
Abstract
About one-quarter of women diagnosed with gynecologic cancer experience depressive symptoms. While the precise mechanism remains unclear, little is known about the association between gut microbiota and depressive symptoms in gynecologic cancer. Thus, this study aimed to evaluate the associations between gut microbiota and depressive symptoms in women with gynecologic cancer over cancer treatment. Thirty-seven women with cervical or endometrial cancer were followed at pre-treatment (T0), 6–8 weeks (T1), and 6 months post-radiation (T2). Depressive symptoms were assessed using the Patient Health Questionnaire-9 (PHQ-9). Rectal swabs were collected at each visit and sequenced for the V4 region of the 16S rRNA gene. MaAsLin2 models evaluated cross-sectional associations between gut microbial taxa and depressive symptoms at each time point, whereas GEE models assessed longitudinal associations over the course of cancer treatment. The patients had an average age of 60 years, and 43% were Black. At baseline (T0), 24% of patients exhibited depressive symptoms, which decreased to 21% at T1 and further to 13% at T2. GEE models showed that lower α-diversity (Shannon index, p = 0.05), dissimilar β-diversity (Bray-Curtis distance, p = 0.02), and reduced abundance of the genus Ruminococcus (p = 0.02) were predictive factors associated with depressive symptoms throughout cancer treatment. Higher depressive symptoms were longitudinally associated with lower gut microbial Shannon diversity, dissimilar microbial community composition, and lower abundance of the genus Ruminococcus. Larger longitudinal studies using shotgun metagenomic sequencing are needed to validate these findings and further elucidate the microbial mechanisms underlying depressive symptoms in women with gynecologic cancers.
Introduction
In the United States, approximately 116,930 women are diagnosed with gynecologic cancer annually (Siegel et al., 2024). Significant advancements in diagnosis and treatment have improved the overall survival rates for these patients (Barandouzi et al., 2023). By 2030, the number of gynecologic cancer survivors in the United States is expected to exceed 1.61 million (Salani et al., 2026). However, these survivors often face a myriad of persistent symptoms, including depressive symptoms (Barandouzi et al., 2023). Depressive symptoms are one of the most frequently reported and prevalent issues among gynecologic cancer patients (Lee et al., 2023). Research shows that between 13% and 32% of cancer patients experience depressive symptoms both during and after their treatment (Klapheke et al., 2020; Watts et al., 2015). Moreover, studies have found that the rate of depressive symptoms is higher in women with gynecologic cancers compared to those with other cancer types (Klapheke et al., 2020). The burden of depressive symptoms can result in a significant decrease in quality of life in cancer patients (Yu et al., 2020). Therefore, understanding the mechanisms underlying depressive symptoms in women with gynecologic cancers is crucial to addressing this issue.
While the precise mechanisms underlying depressive symptoms in cancer patients are not fully elucidated, cancer treatments such as radiotherapy and chemoradiation can induce cancer-related toxicities, including depressive symptoms (Yusof et al., 2016). Cancer treatments may alter the composition of the gut microbiota and disrupt the gut-brain axis. The gut-brain axis is a complex communication network encompassing neural, immune, and endocrine pathways (Song & Bai, 2021). Specifically, cancer treatments can alter the composition of bacteria in the gut by reducing beneficial bacteria, decreasing bacterial diversity, and contributing to dysbiosis (Barandouzi, Eng et al., 2023; Maddern et al., 2023). Dysbiosis of the gut microbiome may lead to a reduction in short-chain fatty acids (SCFAs) such as butyrate, acetate, and propionate, which are critical for maintaining gut health, immune function, and preserving the integrity of the epithelial barrier (Al-Qadami et al., 2022; Maddern et al., 2023; Morrison & Preston, 2016). Reduced SCFAs compromise tight junction integrity, increasing gut permeability and allowing bacterial products, such as lipopolysaccharide (LPS), to translocate into systemic circulation. This triggers immune activation, increases oxidative stress, and promotes the expression of proinflammatory cytokines, which can disrupt neural activity and contribute to depressive symptoms in cancer patients (Barandouzi, Bruner et al., 2023; Ghosh et al., 2020; Kelly et al., 2015; Maddern et al., 2023).
Growing clinical studies also suggest that gut microbiome dysbiosis is associated with depressive symptoms and broader psychoneurological symptom burden in cancer patients, although the literature remains methodologically limited and largely exploratory. Cross-sectional studies in rectal and breast cancer populations have consistently reported associations between depressive symptoms and alterations in gut microbial diversity and composition. In rectal cancer patients undergoing chemoradiation, higher depressive symptom scores were associated with lower abundances of several taxa, including Coprobacter, Intestinibacter, Lachnospiraceae, Intestimonas, Phascolarctobacterium, Ruminiclostridium, Parasutterella, Tyzzerella, and Ruminococcaceae, several of which belong to microbial groups involved in SCFAs production and immune regulation (Gonzalez-Mercado et al., 2021). Similarly, breast cancer patients with depressive symptoms demonstrated lower gut microbial diversity and higher relative abundances of Proteobacteria and Escherichia-Shigella compared to non-depressed breast cancer patients (Maitiniyazi et al., 2022). Additional evidence in breast cancer patients undergoing chemotherapy showed altered gut microbiome composition characterized by lower abundance of the beneficial mucin-degrading genus Akkermansia compared to healthy controls, alongside higher depressive symptoms and cognitive dysfunction (Bilenduke et al., 2022). Longitudinal evidence, although limited by small sample sizes, further supports a relationship between the gut microbiome and psychoneurological symptoms across the cancer treatment trajectory. In head and neck cancer patients receiving radiotherapy, individuals with higher psychoneurological symptom burden exhibited greater decreases in microbial evenness and distinct microbial community composition over time, including higher abundance of Ruminiclostridium9, Tyzzerella, Eubacterium_fissicatena, and DTU089 (Bai et al., 2020). Similarly, among women with gynecologic cancers receiving radiotherapy or chemoradiation, higher psychoneurological symptom burden, including depressive symptoms, was associated with lower microbial diversity and reduced abundances of Fusicatenibacter and Ruminococcus over time (Barandouzi, Eng et al., 2023).
The limited number of longitudinal studies examining the impact of cancer treatment on treatment-related toxicities, including depressive symptoms, and the potential role of the gut microbiome in these relationships underscores the need for further research. Therefore, this longitudinal study aimed to examine the relationship between the gut microbiome and depressive symptoms in women with gynecologic cancers throughout their cancer treatment. These findings could enhance our understanding of the mechanisms underlying depressive symptoms via the gut-brain axis and potentially contribute to the development of more targeted interventions.
Methods
Setting and Participants
The present longitudinal study is a secondary data analysis of an ongoing parent study titled “Gynecologic Cancer, the Vaginal Microbiome and Patient Symptom Experience” (Bai et al., 2021; Garcia et al., 2018). The parent study prospectively collects data at three time points, including baseline (T0), 6-8 weeks post-treatment (T1), and 6 months after T1 (T2), which were used for this analysis. The inclusion criteria for enrollment in the parent study are: (1) age ≥18 years; (2) diagnosed with cervical or endometrial cancer and scheduled for curative radiation therapy (RT) or chemoradiotherapy; (3) able to read English and willing to provide informed consent. The exclusion criteria are: (1) existence of co-morbidities (e.g., autoimmune diseases); (2) history of metastatic cancer; (3) current use of interferon; and (4) use of antibiotics/corticosteroids within four weeks prior to the baseline session. Approximately 170 cancer participants were enrolled in the parent study. Over the 1-year follow-up period, approximately 81 participants (48%) either withdrew or were lost to follow-up. The overall study consent rate was 73.3% among participants with cancer. For participants with cervical cancer, the average time from diagnosis to initiation of treatment was approximately 4–8 weeks. For participants with endometrial cancer, treatment (typically surgery) generally occurred within 2–8 weeks following diagnosis. When adjuvant therapy was indicated, treatment was typically initiated approximately 4–6 weeks after surgery to allow for postoperative recovery.
Data Collection
According to the protocol of the parent study, eligible participants were identified during their outpatient clinic visits and enrolled after providing informed consent. They were then asked to complete paper-based questionnaires at the three time points during regular patient appointments. Rectal swabs were taken at regular patient appointments or, if necessary, self-collected at home, approximately one inch inside the rectum using a CerviSoft foam-tipped collection device. These swabs were placed in sterile MoBio PowerBead Tubes on ice, mailed to the Emory University School of Nursing Biobehavioral Laboratory, and stored at −80°C until DNA extraction. Both physician-collected and participant self-collected vaginal samples were included in the study. Among participants with cancer, 416 timepoints were collected by physicians and 17 by participants. Two replicate samples were collected at each study time point.
Outcome Measures
Demographic and Clinical Characteristics
Demographic and clinical characteristics were obtained from patient-reported forms or electronic medical records. Demographic characteristics include age, race (Black, White, others), marital status (married vs. single/divorced), education, and income. Clinical characteristics include cancer site (endometrial vs. cervical), cancer stage (I vs. II≤), and treatment regimen (radiotherapy vs. chemoradiation).
Depressive Symptoms
Depressive symptoms were measured using the Patient Health Questionnaire (PHQ-9), (Arroll et al., 2010) a validated tool for measuring depressive symptoms in cancer patients (Hinz et al., 2016). The PHQ-9 consists of nine questions, each with four possible response options ranging from 0 (not at all) to 3 (nearly every day), yielding a maximum total score of 27, with higher scores indicating more severe depressive symptoms (Kroenke et al., 2001). A cutoff score of PHQ-9 ≥ 10 was used to indicate the presence of depressive symptoms (Barandouzi, Eng et al., 2023).
DNA Extraction and Microbiome Sequencing
DNA extraction was performed using the Mag-Bind® Environmental DNA 96 Kit, with samples processed on a KingFisher Flex automated instrument according to the manufacturer’s protocol (Omega Bio-Tek, GA). This method allowed for high-throughput and consistent extraction of high-quality DNA. The bacterial 16S rRNA gene was amplified using dual-barcoded primers targeting the V4 hypervariable region, specifically the sequences 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′), as outlined by Kozich et al. (2013) (Kozich et al., 2013). Dual barcoding enabled multiplexing, allowing multiple samples to be processed simultaneously while reducing cross-contamination risks. After PCR amplification, the amplicons were sequenced on an Illumina MiSeq platform using a v3 600-cycle kit, generating 250 bp paired-end reads. The raw reads were then processed using the Mothur software package (v. 1.48.1), following the MiSeq SOP (https://www.mothur.org/wiki/MiSeq_SOP). This entire process was conducted by Microbiome Insights (https://microbiomeinsights.com) to ensure methodological rigor and accuracy.
Batch Effect Correction
To address the batch effects resulting from 16S rRNA gene sequencing performed at two separate times in this study, we employed the R package “sva” (Surrogate Variable Analysis). This package is specifically designed for batch effect adjustment in high-throughput data. We utilized the ComBat function within the “sva” package, which adjusts for known batch effects using empirical Bayes models. This ensures that the observed variances in the microbiome data are more representative of true biological differences rather than technical artifacts.
Statistical Analysis
The demographic and clinical characteristics were calculated using descriptive statistics, including the mean for continuous variables and the frequency for categorical variables. After sequencing the gut microbiome data, the sequence data were trimmed at 0–220 for the forward and 0–190 for the reverse base to obtain high-quality sequences using DADA2. After quality control, an average of 27,543 high-quality sequences per sample was retained. Pre-trained Silva 138 classifier with a 99% threshold was used to determine the taxonomic annotation of the gut microbiota (docs, 2024). To normalize the data, rarefaction was conducted based on our sample sequencing depth. Quantitative Insights Into Microbial Ecology (QIIME) 2 was used for the gut microbiome diversity analysis (Bolyen et al., 2019). The α-diversity (evenness and richness within samples) was calculated using Shannon and Evenness indices. The β-diversity (dissimilarities between samples) was measured using Bray–Curtis distance and Jaccard metrics. Differential abundance analyses of gut microbial taxa at each time point were performed using MaAsLin2 multivariable linear models adjusted for batch effects. For the longitudinal analysis, significant α-diversity and β-diversity indices, and clinically relevant taxa were included in the generalized estimating equation (GEE) models to evaluate associations between the gut microbiome and depressive symptoms over cancer treatment.
Results
Patients’ Demographic and Clinical Characteristics
Demographic and Clinical Characteristics of the Patients at Baseline
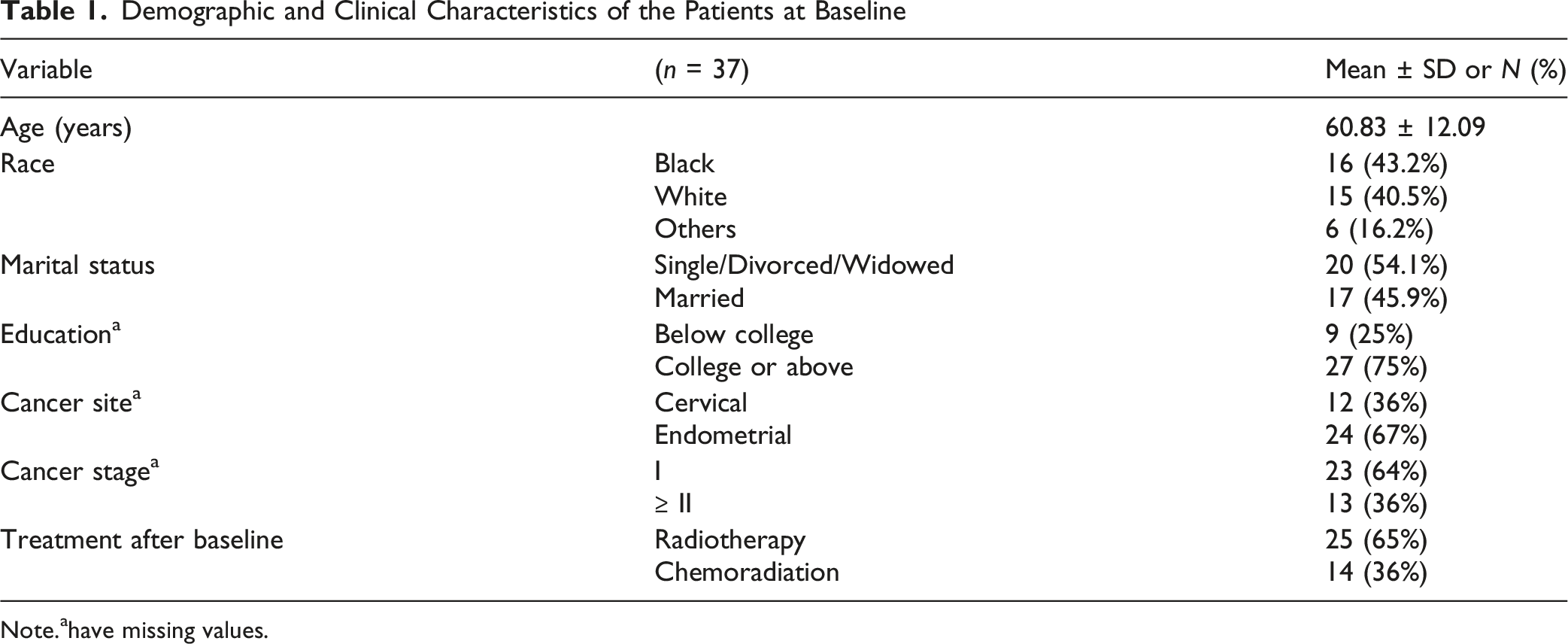
Note.ahave missing values.
Mean of Depressive Symptoms at Each Time Point
Depressive Symptoms at Each Time Point

Note. T0 = pre-treatment; T1 = 6–8 weeks post-treatment; T2 = 6 months post-treatment.
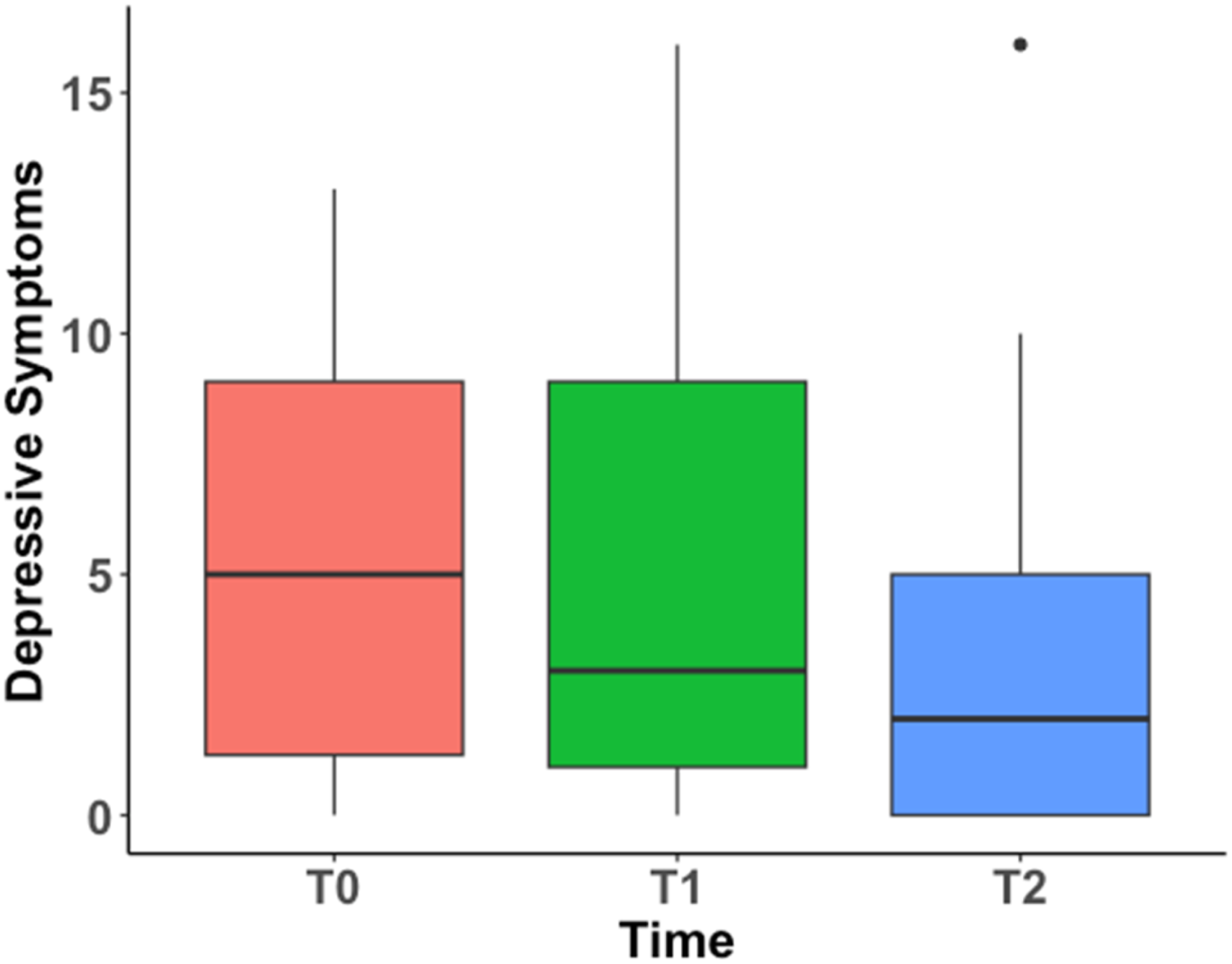
Change of depressive symptoms over time
Change of Depressive Symptoms Over Time (Using Mixed Effect Model)

Note. T1 = 6–8 weeks post-treatment; T2 = 6 months post-treatment. Bold indicates the significant values.
Diversity and Composition of Gut Microbiome Based on Depressive Symptoms at Each Time Point
Figure 2 presents boxplots of Shannon diversity and evenness by depression group at each time point. Although the group differences were modest and not statistically significant, the figures generally show slightly lower median alpha diversity values among participants with higher depressive symptoms across several T0, T1, and T2 (Figure 2). Alpha diversity based on depressive symptoms at each time point, T0 = pre-treatment; T1 = 6–8 weeks post-treatment; T2 = 6 months post-treatment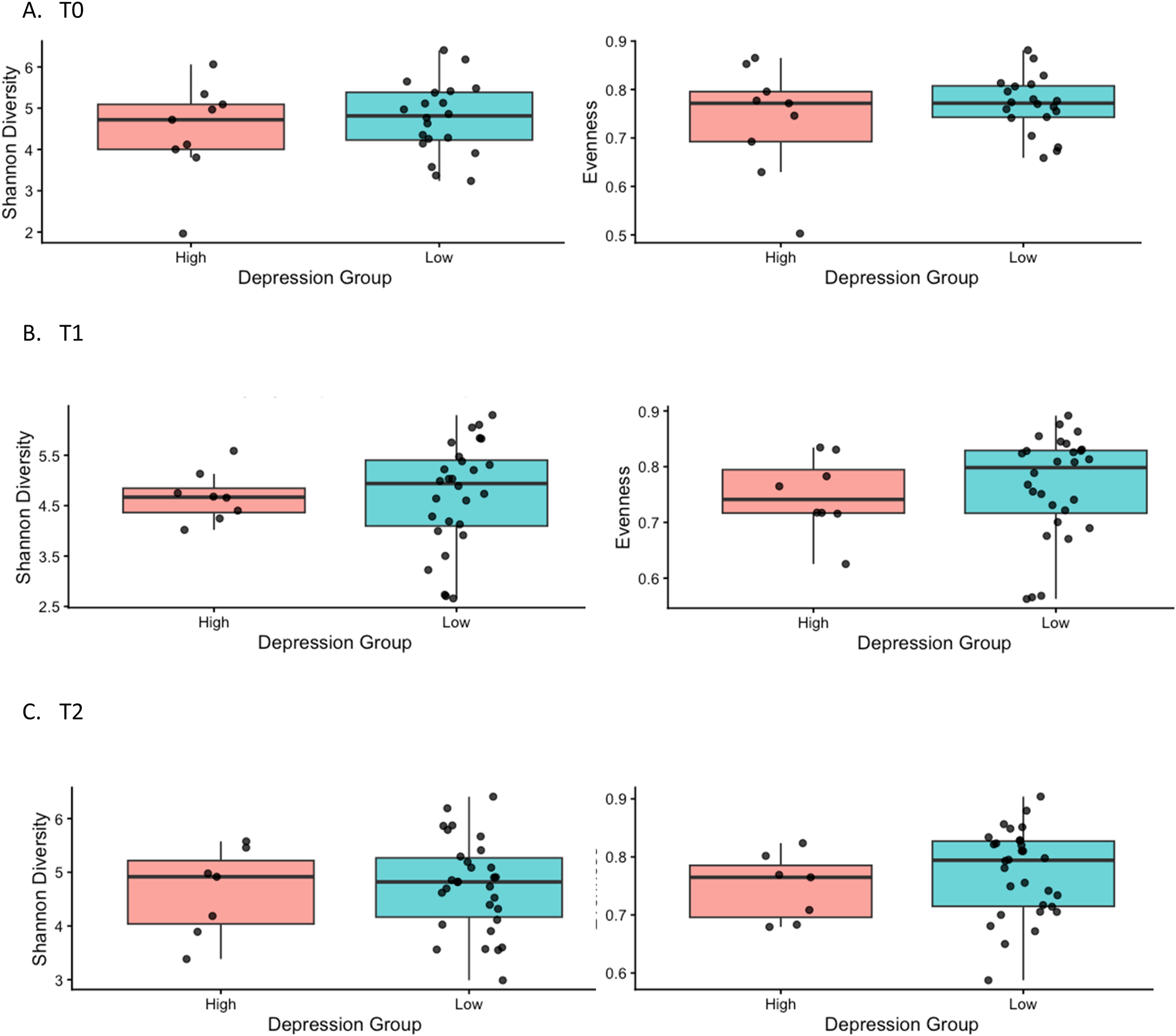
PERMANOVA models adjusted for batch effects were used to examine whether gut microbiome community composition differed by depressive symptom severity across T0, T1, and T2 using Bray–Curtis and Jaccard beta diversity indexes. No significant associations were identified between depressive symptoms and beta diversity at any time point. The PCoA plots (Figure 3). Beta diversity based on depressive symptoms at each time point, T0 = pre-treatment; T1 = 6–8 weeks post-treatment; T2 = 6 months post-treatment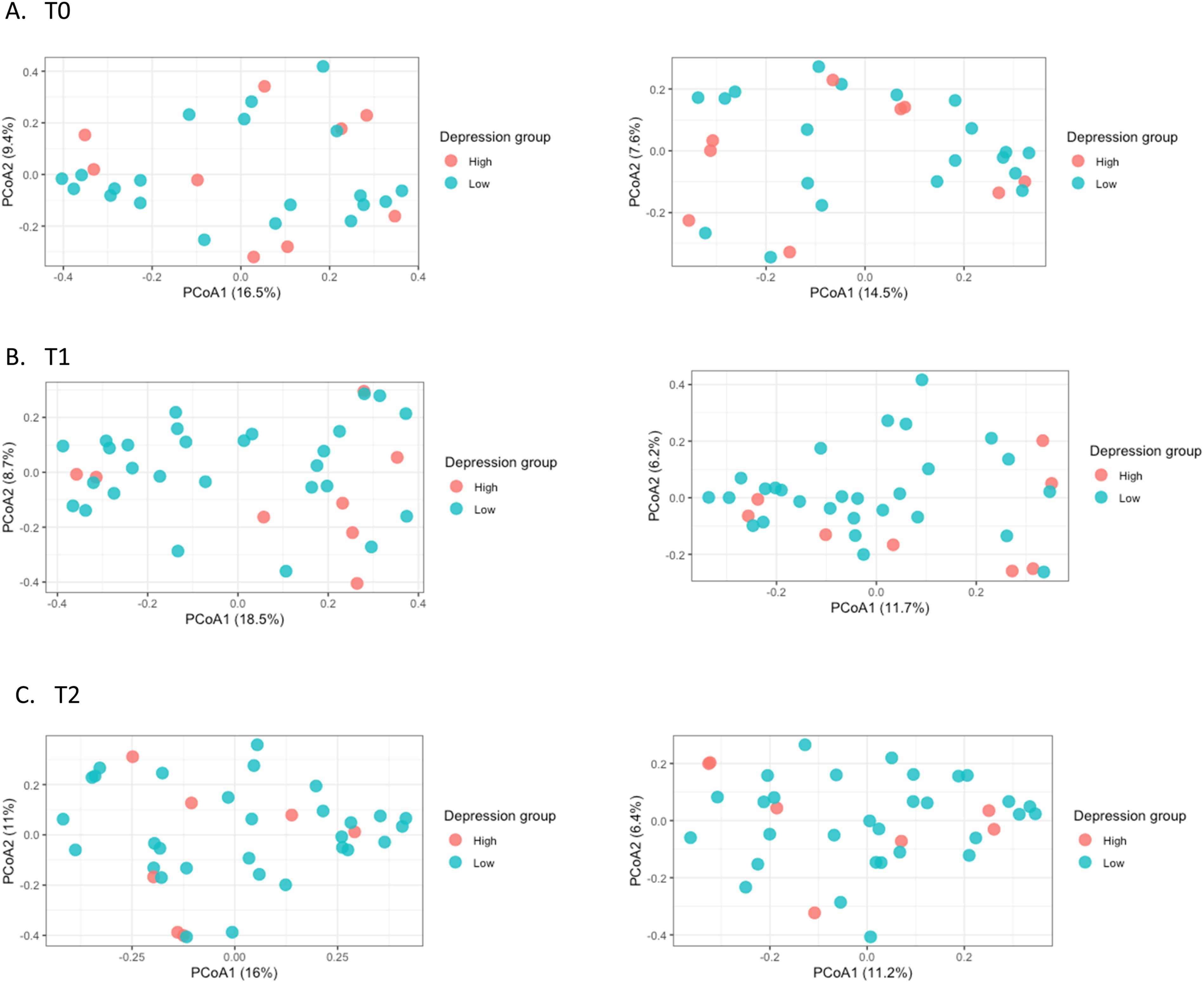
At T0, exploratory differential abundance analysis using MaAsLin2 identified lower relative abundance of Peptoniphilus, Anaerococcus, and Atopobium in the low depressive symptom group, whereas UCG-005 and UCG-002 showed higher relative abundance in the low depressive symptom group. Several Ruminococcus-related taxa also showed borderline associations at T0 (lowest unadjusted p ≈ 0.06). At T1, lower relative abundance of Anaerococcus, Sneathia, Gardnerella, Atopobium, Staphylococcus, Lactobacillus, and Prevotella was observed in the low depressive symptom group, whereas Bacteroides, Alistipes, Parabacteroides, Monoglobus, and Dialister showed higher relative abundance. Ruminococcus-related taxa demonstrated weak but non-significant associations (lowest p ≈ 0.08). At T2, Gardnerella, Staphylococcus, Prevotella, Dialister, Atopobium, and Peptoniphilus showed lower relative abundance in the low depressive symptom group, whereas Anaerococcus, Agathobacter, and Faecalibacterium, showed higher relative abundance. Also, Ruminococcus-related taxa again showed only borderline associations (lowest p ≈ 0.096). Although multiple associations reached nominal significance (unadjusted p < 0.05), none remained significant after false discovery rate correction (all q > 0.05); therefore, these results should be interpreted as exploratory.
Association Between Gut Microbiome and Depressive Symptoms Over Time
Generalized Estimating Equation Analysis of the Association Between Α-Diversity Indices and Depressive Symptoms Over Time Adjusting for Covariates
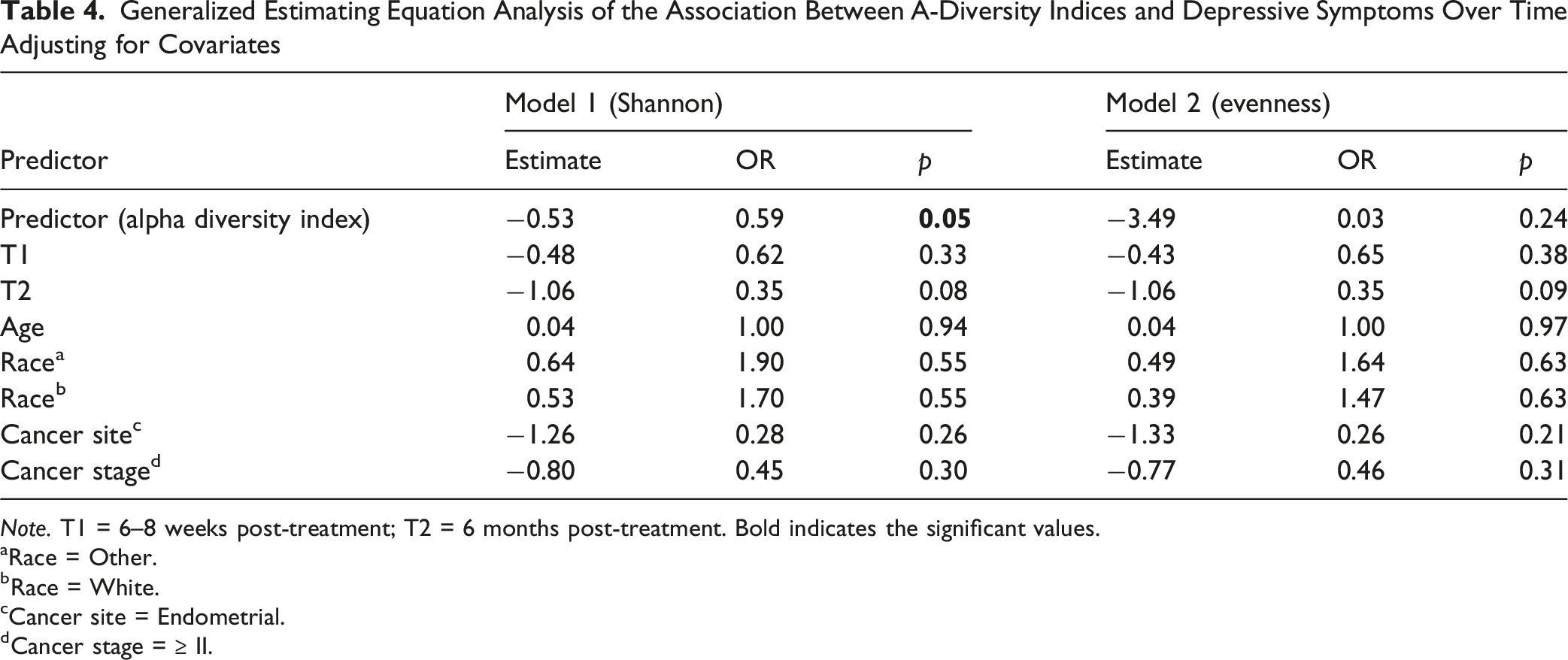
Note. T1 = 6–8 weeks post-treatment; T2 = 6 months post-treatment. Bold indicates the significant values.
aRace = Other.
bRace = White.
cCancer site = Endometrial.
dCancer stage = ≥ II.
Generalized Estimating Equation Analysis of the Association Between Β-Diversity Indices and Depressive Symptoms Over Time Adjusting for Covariates
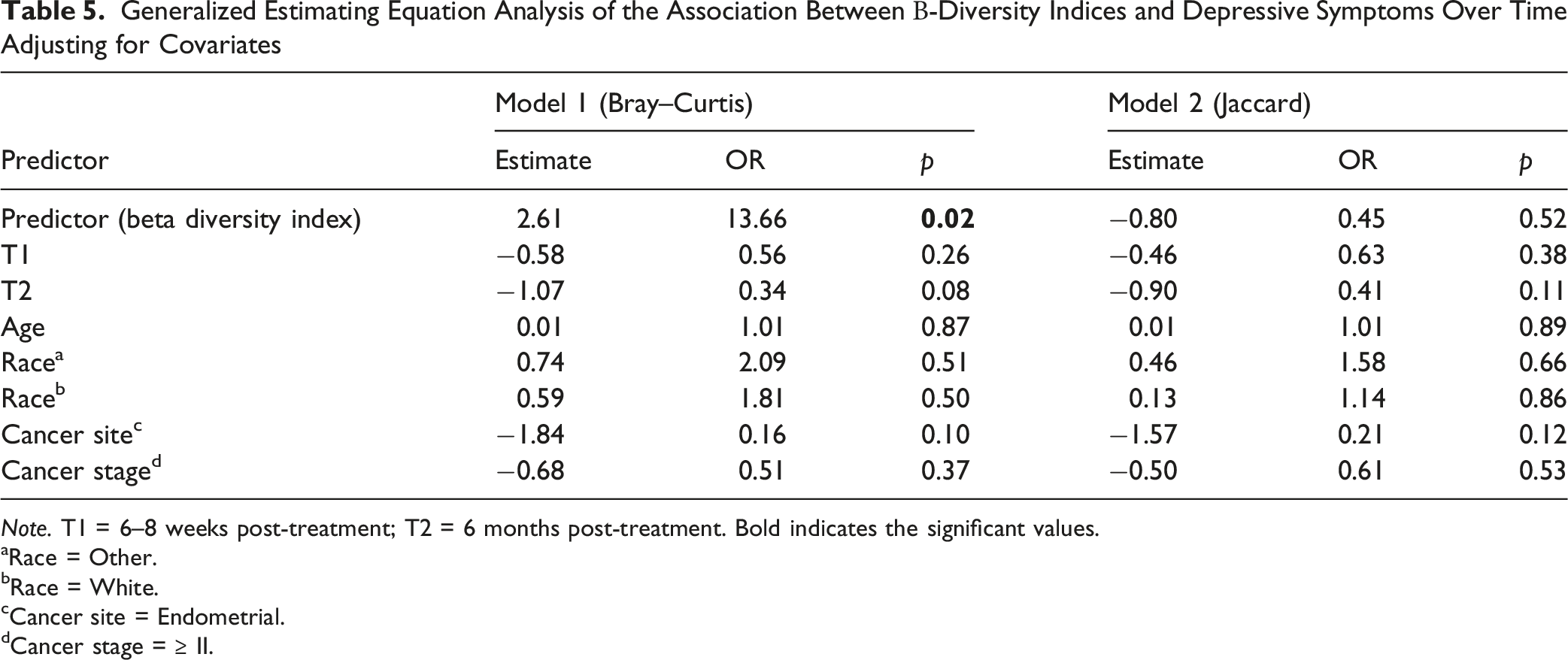
Note. T1 = 6–8 weeks post-treatment; T2 = 6 months post-treatment. Bold indicates the significant values.
aRace = Other.
bRace = White.
cCancer site = Endometrial.
dCancer stage = ≥ II.
Generalized Estimating Equation Analysis of the Association Between Genera and Depressive Symptoms Over Time Adjusting for Covariates
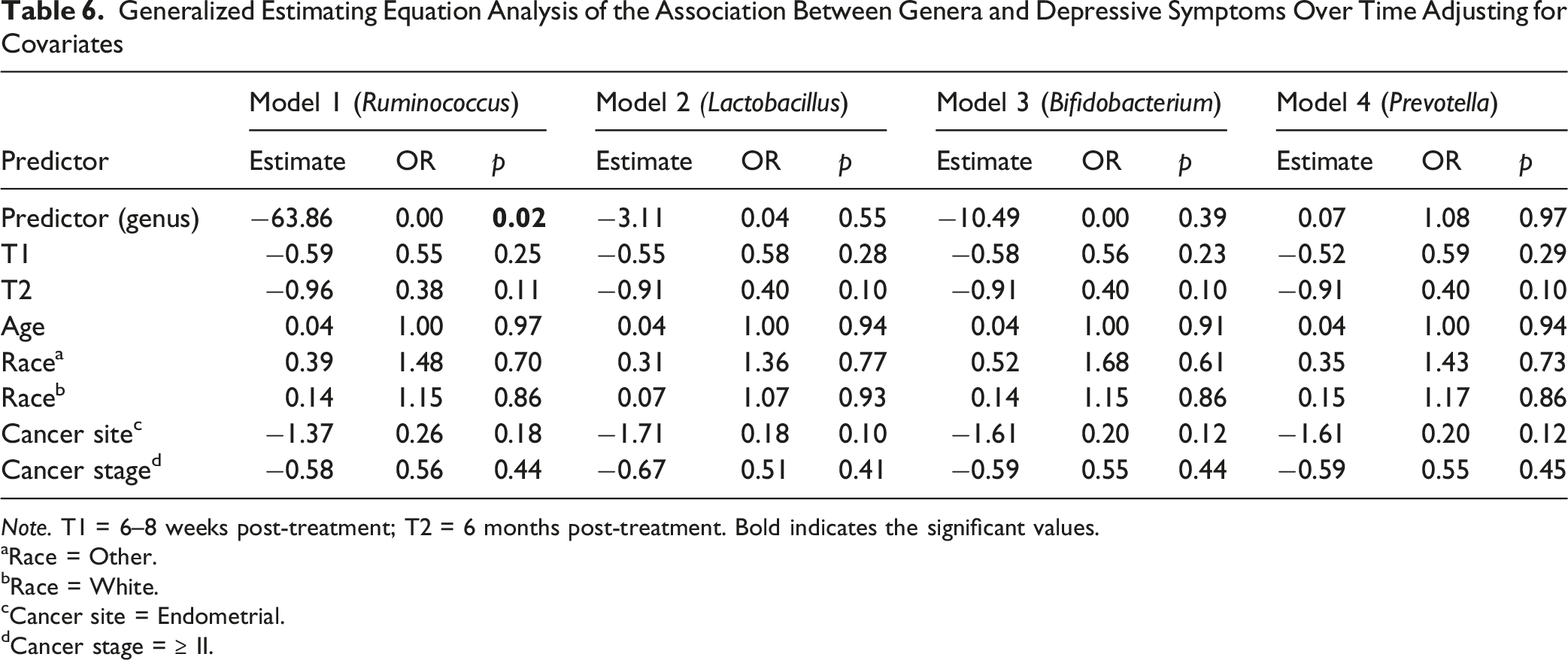
Note. T1 = 6–8 weeks post-treatment; T2 = 6 months post-treatment. Bold indicates the significant values.
aRace = Other.
bRace = White.
cCancer site = Endometrial.
dCancer stage = ≥ II.
Discussion
To our knowledge, this is the first longitudinal study to examine the relationship between the gut microbiome and depressive symptoms among women with gynecologic cancers throughout cancer treatment and early survivorship. We observed that depressive symptoms were most prevalent before treatment and gradually declined over the six months following treatment completion. Longitudinal analyses further demonstrated that higher depressive symptoms were associated with lower gut microbial Shannon diversity and differences in microbial community composition, as reflected by the Bray–Curtis index, over time. In addition, lower abundance of Ruminococcus was significantly associated with higher depressive symptoms across the treatment trajectory. Although several other taxa were identified in exploratory analyses at individual time points, these associations did not remain significant after correction for multiple testing. Taken together, these findings suggest that depressive symptoms in gynecologic cancer patients may be accompanied by longitudinal changes in gut microbial diversity and composition.
Depressive symptoms are frequently reported in the cancer population, significantly impacting their quality of life (Klapheke et al., 2020; Yu et al., 2020). According to our defined PHQ-9 cut-off of ≥10 in this study, 24% of gynecologic cancer patients experienced depressive symptoms before treatment, with 21% continuing to experience them post-treatment, decreasing to 13% six months post-treatment. In line with these findings, a cross-sectional study involving 274 gynecologic cancer patients who were planning or currently undergoing chemotherapy revealed that approximately 24% experienced moderate to severe depressive symptoms as measured by the PHQ-9 (Yu et al., 2020). Additionally, among 1,977 older women (aged ≥65 years) diagnosed with gynecologic cancers identified from the SEER-Medicare Health Outcomes Survey database, the prevalence of depressive symptoms was 31.9% for cervical cancer and 25.3% for uterine cancer (Klapheke et al., 2020). Although the prevalence of depressive symptoms among cancer patients differs, studies show that women with gynecological cancers have a higher occurrence of these symptoms compared cancer-free women in the general population (Klapheke et al., 2020; Massie, 2004). The onset of depressive symptoms even before the start of cancer treatment highlights their high prevalence and underscores the need for standardized screening in patients with gynecologic cancer.
There is still debate on the trajectory of depressive symptoms over time in the cancer population. Our results showed that depressive symptoms decreased over time among gynecologic cancer patients. Consistent with our findings, Bickel et al. (2022) reported a significant decrease in depressive symptoms over a three-month follow-up period among cancer patients diagnosed within the past five years who had moderate to severe depressive symptoms at baseline and were not receiving formal psychological care. However, despite this statistically significant decrease, the average level of depressive symptoms at follow-up remained in the moderate to severe range, suggesting that meaningful symptom burden persisted even among those showing improvement (Bickel et al., 2022). This finding suggests that reductions in depressive symptoms do not necessarily indicate full clinical recovery. Although depressive symptoms may improve during cancer treatment and survivorship, many patients continue to experience clinically significant symptoms. These results highlight the importance of regular screening for depressive symptoms throughout the cancer continuum, from diagnosis through survivorship, to identify patients who may benefit from timely mental health support and intervention.
In the present study, cross-sectional analyses conducted at individual time points did not identify significant differences in alpha or beta diversity according to depressive symptom status. However, when repeated measures were analyzed longitudinally, higher depressive symptoms were significantly associated with lower Shannon diversity and differences in microbial community composition measured by the Bray–Curtis index. These findings likely reflect the increased statistical efficiency of longitudinal analyses, which leverage repeated observations and account for within-subject correlation across the treatment trajectory. Related findings have been reported in other cancer populations. Among breast cancer patients, non-depressed breast cancer patients demonstrated greater gut microbial diversity compared to depressed patients, with alpha diversity serving as a significant mediator between diet quality and depressive symptoms (Maitiniyazi et al., 2022). Similarly, in head and neck cancer patients undergoing radiotherapy, those with higher psychoneurological symptom burden, including depressive symptoms, showed greater decreases in microbial evenness and more distinct community composition over time compared to those with lower symptom burden (Bai et al., 2020). Among women with gynecologic cancers specifically, lower alpha diversity was associated with higher psychoneurological symptom burden across the treatment period (Barandouzi, Bai et al., 2023). Collectively, these findings highlight the importance of longitudinal study designs for understanding microbiome–symptom relationships during cancer treatment and suggest that associations between the gut microbiome and depressive symptoms may become more apparent when evaluated over time rather than at a single assessment point.
At the taxon level, GEE analyses identified a lower abundance of Ruminococcus as significantly associated with higher depressive symptoms across the treatment trajectory. In exploratory MaAsLin2 analyses conducted at individual time points, participants with lower depressive symptoms had higher relative abundances of Ruminococcaceae UCG-005 and UCG-002 at T0 and higher abundances of Faecalibacterium and Agathobacter at T2; however, none of these associations remained significant after FDR correction. The observed depletion of Ruminococcus among participants with higher depressive symptoms is broadly consistent with previous studies linking members of the family Ruminococcaceae to depression-related outcomes. For example, in rectal cancer patients, Ruminococcaceae UCG-005 and uncultured Ruminococcaceae were negatively correlated with HAM-D depression scores (Gonzalez-Mercado et al., 2021). Similarly, individuals with anxiety and depressive symptoms undergoing gastrointestinal cancer screening had lower relative abundances of Ruminococcus compared with controls (Zhu et al., 2021). Although the specific taxa differed across studies, the overall pattern suggests that a lower abundance of Ruminococcaceae-related taxa may be associated with depressive symptoms.
The mechanisms of depressive symptoms in cancer patients are largely unknown. Our longitudinal data revealed that reduced α-diversity, as indicated by the Shannon index, as well as a dissimilar Bray-Curtis index, were associated with increased depressive symptoms over time. This reduced diversity suggests a potential imbalance in the gut microbial community, which may hinder its ability to produce essential metabolites like SCFAs that contribute to behavior regulation (Cheng et al., 2024). SCFAs can also modulate inflammatory pathways, and chronic inflammation has been implicated in the pathophysiology of depressive symptoms (Barandouzi, Bruner et al., 2023; Cheng et al., 2024; Feitelson et al., 2023).
Furthermore, our study identified a negative association between Ruminococcus abundance and depressive symptoms. One potential explanation for this finding relates to the role of Ruminococcus in carbohydrate fermentation and acetate production, which can support butyrate synthesis through cross-feeding interactions with other butyrate-producing bacteria, including Faecalibacterium prausnitzii and Roseburia spp (Baxter et al., 2019; Nogal et al., 2021; Rivière et al., 2016). SCFAs, particularly butyrate, have been implicated in gut–brain communication and may influence mood and behavior through multiple pathways. For example, SCFAs have been shown to regulate brain-derived neurotrophic factor (BDNF), which plays an important role in mood regulation and cognitive function (Suda & Matsuda, 2022). SCFAs also exhibit anti-inflammatory properties that may help mitigate the chronic inflammatory processes often associated with depressive symptoms (Mundi et al., 2023). Consequently, lower abundance of Ruminococcus may reflect a microbial environment with reduced capacity for SCFA production and altered gut–brain signaling. However, because SCFAs were not directly measured in this study, the underlying mechanisms remain hypothetical and warrant further investigation. Future studies are needed to clarify how Ruminococcus and related microbial taxa may contribute to depressive symptoms.
Limitations
This study has several limitations which must be considered when interpreting the results. The sample size was relatively small, with a majority of patients diagnosed with endometrial cancer at an early stage, which may limit the generalizability of the findings to the broader population of women with gynecologic cancers. We focused on one axis of beta diversity to simplify the complexity of the data. However, it’s important to note that this approach led to a loss of detail by not accounting for other dimensions of bacterial community variation when interpreting the results. Additionally, the present study identified associations between gut microbiota diversity, specific genera, and depressive symptoms but did not establish causality. Another important methodological consideration is the use of rectal swab samples rather than fecal specimens for microbiome analysis. Rectal swabs capture both luminal and mucosal-associated microbial communities and may differ from fecal samples in microbial composition and diversity (Jones et al., 2018; Sun et al., 2021). Therefore, findings should be interpreted within the context of specimen type when comparing results across microbiome studies. Information on participants’ prior history of depression and whether they were receiving treatment for depression during the study was not collected, which may have influenced the assessment of depressive symptoms. Another limitation is that immune-related markers, such as cytokines, were not measured alongside the microbiome, which limits our ability to fully understand the role of immune pathways in these relationships. Future research should aim to explore the causal relationships between gut microbiota alterations and depressive symptoms and incorporate measures of immune function to better understand the underlying mechanisms.
Conclusion
This longitudinal study reveals significant associations between the gut microbiota composition and depressive symptoms in women with gynecologic cancers over time. Key findings indicate that a lower α-diversity and dissimilar β-diversity of gut microbiota, along with a reduced abundance of the genus Ruminococcus, were predictors of depressive symptoms throughout cancer treatment. The study highlights the importance of gut microbiota diversity and specific taxa in the development of depressive symptoms in cancer patients. These insights highlight the need for future longitudinal research using advanced gut microbiota sequencing techniques to better understand both the mechanisms and functions of specific taxa and their association with depressive symptoms. Such understanding could lead to the development of targeted interventions aimed at restoring gut microbiota balance and alleviating depressive symptoms among women with gynecologic cancer.
Footnotes
Acknowledgments
The authors would like to express their gratitude to the research staff and patients who took part in the study.
Ethical Considerations
Ethics approval was obtained from the Emory University Institutional Review Board (IRB #00085823). This study was carried out in accordance with the principles of the Helsinki Declaration.
Consent to Participate
Patients provided informed consent to be involved in the study and to have the results published.
Author Contributions
Barandouzi, Z. Conception Design Data acquisition data analysis interpretation drafted manuscript critically revised manuscript gave final approval agrees to be held accountable for all aspects of work, ensuring integrity and accuracy Eng, T. Data acquisition critically revised manuscript gave final approval agrees to be held accountable for all aspects of work, ensuring integrity and accuracy Khanna, N. Data acquisition critically revised manuscript gave final approval agrees to be held accountable for all aspects of work, ensuring integrity and accuracy Shelton, J. Data acquisition critically revised manuscript gave final approval agrees to be held accountable for all aspects of work, ensuring integrity and accuracy Scott, I. Data acquisition critically revised manuscript gave final approval agrees to be held accountable for all aspects of work, ensuring integrity and accuracy Patel, P. Data acquisition critically revised manuscript gave final approval agrees to be held accountable for all aspects of work, ensuring integrity and accuracy Remick, J. Data acquisition critically revised manuscript gave final approval agrees to be held accountable for all aspects of work, ensuring integrity and accuracy Jin, R. critically revised manuscript gave final approval agrees to be held accountable for all aspects of work, ensuring integrity and accuracy Meador, R. Data acquisition critically revised manuscript gave final approval agrees to be held accountable for all aspects of work, ensuring integrity and accuracy Bruner, D. Conception Design data analysis interpretation drafted manuscript critically revised manuscript gave final approval agrees to be held accountable for all aspects of work, ensuring integrity and accuracy.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We received partial financial support from the Sigma Theta Tau Alpha Epsilon Chapter Research Grant Award and the Southern Nursing Research Society Research Grant Award.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.