
Abstract
Glaucoma presents an epidemiological burden as the leading cause of irreversible blindness globally and the most common cause of preventable blindness. While elevated intraocular pressure is the strongest modifiable risk factor, the exact mechanisms of retinal ganglion cell damage leading to progressive vision loss are not entirely understood. Studies of other neurodegenerative diseases show a potential for human gut microbiome dysbiosis to play a pathogenic role. An investigation into whether the microbiome, a potential modifiable risk factor, has significance in glaucoma enables exploration of prophylactic or additive treatments. Elevated population levels of specific bacterial species have been noted in glaucoma patients, particularly Prevotellaceae, Enterobacteriaceae and Escherichia coli, while Megomonas is speculated to be protective. Evidence also points to systemic neuro-inflammation and disruption of autoimmune processes as a result of imbalances in both human and animal models, where heat shock proteins may contribute to pathogenesis. Further research into the influence of gut microbiome on pathogenesis offers a chance to minimise irreversible vision loss in glaucoma.
Introduction
Glaucoma, the leading cause of irreversible blindness globally and the commonest cause of preventable blindness, is a neurodegenerative condition that causes progressive and irreversible death of retinal ganglion cells (RGCs) 1 ; primary open angle glaucoma (POAG) is the most prevalent subset. 2 As neurones that relay visual signals to the optic nerve, loss of RGCs results in visual loss.
The pathways that lead to RGC stress and apoptotic cell death remain a mystery. As the primary modifiable risk factor, elevated intraocular pressure (IOP) is thought to contribute to RGC stress through mechanical stress on RGC axons and axonal transport at the lamina cribrosa, effects on retinal vascular supply, metabolic stress and mitochondrial dysfunction and the induction of pathological immune or autoimmune responses. 1 The significant proportion of normotensive glaucoma and subset of patients that progress despite apparent IOP control point to a more complex neurodegenerative pathogenesis to which raised IOP contributes. 3
What is the influence of the human microbiome in neurodegenerative disease? The gastrointestinal tract and oral cavity (the ‘gut microbiome’) host a vast number and diversity of microbiota, including millions of bacteria from thousands of bacterial species. 4 The composition and activity of the gut microbiome is dynamic. Important to age-related disorders such as glaucoma, it changes during life and responds to genetic and environmental exposures. 5 Multiple lines of evidence point to changes in the microbiota of individuals with neurodegenerative conditions such as Parkinson's and Huntington's diseases and animal models of these diseases. 5 Could a similar effect be seen in patients with POAG?
Autoimmunity and neuroinflammation are thought to induce cellular stress and contribute to the pathogenesis of POAG. 6 While the mechanisms for the influence of the microbiome in disease remain unclear, emerging evidence suggests the gut microbiome elicits autoimmune responses, raising the possibility that the gut microbiome may affect neurodegeneration of RGCs in a ‘gut-glaucoma axis’. As the microbiome is modifiable, this is an area to research for possible targeted treatments. We briefly review the evidence for neuroinflammation in POAG, links between autoimmunity and microbiome composition, and the nascent evidence for the contribution of microbiome dysbiosis to glaucoma pathogenesis.
Neuro-inflammation and autoimmunity in glaucoma
In order to understand the possible role of the microbiome in glaucoma, we need to understand the long-standing research into the role of autoimmunity. The presence and location of optic disc haemorrhage, a clinical sign of glaucoma progression, 7 indicates a breach in the normally otherwise impermeable blood-retina barrier and the infiltration of immune cells such as T cells. 8 Growing evidence of the presence of these cells, as well as components of the inflammatory complement system and Tumour Necrosis Factor Alpha (TNFα) in tissues surrounding the optic nerve head, support the notion that neuroinflammatory processes may contribute to glaucoma.8–10
In the absence of any infection, such neuroinflammation suggests an autoimmune process associated with the risk of tissue self-destruction. Self-antigen T helper (Th) cells produced by the thymus (with antigens for all the tissues within the body including the eye) are crucial to develop ‘central tolerance’ as a safeguard against autoimmunity. 11 Within the thymus, Th cells that express a high affinity to self-antigens are negatively selected and terminated, while those with a low affinity are converted to regulatory T (Treg) cells before being released into the circulation. 11 These Treg cells are responsible for ‘peripheral tolerance’ by killing or suppressing high-affinity self-reactive Th cells which have survived negative selection and escaped the thymus prior to conversion. 11
While the causes of these autoimmune changes in glaucoma remain obscure, both human and animal models suggest that disruption and changes to the microbiome of the gut may contribute.11–14 Understanding whether this relationship has a direct relevance to a human context may illuminate our understanding of immune-mediated processes in the progression of glaucoma.
The microbiome and glaucoma
Human microbiota, referring to bacteria, viruses and fungi which commensally live on or within a host organism,15–17 occupies niche areas such as the skin, oral and nasopharyngeal cavities, gastrointestinal tract and the urogenital area. 16 The gastrointestinal tract is the principal site, with the largest number and diversity of microbiota, including millions of bacteria from thousands of species. 4 These have a direct effect on pro-inflammatory Th cells within the gastrointestinal tract, including Th17 cells and their cytokine product interleukin 17 (IL-17), as well as Treg cells, which prevent inflammation through secretion of IL-10 among other factors. 18 For this literature review, we refer to the oral cavity and gastrointestinal tract as one collective niche area called the ‘gut’.
Microbiome
The gut microbiome is variable amongst individuals owing to factors such as ethnicity, genetics, diet, health, hygiene habits, age and gender.4,14 Evidence even points to long-term stress as an altering factor for gut microbiome. 19 A point of investigation lies in whether adjusting the gut microbiome may alter disease pathogenesis,4,20 especially as the microbiome is relatively modifiable with changes in diet, antibiotic treatment, and, in extreme cases, faecal transplants.21,22
Normally, the human gastrointestinal microbiome assists with digestion and produces some key nutrients such as vitamins B and K.4,22 Given that 70% of the immune system is present in the gastrointestinal tract, the microbiome plays an active role in its regulation. 20 Microbiota such as Clostridia and Bacteroides fragilis can enhance the differentiation of Treg cells in the colon,22,23 thereby supporting the immune system, particularly through the fermentation of fibre via production of short chain fatty acids (SCFA). 22 In order to function optimally, microbiota microbes require their constituent population to be balanced both in size and type, prompting the question as to whether an imbalance precipitates a problematic immune system with systemic pathological consequences.
Microbiome dysbiosis and autoimmunity
Gut microbiome dysbiosis, resulting from an imbalance in microbiota abundance and/or diversity,4,20 has been associated with various immune and neurodegenerative diseases. Such conditions include multiple sclerosis, rheumatoid arthritis, ankylosing spondylitis, atherosclerosis and diabetes.17,20,24 In ocular conditions, some evidence points to microbial dysbiosis being involved in non-infective uveitis11,25 and in microglial activation resulting in pathologies of the central nervous system. 26 Could the same dysbiosis play a role in neurodegenerative ocular conditions such as glaucoma?
Dysbiosis may increase intestinal permeability, releasing activated pathogenic Th cells, inflammatory factors and microbial products into the blood. Rosenbaum and Asquith postulate that the translocation of these products may also be involved in ocular autoimmune diseases, particularly in the case of acute anterior uveitis. 25 In another study, Rosenbaum et al. were able to induce acute anterior uveitis in rats by injecting bacterial endotoxins into their footpads. 27 Though the mechanisms of intestinal permeability changes due to dysbiosis remain unknown, a similar underlying process may occur in glaucoma. Blocking bacterial products originating in the gut from translocating to the eye may offer a means to avoid autoimmune conditions like glaucoma.
Metabolites from the microbiome appear able to enhance both protective Treg cells and pathogenic Th cells, depending on the microbiota and its metabolites, 22 through molecular mimicry of self-antigens. An example is Prevotella, which has been found to promote pathogenic Th cells in the gut.20,28 Furthermore, the gut microbiome and its metabolites have been shown to affect immunity by activating or inhibiting Toll-like receptors (TLR). For instance, lipopolysaccharide (LPS), a metabolite produced by microbiota such as Bacterioides and Escherichia coli is a known ligand for TLR-4, which affects complement activation. The role of microbes and TLR has been studied in Type 1 diabetes, and is demonstrated in Table 1.22,29

In a study investigating the effect of TLR-4 gene deletion in mice, colonisation of germ-free mice with a variety of intestinal bacteria resulted in an increased incidence of autoimmune diabetes. 29 This implies that TLR-4 has a protective role in autoimmune diabetes by binding to metabolites like LPS. 22 However, its activation has been shown to promote pathogenesis in glaucoma, 30 demonstrating the varying influences of the microbiome, their metabolites and the effects of immunity receptors from one disease to another. This variability is also demonstrated by the colonisation of TLR-2 gene knockout germ-free mice resulting in a reduction in autoimmune diabetes incidence, implying a pathogenic pathway instead. 29 The ability for microbiota to both promote or inhibit autoimmunity through different signalling pathways presents a complex problem in understanding the pathogeneses of these diseases.
Microbiome dysbiosis in neurodegenerative disease
The relationship between other neurodegenerative diseases and gut microbiome is worth considering in the context of glaucoma. Parkinson's disease (PD) and Alzheimer's disease (AD), two of the most prevalent neurodegenerative diseases, have been shown to be associated with changes in microbiome. 5 Mice overexpressing human alpha-synuclein were found to have more severe motor dysfunction when transplanted with the microbiome of human PD patients. 31 In AD models, germ-free reared mice expressing human transgenes for β-amyloid precursor protein, which is strongly associated with Alzheimer's, resulted in not only having a decrease in Aβ aggregates in the brain, but also a 40% reduction in activated microglial cells. 32
Perhaps the common denominator for AD, PD and glaucoma, however, is age and its effects on the microbiome demographic. For various reasons, chronic inflammation and changes to the intestinal barrier and its permeability often accompanies aging, 33 with associated changes in the microbiome. Germ-free mice models transplanted with the microbiome of old mice exhibited increased inflammatory cytokines and intestinal permeability, 34 suggesting that the microbiome substantially changes over time to the point it has tangible effects on physiology. As with PD and AD, age is a risk factor for the progression of glaucoma, and it may be that systemic inflammation with age plays a concurrent underlying role in the development of glaucoma.
Microbiome dysbiosis in glaucoma
Emerging evidence has demonstrated microbiome dysbiosis may be present in patients with glaucoma. The oral microbiome in African Americans has been reported to carry a significantly higher bacterial load in glaucoma subjects compared with sex and age-matched controls.30,35 Polla et al. found that Streptococcus mitis and Neisseria meningitides were significantly higher in the glaucomatous group. 35
Gong et al. analysed microbiota in stool samples of 30 POAG subjects with 30 controls. 24 Matched in age, sex, ethnicity (all from Guangzhou, China) and assumed to have similar diets, subjects were excluded if they had neurological, metabolic, or gastrointestinal diseases or other ocular conditions. While microbiome diversity was not significantly different, the prevalence of certain bacteria was. Prevotellaceae, unidentified Enterobacteriaceae, and Escherichia coli were dominant in the POAG group, while Megamonas and Bacteroides plebeius were dominant in the controls. Gong et al. speculate that Megamonas could be ‘protective’ for glaucoma. 24 Known to be crucial in energy homeostasis and cell metabolism, plasma citric acid was also elevated with Megamonas. Plasma citrate concentration may be a biomarker for glaucoma 36 ; its correlation with Megamonas may be significant. Gong et al. also noted that increased hypoxanthine in the POAG group negatively correlated with Megamonas. 24 Hypoxanthine has been linked with optic atrophy-related disorders. 37 Megamonas would seem worthy of further assessment. Between the two groups, imbalances in serum metabolites (20 increased, 15 decreased) supported microbiome dysbiosis in glaucoma. 24 lipopolysaccharide (LPS) from Escherichia coli was significantly prevalent in the POAG group 24 ; it is responsible for upregulating toll-like receptor 4 (TLR-4) and has been linked with local retinal inflammation and complement activation. 30 Table 2 shows the effects of low dose LPS injections on accelerating glaucomatous progression in mice models, which were designed to either develop or be protective against glaucoma. 30 Given LPS is a by-product of several different gut-associated gram negative microbes including Escherichia coli, 24 this microbe could become a target in glaucoma treatment.
Effects of LPS on glaucoma in mice models. 30
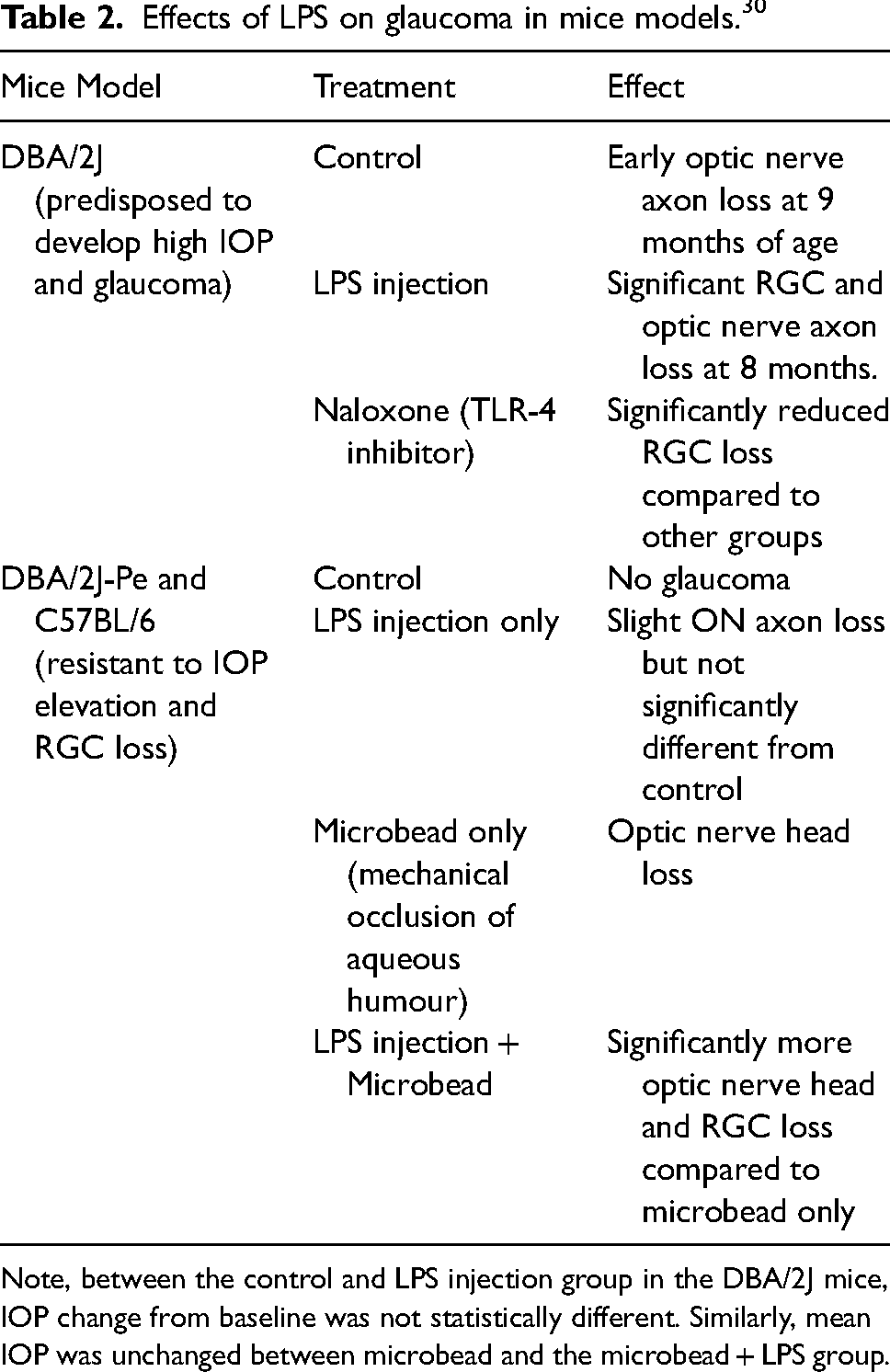
Note, between the control and LPS injection group in the DBA/2J mice, IOP change from baseline was not statistically different. Similarly, mean IOP was unchanged between microbead and the microbead + LPS group.
Glutathione, a major anti-oxidant metabolite, was also found to be reduced in the blood serum levels of glaucomatous rat subjects. 38 Reduced levels of glutathione correlated with significantly increased Bacteriodes and Romboutsia caecal populations compared to control subjects, and positively correlated with RGC loss. 38 A human study by Skrzypecki et al. also found in a cohort of glaucomatous patients significantly higher levels of aqueous trimethylamine, a toxic metabolite of gut bacteria, than their control counterparts. 39 In combination with a negative effect on the smooth muscle of the trabecular meshwork, 40 they hypothesise that trimethylamine may interfere with defence mechanisms against hypoxic stress and thereby accelerate glaucoma. 39 Another study examining the serum of glaucoma patients found that certain central carbon-related metabolites in the gut may have anti-oxidative properties protective against the development of glaucoma. 41 These metabolites were significantly reduced in glaucomatous patients, which also correlated with a significant decrease in populations of Blautia and Fusicatenibacter. 41 Therefore, these species may be protective.
In another study where Chen et al. elevated IOP in DBA/2J model mice, they reported CD4+ T cell markers in the ganglion cell layer. 12 Two weeks after IOP increase, there was a 17% loss in RGCs and axons, rising to 25% after eight weeks, five weeks after IOP normalisation. In contrast, transgenic mice deficient in T cells (Rag1 -/-) exhibiting similar loss initially to IOP elevation showed no ongoing loss of RGCs once the IOP had normalised. When CD4+ T cells from a glaucomatous mouse were injected into Rag1 -/- mice, further RGC and axon loss occurred, suggestive that T cell activity may be needed for continued pathology. The absence of glaucomatous disease in these germ-free transgenic DBA/2J mice models suggests a link between the gut microbiome and glaucoma. 12
Given the potential role for microbes and the immune system in prolonged RGC loss once elevated IOP has been normalised, this is an area of interest for investigation of glaucomatous progression in patients with normal pressure.1,12
Role of microglia and heat shock proteins in glaucoma
While known glaucomatous risk factors such as elevated IOP and metabolic stress including reactive oxygen species (ROS) can cause cellular damage particularly in the elderly with ‘normal’ IOP, 6 the role of microglia also appears to be significant. Both precursor monocytes and microglia are specialised macrophages in the central nervous system which play a crucial role in inflammatory responses and are increased in glaucomatous nerves. 42 Activated microglia present antigens such as heat shock proteins (HSP), a family of proteins produced in response to stressful stimuli, which in turn activate Th cells. 43 Intracellular HSPs appear to be protective, preventing stress-induced protein damage, whilst highly antigenic extracellular HSPs cause strong autoimmune reactions against infected or damaged cells.44,45 Increased microglial activity with HSP expression has also been described in other neurological pathologies such as PD and AD, 46 and may be important for autoimmune mechanisms in glaucoma.
In mice models, two particular HSPs, HSP27 and HSP60, found normally at low levels within ganglion cell layers, were upregulated three to four fold by IOP elevation. 12 In humans, HSP60 and HSP27 serum levels were elevated five-fold in patients with POAG and normal-tension glaucoma (NTG) compared with a control group. 12 HSP27 in particular provokes RGC apoptosis by destabilising the structural actin cytoskeleton. 47 Wax et al. also noted raised HSPs in the serum of glaucomatous mice subjects. 43 Neurodegeneration, from the production of inflammatory cytokine FasL by activated T cells, was able to be induced in mice immunised with HSP. 43
Microglia changed morphologically into a more active state after injection with lipopolysaccharide (LPS), 30 a metabolite from microbiota such as Escherichia coli, recently shown to be prevalent in the guts of patients with glaucoma. 24 lPS from gut microbiome can enable glaucomatous RGC loss in a multi-faceted manner through microglial activation and TLR-4 upregulation. 30 Furthermore, the gut microbiome may express its own HSPs, which can act as microbial mimics of self-HSPs, further enhancing the inflammatory pathway in a positive feedback loop. 6 It is this relationship between LPS, HSPs and microglia which makes this process important in the discussion of glaucoma progression.
Conclusion
Increasing evidence suggests a potential link between the gut microbiome, the immune system and glaucoma progression with or without elevated IOP. While the aetiology remains unknown, the influence of the microbiome on the inflammatory and immune processes systemically appears to extend beyond the enteric system. Despite evidence from mice models supporting this, it may not be relevant to a human context. Further human studies are needed to confirm the role or roles of the microbiome in glaucoma pathogenesis. Greater understanding of this subject might offer additional treatment options for patients with glaucoma both prophylactically and additively to minimise progressive irreversible vision loss.
Footnotes
Author's note
Matthew Krilis is also affiliated with Nuffield Department of Clinical Neurosciences, Oxford Eye Hospital, University of Oxford; Save Sight Institute, Sydney Eye Hospital, University of Sydney and Marrickville Eye Doctors, Marrickville, Sydney.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.