
Abstract
Purpose:
Treatment of corneal injury depends on the self-proliferation ability of human corneal epithelial cells (HCEp). Our previous study revealed kojic acid had the anti-senescence function on human corneal endothelial cells. In this study, we researched the enhancive proliferation effect of kojic acid in HCEp.
Methods:
Cell viability was evaluated by MTT assay. The expression of proliferation-related protein was detected by western blotting and immunofluorescence assay.
Results:
Kojic acid could enhance HCEp proliferation, characterized by promoting cell proliferation rate, decreasing the expression levels of p21, galectin 8 and ki67, and increasing that of p-p38. The p38 signaling pathway inhibitor, SB203580, could reverse the enhancive proliferation function of kojic acid. Furthermore, knockdown of p21 had similar enhancive proliferation effect to kojic acid.
Conclusion:
Kojic acid might enhance HCEp proliferation through p38 and p21 signaling pathways, potentially via reduced expression levels of galectin 8 and ki67. Hence, kojic acid might be a potential drug to accelerate the healing of corneal epithelial injury.
Introduction
Corneal epithelium, the outermost layer of the cornea, is a defensive line to prevent infectious or toxic agents from penetrating the ocular surface. 1 Given its position, the cornea epithelium is sensitive to injuries that can result in permanent visual impairment. 2 The completeness of the corneal epithelium mainly relies on the consistent self-renewing function of corneal epithelial cells.3–5 Proliferation of corneal epithelial cells is involved in the healing mechanism of corneal injury. 6 Clinically, there is great advance in the therapy of corneal injury. However, there are still limited therapeutic drugs that can promote corneal healing. 7 Therefore, it is of great significance to promote proliferation of corneal epithelial cells for maintaining the line of defense.
P38 plays an important role in the physiology and pathophysiology of several biological systems, such as cell differentiation and proliferation.8,9 SB203580, as a p38 signaling inhibitor, dramatically reduced fibroblast proliferation.10,11 Cyclin-dependent kinase inhibitor, p21, plays a crucial role in suppression of cell proliferation. 12 Additionally, p21 regulates the cell cycle and participate in the regulation of cell proliferation in response to wounding of the corneal epithelium. 13 Galectin 8, as a part of the evolutionary conserved family of galectins, 14 inhibits cell cycle progression and induces growth arrest through up-regulation of p21. 15 Furthermore, ki67, as a marker of tumor proliferation, increased in mouse model of dry eye cornea. 16 Therefore, p21, p38, galectin 8, and ki67 might be correlated with the proliferation of human corneal epithelial cells (HCEp).
Kojic acid, one such tyrosinase inhibitor, is a naturally occurring fungal metabolite derived from Aspergillus, Penicillium, and Acetobacter genuses. 17 Kojic acid has various kinds of applications in different fields such as medicine, cosmetic industry, agriculture, food industry, and chemical industry. 18 In humans, kojic acid verified chronic therapy of as long as 14 years lead to no adverse local or systemic effects 19 and not to possess initiation and promotion activity of skin carcinogenesis. 20
Kojic acid and its ramification has various kinds of biological such as anti-oxidant, anti-tumor and anti-inflammatory.20–23 In our previous study, kojic acid inhibited senescence of human corneal endothelial cells, simultaneously activated NF-κB and p21 signaling pathways. 24 In this study, we will attempt to investigate the function of kojic acid on the proliferation of HCEp and its mechanism.
Materials and methods
Culture of HCEp
Human corneal epithelial cell line HCEp was purchased from the American Type Culture Collection (Manassas, VA, USA). HCEp were cultivated as monolayer in high-glucose DMEM (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS) (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100 µg/mL streptomycin and 100 U/mL penicillin (Beyotime Institute of Biotechnology, Haimen, China) at 37°C in humidified atmosphere with 5% CO2.
MTT assay
Cell proliferation of HCEp treated with different stimulating factors was evaluated by MTT assay. In this study, HCEp were treated with various concentrations of kojic acid from Sigma for 3 days (0–10,000 μM); or HCEp were treated with kojic acid (2000 μM) for 1, 3, and 5 days. Briefly, a density of 1.0 × 105 cells/well allowed to seeded in 96-well plate for 24 h. After culture with required days, HCEp were incubated with 20 μL of MTT solution (5 mg/mL in phosphate-buffered saline) each well and stored at 37°C for 4 h. Then, we removed 150 μL/well of supernatant and the formazan crystals were dissolved in 150 μL/well of DMSO. The optical density was measured at 570 nm using a microplate reader (BioTek, Winooski, VT). Each group was researched in duplicate and each experiment was repeated three times. 24
Antibodies
P-p38 rabbit monoclonal antibody (1:1000, cat no., 4511T; Cellsignaling, Boston, USA), p21 rabbit polyclonal antibody (1:1000, cat no. 10355-1-AP; proteintech, Chicago, USA), galectin 8 rabbit polyclonal antibody (1:1000, cat no. NFP1-66520; Novus Biological, Littleton, Colorado, USA), β-actin mouse monoclonal antibody (1:1000, cat no. AF0003; Beyotime Institute of Biotechnology, Haimen, China) were applied to this research.
Western blotting
The expression of proliferation relative protein was measured by western blotting. Briefly, cell lysate was added to cell lysis buffer [0.1 M Tris-Cl (pH 6.8), 0.05 M dithiothreitol, 4% sodium dodecyl sulfate, 0.1% bromophenol blue, 20% glycerol], and boiled for 5 min. Subsequently, protein samples were separated by SDS-PAGE and the separated proteins were electroblotted onto a poly (vinylidene difluoride) membrane (EMD Millipore, Billerica, MA, USA). After blocking the proteins in Tris-buffered saline containing 0.05% Tween-20 (TBS-T) and 5% skimmed milk for 1 h at room temperature, membranes were covered by the mentioned antibodies diluted in solution 1 (Toyobo Life Science, Osaka, Japan) at 4°C for 2 h. Then the membranes were washed with TBS-T for three times and covered with HRP-conjugated goat anti-mouse IgG or HRP-conjugated goat anti-rabbit IgG (1:5000; Thermo Fisher Scientific, Inc.) antibodies diluted in solution 2 (Toyobo Life Science) for 1 h at room temperature. Then, the antibody-antigen compound was measured through an enhanced chemiluminescent kit (Beyotime Institute of Biotechnology). 25 To normalize the data, β-actin was severed as an internal control of proteins. Image-J software (version 1.47, NIH) was used to measure band intensities. 24
Immunofluorescence assay
Immunofluorescence assay was used to evaluate the expression level of ki67. HCEp were seeded in 24-well plate for 24 h, then treated with 2000 μM of kojic acid for another 3 days. In brief, cells were fixed with 4% paraformaldehyde solution including 0.18% Triton X-100 at 4°C for 30 min. Following blocked within 2% goat serum (Shang hai yuan mu, Shanghai, China) diluted in PBS for 1 h, the cells were covered with the rabbit pAb against ki67 (1:1000, cat nos. ab92742; abcam, Cambridge, UK) at 37°C for 1 h. Then cells were covered with FITC-anti-rabbit IgG (1:250, cat nos. A11008; life technologies, New York, USA) at 37°C for 30 min. To clearly visualize cell nuclei, cells were counterstained with DAPI (1:5000, cat nos. C0060; Solarbio, Beijing, China). Finally, cells were visualized by immunofluorescence microscope and photographed with a camera (Olympus Corporation, Tokyo, Japan; magnification, ×100). Image-J software (version 1.47, NIH) was used to measure fluorescence intensities. 24
SB203580 administration and siRNA transfection
SB203580 (MedChempExpress, Monmouth Junction, USA), a p38 signaling inhibitor, was used to investigate the enhancive proliferation mechanism of kojic acid. HCEp were seeded in 96-well plates at 1.0 × 105 cells/well for 24 h. One well was co-treated with SB203580 (10 or 5 μM) plus kojic acid (2000 μM), and another well was only treated with kojic acid (2000 μM) or SB203580 (10 or 5 μM). The p21 specific siRNA (5′-AAUGGCGGGCUGCAUCCAGGATT-3′), galectin 8 specific siRNA (5′-GGCCUUUCAUUUCAAUCCUTT-3′), and control siRNA (5′-UUCUCCGAACGUGUCACGUTT-3′) were attained from GenePharma (Shanghai, China). For transfection of siRNA, HCEp were seeded in 6-well plates at 1.0 × 105 cells/well for 24 h. According to the manufacturer’s instruction, 100 pM of control siRNA, 100 pM of p21 siRNA or 100 pM of galectin 8 siRNA was transfected into HCEp with lipofectamine 2000 (Thermo fisher scientific, CA) according to the manufacturer’s instruction. After 6 h, the supernatant was replaced with fresh cell culture medium. Three days after transfection, cells were applied for MTT assay mentioned above, or cell lysate was collected for western blotting analysis. 24
Statistical analysis
SigmaPlot 12.0 (Hulinks, Inc., Tokyo, Japan) was used for statistical analysis. ANOVA with calculation of Tukey’s Honest Significant Difference test was performed to make pairwise comparisons within every experiment. In any case, p < 0.05 were considered statistically significant. 24
Results
Kojic acid enhances cell proliferation of HCEp
MTT assay was used to detect the kojic acid’s effect on the cell proliferation of HCEp. As shown in Figure 1(a), with the increase of kojic acid treatment concentration for 3 days, cell proliferation rate of HCEp increased and reached a maximum at 2000 μM (p < 0.001, F = 327.385). Furthermore, compared with 2000 μM, kojic acid at 5000 μM and 10,000 μM reduced the cell proliferation rate of HCEp after 3 days’ treatment. As shown in Figure 1(b), 2000 μM of kojic acid increased cell proliferation rate of HCEp after 3 days’ (p < 0.001, F = 238.953) 5 days’ treatment (p < 0.001, F = 329.958). As expected, there are more numbers of HCEp treated with 2000 μM of kojic acid than cells cultured for 3 days (Figure 1(c)). From the above, we will next enhance HCEp proliferation by 2000 μM of kojic acid treatment for 3 days, with HCEp cultured 3 days as control, and use 2000 μM of kojic acid to research its mechanism on promotion of HCEp proliferation.
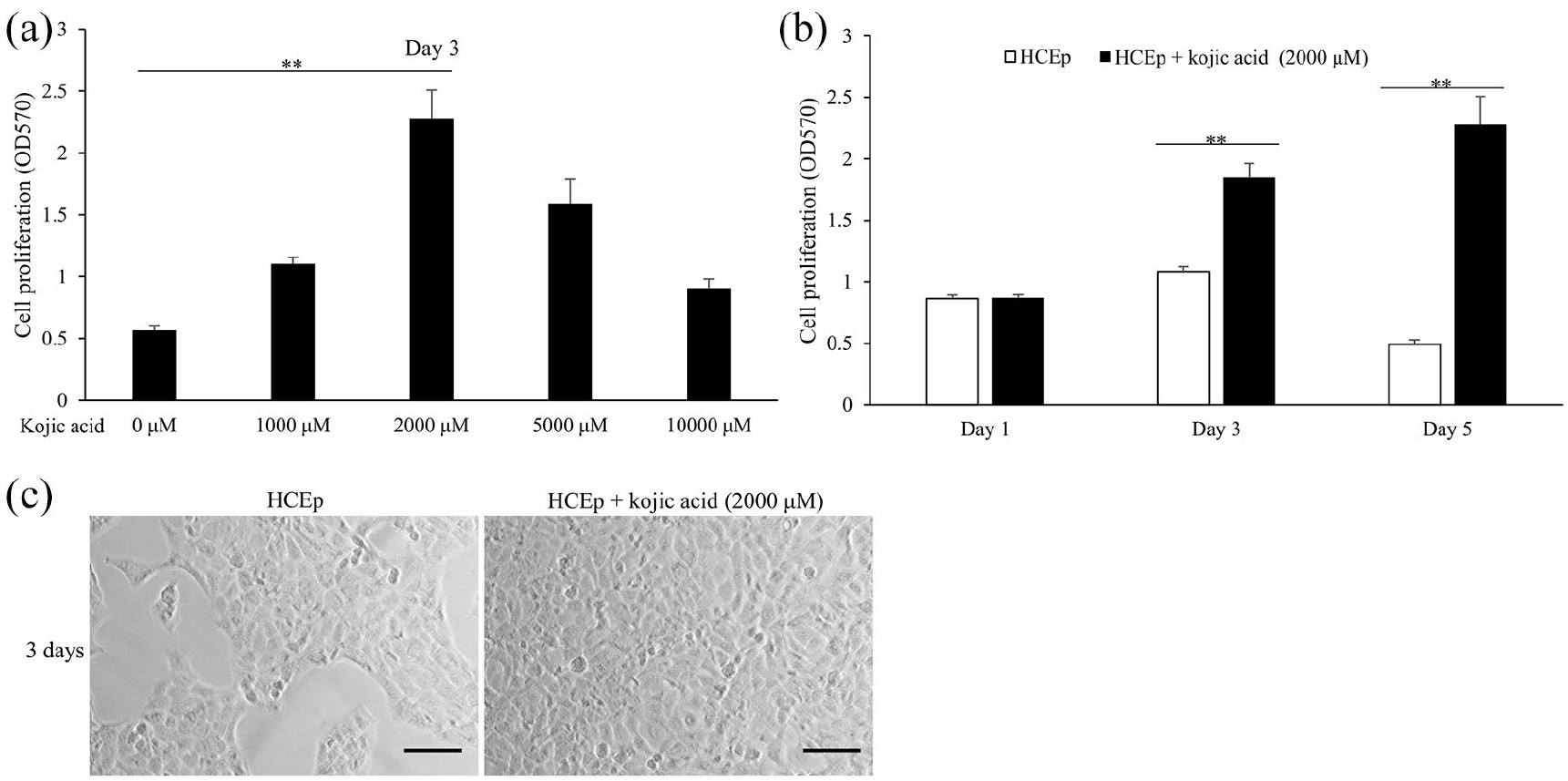
Kojic acid enhances the proliferation of HCEp. (a) Cell proliferation of HCEp treated with different concentrations of kojic acid (0 μM, 1000 μM, 2000 μM, 5000 μM, 10,000 μM) for 3 days by MTT assay. (b) Cell proliferation of HCEp treated with 2000 μM of kojic acid for 1 day, 3 days, 5 days by MTT assay. (c) Represented pictures of HCEp treated with 2000 μM of kojic acid or no treatment for 3 days. Scar bar: 100 μm.
Kojic acid influences the expression levels of proliferation relative proteins in HCEp
To research the enhancive proliferation mechanism of kojic acid in HCEp, we detected some proliferation relative proteins and signaling proteins by western blotting. As shown in Figure 2(a) and (b), compared to HCEp cultured for 3 days, cells treated with 2000 μM of kojic acid had superior expression level of p-p38 (p = 0.018, F = 14.758), lower expression levels of p21 (p = 0.011, F = 19.641) and galectin 8 (p < 0.001, F = 63.414). According to the results of immunofluorescence assay, HCEp treated with 2000 μM of kojic acid for 3 days expressed lower expression level of ki67 in cell nucleus (p = 0.006, F = 28.542) (Figure 2(c) and (d)). These results revealed that p-p38, p21, galectin 8 and ki67 might participate in the mechanism of kojic acid promoting cell proliferation in HCEp.
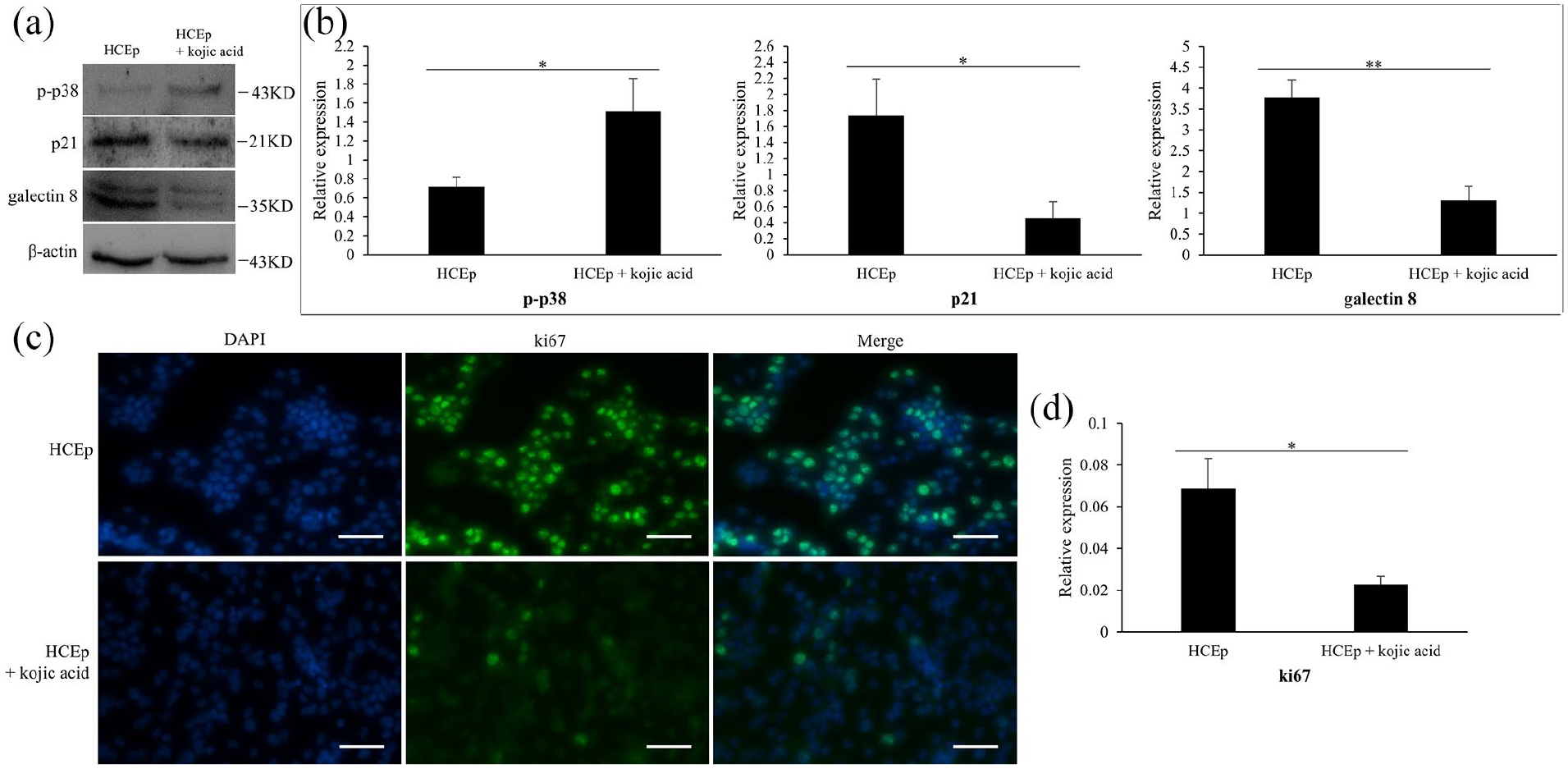
Kojic acid influences the expression levels of proliferation relative proteins in HCEp. (a) The expression of p-p38, p21 and galectin 8 in HCEp, and that treated with 2000 μM of kojic acid for 3 days by western blotting. (b) Quantitative analysis of western blotting results about p-p38, p21 and galectin 8. (c) The expression of ki67 in HCEp, and that treated with 2000 μM of kojic acid for 3 days by immunofluorescence. Scar bar: 100 μm. (d) Quantitative analysis of immunofluorescence result about ki67. Band intensities were normalized to β-actin. Scar bar: 100 μm.
SB203580 inhibits the enhancive proliferation effect of kojic acid on HCEp
To investigate the effect of p38 signaling pathway on the enhancive proliferation of kojic acid in HCEp, we introduced p38 signaling pathway inhibitor, SB203580, HCEp treated with SB203580 alone was considered as negative control. The result was shown in Figure 3, compared to HCEp treated with kojic acid alone, HCEp treated with kojic acid plus 10 μM of SB203580 (p < 0.001, F = 96.146) or 5 μM of SB203580 (p < 0.001, F = 804.838) had significantly lower cell proliferation rate by MTT assay. Additionally, compared to HCEp cultured for 3 days, cells treated with SB203580 alone could not influence the cell proliferation. These results confirmed that p38 signaling pathway participated in the enhancive proliferation mechanism of kojic acid in HCEp.
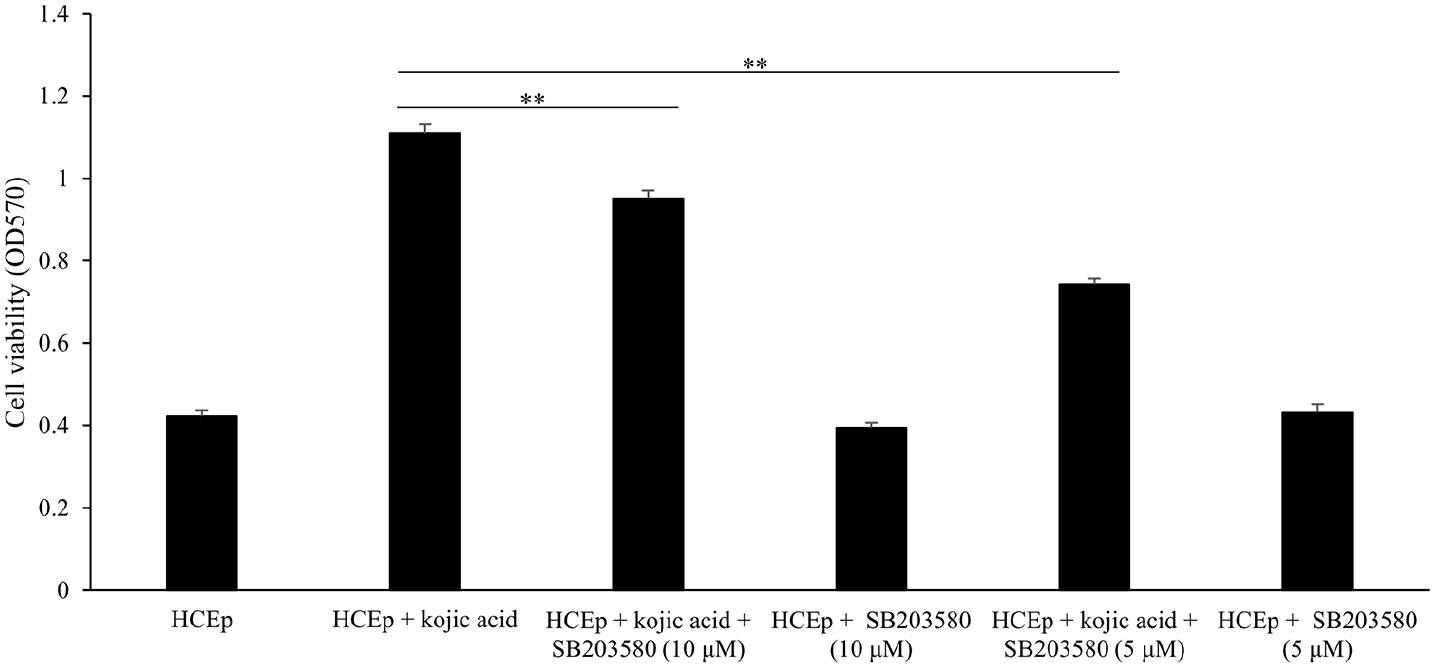
SB203580 inhibits the proliferation effect of kojic acid on HCEp. Cell proliferation of HCEp treated with 2000 μM of kojic acid, treated with kojic acid plus 10 μM of SB203580 and treated with kojic acid plus 5 μM of SB203580.
P21 siRNA enhance HCEp proliferation
To detect the effect of p21 signaling pathway on the enhancive proliferation of kojic acid in HCEp, we transfected p21 siRNA into cells. As shown in Figure 4(a), HCEp transfected with p21 siRNA could decrease the expression level of p21 compared to control siRNA. According to the result of MTT assay, HCEp transfected with p21 siRNA, but not galectin 8 siRNA, had significantly higher cell proliferation rate (p < 0.001, F = 72.883) (Figure 4(b)). The results by western blotting showed that HCEp transfected with p21 siRNA could reduce the expression level of galectin 8 (p = 0.007, F = 26.276) (Figure 4(c) and (d)). Moreover, p-p38 was not detected in HCEp transfected with p21 siRNA (Figure 4(c)). These results indicated that p21 signaling pathway might participate in the enhancive proliferation mechanism of kojic acid promoting cell proliferation in HCEp, and influence the expression level of galectin 8.
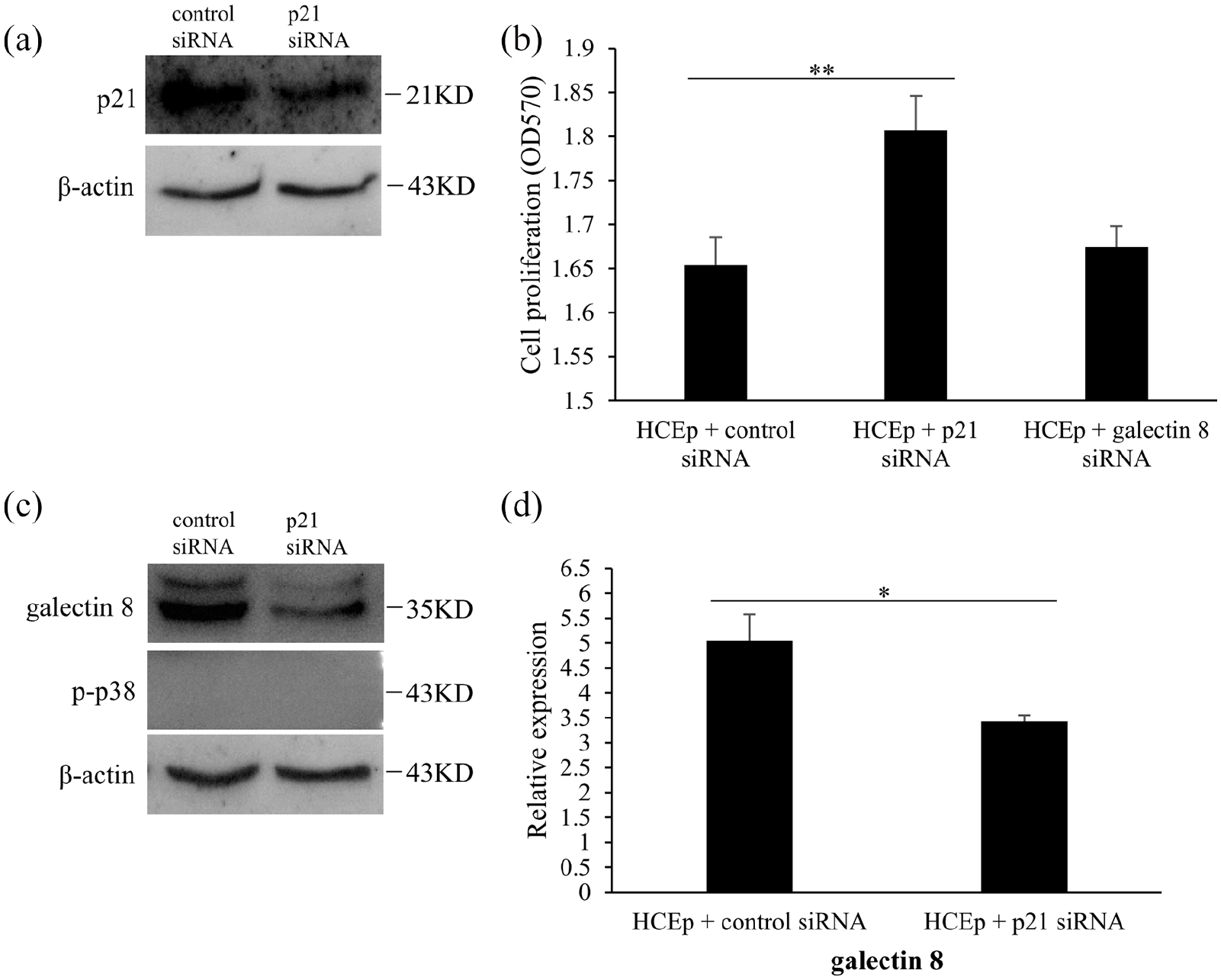
P21 siRNA enhances the proliferation of HCEp. (a) The expression of p21 in HCEp transfected with 100 pM of control siRNA, 100 pM of p21 siRNA for 3 days by western blotting, (b) Cell proliferation of HCEp transfected with 100 pM of control siRNA, 100 pM of p21 siRNA and 100 pM of galectin 8 siRNA for 3 days by MTT assay. (c) The expression of galectin 8 and p-p38 by western blotting. (d) Quantitative analysis of western blotting results about galectin 8 and p-p38. Band intensities were normalized to β-actin.
Discussion
In this study, we demonstrated that kojic acid could enhance HCEp proliferation, simultaneously, decrease the expression levels of p21, galectin 8 and ki67, and increase the expression level of p-p38 in HCEp. The p38 signaling pathway inhibitor, SB203580, could reverse the enhancive proliferation function of kojic acid, and knockdown of p21 in HCEp had similar enhancive proliferation effect with kojic acid. These results supposed that kojic acid may enhance HCEp proliferation via p38 and p21 signaling pathways.
Kojic acid is known to have various bioactivities. 17 In our previous study, kojic acid decreased the expression levels of p21 and galectin in human corneal endothelial cells16. In this study, we firstly detect the function and mechanisms involved of kojic acid in HCEp. The result revealed that, as expected, kojic acid also via p21 signaling pathway enhanced proliferation of HCEp and influenced the expression level of galectin 8. Furthermore, p38 signaling pathway and ki67 took part in the enhancive proliferation process of kojic acid in HCEp.
In addition, we found that galectin 8 was downregulated in HCEp treated with kojic acid as well as cells transfected with p21 siRNA. This result was consistent with our previous research which galectin 8 took part in the anti-senescence process of kojic acid in human corneal endothelial cells and was regulated by p21 signaling pathway. Therefore, the interaction between p21 signaling pathway and galectin 8 plays a significant role in the research of kojic acid’s function in cornea.
Ki67 is a well-known proliferation marker that is widely used in the diagnosis of several pathologic lesions, especially human tumors and malignancies. 26 A recent research revealed that the expression level of ki67 increased in mouse model of dry eye cornea. 16 In our study, kojic acid enhanced cell proliferation of HCEp, however, decreased the expression of ki67. These results might suggest that the mechanism by which kojic acid promoted the proliferation of HCEp may be a more complex process involving ki67. In addition, these results also enlighten us that kojic acid may be a potential drug composition in the treatment of dry eye disease, but this possibility needs to be verified by more studies.
The p38 signaling pathway plays a significant important role in the process of cell proliferation.8,9 In this study, we introduced SB203580, a signaling inhibitor of p38, to prove that p38 signaling pathway was involved in the process of kojic acid promoting the proliferation of HCEp. Furthermore, we found p-p38 in HCEp could not be detected after p21 was knocked out.
HCEp proliferation is involved in the healing mechanism of corneal injury. Promoting the proliferation of HCEp can accelerate the healing of corneal injury. In our research, kojic acid could enhance proliferation of HCEp. Hence, kojic acid may be a potential drug for therapy of corneal injury.
There were two major boundedness in this study. Firstly, we used only HCEp line to investigate the proliferative effect of kojic acid. Secondly, there is no further study on the contradiction between the enhancive proliferation effect of kojic acid and the reduced expression of ki67 in HCEp, which will not only accelerate the function research of kojic acid, but also benefit to detect new targets for therapy of corneal injury.
Conclusion
Kojic acid might enhance HCEp proliferation through p38 and p21 signaling pathways, possibly via reduction of galectin 8 and ki67. Hence, kojic acid is a potential drug to promote the healing of corneal injury.
Footnotes
Author contributions
P.L., X.L.T., X.G.L., and H.Q. designed this research. X.W., X.G.L., and H.Q., L.Y.S., C.W., F.Q.S., L.L.D., and X.G. performed the experiments, data collection, analysis, and interpretation. X.G.L. and H.Q. guided and supervised this research. X.W., X.G.L., and H.Q. wrote the manuscript. P.L. and X.L.T. revised the manuscript. All authors checked and approved the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Nature Science Foundation of China (no. 81470618), Scientific Research Innovation Fund of the First Affiliated Hospital of Harbin Medical University (no. 2017B003).