
Abstract
Introduction
The cause of multiple sclerosis (MS) is unknown and evidence suggests a complex and multifactorial aetiology. Chronicity and progression of the disease are characteristic features, making it difficult to study with autopsy material alone, where the majority of lesions are end-stage. Fortuitous demonstration of demyelination in biopsied brain lesions provides a unique opportunity to study early-stage demyelination. For example, a number of biopsy-based studies have suggested that pathological heterogeneity of demyelination may exist between MS patients. 1 – 3
Endoplasmic reticulum (ER) stress-mediated neurodegeneration has been proposed in Alzheimer’s, 4 Parkinson’s 5 and prion diseases. 6 We have shown that elevated levels of the ER stress pathway proteins, C/EBP homologous protein (CHOP), immunoglobulin heavy chain binding protein (BiP) and X-box-binding protein-1 (XBP1) are strongly associated with MS lesions. Furthermore, in a proportion of the patients studied, activation of an ER stress response in active lesions coincided with raised levels of D-110, an established marker of hypoxia. 7,9
A third of proteins synthesized by eukaryotic cells are trafficked through the ER, destined for secretion or insertion into cellular membranes. An oxidizing environment within the ER lumen supports the activity of protein-folding enzymes, assisted by ER chaperone proteins. 10 Disturbances in redox or calcium homeostasis in the ER trigger a cellular response known as the unfolded protein response (UPR), centred on the activation of the PKR-like endoplasmic reticulum kinase (PERK), inositol-requiring enzyme (IRE1) and activating transcription factor 6 (ATF6). Key identifiers of UPR activation are increased expression of CHOP and BiP. 10,11 The UPR is generally a pro-survival mechanism, mediated by translation arrest and the induction of a number of transcription factors and chaperone proteins that function to restore the ER to its normal physiological state. However, if ER stress persists and protective mechanisms fail, switches to a pro-apoptotic phase are thought to involve activation of CHOP and Jun-kinase. 12,13
In order to determine if ER stress and/or hypoxia signalling could be relevant to events in early demyelination, we examined the expression of CHOP and BiP in seven biopsy samples obtained from three patients with clinically proven MS and four with an inflammatory demyelinating episode of unknown aetiology. Since Aboul-Enein et al. first reported hypoxic-like oligodendrocyte damage in MS lesions 14 and the subsequent detection of a marker of hypoxia in a quarter of biopsy and acute MS tissue samples by Lucchinetti et al., 2 further confirmed by our recent study of post-mortem tissue samples, 7 we also assessed biopsy samples for the presence of the hypoxia marker D-110.
While our previous molecular analysis of immunohistochemical data, obtained using MS tissue, provided a crude indication that alterations in ER stress gene transcription had occurred, 7 in the second phase of the work reported here we exploited laser capture microdissection and real-time PCR technology in order to quantify the gene expression of ER stress transcription factors CHOP, activated transcription factor 4 (ATF4) and the ER chaperone BiP. Previous work from our group has demonstrated the feasibility and advantages of using laser capture microdissection to isolate cerebral blood vessels from MS tissue for gene expression analysis. 15 In the present study, differential expression of the ER stress genes in five different categories of MS tissue were compared with levels seen in white matter from normal controls.
Materials and methods
Biopsy study
Clinical details of patients from demyelinating biopsy series and post-mortem tissue used in laser capture microdissection
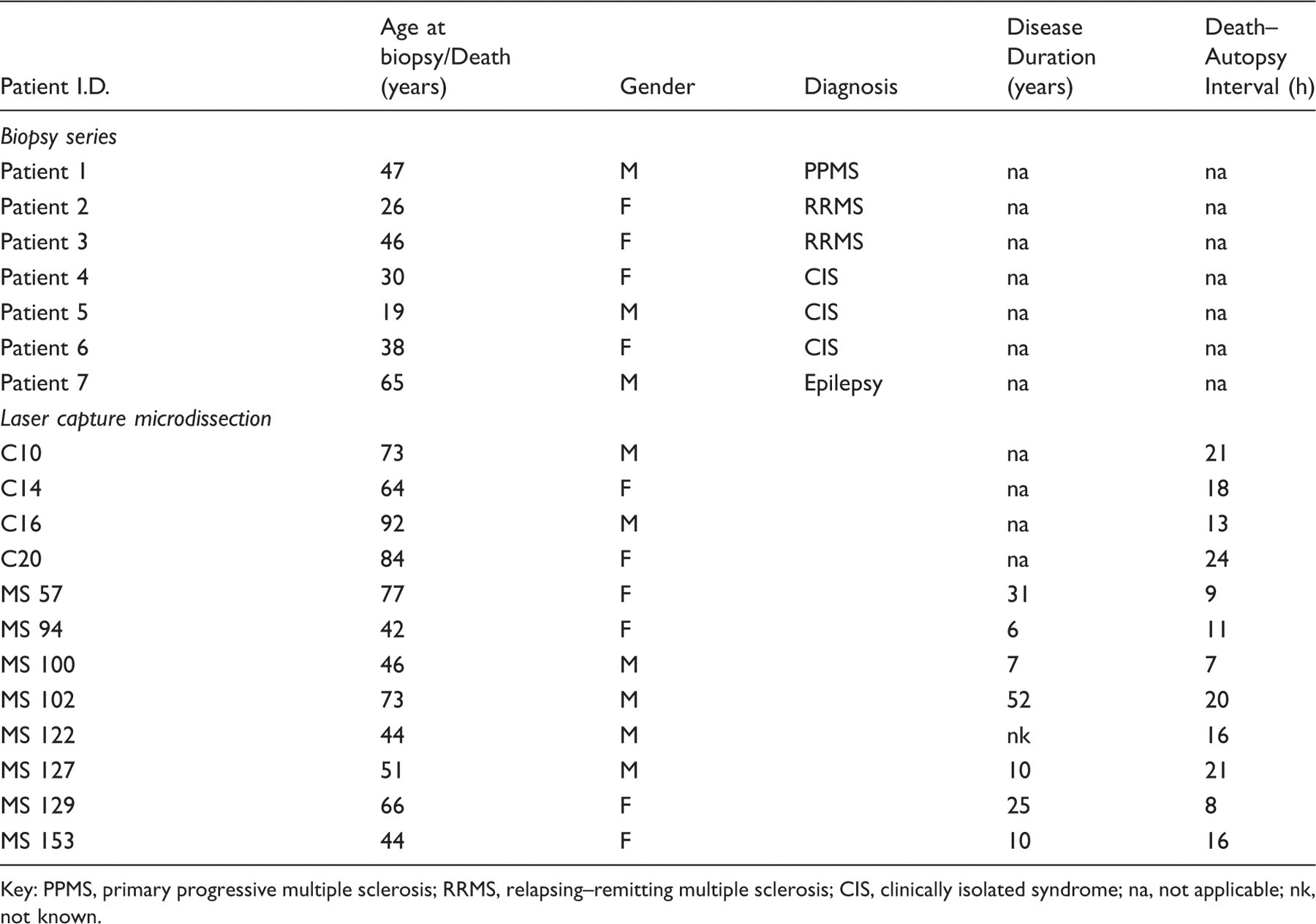
Key: PPMS, primary progressive multiple sclerosis; RRMS, relapsing–remitting multiple sclerosis; CIS, clinically isolated syndrome; na, not applicable; nk, not known.
Antibodies against ER stress-related proteins were used to detect CHOP (sc793, 1:400, Santa Cruz Biotechnology Inc, Heidelberg, Germany) and BiP (ab32618, 1:200, Abcam, Cambridge, UK). The presence of hypoxia in tissue sections was identified using an antibody to the D-110 epitope (1:2000, kindly supplied by Professor Hans Lassmann, Medical University of Vienna, Vienna, Austria). Sections were pre-treated, incubated and immuno binding sites detected as previously described. 7 A semi-quantitative analysis of antigen expression was performed on serial sections. The following scoring system was applied: 0 = no antigen expression, 1 = low expression (present in 0–30% of cells); 2 = moderate expression (30–60%) and 3 = abundant expression (60–100%). In addition to cell-specific staining, a diffuse pattern of D-110 staining was occasionally observed and was assigned a score of 1. In order to describe inherent variation in expression of these molecules within lesion-positive sections, three different anatomical areas were assessed: lesion centre including edge (LC/E), zone of myelin destruction, and perilesional white matter (PL), comprising white matter surrounding the lesion. Selected sections were digitally scanned using an Aperio Scanscope T3 (www.aperio.com) with a × 40 objective and were then displayed at a range of magnifications. Images of immunostained slides (×200 magnification) from one of the clinically isolated syndrome (CIS) cases (patient 6) were obtained using an Olympus BX51 microscope and Olympus DP70 colour camera, in conjunction with Cell B software.
Tissue cohort for laser microdissection study
Clinical details of cases used in the laser microdissection study are listed in Table 1. Three 10 µm sections from each of eight blocks per tissue category (control white matter, normal-appearing white matter (NAWM), active and chronic plaque) were microdissected. For each tissue sub-region, the eight blocks used were derived from four or five different patients. Frozen tissue was supplied by the UK Multiple Sclerosis Tissue Bank (London).
Rapid staining of tissue prior to microdissection
Cryostat sections of 10 µm were cut and collected onto MMI RNase-free membrane slides (Mason Technology, Dublin, Ireland) before fixing in ice-cold ethanol and storing at −80°C. After thawing, sections were immersed in 70% ethanol for 1 min followed by Mayer’s Haematoxylin (TCS Biosciences Ltd, Buckingham, UK) for 45 s. After de-staining with DEPC-treated milli-Q water for 45 s, sections were immersed in 70% ethanol for 30 s. Sections were then stained with MMI kit Eosin (Mason Technology, Dublin, Ireland), diluted 1:2, for approximately 5 s. Following a 30 s incubation in 70% ethanol and two immersions in 100% ethanol for 1 min each, sections were air-dried. Sections were stained immediately prior to laser microdissection to minimize RNA degradation.
Laser capture microdissection
To aid localization of lesions within the rapidly stained sections, parallel sections used for initial block characterization were observed. These had been stained by H&E and Oil-Red-O or MOG, as previously described. 7 To reduce possible degradation of RNA, sections were laser microdissected and tissue was collected within 30 min of staining. Tubes with adhesive caps (Mason Technology, Dublin, Ireland) were used to collect the microdissected tissue and were then processed for RNA extraction. Due to low yields of RNA generated following single-region sampling, each microdissected tissue sub-region was collected from 2–3 serial sections from each tissue block and pooled. This resulted in eight samples extracted from each sub-region (control white matter, NAWM, active plaque perilesion, active plaque, chronic plaque perilesion and chronic plaque) for RNA analysis. Figure 3a depicts section staining before and after removal of targeted sub-region.
RNA isolation from laser microdissected tissue
The QIAGEN RNeasy Micro kit was used to extract RNA from the microdissected tissue sections according to manufacturer’s instructions and RNA concentration and integrity was measured as previously described. 15
cDNA synthesis and real-time PCR
The QIAGEN Quantitect cDNA synthesis kit (Qiagen Ltd., West Sussex, UK) was used to generate cDNA according to the manufacturer’s instructions. Real-time PCR was carried out using the SYBR green method, according to Applied Biosystems’ guidelines. The primer sequences used for ER stress and housekeeping genes were, respectively: CHOP forward primer, 5′− agctggaacctgaggagaga −3′; CHOP reverse primer, 5′− tggatcagtctggaaaagca −3′; ATF4 forward primer, 5′− ccactaggtaccgccagaag −3′, ATF4 reverse primer, 5′− gccttgcggacctcttctat −3′; BiP forward primer, 5′− gccgtcctatgtcgccttc −3′, BiP reverse primer, 5′− tggcgtcaaagaccgtgttc −3′; 18s forward primer, 5′− cctgcggcttaatttgactca −3′, 18s reverse primer, 5′− agctatcaatctgtcaatcctgtcc −3′.
Each real-time PCR reaction contained 0.5 µl of cDNA, forward and reverse primers at 300 nM final concentration and Power SYBR green PCR master mix (Applied Biosystems), and was run on the ABI 7000 (Applied Biosystems). Results were calculated using the ΔΔCt method, graphs plotted using Microsoft Excel and statistical analysis carried out using GraphPad Prism. Groups were compared using the Mann–Whitney non-parametric test. p-values of less than 0.05 were considered significant.
Results
Characterization of biopsy tissue
At clinical follow-up, patient 7 was diagnosed with lesional epilepsy, after a seizure-induced isolated demyelination episode, while patients 1–3 were confirmed as primary progressive MS (patient 1) or relapsing–remitting MS (patients 2 and 3). Patients 4–6 were diagnosed with CIS, presenting with a single attack, showing one demyelinating lesion on magnetic resonance imaging (MRI) analysis. Digital Aperio scans of biopsy sections from the patient with primary progressive MS, the epilepsy patient, and one patient with CIS (patient 6), are shown in Figures 1 and 2.
Expression of endoplasmic reticulum stress proteins and hypoxia marker in biopsy material from case of primary progressive multiple sclerosis (patient 1). Characterization of demyelinating activity was carried out using LFB/H&E stain for myelin (A: 0.65 × magnification) and presence of microglia/macrophages determined using HLA-DR immunostaining (B: 0.65 × magnification). Actively demyelinating lesion centres ( Expression of endoplasmic reticulum stress proteins and hypoxia marker in biopsy material from cases of lesional epilepsy (patient 7) and clinically isolated syndrome (patient 6). Panels A (LFB/H&E) and B (HLA-DR) show 1× Aperio scans of sections from the case of lesional epilepsy which demonstrate the overall pathology in the biopsy tissue and allow demarcation of demyelinated and perilesional areas. Panels C–K show CHOP (C–E), BiP (F–H) and D110 (I–K) immunostaining from lesional material in clinically isolated syndrome (C, F, I), and perilesion (D, G, J) and lesion centres (E, H, K) in epilepsy. Scale bars: A, B = 1 mm; C–K = 200 µm. * = lesion centre; ▾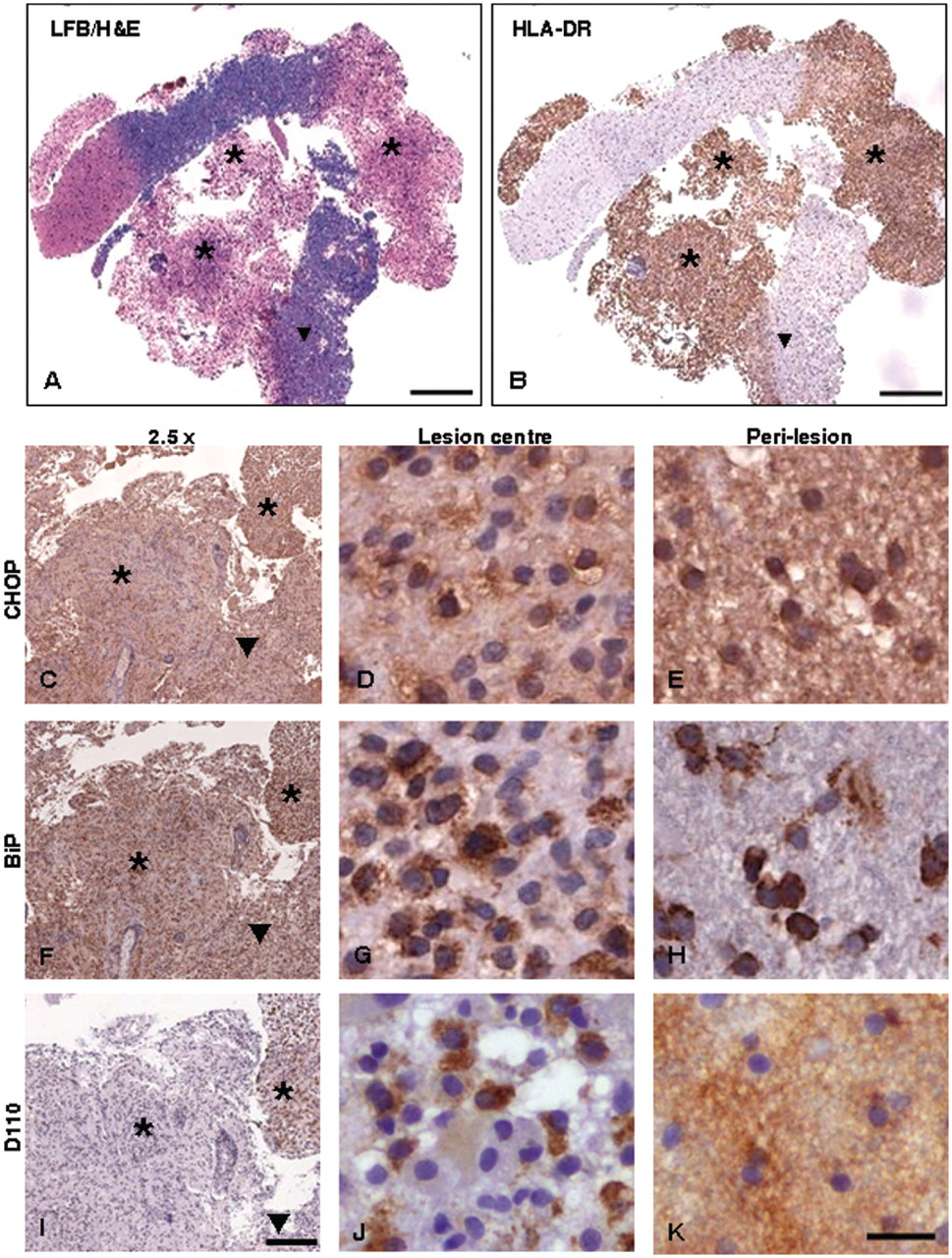
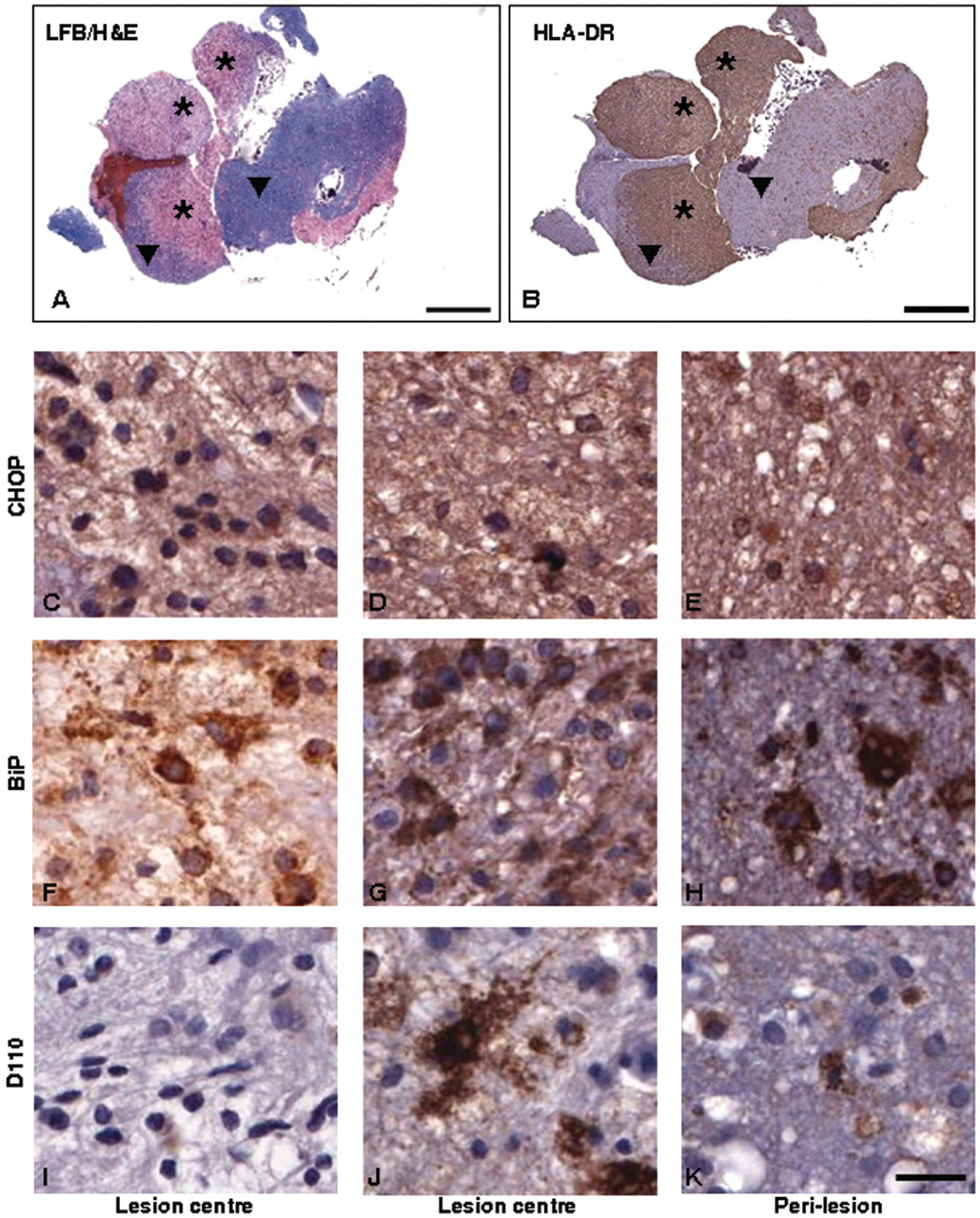
In all cases the H&E/LFB stain demarcated the actively demyelinating areas from the perilesional white matter. Sample images obtained from patients 1 and 7 are shown in part A of Figures 1 and 2. Intense and widespread expression of HLA-DR was also observed within all lesion centres (see panel B of Figures 1 and 2 for sample images).
Detection of ER stress and hypoxia markers in biopsied demyelinated lesions
Expression profile of the endoplasmic reticulum stress proteins CHOP and BiP and the hypoxia marker, D-110 in a series of demyelinating biopsies
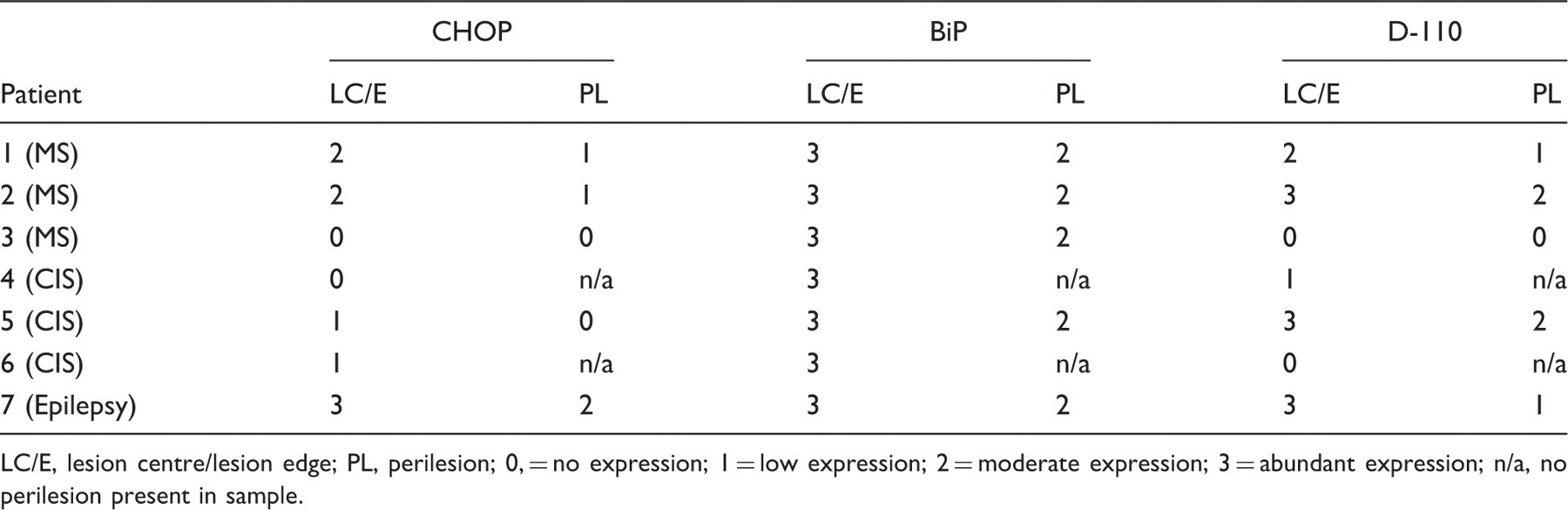
LC/E, lesion centre/lesion edge; PL, perilesion; 0, = no expression; 1 = low expression; 2 = moderate expression; 3 = abundant expression; n/a, no perilesion present in sample.
Samples obtained from patients with CIS generally contained lower levels of CHOP when compared with the epilepsy and MS patients, although BiP levels were abundant across all CIS lesions and in patient 6 were moderate in the PL (see Figures 2c and 2f for sample CHOP and BiP staining of the CIS patient 6). D-110 expression was essentially absent from the same sample, although occasional faint staining of cells was evident (Figure 2i). Abundant levels of CHOP expression were present at the lesion centre and edge of demyelinating zones derived from the epilepsy patient biopsy, with reduction to moderate levels in the surrounding perilesion (Figures 2d and 2e). In both lesion centre and perilesion, CHOP was found in the nucleus of a small population of cells. Expression of BiP (Figures 2g and 2h) and D-110 (Figures 2j and 2k) was also detected at abundant levels in the epilepsy lesion centre and edge, being reduced in the perilesion. D-110 expression was associated with cells morphologically identified as foamy macrophages and microglia (Figures 2i and 2j).
Differential expression of CHOP, ATF4 and BiP in laser-microdissected regions of post-mortem MS tissue samples
To preserve RNA integrity, frozen tissue sections were rapidly stained with H&E in order to enable targeted laser microdissection of desired section sub-regions (see Figure 3a). Real-time PCR analysis of resultant RNA preparations showed differences in the expression of BiP, CHOP and ATF4 genes within the various sub-regions isolated (Figure 3b). For all genes analysed, statistically significant increases in expression were observed in the NAWM compared with control white matter (BiP p < 0.01, CHOP p < 0.05 and ATF4 p < 0.05). In the case of CHOP, a significant increase in expression (p < 0.01) was additionally detected in the perilesion of active plaques compared with control white matter. Furthermore, a significant increase was also observed for CHOP in the perilesion of the chronic plaque compared with control white matter (p < 0.05). Minor increases in BiP expression in all sub-regions sampled compared with control white matter, with a significant increase detected in the perilesion of the chronic plaque samples, were also noted. Analysis of ATF4 expression also showed significant increases in expression in the perilesion of the active and chronic plaques (p < 0.05), with small increases in expression observed in the isolated plaques compared with control white matter. Differential expression of BiP, CHOP and ATF4 in laser-dissected multiple sclerosis tissue sub-regions. 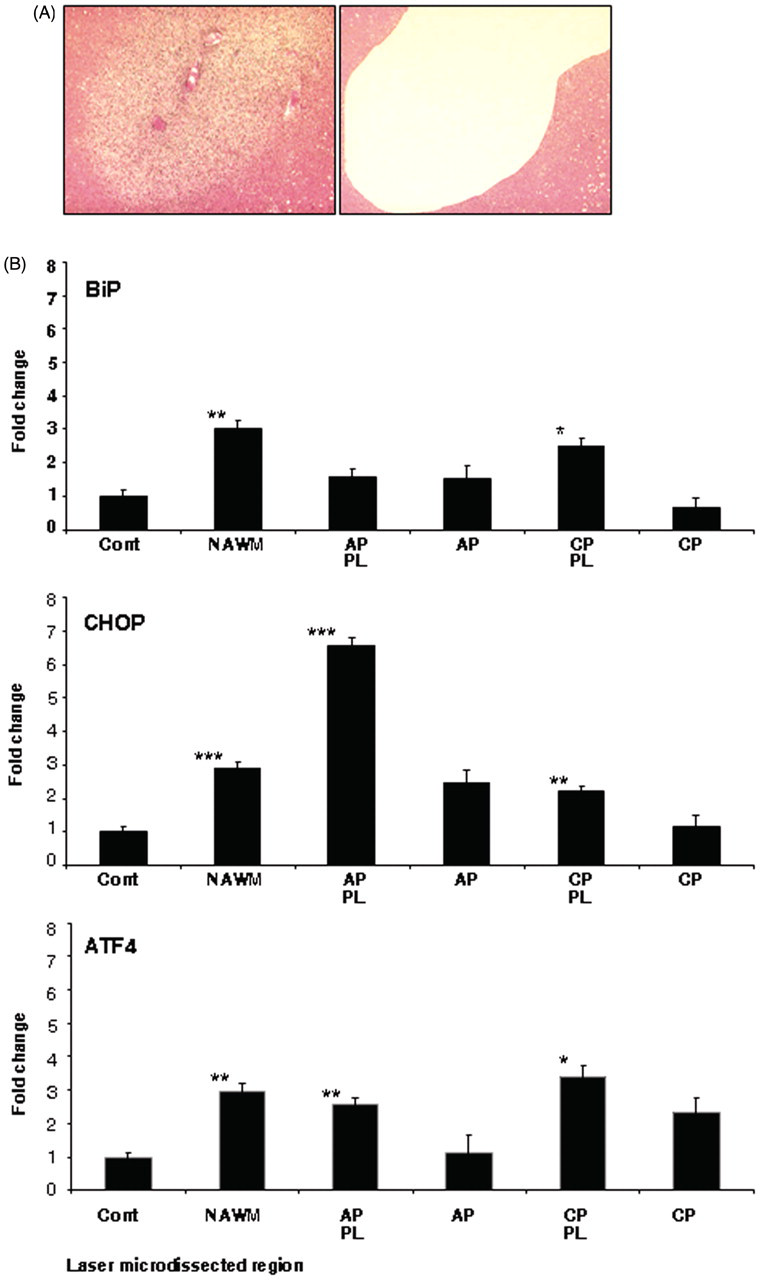
Discussion
The case for targeting of the ER stress pathway in the treatment of MS is gaining momentum. 16 – 18 In animal studies, Lin and colleagues provided the first evidence that the ER stress kinase PERK could be required to enhance oligodendrocyte survival in an inflammatory demyelinating environment. 19 – 21 In a subsequent detailed immunocytochemical analysis of post-mortem human MS tissue, increased levels of CHOP, BiP, and XBP1 were demonstrated, particularly at the edge of chronic active MS lesions. 7 The present study expands on our previous work, demonstrating increased expression of CHOP and BiP at both the centre and perilesional region in five out of seven actively demyelinating biopsied brain tissue samples. Detection of ER stress molecules in the early stages of demyelination calls for further investigations into the role of ER stress in MS pathogenesis. In addition, we detected significantly elevated levels of CHOP, BiP and ATF4 mRNA in NAWM laser-microdissected from post-mortem human MS brain, and significantly increased CHOP expression was also detected in the perilesion of active plaques (p < 0.01) compared with control white matter (Figure 3b).
The detection of raised levels of CHOP and BiP at the centre and edge of actively demyelinating lesions implicates the ER stress signalling pathway as a central mediator of early myelin destruction occurring in at least three kinds of clinically distinct demyelination. Here, we show that ER stress signalling is occurring at the earliest stages of demyelination within biopsy tissue obtained from patients with clinically definite MS, CIS and one unusual patient with demyelination of unknown aetiology. Detection of moderate levels of CHOP (cytoplasmic and/or nuclear) in a variety of cell types within demyelinating zones of two out of three MS biopsy samples (patients 1 and 2) strengthens our hypothesis that the ER stress signalling molecules are relevant to the earliest stages of demyelination. Strikingly, moderate or abundant levels of D-110 were also found at the lesion centre/edge of the same lesions, supporting the proposal 1,14 that for some MS patients, a response to hypoxic-like conditions may be activated at the earliest phase of demyelination. In a corollary to this, examination of the third MS biopsy sample (patient 3) suggests that ER stress or hypoxia may not be sufficient to explain all MS-related demyelinating triggers, as neither CHOP nor D110 were detectable at the lesion centre/edge, although moderate levels of D110 were detected in the perilesion.
ER stress activation in MS NAWM may represent ‘pre-lesional’ activity, suggested by abnormalities observed in the NAWM of MS tissue. Kutzelnigg and colleagues described diffuse white matter damage consisting of axonal abnormalities and ‘pale’ LFB staining, which correlated with cortical demyelination. 22 Allen et al. reported frequent microglial activation in the NAWM of 10 MS patient samples. 23 In addition, differential expression of ER stress genes in MS NAWM has been documented in a small number of studies. Graumann and colleagues detected elevated CHOP expression in NAWM from nine patients with MS when compared with non-neurological controls. 24 An additional study by Antony et al. also investigated the gene expression profile of a number of ER chaperone genes in MS white matter versus non-MS controls, showing increases in ER chaperones in MS tissue. 25 Our data, showing simultaneous increased expression of CHOP, BiP and ATF4 in NAWM compared with non-disease control tissue, together with previous data, strengthen the idea of some form of pre-lesional activity occurring in the NAWM. The previous studies, however, restricted their investigations to NAWM and did not consider gene expression changes in active or chronic active perilesions or lesions. Here, we demonstrate elevations in ER stress gene expression in active or chronic inactive lesions compared with non-MS controls. The fact that the greatest increase in CHOP expression was associated with the perilesional zones of active plaques suggests a possible role for CHOP in lesion development. The expression of pro-apoptotic CHOP is highly regulated during ER stress, therefore deciphering its precise role in lesion development is important.
The relationship between inflammation and ER stress is a complex one. While the pro-inflammatory molecules nitric oxide and tumour necrosis factor have been shown to induce the UPR in β-cells, macrophages, neurons and fibroblasts, 26 – 28 Zhang and Kaufman have recently summarized a variety of ways in which ER stress itself may act as the initiating trigger of an inflammatory response. 29 For example, excessive protein folding mediated by protein disulphide isomerase and ER oxidoreductase 1 within the ER generates reactive oxygen species (ROS) which can provoke inflammation. 30 PERK and IRE1 activation causing translation arrest or TRAF2 clustering, respectively, can interfere with NFκB inhibitory signals, thereby promoting a pro-inflammatory response. 31,32 Finally, two-way communication between the ER and mitochondria may be triggered by ER calcium depletion, leading to ROS generation and inflammation. 33 The idea that ROS are released following the accumulation of misfolded proteins within the ER of brain cells is intriguing, given the positive expression of the hypoxia marker D-110 at the centre of primary progressive MS lesions. It is possible that D-110 expression followed a hypoxic-type response induced after ROS release from CHOP-positive cells in lesion centres.
As inflammation has long been accepted as a trigger of demyelination, particularly within central nervous system white matter, deciphering the relationship between inflammation and ER stress within oligodendrocytes is essential. Lin and colleagues have made some progress in this area, demonstrating that the effect of ER stress on oligodendrocytes is context- and oligodendrocyte maturation-dependent. 19 – 21
To increase evidence that ER stress has a definitive role in MS lesion pathogenesis, and is therefore amenable to therapeutic intervention, the following questions still need to be addressed: (1) Should ER stress in pre-lesional areas be promoted or inhibited? (2) Can the ER stress response in individual cell types or in oligodendrocytes at different stages of development be manipulated through exogenous drug delivery? (3) In the context of MS, which ER stress molecules are the ‘master regulators’ and therefore potential therapeutic targets? (4) During demyelination, is inflammation triggering ER stress or is ER stress triggering inflammation? Answering these important questions requires much greater investigation into the mechanisms of ER stress in myelinating cells, to achieve a more complete understanding of the role of ER stress in the pathogenesis of MS.
Footnotes
Acknowledgements
Human biopsy samples were obtained from the files of the Neuropathology laboratory, Beaumont Hospital, Dublin, Ireland. Frozen tissue was supplied by the UK Multiple Sclerosis Tissue Bank, funded by the Multiple Sclerosis Society of Great Britain and Northern Ireland, registered charity 207495.
Funding
Conflict of interest
The authors declare that they have no conflicts of interest.