory properties, IL-2 is primarily an anti-inflammatory cytokine (Banchereau et al., 2012). Similarly, while IL-1β may not be a direct measure of chronic inflammation, it is often elevated during chronic inflammatory responses. Therefore, IL-2 and IL-1β may be better considered complementary, rather than direct, measures of chronic inflammation.
C-Reactive Protein
C-reactive protein (CRP) is a plasma protein that increases during the inflammatory response and acts as a pattern recognition molecule. CRP is part of a larger change in gene expression, termed the acute phase response, which happens in response to injury and other acute inflammatory stimuli. CRP is also associated with chronic inflammation evidenced by elevations in baseline CRP levels which indicate excessive inflammation or failure to resolve inflammatory response (Pepys & Hirschfield, 2003). While context dependent, CRP has established pro-inflammatory as well as anti-inflammatory properties; however, as a biomarker its use is classically reserved for increases in inflammation giving rise to increases in the plasma (Black et al., 2004).
CRP is commonly used to measure both acute as well as chronic inflammation and inform providers on treatment efficacy as it rapidly increases 4–6 hours following injury and remains elevated while inflammation is present. However, because CRP has a half-life of only 4–7 hours, it can also be used to measure the resolution of inflammation during treatment or lifestyle interventions designed to reduce inflammation. CRP is commonly measured using nephelometry and offers sensitivity down to 0.04 mg/L. While it is readily detected in multiple bodily fluids (CSF, amniotic, pleural, synovial), CRP is still most commonly measured in the plasma although newer techniques use salivary specimens (Ouellet-Morin et al., 2011).
High-sensitivity CRP (hsCRP), an analysis to specifically detect low levels of CRP, can be measured in venous blood and salivary samples, although considerations must be made when using salivary CRP to assess chronic low-grade inflammation. The oral environment will acutely increase CRP in response to local trauma, thereby complicating the interpretation of results. Additionally, the patient dependent salivary flow, which is largely dependent on hydration status, can concentrate or dilute the salivary samples thus impacting the relative levels of CRP (Pay & Shaw, 2019).
TNF-α
Tumor Necrosis Factor alpha (TNF-α) is so named because of its original discovery as a soluble factor that was present in the crude bacterial extracts (the product initially obtained from extraction) with the ability to cause necrosis of tumor cells. TNF-α is known to be a critical mediator of both acute and chronic inflammation by inducing its own secretion to begin the inflammatory immune response (Chu, 2013). Although, TNF-α can be measured in multiple bodily fluids, the relative levels of TNF-α are, like many other cytokines, somewhat specific to the area being measured. For example, children who have inflammatory bowel disease are known to not only have increased levels of TNF-α in the serum but also in the stool (Nicholls et al., 1993). All of these measurements are generally made using traditional ELISA assays which are inexpensive, accurate, and relatively quick for use in clinical diagnostic testing. However, care must be taken when interpreting results as discrepancies have been noted when using bioassays and immunoassays in determining quantitatively the levels of TNF-α in biological fluids (Corti et al., 1994).
IL-6
Interleukin-6 (IL-6) classically plays 2 roles in mediating the immune response to stress and inflammation. During acute stress or acute inflammation, IL-6 is considered to be anti-inflammatory as its main role is to suppress the levels of several pro-inflammatory cytokines by stimulating the production of acute phase proteins (Kaplanski, 2003; Xing et al., 1998). However, if levels of IL-6 persist past the initial inflammatory reaction, they bind with the soluble IL-6 receptor and through a series of loops are able to begin the recruitment of chronic inflammatory monocytes through secretion of monocyte chemoattractant protein-1 (MCP-1). This recruitment leads to continuation of the inflammatory process leading to chronic inflammation (Gabay, 2006).
This process of chronic inflammation, especially in the pediatric population, is triggered by a number of acute and autoimmune inflammatory conditions such as juvenile idiopathic rheumatoid arthritis (Akioka, 2019). These chronic inflammatory conditions are associated with increases in circulating levels of IL-6 as well as impaired growth. A typical detection range is 0.01–0.03 pg/mL and detection in the lower end of the range offers increased sensitivity for childhood inflammatory conditions (Rivnak et al., 2015).
Complementary Measures of Chronic Inflammation
While the biomarkers discussed above reflect direct assessment of chronic inflammation, there are biomarkers that may be indirectly reflective of chronic inflammation. Among those are IL-2 and IL-1β. IL-2 is a well-known anti-inflammatory cytokine (Banchereau et al., 2012) that has been shown to be reduced in chronic inflammatory conditions such as insulin-dependent diabetes mellitus (IDDM). This reduction in IL-2 in children with IDDM demonstrates a role for IL-2 reduction as a marker of sustained chronic inflammation (Dogan et al., 2006). On the other hand, IL-1β is a well-known pro-inflammatory cytokine that acts to resolve acute inflammation by recruiting effector immune cells and initiating tissue repair (Dinarello, 2014). However, during stressful situations (such as obesity; nutritional excess) an excess of IL-1β is noted, and IL-1β receptor antagonists have had success in treating obesity associated type-2-diabetes (Larsen et al., 2007). This calls for careful interpretation of high levels of IL-1β in clinical settings as both acute and chronic inflammation can cause an increase.
Considerations for Data Collection
Biomarkers may be obtained through various mediums such as blood, saliva, hair, and urine. When obtaining specimens from children in community and school settings, saliva, hair and nails are better choices because they are less invasive to collect. However, there are unique characteristics of each medium that researchers must consider when collecting biomarkers to assess chronic stress and inflammation in children.
Blood/Serum
While venipuncture offers the ability to measure a wide array of biomarkers, it is often difficult to perform in community settings and has increased participant burden with low compliance in children. In community and school settings, blood samples are best collected by fingerstick with capillary blood. While this blood may be applied to paper as dried blood spots (DBS) or dripped into a specimen tube, new technology in microsampling, such as Neoteryx’s MITRA devices (Neoteryx, 2022), have simplified the collection of blood specimens in outpatient and community settings. DBS are relatively easy and inexpensive to collect, and the amount of blood needed to complete DBS analysis is small, requiring only 50–70 μL per DBS with 4–6 spots typically collected. However, there are some limitations in measurement with DBS. For example, concentrations of analytes in DBS can be easily influenced by patient hematocrit levels, and correlations with serum/plasma samples can vary (Garza & Clarke, 2022). Conversely, while MITRA devices are somewhat more expensive, there is less dependency on hematocrit levels and more consistency in sample collection. Like DBS, MITRA devices also collect small samples of blood: 10–30 μL per absorbent tip with 1–4 tips collected at a time (Neoteryx, 2022).
Along with the methods used to collect blood, there are specific considerations for storage and transportation of blood. If collecting capillary blood that is dripped into a tube, considerations must be made for transferring the blood to the laboratory for analysis. In certain situations, the tubes with blood samples may need to be transferred in a cooler on ice or dry ice within a certain time frame to prevent clotting and degradation of the sample. For example, some inflammatory markers (IL-6 and TNF-α) can begin to degrade after 4–6 hours at room temperature. Therefore, it is recommended to process samples immediately, or to store at them at lower than −70°C, especially if stored for an extended time (Tworoger & Hankinson, 2006). This may impact the ability and cost to mail or transport samples for analysis if necessary. Additionally, special tubes may need to be used that include compounds to prevent clotting of the blood sample which may incur additional costs. DBS and blood collected with MITRA devices are more flexible when collecting in community settings. Once dried (requires, approximately 90 minutes to 24 hours), DBS may be stored, transported or mailed at room temperature (CDC, 2017). Similarly, blood collected via MITRA devices are stable at room temperature for up to 3 weeks, require no dry time, and are able to be mailed to a laboratory for analysis (Neoteryx, 2022).
Emerging technology allows for the analysis of multiple inflammatory biomarkers from relatively small samples of blood. For example, highly sensitive multiplex immunoassay has been used to successfully measure multiple inflammatory cytokines with only 70 μL of blood (McDade et al., 2021) and multiplex data (such as cytokine panels with 25 + analytes) have been measured on high-sensitivity platforms, such as Luminex and Olink, using dried blood spots (Rosenberg-Hasson et al., 2020). Further, novel methods may promise quick results, under 30 minutes, using small volumes of blood for certain biomarkers such as IL-6 (Koyama et al., 2017). Despite these advances in technology and measurement methods, there may be limitations in what equipment is needed to adequately detect certain biomarkers from small samples of blood. For instance, equipment specifications may require specialized training to perform assays and can inflate the cost of assays and analysis.
Saliva
Saliva is relatively non-invasive to collect, making it a good medium to measure biomarkers in children. Additionally, salivary samples can be collected repeatedly in a short amount of time or spaced throughout the day. However, there are certain considerations for collecting saliva from children. Saliva may be collected using a passive drool technique in which a child drools into a tube. This collection may or may not be timed depending on if flow rate must be calculated for the specific biomarker (Salimetrics, 2022). Additionally, a cotton swab (with strings to prevent inadvertent swallowing) placed in the mouth for a certain amount of time may be used to collect saliva. This is more appropriate for very young children or for children with developmental delays who may not understand instructions for passively drooling into a tube. When collecting salivary samples, children should not eat or drink anything for 30 minutes or more before the sample is collected. It is also important to consider timing of salivary sample collection throughout the day (Szabo & Slavish, 2021). Because cortisol follows a diurnal rhythm, cortisol concentrations are often higher in morning than afternoon samples. Therefore, one must note the time that samples are collected or to collect them at specific times throughout the day to assess diurnal changes in biomarkers.
Regardless of the biomarker, children and parents must be provided with specific instructions for the collection, storage, and transport of saliva, especially if they are collecting it independently (Szabo & Slavish, 2021). Once collected, saliva may be transported at room temperature, or may need to be transferred in a cooler on ice or dry ice and stored in a freezer until analysis. This is dependent on the specific biomarker that is being examined (Salimetrics, 2022) and may increase costs to collect, store, and transport samples. For example, if collecting saliva to measure cortisol levels, it is recommended that these samples are transferred on ice and stored in a −80°C freezer until analysis. This may not be feasible for collection in all community samples.
There are limits to which inflammatory markers are measurable in salivary samples (Salimetrics). While cortisol levels in saliva correlate with serum levels (Ma et al., 2022), this is not necessarily true for other biomarkers. For example, inflammatory biomarkers from saliva may represent local rather than systemic inflammation in individuals with poor oral health (Szabo & Slavish, 2021). However, other biomarkers collected from saliva, such as CRP, may more accurately represent systemic inflammation because it is produced in the liver and enters saliva through passive diffusion and filtration (Szabo & Slavish, 2021).
Hair
Hair is an emerging non-invasive collection technique that is feasible for community data collection. While somewhat new, hair is most commonly collected to measure cortisol and represents total cortisol concentrations over the last 3–4 months (Wright et al., 2015). When analyzing hair biomarkers, it is necessary to standardize the amount of hair that is used for assays. Generally, 1 cm of hair equates to 1 month of growth (Gonzalez et al., 2019; Wright et al., 2015). Therefore, assays will provide an average value of a biomarker over a certain period of time depending on the length of hair (Stress Measurement Network, 2018).
In order to collect this specimen, hair is often cut from the back of the scalp approximately 2 cm below the cranial bone, taped or secured to aluminum foil, and transferred to the lab for analysis (Stress Measurement Network, 2018). The amount of hair collected is approximately half the diameter of a pencil. Usually, a minimum of 3 cm in length is needed for analysis. It is important to note which end of the hair collected was closest to the scalp for assay purposes. When collecting hair samples, researchers should assess for frequent hair washing and recent coloring, bleaching, or processing since these may affect biomarker concentrations in hair (Gonzalez et al., 2019). Parental concern about cutting hair from their child may also limit the use of this specimen. Hair is stable at room temperature, so there are often no significant considerations for transport; however, samples should be stored in a dark, dry location.
Nails, Urine, Tears, and Sweat
While tears are an emerging method to collect a variety of inflammatory biomarkers (Hagan et al., 2016), they may not be applicable to children in community settings due to invasiveness of collection (Bachhuber et al., 2021; Ku et al., 2020). For example, a contact lens, capillary tube, or strip may need to be placed on the eye or very close to the eye to collect a sufficient sample for analysis. Along with tears, a recent development has been the use of sweat to measure cortisol. Innovations in biomarker collection have led to the development of patches with wireless sensors that detect cortisol in sweat (Parlak et al., 2018). These patches exhibit strong correlations between sweat and serum cortisol levels. While this is a non-invasive method that could allow for the detection of diurnal changes in cortisol in community settings, to our knowledge these patches have not been used with children. However, urine and nail samples have been used to evaluate biomarkers in children, albeit not as commonly as other methods.
Urine may be used to measure select stress and inflammatory biomarkers, such as cortisol and TNF-α, but stability of inflammatory biomarkers may fluctuate over time (Schenk et al., 2019). Additionally, there may be wide fluctuations between samples depending on how and when the urine is collected (Thomas et al., 2010). There are multiple methods to collect urine: over 24 hours, first morning samples, and spot collections (Thomas et al., 2010). Urine degrades at room temperature so refrigeration and/or freezing of samples is necessary, and this may pose extra challenges when collecting 24-hour urine samples. While it may be possible to collect samples of urine at home with established protocols (National Health and Nutrition Examination Survey, 2009) and with parental assistance, urine may be difficult to collect, store, and transport in community settings, and thus may not be the most feasible option to measure biomarkers in children. In contrast, it may be more feasible to collect, store, and transport nail samples from children in community settings. Once collected, nail clippings are easy to store and transport in community settings. Like hair samples, determining the rate of nail growth is imperative to establish biomarker concentration rates (Liu & Doan, 2019). However, it may take longer to achieve adequate nail growth to collect a sufficient sample for testing. Additionally, there have been relatively few stress and inflammatory biomarkers found in nail samples.
Other Considerations for Collecting Biomarkers
Cost must be considered when choosing which biomarkers to collect because the cost can vary based on biomarker, sample, and access to laboratories with high-sensitivity multiplex platforms used for analysis. The child’s age and developmental level must also be considered when choosing the methods of biomarker collection. Although non-invasive methods have emerged to measure biomarkers, the child must be a willing and cooperative participant. Age and developmentally appropriate instructions should be given to the child and parent to ensure that samples are collected according to specifications. When interpreting the results, remember that previous research demonstrates that there may be differences in biomarker responses between male and female children (Casimir et al., 2010). Further, it is essential to consider the specimen used to collect the biomarker. The reliability among biomarkers collected from different mediums may vary based on collection and analysis methods.
Next Steps for Research
There are emerging biomarkers to consider when examining stress and inflammation in children. While these biomarkers have been used extensively in adult research, their use in child-centered research has been limited. However, there are several biomarkers that may be applicable to stress and inflammation in children. These include leptin, adiponectin, and chemokines (Rivera et al., 2019). Additionally, it would be extremely beneficial to standardize biomarker levels in children, especially in consideration of age. This would help by providing objective measurement references in which researchers could compare stress and inflammatory levels across studies. Along with standardizing biomarker levels, it would also be helpful to calculate an allostatic load index based on immune biomarkers as well as cardiovascular and metabolic biomarkers (Liu et al., 2021). While allostatic load indices have been formulated for adults (Guidi et al., 2021; Juster et al., 2010; Sabry et al., 2020), there is less work on measurement of allostatic load in children. Lastly, a better understanding of stress and inflammatory biomarkers in children would benefit interventional studies. Researchers could use biomarker levels to develop and evaluate interventions targeted at reducing systematic stress and inflammatory responses in children, and, thus, ultimately improve overall health outcomes.
Conclusion
Strengths and Limitations in Measurement of Biomarkers for Chronic Stress and Inflammation in Children.
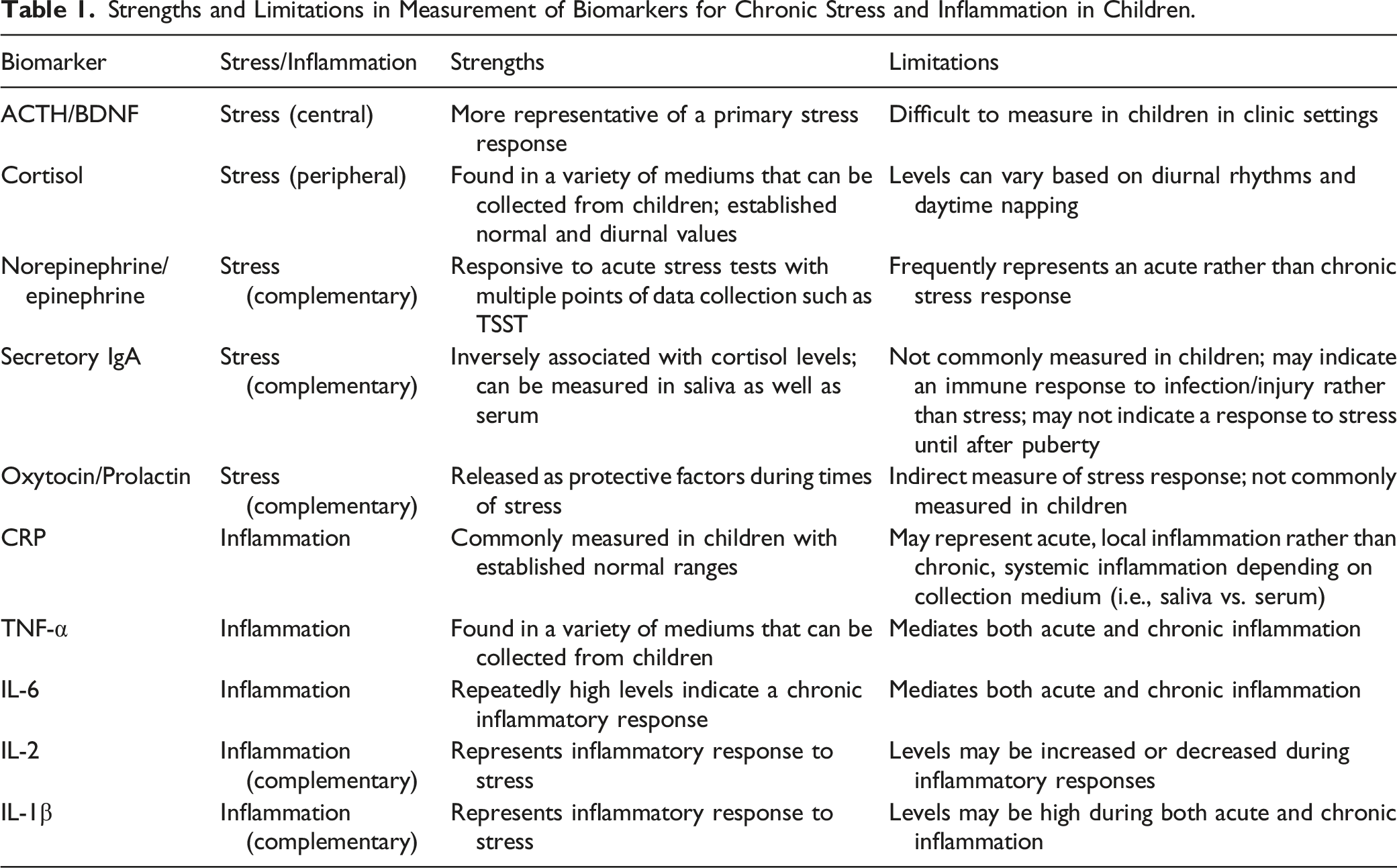
Stress biomarkers can be both centrally or peripherally derived; however central biomarkers, such as ACTH and BDNF, are difficult to obtain in the clinical or community settings. Thus, peripheral biomarkers, such as cortisol, are more easily obtained and can involve blood, saliva, urine, hair or nail specimens. Indirect biomarkers of stress, such as oxytocin and prolactin, can also be obtained to display a holistic understanding of the stress response in children. Along with stress biomarkers, common biomarkers of chronic inflammation in children, such as CRP, TNF-α and IL-6, can be measured through both blood and saliva. CRP and IL-6 have both anti-inflammatory and pro-inflammatory properties and are considered direct measures of chronic inflammation. Complementary or indirect measures of chronic inflammation such as IL-2 and IL-1β can also be measured to give a more complete picture of the inflammatory process.
Methodological considerations such as type of specimen, storage issues, and types of assays may make measuring biomarkers for stress and inflammation more problematic. Newer approaches to the collection and analysis of biomarkers, such as MITRA devices and high-sensitivity multiplex platforms, may make measurement of these biomarkers possible in children. Efforts to measure and intervene on chronic stress and inflammation in children is critical to preventing negative health outcomes both in children and adults.
Footnotes
Author’s Contribution
All authors contributed to the conception and design of the methodological review paper, preparation of the manuscript, and final editing and approval of the article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Research reported in this publication was supported by the National Center for Advancing Translational Sciences of the National Institutes of Health under award number KL2TR003097. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.