
Abstract
Purpose
This study aimed to explore the pterygium formation and recurrence, by detecting the expression of Nod-like receptor pyrin domain 3 (NLRP3) and Nod-like receptor pyrin domain 6 (NLRP6) in pterygium and evaluate the correlation between NLRP3 and NLRP6 in pterygium.
Methods
In this prospective study, the expression levels of NLRP3 and NLRP6, with their related effectors, were evaluated in primary pterygium (n = 40) and recurrent pterygium (n = 32) tissue samples and compared with normal conjunctiva (n = 11) tissue samples by immunohistochemistry.
Results
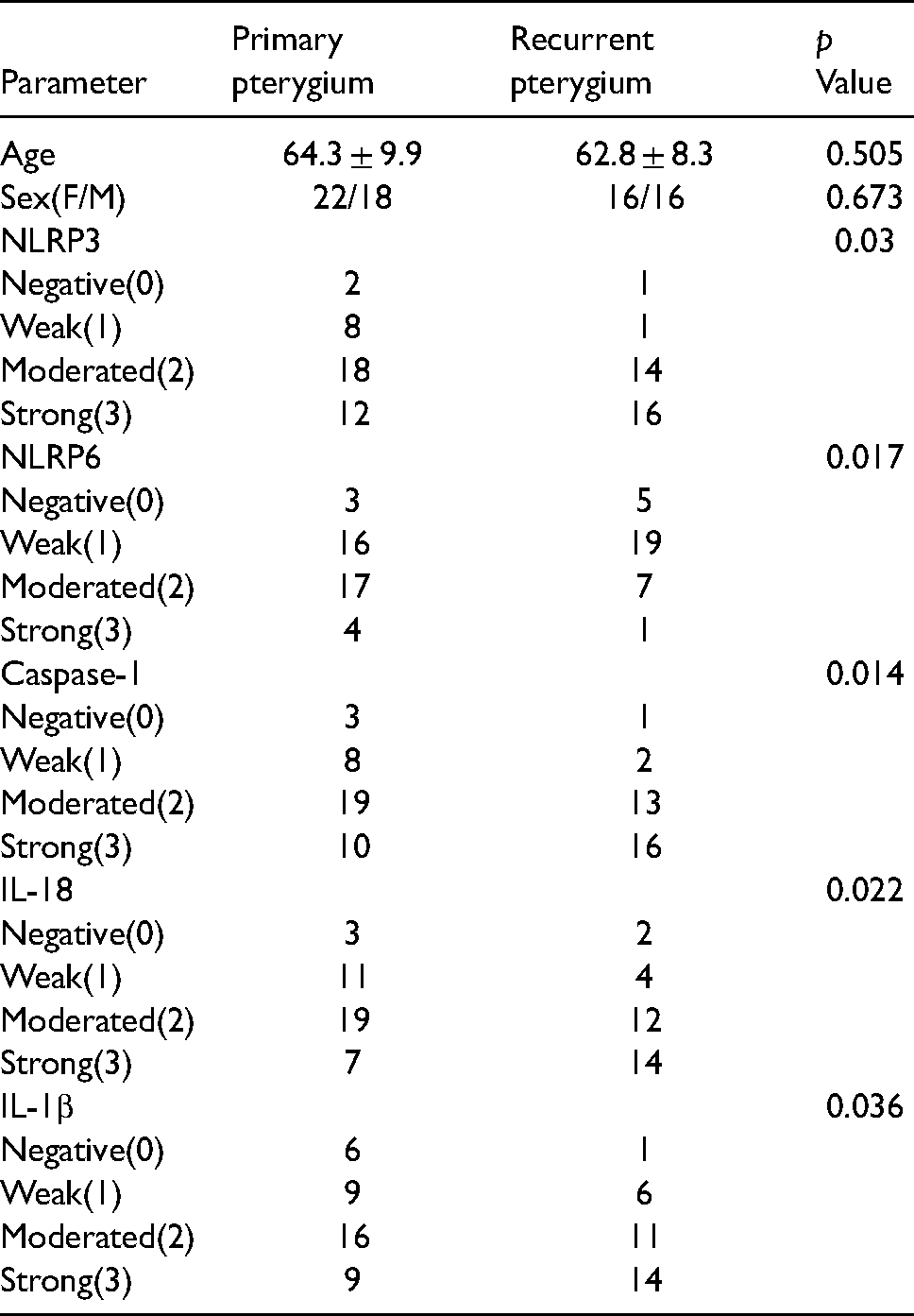
Compared to the normal conjunctiva group, the expression levels of NLRP3, caspase-1, IL-18, and IL-1β, were significantly higher, and NLRP6 showed an expression that was significantly lower in pterygium tissue samples (P < 0.05, respectively). Compared to the primary pterygium group, the expression levels of NLRP3, caspase-1, IL-18 and IL-1β were significantly higher, and NLRP6 showed an expression that was significantly lower in recurrent pterygium tissue samples (P < 0.05, respectively).There was a negative correlation between NLRP3 expression and NLRP6 expression in normal conjunctival (r = −0.739, P = 0.009) and pterygium (r = −0.533, P = 0.000).
Conclusions
NLRP3 and NLRP6 may be involved in the formation and recurrence of pterygium. NLRP6 may play an anti-inflammatory role in normal conjunctival tissue to maintain conjunctival homeostasis.
Introduction
Pterygium is a common disease with a high incidence in ophthalmology. In later stages of pterygium, the pterygium tissue can invade the cornea, covering the pupil area, and seriously damage vision. 1 The pterygium recurrence is still the most common problem after surgery. Recent studies have shown that lymphocyte infiltration and immunoglobulin deposition can be found in pterygium tissues which are accompanied by abnormal secretion of IL-1, IL-6, IL-8, and TNF-γ. 2 The interaction between immune cells and inflammatory factors leads to fibroblast proliferation and angiogenesis, which induces the progression and recurrence of pterygium.3,4
NLRs-mediated inflammatoryosomes have been reported to take part in innate immune-induced inflammation, which usually contains adapter apoptosis-related spot-like protein of CARD (ASC). NLRs combine with proto-cysteine-1 to form a multiprotein known as the inflammatory complex,5,6 which activates one or more cysteine proteases for the secretions of proto-inflammatory cytokines, interleukin-1β (IL-1β) and IL-18.7,8 NLRP3, an important member of inflammasom, is most widely studied at present, and can be activated by multiple stimuli.9–11 In the NLRP family, NLRP6 is the first reported to inhibit an innate immune response and negatively regulate inflammatory signals. 12 However, NLRP6 has been reported to be up-regulated in inflammation-related diseases, such as pneumonia, Crohn's disease, and periodontitis.13–15 Therefore, the effect of NLRP6 remains controversial in inflammation.
Most studies have shown that a low dose of mitomycin C decreases the postoperative recurrence rate of pterygium by perturbing the NLRP3 inflammatory signaling pathway and suppressing the expression of inflammatory factors. 16 However, the mechanism in pterygium NLRP6 expression has not been reported. Additionally, NLRP3 and NLRP6 have not been reported in recurrent pterygium.
For the previously stated reasons, we aimed to explore whether NLRP3 and NLRP6 are involved in the formation or recurrence of pterygium by evaluating the biomarkers NLRP3, NLRP6, caspase-1, IL-18, and IL-1β in pterygium tissue samples.
Materials and methods
72 outpatient subjects underwent single pterygium surgery due to 40 primary pterygia and 32 recurrent pterygia and were included from May 2020 to April 2021. The normal conjunctiva was from 11 patients as a control group during conjunctivochalasis surgery. The excluded symptoms included infectious conjunctivitis, keratitis, allergic conjunctivitis, xerophthalmia, blepharitis, uveitis, strabismus, amblyopia, glaucoma, nystagmus, eye surgery or trauma within the last 3 months, previous use of systemic or local corticosteroids, mitomycin C, or immunosuppressive agents. This research was conducted following the principles of the Declaration of Helsinki and approved by the Ethics Committee. Informed consent was obtained from all participants.
Immunohistochemistry
All tissue samples were immediately immobilized with a 4% formalin, and then treated and embedded by paraffin according to the immunohistochemical procedures. The 4 mm thick section was dewaxed with xylene and treated with 0.01 M citric acid buffer (pH 6.0) in the microwave for 10 mins so that the antigens were extracted. Sections were treated with 3% hydrogen peroxide for 10 mins to stop endogenous peroxidase activity and then incubated with normal goat serum at room temperature for 30 mins to prevent non-specific background. Washed with buffer solution, the sections were incubated with antibodies at 4°C overnight, including anti-NLRP3, anti-NLRP6, anti-Caspase-1, anti-IL-18, and anti-IL-1β (1:100 dilution). After that, the slices were incubated with the EnVision horseradish peroxidase system at room temperature for 30 mins and DAB for 5 mins, which were stained with hematoxylin dehydrated, and finally packed.
The expression levels of NLRP3, NLRP6, and their related effector proteins were detected in all samples. Cells with brown cytoplasm and nuclei were identified as positive. The protein staining results based on the quantity of staining intensity (0: negative, 1: weak, 2: medium, 3: strong) were evaluated by two pathologists who did not know the clinical information of the patients.
Statistical analysis
The SPSS program, version 19.0, was used for statistical analysis. A student's t-test was used for age data analysis. An X2 test was used for gender comparison, and the non-parametric Mann-Whitney test was used for staining intensity. Spearman's correlation coefficient for ranked data was used for group correlation. A p-value of <0.05 was considered significant.
Results
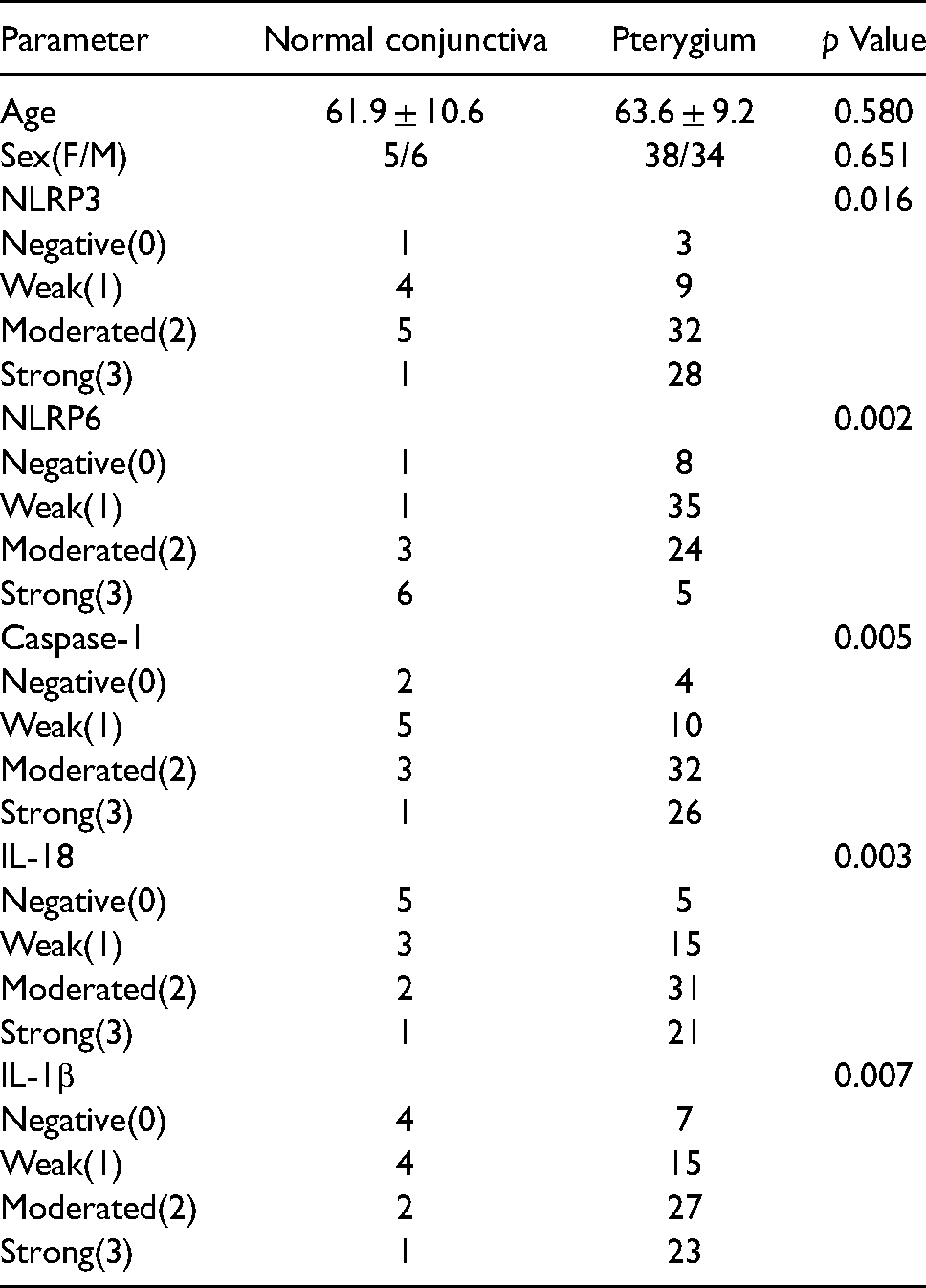
The control group included 11 patients (6 males and 5 females), whose mean age was (61.9 ± 10.6) years. The primary pterygium group included 40 patients (18 males and 22 females), whose mean age was (64.3 ± 9.9) years. The recurrent pterygium group included 32 patients (16 males and 16 females), whose mean age was (62.8 ± 8.3) years. There was no significant difference between the groups regarding age and gender (P > 0.05, respectively). (Table 1).
Expression of NLRP3, NLRP6, caspase-1, IL-18 and IL-1β in normal conjunctiva and pterygium.
Expression of NLRP3, NLRP6, caspase-1, IL-18, and IL-1β in normal conjunctiva and pterygium (Table 1)
Compared to the normal conjunctiva group, the expression levels of NLRP3, caspase-1, IL-18, and IL-1β were significantly higher, and NLRP6 displayed a significantly lower expression in pterygium tissue samples (P < 0.05, respectively). The high expressions of NLRP3, caspase-1, IL-18, and IL-1β in pterygium were primarily located in epithelial cells and fibroblasts, and the high expression of NLRP6 in normal conjunctiva was mainly located in epithelial cells and fibroblasts as well (Figure 1).

(a,b,c):NLRP3 Expression in normal conjunctiva,primary pterygium and recurrent pterygium( × 400)
Expression of NLRP3, NLRP6, caspase-1, IL-18, and IL-1β in primary and recurrent pterygium (Table 2)
Compared to the primary pterygium group, the expression levels of NLRP3, caspase-1, IL-18, and IL-1β were significantly higher, and NLRP6 had an expression that was significantly lower in recurrent pterygium tissue samples (P < 0.05, respectively).
Expression of NLRP3, NLRP6, caspase-1, IL-18 and IL-1β in primary and recurrent pterygium.
The correlation between NLRP3 and NLRP6
There was a negative correlation between the NLRP3 expression and the NLRP6 expression in normal conjunctival (r = −0.739, P = 0.009) and pterygium (r = −0.533, P = 0.000).
Discussion
Primary and recurrent pterygium is associated with inflammation. 17 In this study, the expression of NLRP3, caspase-1, IL-18, and IL-1β was higher in pterygium tissues compared with the normal conjunctival tissues. While the expression of NLRP6 was higher in normal conjunctival tissues as compared to pterygium tissues. It is considered that NLRP3 may be involved in the formation of pterygium by activating caspase-1, IL-18, and IL-1β, as well as NLRP6, which plays a protective and inhibiting inflammatory role in normal conjunctiva.
The expression levels of NLRP3 and its interacting proteins were significantly increased in pterygium, which was consistent with previous studies. 16 Currently, it has been found that a variety of stimulants in vivo and in vitro can abnormally activate NLRP3 inflammasom. As ATP opens the ion channel by activating the P2X7 receptor on the surface of the cellular membrane to change the intracellular and extracellular ion environment, NLRP3 can recognize the exogenous molecules to active inflammasome. 18 When the inflammasome is out of control, excessive inflammatory responses will break out in our bodies to lead to autoimmune diseases. 19 Pterygium is generally considered to be a chronic inflammatory injury caused by external stimulation, which can metastasize from the conjunctiva to the cornea. The pathogenesis of pterygium mainly related to ultraviolet radiation is still controversial. Inflammation, viral infection, oxidative stress, and other factors are all considered to be epidemic factors. 20 Therefore, we believe that NLRP3 might be activated by the stimuli to cause the activation of the downstream signaling pathway mediated by caspase-1, which can promote the secretion of inflammatory cytokines IL-1β and IL-18, damaging the normal conjunctival tissue, and lead to the occurrence of pterygium.
NLRP6 has been reported to promote inflammation by activating proinflammatory cytokines,13,21,22 and to reduce inflammation by inhibiting NF-κB, ERK, and TNFα signaling pathways.23–26 In normal conjunctiva, NLRP6 had an expression level that was higher, while the expression levels of caspase-1, IL-18, and IL-1β were lower. It is suggested that NLRP6 might inhibit inflammation through NF-κB and ERK signaling pathways. Low expression of NLRP6 in pterygium may lead to the incidence of pterygium due to the weakened inhibition to inflammation.
Additionally, our study found that the expression levels of NLRP3, caspase-1, IL-18, and IL-1β were higher in recurrent pterygium than primary pterygium, which suggests that the inflammation effect is more obvious in recurrent pterygium than primary pterygium. A high expression of NLRP3 may cause the recurrence of pterygium. A similar previous study showed the activation of NLRP3 and caspase-1 pathways might be a strong driving force for promoting angiogenesis and fibroblast proliferation to induce pterygium recurrence. 16 Moreover, this study confirmed that low-dose mitomycin C could inhibit the expression of inflammatory factors by interfering with the NLRP3 inflammatory signaling pathway, and reducing the recurrence rate of pterygium after surgery. Therefore, NLRP3 has become a potential therapeutic target to inhibit the expression of inflammatory factors, and reduce the recurrence rate of pterygium after surgery.
The expression level of NLRP6 was lower in recurrent pterygium than the primary pterygium, and higher in normal conjunctiva than pterygium. They are consistent and suggested that the low expression of NLRP6 is potentially not only involved in the formation, but also in the recurrence of pterygium. The possible reason is that NLRP6 plays a protective role in normal conjunctiva, and NLRP6 had an expression that is reduced due to inflammation in the pterygium. Recurrent pterygium inflammation is more obvious than primary pterygium inflammation, so the NLRP6 expression is further reduced. However, the mechanism of pterygium recurrence is complex, and our hypothesis needs to be confirmed by further prospective studies in a larger patient population. Therefore, NLRP6 may be a new therapeutic target to reduce the recurrence of pterygium.
Another finding of our study was, that there is a negative correlation between NLRP3 expression and NLRP6 expression in normal conjunctival and pterygium, which suggests the pro-inflammatory effect of NLRP3 may be related to the anti-inflammatory effect of NLRP6 in normal conjunctival tissues and pterygium tissues. Chi found that Cysteinase-1- dependent inflammasomes may inhibit the production and activity of NLRP6. 27 Therefore, we suggest the activation of NLRP3 may inhibit the activation of NLRP6 in normal conjunctival and pterygium tissues.
In our study the total number of samples is small. If we can collect more samples, we can better study the expression levels of NLRP3 and NLRP6. In addition to terms of correlation, NLRP3 is up-regulated and NLRP6 is down-regulated, but this study would be better able to confirm NLRP3 and NLPR6 expression if an indicator could be found to activate NLRP3 and inhibit NLRP6.
Above all, NLRP3 and NLRP6 may be involved in the pathogenesis and recurrence of pterygium. NLRP6 plays an anti-inflammatory role in normal conjunctival tissues to maintain conjunctival tissues homeostasis. The immune mechanism revealed in this study can provide a theoretical basis in the exploration for prevention and treatment methods of pterygium.
Footnotes
Ethical approval
This study was performed on the basis of the principles of the Declaration of Helsinki and approved by the Ethics Committee of Tianjin Eye Hospital.
Informed consent
Informed consent was obtained from all participants.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (grant number NO.81870638,81970772,81670817,81670816).