
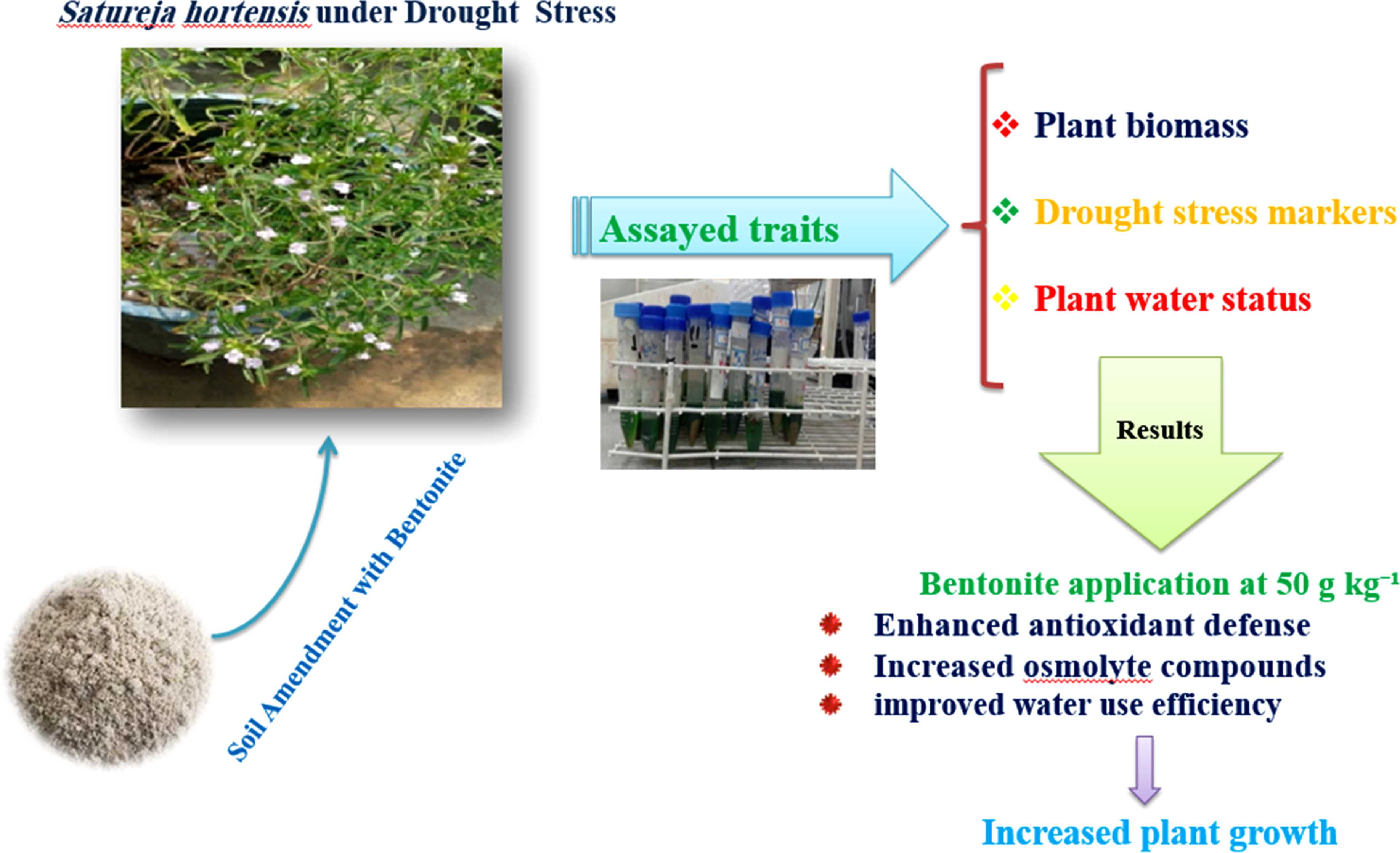
Abstract
Summer savory (Satureja hortensis L.) is a valuable annual herb sensitive to drought stress. This greenhouse study investigated whether amending soil with natural bentonite clay (0, 50, or 100 g kg-1) could mitigate the effects of three irrigation regimes (100, 75 and 50% of field capacity, FC). Growth, physiological and biochemical parameters were measured using a factorial based on completely randomized design to assess plant respons. Results showed that severe drought (50% FC) reduced shoot biomass by 25.8% and leaf relative water content (RWC) by 37.2%, while increasing root growth and oxidative stress markers (malondialdehyde by 50.8%). Bentonite amendment at 50 g kg-1 significantly alleviated these stress symptoms: it restored leaf RWC by 42.4% in stressed plants, increased the activity of key antioxidant enzymes (guaiacol peroxidase and polyphenol oxidase) by up to 45.5% and enhanced the uptake of essential nutrients (e.g., N, K, Zn). Consequently, this optimal bentonite rate improved the biomass of drought-stressed plants by up to 19.0%. The higher application rate (100 g kg-1) was less effective or even detrimental, likely due to reduced soil aeration. Our findings demonstrate that moderate bentonite application (50 g kg-1) effectively enhances drought tolerance in summer savory by improving soil water retention, nutrient availability and the plant’s antioxidant capacity. This strategy offers a practical and sustainable approach to cultivate this high-value medicinal herb under water-limited conditions.
Introduction
Satureja genus belonging to the Lamiaceae family is best known for its distinctive flavor. Summer savory (Satureja hortensis L.) as the most important species in this genus, is an annual crop that has been used as a vegetable and medicinal plant for a long time (Ejaz et al., 2023). The aerial parts of S. hortensis are harvested at the flowering stage and used for facilitating digestion, as a stomach tonic, carminative, diuretic, astringent, anti-diarrhea and anti-worm agent (Hagh et al., 2019). The therapeutic potential of this herb is attributed to its diverse phytochemical composition. Phenolic compounds (flavonoids, phenolic acids, tannins and pyrocatechols), gums, steroids and mucilage are the main components of S. hortensis leaf extract (Abou Baker et al., 2019). These bioactive constituents possess a high potential in treating serious ailments such as diarrhoeal, cardiovascular disorders, cancer and Alzheimer’s diseases (Ejaz et al., 2023). Furthermore, the essential oil of S. hortensis foliage is extensively used in food industries as a preservative and in beverages, cosmetic and pharmaceutical products due to its antioxidant, anti-inflammatory, antimicrobial and antifungal effects (Ejaz et al., 2023).
However, the growth and accumulation of these valuable secondary metabolites in plants are significantly influenced by environmental factors. Genetic and environmental factors can considerably affect plant growth with regard to morphological and physiological parameters (Chaki et al., 2020). Variation of environmental conditions can be the main cause of changes in plant morphological characteristics, which is able to affect a plant’s phenotype in a short time as well as a plant’s genotype in a long time (Estaji et al., 2018). Among various abiotic stresses, drought stress is considered the most important environmental stress, which reduces the growth and performance of plants by disrupting key physiological and biochemical processes (Chaki et al., 2020). A common occurrence under drought stress is the accumulation of reactive oxygen species (ROS) in plant cells. This ROS accumulation can play a fundamental role in reducing photosynthetic capacity due to the degradation of the photosynthetic system and chloroplast membranes (Khan et al., 2025). Additionally, stomatal closure and reduced transpiration under drought disturb plant water potential, leading to the inhibition of growth and ion uptake. These physiological alterations ultimately manifest as reduced plant height and total leaf area (Li et al., 2025).
To cope with the destructive effects of drought stress, plants employ a chain of defense mechanisms at the cellular level. Different defense mechanisms are involved in plant cells, including the activation of antioxidant enzymes and the synthesis of protective compounds as well as osmoregulators (Mergo & Ali, 2025; Mishra et al., 2023). Nevertheless, an imbalance between plant antioxidant capacity and ROS accumulation under prolonged or severe stress leads to oxidative stress, which damages cell membranes and other organelles (Afzal et al., 2024; Khan et al., 2025). Addressing the challenge of water scarcity is crucial for sustainable agriculture. Due to the limitation of water sources all over the world and the destructive effect of drought stress on crop performance, improving the soil’s ability to retain water, increasing water use efficiency and reducing evapotranspiration are essential strategies for sustainable plant production under drought stress conditions (Kamran et al., 2025; Nasser et al., 2016). In this context, the use of soil amendments has gained considerable interest. Recently, water-saving soil amendments such as superabsorbent polymers have attracted more attention for soil water management. It was reported that the application of superabsorbent increased the capacity of soil water holding and crops water use efficiency as well as reduced evapotranspiration (Mi et al., 2017). Beyond water retention, such amendments can improve overall soil health; also, the reduction of soil erosion by stabilizing soil structure is another benefit of soil amendments (Alqahtani et al., 2026). These effects are attributed to increasing soil aggregation and decreasing soil bulk density, which enhance soil porosity, water-holding capacity, soil fertility and ultimately crop growth (Kayama et al. 2016, 2020).
Bentonite represents a promising natural alternative to synthetic amendments. Bentonite, as a natural and non-toxic superabsorbent, is a phyllosilicate clay with high cation exchange capacity (CEC) that has been used in various industrial applications such as sealing well casings and sorption of heavy metals (Lerma, Chamorro, & Palencia, 2024). For agricultural use, its mode of action involves absorbing moisture from irrigation or precipitation and preventing its leaching (Ramesh & Raghavan, 2025). Under water-limiting conditions, the trapped moisture in bentonite is gradually released, making water available to the crop for a longer period (Nasser et al., 2016). Despite its potential, the performed researches specifically examining the effect of bentonite on the physiological and biochemical responses of plants under controlled drought stress is limited. Therefore, this study was designed to address this knowledge gap. The present study was conducted to evaluate the S. hortensis response with regard to morphological, biochemical and yield characteristics to different levels of water limitation and to determine the possible effect of bentonite on the mitigation of the destructive effects of drought stress. By investigating a range of physiological and biochemical markers, this research aims to provide a mechanistic understanding of how bentonite amendment can enhance drought tolerance in S. hortensis plant.
Materials and Methods
Plant Material and Experimental Design
This study was performed to investigate how the addition of bentonite clay influences the growth and physiological responses of summer savory subjected to water deficit conditions. The research took place in a greenhouse setting at the Faculty of Agriculture, Ferdowsi University of Mashhad, which is situated at an elevation of 985 meters (coordinates: 36° 18' N, 59° 31' E).
The experimental design was a factorial arrangement using a completely randomized design with three replicates. The treatments combined three irrigation regimes (100, 75 and 50% of field capacity, FC) with three rates of bentonite amendment (0, 50 and 100 grams per kilogram of soil). These specific treatment levels were chosen based on prior screening tests that assessed the plant’s tolerance.
The plants were grown in plastic pots (30 cm diameter, 40 cm height) filled with a sandy loam substrate. The soil’s physicochemical properties were as follows: pH 7.7, containing 70.2% sand, 21.1% silt and 9.0% clay, with an electrical conductivity (EC) of 1.3 dS m-1. Its nutrient profile included 0.2% total nitrogen, 61 mg kg-1 of available phosphorus and 184 mg kg-1 of available potassium. The predetermined amount of bentonite was thoroughly incorporated into the soil before planting. Seeds of summer savory were sterilized in a 5% sodium hypochlorite solution and then sown. After germination, seedlings were thinned to maintain six uniform plants per each pot.
Plant Material, Experimental Design and Growth Conditions
The plants were grown in a greenhouse to control environmental variables. The average daily climatic conditions inside the greenhouse throughout the experiment were as follows: daytime temperature of 25 ± 2 °C, nighttime temperature of 18 ± 2 °C and relative humidity ranging from 60 to 80%. Natural sunlight was the light source. These conditions remained consistent, with minimal weekly variability, as the greenhouse environment was regulated. Therefore, all treatment groups experienced the same climatic background, ensuring that observed differences were due to the irrigation and bentonite treatments.
Irrigation Management and Stress Imposition
Soil moisture was monitored daily using a Time-Domain Reflectometer (TDR; Spectrum Technologies Inc.). Pots were weighed periodically to corroborate the TDR readings and ensure accuracy in maintaining the target soil water levels (100, 75 and 50% FC). The field capacity (100% FC) was determined as the stable water content after saturating the pots and allowing free drainage for 48 hours while covered to prevent evaporation. The 75 and 50% FC levels were then calculated as proportional reductions from this weight. Irrigation was applied whenever the TDR reading or pot weight indicated a drop below the target threshold, restoring the soil to its predetermined weight for the specific FC level. This method ensured the soil moisture levels were maintained within a defined range. The three soil moisture levels—100%, 75% and 50% FC—were designated as the control, moderate drought stress and severe drought stress conditions, respectively. The water source for irrigation was tap water with an EC of 1.01 dS m-1 and a pH of 8.2. The total duration of the study, from sowing to final harvest at the flowering stage, was approximately 75 days. The drought stress treatments (maintenance of soil moisture at 75 and 50% FC) were initiated once seedlings were fully established and were maintained until plant harvesting time.
All measurements and plant samplings for subsequent laboratory analyses were conducted at the flowering stage of the summer savory plants.
Cation Exchange Capacity of Soil and Mineral Nutrient of Plant
The CEC of soil was determined based on the ammonium acetate method at pH 7 (Schollenberger & Dreibelbis, 1930). The oven-dried leaves of S. hortensis were used to assay the macro and micro nutrient analysis of the leaves. The dry leaf samples (0.3 g) were powdered and homogenized with 5 mL H2SO4. The solution was placed in a digesting apparatus for 2 h. After cooling, 20 mL Na2OH and 30 mL HBO3 were added and then titrated with H2SO4. According to consumed H2SO4, the nitrogen content of samples was calculated (Bremner, 1996).
For assay the phosphor content of leaves, the samples were powdered and burned at 450 °C for 4 h. The cooling ashes were digested in 10 mL HCl 0.1 N. The solution was heated at 80 °C for 10 min and added distilled water to the final volume of 50 mL. Then the absorbance was read at 660 nm (Kuo, 1996). Potassium content was measured by flame photometry (Waling et al., 1989). After calibration with relevant standard solutions, the concentration of Mg2+, Ca2+, Fe2+, Zn2+ and Mn2+ was determined by ICP (ICP-OES).
Total Phenol Content
Total phenol content was measured by the Folin-Ciocalteu reagent (Singleton & Rossi, 1965). Briefly, the leaf samples were extracted with 99% methanol at room temperature and centrifuged at 12,000 g for 10 min. The supernatant (200 μL) was diluted with 300 μL of distilled water and subsequently, 2.5 ml of freshly prepared 50% Folin-Ciocalteu reagent was added. The solution was incubated in darkness for 3 min at room temperature. After that, 2 mL of 7.5% sodium carbonate solution was added. The absorbance of the solution was read at 765 nm. Different concentrations of gallic acid were used as a standard and the total phenol content was expressed as mg of gallic acid equivalent per g leaf fresh weight.
Soluble Protein and Antioxidant Enzyme Activities
For enzymatic analysis, 0.5 g of plant tissue was homogenized in 5 mL of an ice-cold 50 mM potassium phosphate buffer (pH 6.8). The extraction buffer was supplemented with 1% (w/v) polyvinylpyrrolidone (PVP) and 1 mM EDTA to inhibit polyphenol complexation and chelate metal ions, respectively. The resulting homogenate was centrifuged at 12,000 × g for 20 minutes at 4°C. The supernatant was collected and served as the enzyme extract for subsequent assays. The soluble protein concentration in this extract was quantified according to the Bradford (1976) method, using bovine serum albumin (BSA) to generate the standard curve.
Guaiacol peroxidase (GPX, EC 1.11.1.7) activity was determined by monitoring the oxidation of guaiacol (Plewa et al., 1991). The assay mixture, with a total volume of 3 mL, contained 2.77 mL of 50 mM potassium phosphate buffer (pH 7.0), 100 μL of 2% (v/v) guaiacol, 100 μL of 1% (v/v) hydrogen peroxide (H2O2) and 30 μL of the enzyme extract. The increase in absorbance at 470 nm, resulting from the formation of tetraguaiacol was recorded spectrophotometrically for 3 minutes. Enzyme activity was calculated using the extinction coefficient for tetraguaiacol (25.5 mM-1 cm-1) and expressed in units (U) per mg of soluble protein, where one unit corresponds to the amount of enzyme required to produce 1 mmol of tetraguaiacol per minute.
The activity of polyphenol oxidase (PPO, EC 1.14.18.1) was assayed using pyrogallol as the substrate (Kar & Mishra, 1976). A reaction mixture containing 2.5 mL of 20 mM phosphate buffer (pH 6.8) and 200 μL of 2 mM pyrogallol was pre-incubated in a water bath at 40°C. The reaction was initiated by adding 100 μL of the enzyme extract. The change in absorbance at 430 nm was measured over a 4-minute period. Specific activity was determined using the molar extinction coefficient for the oxidation products of pyrogallol (2.47 mM-1 cm-1) and is reported as U mg-1 protein, with one unit defined as the oxidation of 1 mmol of pyrogallol per minute.
Antioxidant Activity
The antioxidant capacity of the leaf extracts was evaluated based on their ability to scavenge the stable free radical, 2,2-diphenyl-1-picrylhydrazyl (DPPH). Initially, plant extracts were obtained by dissolving the leaves in 99% methanol. For the assay, the methanolic extract was mixed with 4 mL of a freshly prepared DPPH solution in methanol (40 mg L-1). This mixture was kept in darkness at ambient temperature for a 30-minute reaction period. Following this incubation, the absorbance of both the plant sample and a control solution (containing only the DPPH reagent) was measured at a wavelength of 517 nm (Moon & Terao, 1998). The radical scavenging activity, expressed as a percentage of DPPH inhibition, was calculated using the formula:
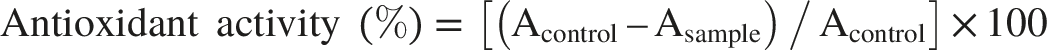
Soluble Sugars
To measuring the content of soluble sugars, 0.5 g of fresh leaves was homogenized with 5 mL of 99% methanol. The supernatant (0.2 mL) was mixed with 3 mL anthrone (150 mg anthrone, 100 mL of 72% sulphuric acid, W/W). The samples were placed in a boiling water bath for 10 minutes. After cooling samples, the absorbance was estimated at 625 nm (Paquin & Lechasseur, 1979). Contents of soluble sugars were determined using glucose standard and expressed as mg g-1 FW of leaves.
Malondialdehyde Accumulation and Electrolyte Leakage
The extent of lipid peroxidation in leaf tissues was quantified by measuring the concentration of malondialdehyde (MDA), a key byproduct of polyunsaturated fatty acid degradation (Davey et al., 2005). Following the procedure, 0.3 g of fresh leaf material was homogenized in 3 mL of 0.1% (w/v) trichloroacetic acid (TCA). The homogenate was centrifuged at 10,000 × g for 20 minutes at 4°C. A 0.5 mL aliquot of the resulting supernatant was then reacted with 300 µL of a solution containing 0.5% (w/v) thiobarbituric acid (TBA) and 20% (w/v) TCA. The mixture was incubated at 90°C for 30 minutes to develop the MDA-TBA adduct. The reaction was terminated by rapidly cooling the samples in an ice bath. The absorbance of the supernatant was measured at 532 nm, with a correction for non-specific turbidity performed by subtracting the absorbance at 600 nm. The MDA concentration was calculated using an extinction coefficient of 155 mM-1 cm-1 and reported as nanomoles of MDA per gram of fresh leaf weight (nmol MDA g-1 FW).
Membrane stability was evaluated by measuring electrolyte leakage from leaf discs (Teutonica et al., 1993). Leaf sections (2 cm in length) were prepared, rinsed with distilled water to remove surface ions and placed in test tubes containing 10 mL of distilled water. The tubes were kept in darkness for 24 hours at room temperature, after which the initial electrical conductivity (EC1) of the bathing solution was recorded. The samples were then autoclaved at 121°C for 20 minutes to release all intracellular electrolytes. Once the solutions had cooled to room temperature, the final electrical conductivity (EC2) was measured. The percentage of electrolyte leakage (EL), representing membrane damage, was calculated using the formula:
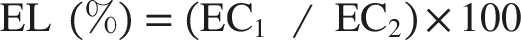
Transpiration Rate and Relative Water Content
The transpiration rate of fully expanded leaves from the upper canopy of S. hortensis plants was measured with a portable infrared gas analyzer (IRGA; LCA-4, ADC). To ensure consistent and maximal physiological activity, all measurements were taken during a one-hour window between 11:00 AM and 12:00 PM under saturating light conditions.
The hydration status of the leaves was assessed by determining their Relative Water Content (RWC). Following the standard protocol, leaf samples were first weighed to obtain their fresh weight (FW). To ascertain their turgid weight (TW), the same leaves were fully hydrated by floating them on distilled water for 24 hours at room temperature and then weighed again. Finally, the samples were oven-dried at 75°C for 48 hours to determine their dry weight (DW). The RWC was calculated using the formula:
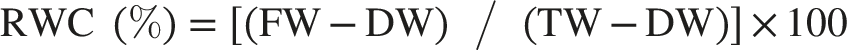
Photosynthesis Pigments
Chlorophylls and carotenoids were extracted from leaves using 99% methanol. After centrifuging at 12,000 g for 5 min, the absorbance of the solution was measured by spectrophotometer at 470, 653 and 666 nm. The contents of chlorophyll a, b and carotenoid were estimated using the equations proposed by Dere et al. (1998) and expressed as mg of each pigment per g leaf fresh weight.
Plant Growth and Biomass
At the flowering stage, plant height, length of intermediate and branch, number of nodules and branches, stem diameter and leaf area as well as length and width of leaves were measured (three plants of each pot were randomly selected). In addition, three plants were removed from each pot and the number, length and volume of roots were recorded. Aerial parts and root were separated and the dry weight of them was determined by drying each organ at 80 °C for 48 h.
Statistical Analysis
Data were subjected to analysis of variance using the statistical analysis program (Minitab ver.17.0) and the mean values were compared by Bonferroni test at P ≤ 0.05. The figures were drawn by the Excel program.
Results
Cation Exchange Capacity of Soil and Mineral Nutrient of Plant
Effect of Bentonite on Cation Exchange Capacity of Experimental Soil and Mineral Contents of S. hortensis Leaves Under Different Levels of Irrigation Regimes
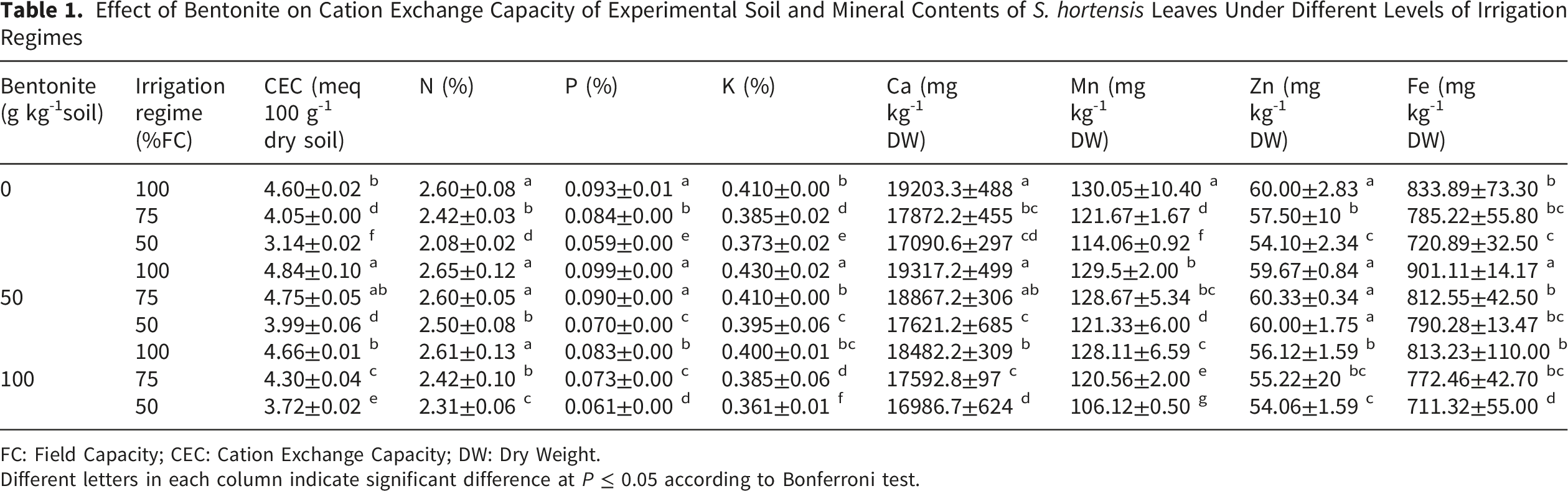
FC: Field Capacity; CEC: Cation Exchange Capacity; DW: Dry Weight.
Different letters in each column indicate significant difference at P ≤ 0.05 according to Bonferroni test.
Analysis variance showed that the irrigation regimes and bentonite application significantly affected the content of mineral elements of summer savory leaves. The interaction effect of both treatments was significant on all studied elements with exception of Mg. With increasing the water limitation, the contents of N, P, K+, Ca2+, Mn2+, Zn2+, Fe2+ (Table 1) and Mg2+ (Figure 1A) were decreased and the most prominent effect of drought stress was observed at 50% FC. It is interesting that the content of mineral elements of non-stressed plants was not influenced by bentonite application. In drought-stressed plants, application of bentonite at 50 g kg-1 increased the content of studied elements. However, bentonite at 100 g kg-1 did not show a significant effect on cations content compared with the untreated conditions. The content of all studied elements in grown plants under 75% FC and bentonite application was similar to their content in full-irrigated plants. Also, the content of N, K+ and Zn2+ in severely stressed plants that treated with 50 g kg-1 bentonite did not differ from those contained in unstressed plants (Table 1). Effect of different levels of irrigation regimes (A) and bentonite (B) on magnesium content of S. hortensis leaves. Bars represent mean ± SE (n=3). Different lowercase letters above bars within a panel indicate significant differences among treatments according to Bonferroni post-hoc test following two-way ANOVA (two-sided test). The specific P-values for the comparisons indicated by letters are as follows: (A) between 100% FC and 50% FC: P = 0.003; between 75% FC and 50% FC: P = 0.012. (B) between 0 and 50 g kg-1 bentonite under drought: P = 0.021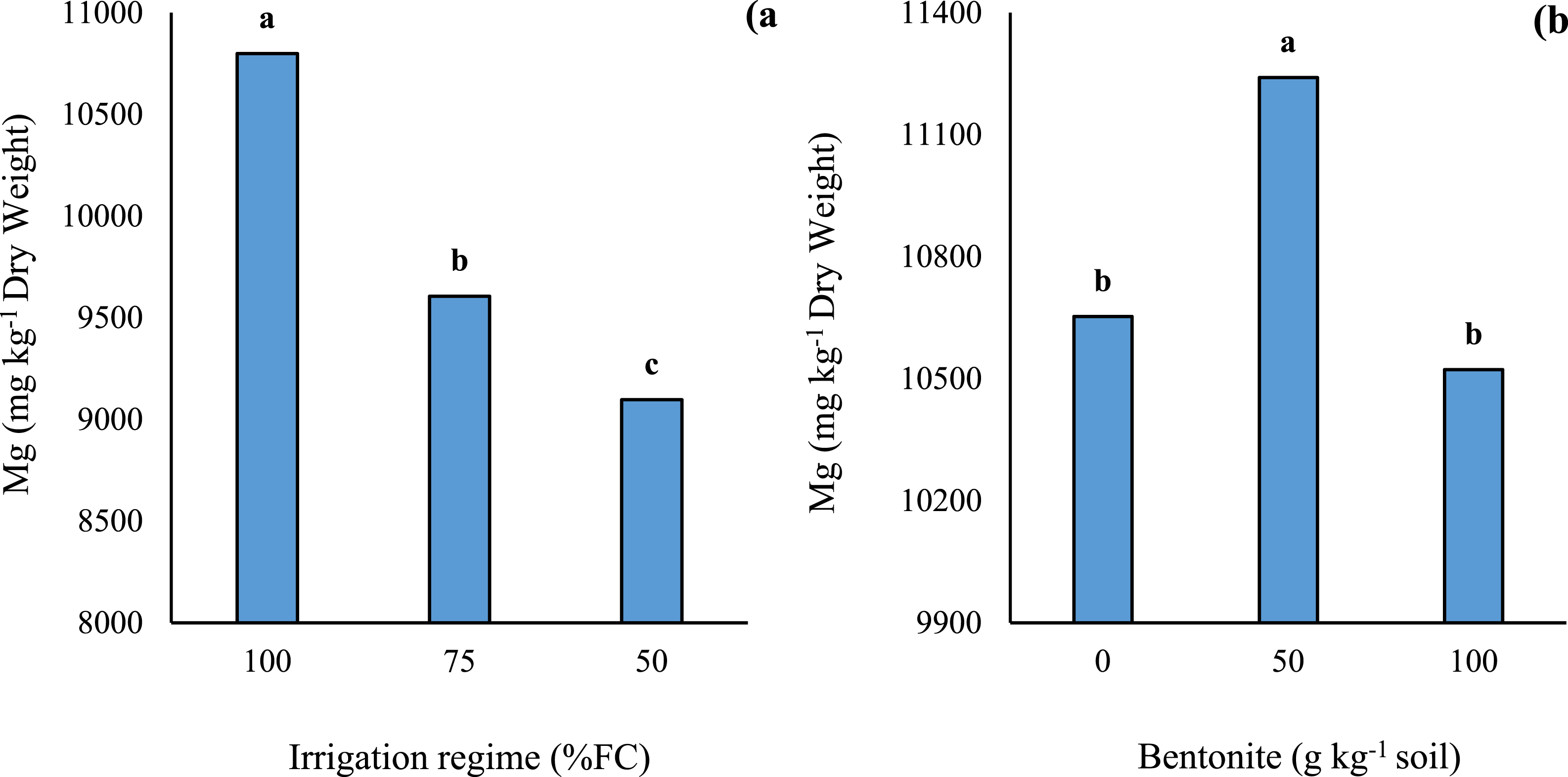
Total Phenol Content and Antioxidant Activity
Effect of Bentonite on Some Physiological Characters of S. hortensis Under Different Levels of Irrigation Regimes
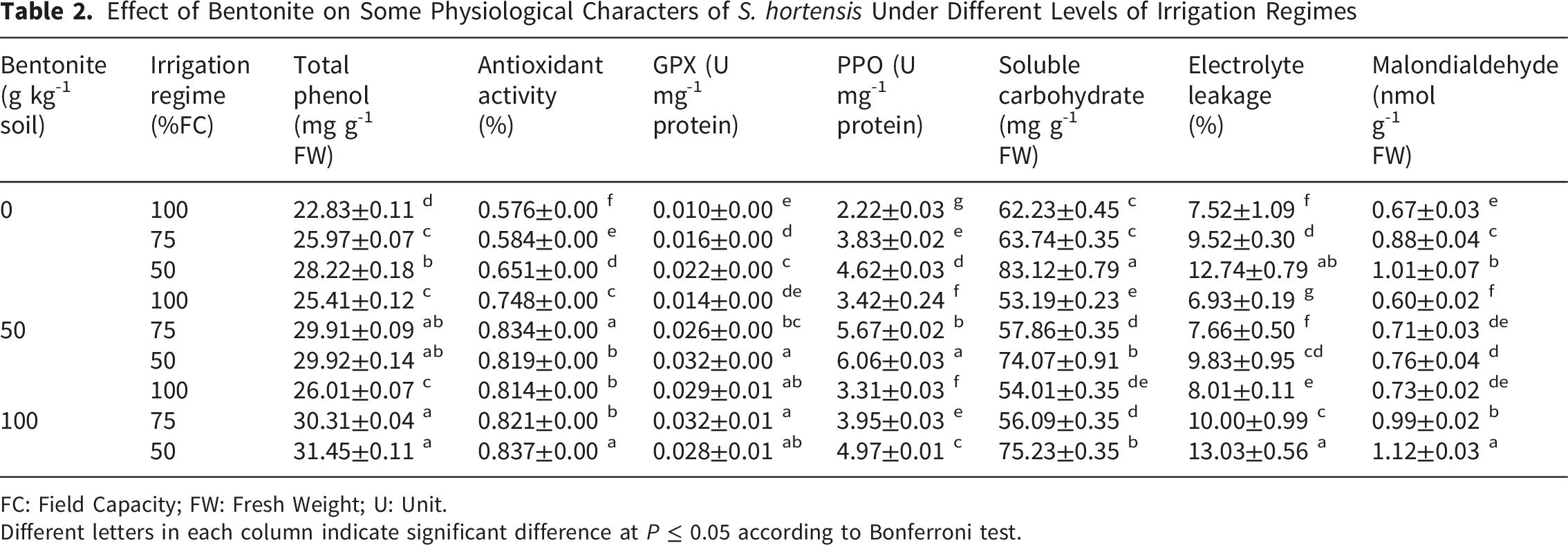
FC: Field Capacity; FW: Fresh Weight; U: Unit.
Different letters in each column indicate significant difference at P ≤ 0.05 according to Bonferroni test.
Antioxidant capacity of summer savory extract regards antioxidant activity, PPO and GPX activity significantly influenced by interaction effects of irrigation regimes and bentonite application. Water limitation increased antioxidant activity up to 13.02% and its content was increased by 41.32% in response to bentonite application. The effective concentration of bentonite was dependent on drought stress intension. The application of bentonite increased antioxidant activity by 42.81 and 28.57% in moderate and severe drought-stressed plants, respectively (Table 2). PPO and GPX activity significantly increased with decreasing available water and under severe drought stress, their activity was two times more than unstressed plants. Bentonite application increased PPO activity in unstressed and stressed plants. PPO activity in treated control, moderate and severe drought-stressed plants by 50 g kg-1 bentonite was increased up to 54.05, 48.04 and 31.17%, respectively. GPX activity in control and moderately stressed plants treated with 100 g kg-1 bentonite was two times higher than untreated plants. Under severe drought stress, bentonite application at 50 g kg-1 increased GPX activity by 45.45% compared with control (Table 2).
Protein and Soluble Sugars Content
Protein content was decreased in response to drought stress by 7.85% compared with control (Figure 2A). Bentonite application especially at 100 g kg-1 slightly enhanced the protein content but was not significant (Figure 2B) also the interaction of studied treatments was not significant on protein content. Effect of different levels of irrigation regimes (A) and bentonite (B) on protein contents of S. hortensis leaves under. Bars represent mean ± SE (n=3). Different lowercase letters above bars within a panel indicate significant differences among treatments according to Bonferroni post-hoc test following two-way ANOVA (two-sided test). The specific P-values for the comparisons indicated by letters are as follows: (A) between 100% FC and 50% FC: P = 0.047. (B) No statistically significant differences were found between bentonite treatments within the same irrigation regime (all P > 0.05)
Soluble sugar content was increased up to 33.57% under severe water deficit stress. Bentonite application decreased soluble sugar content in stressed and unstressed plants and there was no significant difference between 50 and 100 g kg-1 (Table 2).
Electrolyte Leakage and Malondialdehyde Accumulation
Electrolyte leakage and MDA increased with increasing stress intension by 69.41 and 50.75%, respectively. Bentonite application at 50 g kg-1 decreased electrolyte leakage by 40.55% in drought-stressed plants. However, applied bentonite at a high amount (100 g kg-1) significantly increased electrolyte leakage in unstressed and stressed plants (Table 2). Applied bentonite at 50 g kg-1 decreased MDA content by 10.45 and 24.75% in the control and stressed plants, respectively. Similar to EC, bentonite in 100 g kg-1 intensified the drought stress effect on the MDA accumulation in S. hortensis plants (Table 2).
Transpiration Rate and Relative Water Content
Effect of Bentonite on Relative Water Content, Transpiration Rate and Photosynthesis Pigments of S. hortensis Under Different Levels of Irrigation Regimes
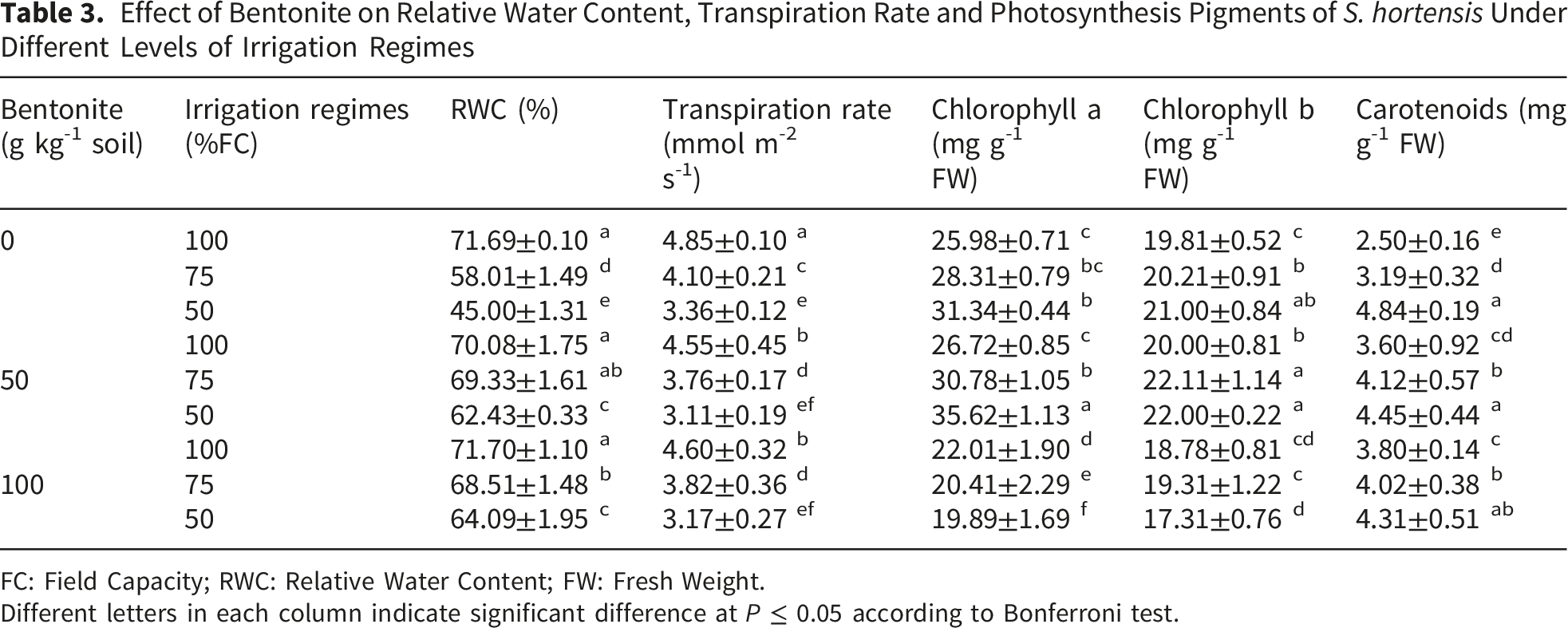
FC: Field Capacity; RWC: Relative Water Content; FW: Fresh Weight.
Different letters in each column indicate significant difference at P ≤ 0.05 according to Bonferroni test.
RWC of S. hortensis leaves significantly reduced under severe drought-stressed plants by 37.23%. However, the RWC of unstressed plants was not affected by bentonite but its application effectively increased RWC in stressed plants by 42.42% and there was no significant difference between 50 and 100 g kg-1 (Table 3).
Photosynthesis Pigments
In the present study, photosynthesis pigments including chlorophyll a, b and carotenoids increased with decreasing available water by 20.63, 6.01 and 93.6%, respectively. The contents of chlorophyll a and b were slightly increased by bentonite application at 50 g kg-1 but its application at 100 g kg-1 decreased chlorophylls content in unstressed and stressed plants (Table 3). Carotenoid content in unstressed and stressed plants was significantly increased in response to bentonite application and there was no significant difference between applied bentonite concentrations (Table 3).
Plant Growth and Biomass
Effect of Bentonite on Morphological Characters of S. hortensis Under Different Levels of Irrigation Regimes
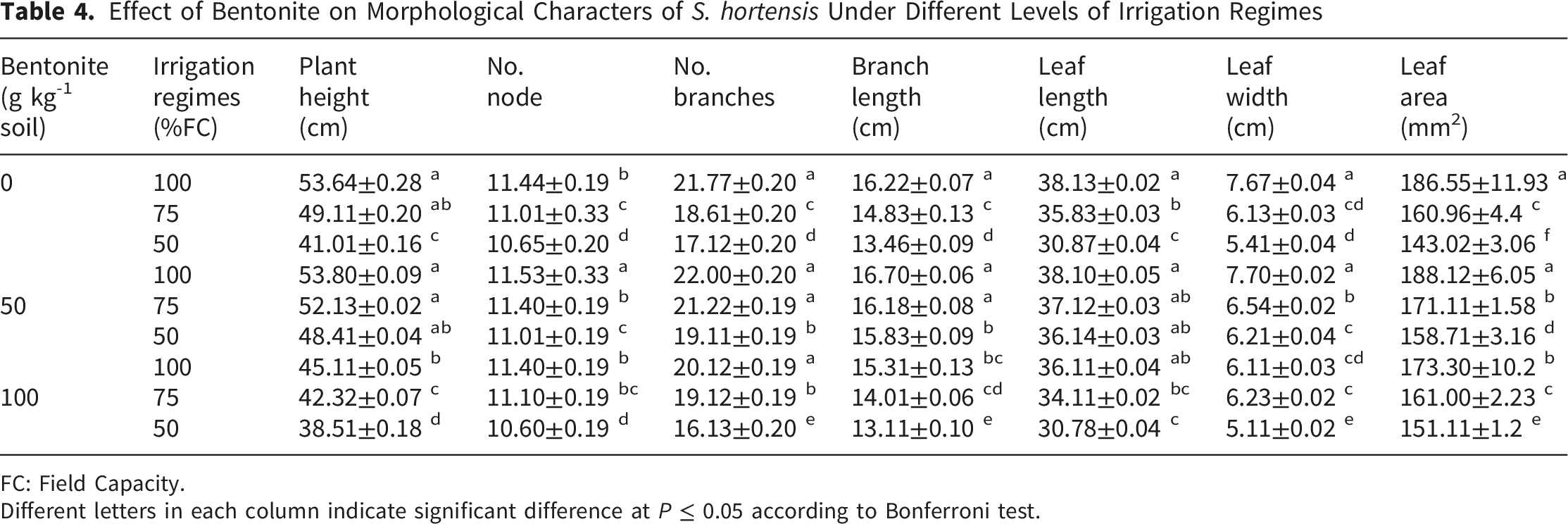
FC: Field Capacity.
Different letters in each column indicate significant difference at P ≤ 0.05 according to Bonferroni test.
Effect of Bentonite on Plant Biomass and Root Growth of S. hortensis Under Different Levels of Irrigation Regimes
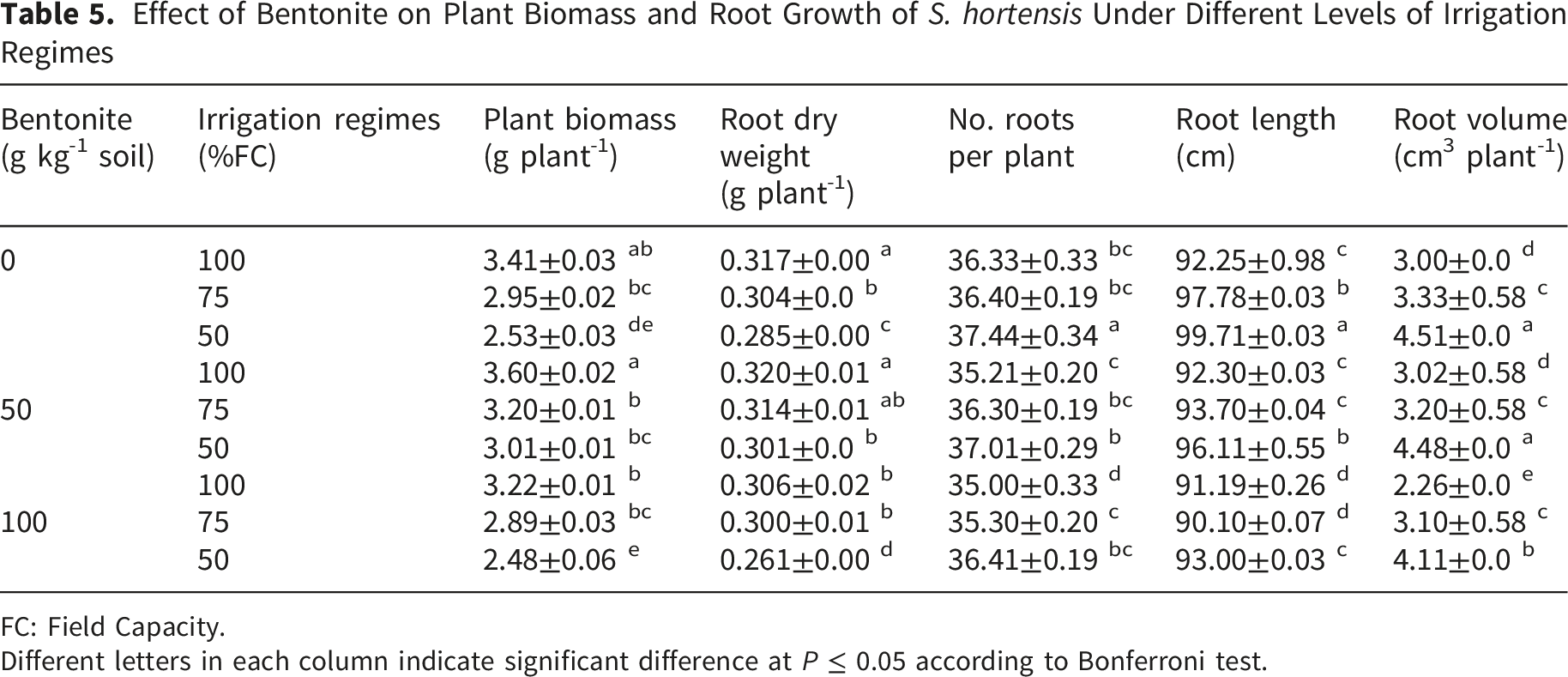
FC: Field Capacity.
Different letters in each column indicate significant difference at P ≤ 0.05 according to Bonferroni test.
Plant biomass was decreased in response to drought stress by 25.81%. Plant biomass of unstressed plants was slightly increased in response to bentonite application at 50 g kg-1. Also applied bentonite (50 g kg-1) significantly increased biomass of stressed plants up to 18.97% (Table 5).
Discussion
The significance of soil amendment strategies in arid and semi-arid regions cannot be overstated, as they directly address the primary constraint of water scarcity. Bentonite clay, as investigated in this study, represents a promising amendment due to its innate ability to modulate the soil physical environment and, consequently, plant physiological responses. Unlike studies that solely report growth yield, our research delineates the specific physiological and biochemical pathways—including nutrient homeostasis, water relations and antioxidant metabolism—through which bentonite exerts its protective effects under drought. This mechanistic understanding is crucial for optimizing its use in sustainable agriculture.
Cation Exchange Capacity of Soil and Mineral Nutrient of Plant
In the present study, soil CEC decreased under water stress (Table 1) due to desertification and degradation processes, which reduce the soil’s ability to retain and exchange cations (Meng et al., 2023; Zhou et al., 2019). This reduction indicates that drought causes not only a physical water deficit but also a chemical fertility crisis (Alqahtani et al., 2026; Kamran et al., 2025; Lerma, Chamorro, & Palencia, 2024). According to obtained results bentonite application increases soil CEC, especially under water shortage (Table 1), by improving soil structure and providing more exchange sites for nutrient retention (Kayama et al., 2016; Lerma, Chamorro, & Palencia, 2024; Mergo & Ali, 2025; Mi et al., 2017).
Similar to present study, the reduction of macro and micro elements in the drought-stressed plant (Table 1) has been reported in different plant species (Bista et al., 2018; Tadayyon et al., 2017). It has been recognized that nutrient uptake under drought stress depends on water uptake by plants. Water shortage reduces nutrient uptake by the roots and their transport to the shoots (Khan et al., 2025). In addition, decrement of nutrient elements in leaves of the drought-stressed plants may be related to decreasing soil microbial activity (Meng et al., 2023), involved enzyme activity in nutrient assimilation and expression of nutrient-uptake proteins in roots (Bista et al., 2018). Recently Bista et al. (2018) reported that the decreasing of nutrients elements especially N and P under drought stress is attributed to the reduction of concentration or activity of the main uptake-proteins of N and P names NRT1, AMT1 and PHT1. Our data on S. hortensis align with this molecular-level explanation, as the reduction in leaf N and P (Table 1) corresponded with the severity of water deficit, suggesting a downregulation of nutrient uptake mechanisms (Alqahtani et al., 2026). The increasing of cations in water-stressedS. hortensis plants in response to bentonite application is consistent with other researches (Ramesh & Raghavan, 2025; Zhou et al., 2019). Zhou et al. (2019) reported that applied bentonite in the soil of the semi-arid region increased the plant access to essential cations and facilitated the cations uptake by the plant. Bentonite belongs to the 2:1 clay family that can enhance soil structural stability through rearrangement, flocculation and cementation of soil particles (Lerma, Paradelo, & Palencia, 2024). The high swelling potential and water-absorbing capacity of bentonite reduce soil evaporation and store water within the silicate interlayer then release slowly when crops need it (Nasser et al., 2016). This character of bentonite improves cations mobility under the low water content of soils (Nasser et al., 2016). Also, the change of physicochemical characteristics of the soil under bentonite treatment was previously reported by Haider et al. (2019). The larger surface of bentonite increases the cation exchange capacity (CEC) of the soil and bentonite cations such as Si4+, Al3+, Ca2+ and Fe3+ form bridges between clay and soil organic matter particles that play a key role in the enhancement of soil holding capacity, soil fungal hyphae formation and fortification of soil (Haider et al., 2019; Ramesh & Raghavan, 2025).
Therefore, the principal role of bentonite in nutrient dynamics appears to be indirect but critical: by mitigating the physical drought stress and improving soil CEC, it sustains the soil-plant continuum necessary for nutrient transport and uptake, thereby preventing the typical drought-induced nutrient deficiencies observed in un-amended soils.
Total Phenol Content
Under water stress, phenolic compounds can either increase or decrease depending on plant species. In some plants, phenols increase as they act as antioxidants scavenging hydroxyl and peroxyl free radicals (Abou Baker et al., 2019; Chaki et al., 2020; Khan et al., 2025; Li et al., 2025; Mishra et al., 2023). However, in the present study the phenolic compounds of stressed S. hortensis were decreased under water limitation (Table 2) as reported in Salvia dolomitica (Caser et al., 2019). The reduction of phenolic compounds under stress conditions can be related to their degradation in reaction to oxidative compounds (Osmolovskaya et al., 2018).
The significant increase in total phenols with bentonite application, even under drought (Table 2), suggests a novel bio stimulant-like effect. Unlike studies focusing solely on drought induction of phenolics, our results indicate that bentonite creates a soil environment that potentiates this biosynthetic pathway. It is hypothesizes that this is a consequence of improved plant vitality and resource allocation, as evidenced by better nutrient status (Table 1), rather than a direct chemical induction by bentonite (Ramesh & Raghavan, 2025). These changes lead to the increasing in plant protective potential in morphology and physiology aspects.
Antioxidant Activity
The S. hortensis antioxidant capacity with antioxidant enzymes activity (GPX and PPO) showed a progressive increase with increasing water limitation (Table 2), which is a typical response of plants to unfavorable environmental condition (Mishra et al., 2023) and adapted strategies by the plants to cope up with stress condition (Bista et al., 2018; Shariat et al., 2018). Biosynthesis of a broad array of drought-protective proteins, predominantly antioxidant enzymes is upregulated under drought stress, which play a central role in detoxification of ROS (Mishra et al., 2023). Increasing the activity of the antioxidant enzymes under drought stress could be the consequence of increasing ROS generation and activating the plant cell-protective mechanism to reduce oxidative damage (Khan et al., 2025; Li et al., 2025). The antioxidant activity shows the plant’s potential to savaging the ROS and protecting the plant’s survival under different conditions which including the antioxidant enzymes and compounds (Mishra et al., 2023). Therefore, in the present study increasing the antioxidant activity of S. hortensis extract (Table 2) was related to increasing GPX and PPO activity and total phenol content.
As results showed bentonite application increased antioxidant activity and GPX and PPO activity (Table 2). The increasing antioxidant capacity of treated plants with bentonite was reported by Snioszek et al. (2018); Haider et al. (2019). The responsible mechanism for this enhancement of antioxidant activities in response to bentonite application is due to the fundamental characteristics of bentonite such as the provision of plant essential nutrients, having a larger surface area and high cation exchange capacity (Haider et al., 2019). Thus, bentonite does not merely alleviate drought stress passively by holding water; it actively enhances the plant’s inherent capacity to manage oxidative stress. This synergistic enhancement of both enzymatic (GPX, PPO) and non-enzymatic (phenolics) antioxidant systems marks a key advantage over inert soil conditioners and aligns with the observed reduction in oxidative damage markers (MDA and EL).
Protein Content
In the present study, the protein concentration was decreased under drought stress (Figure 2A) which attributed to reduced N assimilation, decreased protein synthesis and increased protein degradation from oxidative damage (Bista et al., 2018; Chaki et al., 2020; Mishra et al., 2023). Although in the present study, the effect of bentonite on protein content was not significant (Figure 2B) but Snioszek et al. (2018) reported that bentonite application by supplying water source for nutrient uptake prevents degradation and decreasing of protein content. The discrepancy among the results of the literature is related to differences in the studied plant species (Kayama et al., 2016), plant growth stage (Kayama et al., 2016), applied concentrations (Paradelo et al., 2019), growth media (Zhou et al., 2019) and environmental conditions (Nasser et al., 2016).
Soluble Sugars Content
Under drought conditions, the accumulation of the soluble sugar content was increased in S. hortensis (Table 2). The increment of these compounds in plant cells can provide a situation for the continuance of water absorption from root zone. Metabolic adjustment is a critical mechanism in maintaining the water status and physiological activity of plant cells, especially during water deficit conditions. Accumulation of metabolically neutral and osmotically active solutes such as different sugars, amino acids (proline and glycine) and organic acids under drought conditions is well documented in different plant species (Dbira et al., 2018; Meng et al., 2023; Shariat et al., 2018). However, osmotic regulation based on these organic compounds has its own cost and consumes a part of the plant’s energy (Templer et al., 2017). Under water limitation, a large number of soluble carbohydrates should be used for osmotic regulation which causes a reduction of plant growth. However, some plant species avoided the accumulation of such costly compounds for osmotic regulation (Osmolovskaya et al., 2018).
In the present study the lower accumulation of soluble sugars in bentonite-amended, drought-stressed plants (Table 2) was a result of mitigation of drought stress intensity through increasing the soil water content and antioxidant activity of plants which prevent the production of high-cost compounds such as soluble sugars that consume a large part of plant reduction power (Templer et al., 2017). This trade-off—reduced investment in osmotic adjustment coupled with increased investment in antioxidant defense—illustrates how bentonite helps re-optimize the plant’s metabolic budget under drought.
Electrolyte Leakage and Malondialdehyde Accumulation
In the present study as water deficit stress was intensified, membrane permeability significantly decreased therefore the highest electrolyte leakage was observed in the lowest available water (Table 2). The reduction of cell volume under water limitation increases the viscosity of cytoplasmic components and denaturation of protein membrane. That disruption of cell membrane increases electrolyte leakage (Osmolovskaya et al., 2018). The main cellular components susceptible to damage by free radicals are lipids which peroxidation of unsaturated fatty acids in membranes increased MDA accumulation (Meng et al., 2023) as observed in the present study.
The reduction of MDA levels and EC in treated stressed S. hortensis plants with bentonite may be due to increment in the antioxidant enzyme activity and phenolic compounds (Table 2) as reported by Snioszek et al. (2018) and Haider et al. (2019). Increasing the antioxidant defense system by trapping the lipid alkoxyl radical quenches lipid peroxidation so decreases the fluidity of the membranes (Chaki et al., 2020). In addition, Haider et al. (2019) reported that higher mobility of essential nutrients to plants under bentonite treatment improves plant vigor and thereby reduces the contents of reactive oxygen species and their damages.
Transpiration Rate and Relative Water Content
Decreasing the transpiration (Table 3) through stomata closure is one of the main plant strategies to avoid water loss that results in the enhancement of the water status of plant cells (Kapoor et al., 2020). Water deficit stress occurs when the amount of water lost through transpiration exceeds the amount taken up by the roots, meaning the plant’s water requirements cannot be fully met (Li et al., 2025). In this condition, plants attempt to preserve their hydric status by morpho-physiological and biochemical adjustments to counteracting the loss of water. Bentonite application, decreased transpiration rate in treated S. hortensis plants (Table 3). It seems that soil amendment with superabsorbent compounds along with decreasing plant transpiration rate provides plant water requirement under different levels of drought stress (Kayama et al., 2016; Mi et al., 2017).
Relative water content is a suitable indicator of plant water status and a relevant screening tool for drought tolerance. Under water deficit stress, the plant cell lost intracellular water due to cellular dehydration (Table 3). In this condition, organic compounds are accumulated in plant cells (Table 2) to prevent cell degradation (Table 2) without disturbing intracellular biochemistry (Khan et al., 2025). In the present study, it seems that increasing antioxidant capacity (Table 2) regards to antioxidant enzyme activity and total phenol content as well as accumulation of soluble sugars content (Table 2) prevented the reduction of RWC in stressed S. hortensis plants. Increasing RWC of S. hortensis in response to bentonite application under moderate and severe stress condition (Table 3) may be related to the mineralogical structure of bentonite. The characteristic allows the absorbing and storage of a large amount of water under sufficient water conditions and slowly releases water into the soil under drought condition for plant uptake (Mi et al., 2017; Paradelo et al., 2019; Zhou et al., 2019).
Photosynthesis Pigments
In the present study photosynthesis pigments were increased under induced drought stress (Table 3). The plant potential of photosynthetic activity can be quantitated by the determination of pigments including chlorophylls and carotenoids. In consistency with our results, Mohammadi et al. (2017) reported the increase of chlorophyll content in S. hortensis stressed plants under 50% FC. Dbira et al. (2018) reported that higher chlorophyll content in water-stressed plants is generally associated with higher plant drought tolerance. An additional increase of photosynthesis pigments can be related to photochemical quenching of excess light energy by increased photorespiration (Osmolovskaya et al., 2018). Reduction of chlorophyll level under drought stress, which was reported in S. rechingeri by Shariat et al. (2018) is considered as a symptom of oxidative stress due to photo-oxidation and chlorophyll degradation (Li et al., 2025).
Carotenoids as photosynthesis pigments absorb light and pass the captured energy to the chlorophylls (Chaki et al., 2020). Carotenoids have additional roles that help plants to withstand drought stress and considered as the first line of plant defense that able to detoxify various forms of ROS. Increasing the carotenoid concentration in water-stressed plants releases excessive energy by thermal dissipation to the amplification of the antioxidant defense system (Chaki et al., 2020).
Increasing photosynthesis pigments in response to bentonite application as reported in the present study (Table 3), is due to enhancement of nutrient supply (Table 1) (Ramesh & Raghavan, 2025; Zhou et al., 2019), protein synthesis (Snioszek et al., 2018), antioxidant potential (Haider et al., 2019), osmotic compounds (Table 2) (Haider et al., 2019; Snioszek et al., 2018) and relative water content (Table 3) (Kayama et al., 2016). The observed increasing in chlorophyll and carotenoids in our study, particularly with bentonite at 50 g kg-1 (Table 3), supports the view of S. hortensis as a drought-tolerant species that can maintain photochemical capacity under moderate stress. Bentonite likely aids this by protecting the photosynthetic apparatus from oxidative damage, as reflected in lower MDA levels.
Plant Growth and Biomass
The reduction of plant growth metrics under drought (Table 4) is the integrated morphological outcome of the physiological limitations discussed above, primarily reduced cell turgor and metabolic diversion towards stress defense. To preserve water supply under water stress conditions, plants adopt various strategies to prevent water loss to maintain their growth during unfavorable water regimes (Osmolovskaya et al., 2018). These drought-induced alterations can affect plant physiology and biochemistry characters including antioxidant enzyme activity, accumulation of compatible solutes and phenolic compounds (Table 2), which can take intensive energy and resource from the plant. These changes are accompanied by the reduction of plant morphology (Table 4) in degree depending on plant species, developmental stage, duration and intension of water limitation (Shariat et al., 2018).
Bentonite application at appropriate concentration (50 g kg-1) enhanced plant growth of S. hortensis. Similar results were reported by Kayama et al. (2016); Snioszek et al. (2018); Haider et al. (2019). However, it seems that the application of extra bentonite (100 g kg-1) reduces the available oxygen in the plant growth media due to the maintenance of more water content, which subsequently results in growth reduction as was observed in the present study (Table 4).
Increasing root number and length is the essential plant strategy for avoidance of drought stress, which enhances plant ability to absorb more water from soil depth (Khan et al., 2025; Osmolovskaya et al., 2018). However, the reduction of root biomass in water-stressed plants of S. hortensis (Table 5) was previously reported by Mohammadi et al. (2017). The discrepancy with literature is related to differences in plant species, growth condition, duration and intensity of water stress. Many studies have shown that superabsorbent could enhance soil infiltration capacity, reduce soil surface runoff, prevent water erosion and thereby can improve soil ecosystem (Lerma, Chamorro, & Palencia, 2024; Nasser et al., 2016). Stimulation of root growth of maize in response to bentonite application was reported by Zhou et al. (2019) which can relate to the existence of sufficient available water for plant growth, increasing nutrient uptake and reduction of surface evaporation (Haider et al., 2019). Decreasing root growth under a high concentration of bentonite may be related to the rarity of oxygen in amendment soil.
Decreasing of S. hortensis biomass (Table 5) could be associated with the reduction of mineral elements (Table 1), RWC (Table 3) and plant growth (Table 4). Mohammadi et al. (2017) reported that drought reduced both shoot and root dry weight in S. hortensis, but the effect was more severe on shoots. The increase of plant biomass in response to bentonite application (Table 5) was consistent with Mi et al. (2017) who showed that bentonite application significantly increased millet crop performance under arid and semi-arid conditions. These authors reported that increasing crop yield under bentonite application was due to the improvement of soil water storage capacity, reduction of nutrient loss, increasing cation exchange capacity, enhancement of water and nutrients absorption. According to Zhou et al. (2019) reports, the enhancement of plant biomass under bentonite application was related to increasing soil water storage and the ability of roots to access water stored in the soil.
Comparative Advantages and Practical Implications of Bentonite
Generally, compared to other common soil amendments used for drought mitigation, such as organic compost or synthetic superabsorbent polymers, bentonite offers a distinct profile of advantages and limitations. Its primary advantages are its natural origin, high cation exchange capacity, longevity in soil without rapid decomposition and cost-effectiveness for large-scale application. Crucially, our study identifies a non-linear dose-response, with 50 g kg-1 being optimal. This rate delivers maximum benefit for water retention and nutrient availability while avoiding the soil aeration problems often associated with high application rates of fine-textured amendments—a common drawback noted in other studies (Haider et al., 2019; Kamran et al., 2025; Snioszek et al., 2018). A key limitation is that bentonite provides no direct nutritional value, implying that its greatest agronomic potential may be realized in combination with fertilizers. While synthetic superabsorbent polymers can absorb more pure water, they raise concerns regarding environmental persistence, cost and potential toxicity to plants. For long-term soil health in arid regions, natural, mineral-based amendments like bentonite are therefore a more sustainable and reliable choice.
Conclusion
This study demonstrates that soil amendment with natural bentonite clay, particularly at an optimal rate of 50 g kg-1, is an effective strategy for mitigating drought stress in summer savory. The efficacy of bentonite is rooted in a dual mechanism: modifying the soil environment to enhance resource availability and priming the plant’s internal physiological defenses. Our key finding—the identification of a specific, optimal application rate—carries significant practical importance. While studies on crops like millet and maize have shown the general benefits of bentonite (Mi et al., 2017; Zhou et al., 2019), our research on S. hortensis defines the threshold for benefit. The 50 g kg-1 rate successfully improved soil water retention and cation exchange capacity, leading to better plant water status (RWC) and enhanced uptake of essential nutrients like N, K and Zn under drought. Crucially, this rate avoided the negative effects on soil aeration and growth suppression observed at the higher 100 g kg-1 rate, a drawback noted in other pot studies with fine-textured amendments (Haider et al., 2019). This provides a clear and evidence-based guideline for application. Furthermore, our results extend beyond agronomic parameters to elucidate the physiological mechanism behind the improved resilience. The obtained results confirm that drought triggers a strong antioxidant response in S. hortensis, consistent with findings in related species like S. rechingeri (Shariat et al., 2018). By alleviating the primary water deficit stress, bentonite appears to allow the plant to allocate resources more efficiently, resulting in a coordinated boost in both enzymatic (GPX, PPO) and non-enzymatic (phenolic compounds) antioxidant activity. This enhanced biochemical capacity directly translated into reduced oxidative membrane damage (lower MDA and electrolyte leakage), a key factor in maintaining growth under stress.
Therefore, the applied significance of this work is twofold. First, it proposes a practical and sustainable intervention for cultivating high-value medicinal herbs in water-limited environments. Second, it shifts the understanding of bentonite from a simple soil conditioner to a plant stress-response modulator that operates by improving soil physics and synergistically enhancing the plant’s own biochemical defenses. For future research, field trials are essential to validate this optimal rate under real-world conditions and different soil types. Additionally, investigating the synergistic effect of combining bentonite with organic fertilizers or bio stimulants could address its lack of direct nutrition and may further amplify its beneficial role in sustainable arid-land agriculture.
Footnotes
Author Contributions
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
Data will be made available on request.