
Abstract
Background:
Alterations of synaptic transmission induced by inflammatory activity have been linked to the pathogenic mechanisms of multiple sclerosis (MS). Regulated upon activation, normal T-cell expressed, and secreted (RANTES) is a pro-inflammatory chemokine involved in MS pathophysiology, potentially able to regulate glutamate release and plasticity in MS brains, with relevant consequences on the clinical manifestations of the disease.
Objective:
To assess the role of RANTES in the regulation of cortical excitability.
Methods:
We explored the association of RANTES levels in the cerebrospinal fluid (CSF) of newly diagnosed MS patients with magnetic resonance imaging (MRI) and laboratory measures of inflammatory activity, as well its role in the control of cortical excitability and plasticity explored by means of transcranial magnetic stimulation (TMS), and in hippocampal mouse slices in vitro.
Results:
CSF levels of RANTES were remarkably high only in active MS patients and were correlated with the concentrations of interleukin-1β. RANTES levels were associated with TMS measures of cortical synaptic excitability, but not with long-term potentiation (LTP)-like plasticity. Similar findings were obtained in mouse hippocampal slices in vitro, where we observed that RANTES enhanced basal excitatory synaptic transmission with no effect on LTP.
Conclusion:
RANTES correlates with inflammation and synaptic excitability in MS brains.
Introduction
Multiple sclerosis (MS) is a neuroinflammatory and degenerative disease. It has been proposed that excitatory synaptic transmission of denervated neurons can be adaptively potentiated to minimize the neurological defect caused by a brain damage in MS through mechanisms of long-term potentiation (LTP).1,2 On the other hand, excitotoxic neuronal damage may represent a potential pathogenic mechanism contributing to MS severity. 3
During a relapse, MS patients show increased cortical excitability, 4 while expression of LTP-like plasticity is associated with recovery of the clinical defects related to a relapse 12 weeks after its onset. 2
A large amount of cytokines are released in the cerebrospinal fluid (CSF) and in the brain during neuroinflammation, predominantly by infiltrating T lymphocytes, and by activated astrocytes and microglia. A number of these cytokines influence neuronal excitability and synaptic plasticity, giving rise to a dense neuro-immune crosstalk. For instance, Interleukin 1β (IL-1β) was associated to increased cortical excitability 3 and LTP-like synaptic plasticity; 5 CSF levels of amyloid-β1–42 were associated to increased LTP and better cognitive performance during a relapse;6,7 IL-13 correlated with enhanced Gamma-AminoButyric Acid (GABA) mediated intracortical inhibition and higher axonal and neuronal integrity; 8 CSF levels of platelet derived growth factor (PDGF) were associated to greater LTP-like plasticity and recovery from the clinical deficits related to an acute relapse.1,9
Regulated upon activation, normal T-cell expressed, and secreted (RANTES), also named CCL5, is one of the principal chemotactic agents that induce recruitment of the immune cells to the CNS 10 via interaction with its corresponding receptor CCR5. CCR5 is also expressed in rat hippocampal neurons where its activation induces rapid changes in neuronal Ca2+ signaling 11 and glutamate release.12,13,14 Thus, along with its role in inflammation regulation, RANTES could in principle regulate also synaptic transmission 12 and brain plasticity.15,16
Here, we tested the hypothesis that RANTES, being regulated by inflammation17–20 and regulating neuronal activity,12,15,16 could play an important role in the neuro-immune crosstalk, which is crucial for the development of compensatory neuronal plasticity as well as for excitotoxic neuronal damage associated with acute inflammation. Indeed, our results show that the inflammatory process typical of MS is associated with an increase in the central levels of RANTES, which causes in turn significant changes of synaptic excitability of central neurons.
Patients and methods
The study was approved by the Institutional Review Board of the University Hospital Tor Vergata, Rome, Italy. All subjects gave their written informed consent to participate in the study.
CSF withdrawal
We collected CSF from 136 patients, admitted to the neurology clinic of the University Hospital Tor Vergata of Rome who underwent lumbar puncture and brain magnetic resonance imaging (MRI) for diagnostic purposes. Of these, 92 patients (age: 34.8 ± 0.9 years; sex: 62F, 30M) were later diagnosed as suffering from MS (80 relapsing-remitting MS (RRMS); 12 primary progressive MS (PPMS)). The diagnosis of MS was established by clinical laboratory and MRI parameters, and matched published criteria. 21 In all instances, patients underwent detection of oligoclonal banding in the CSF. In the remaining 44 patients (age: 44.6 ± 4.1 years; sex: 32F, 12M), the diagnosis was not confirmed and were thus considered as controls. They were individuals without inflammatory or degenerative diseases of the central or peripheral nervous system, who underwent lumbar puncture because of a clinical suspect of MS, acute peripheral neuropathy, meningitis, or subarachnoidal hemorrhage, which were not confirmed. In a subgroup of 17 patients who were diagnosed with RRMS and gave consent to transcranial magnetic stimulation (TMS), cortical excitability and plasticity were measured within 24 hours from CSF withdrawal. Corticosteroids or other MS-specific immunoactive therapies were initiated later when appropriate.
Determination of RANTES and IL-1β in the CSF
Immediately after withdrawal, the CSF was centrifuged and stored at −80°C until analyzed.
RANTES and IL-1β concentrations were analyzed using Bio-Plex Multiplex Cytokine Assay (Bio-Rad Laboratories, Hercules, CA, USA), according to manufacturer instructions. 5 Concentrations were calculated according to a standard curve generated for the specific target and expressed as pg/mL. When the concentrations were below the detection threshold, they were assumed to be 0 pg/mL.
Intracortical circuits in the motor cortex
The output of the primary motor cortex (M1) can be objectively measured in the form of a motor-evoked potential (MEP) from surface electromyographic (EMG) recording electrodes, positioned on the skin overlying the targeted muscles in response to a single supra-threshold TMS pulse delivered to M1.
The MEP amplitude elicited by stimulation of M1 can be modulated by a preceding conditioning pulse (CS). Depending on the intensity and inter-stimulus interval used a CS can produce either a facilitation or inhibition of the MEP elicited from the test stimulus (TS) over M1. To explore both excitatory and inhibitory synaptic transmissions, we tested through paired-pulse (pp) TMS, short-interval intracortical inhibition (SICI; mediated by GABA-A pathways), 22 intracortical facilitation (ICF; mediated by glutamatergic pathways),22–25 short-interval intracortical facilitation (SICF; mediated by excitatory cortical interneurons), 26 and long-interval intracortical inhibition (LICI; mediated by local GABA-B pathways) 27 of the left M1.
One figure-of-eight coil, with external diameter of 70 mm, was held tangentially to the scalp over the motor “hot spot” for the first dorsal interosseous muscle (FDI) of the dominant hand. Stimulation intensity for TS was adjusted in each experiment to evoke an MEP of about 1 mV peak-to-peak amplitude in the relaxed FDI.
All ppTMS experiments were conducted according to established protocols. SICI and ICF were tested using ppTMS with a sub-threshold CS preceding a supra-threshold TS. 22 As in previous works,3,28 CS stimulus was set at 80% active motor threshold (AMT). Three conditions were presented in a random order: control (TS given alone) and two pp conditions (TS preceded by CS) at one of two different ISI (2 and 10 ms). SICF was tested using a CS intensity of 90% resting motor threshold (RMT). 26 Six randomly intermixed conditions were presented in a random order: TS given alone and five conditions with the TS followed by the CS at one of five different ISIs (1.5, 2.1, 2.7, 3.7, and 4.3 ms). 26
For LICI, the intensity of the CS was set at 120% RMT. 27 Two conditions were presented in a random order: control (TS given alone) and one paired-pulse condition (TS preceded by CS) at 100 ms ISI. 27 For each experiment, 10 responses were collected for the TS alone and for the TS paired to a CS at each ISI. Changes in MEP amplitude at each ISI were expressed as the percentage of the mean unconditioned MEP amplitude.
EMG
EMG traces were recorded from the FDI of the dominant hand with surface cup electrodes. The active electrode was placed over the muscle belly and the reference electrode over the metacarpo-phalangeal joint of the index finger. Responses were amplified with a Digitimer D360 amplifier (Digitimer, Welwyn Garden City, Hertfordshire, UK) through filters set at 20 Hz and 2 kHz with a sampling rate of 5 kHz, and then recorded by a computer with SIGNAL software (Cambridge Electronic Design, Cambridge, UK). MEPs were evoked through a figure-of-eight coil with external loop diameter of 70 mm connected to a Magstim 2002 magnetic stimulator (Magstim Company, Whitland, UK).
The coil was held tangentially to the scalp surface with the handle pointing posteriorly and laterally at about 45° with respect to the mid-sagittal axis of the head, and positioned over the optimal scalp site to evoke motor responses in the contralateral FDI.
Cortical plasticity measured by means of TMS
Cortical plasticity was explored in patients by means of continuous theta burst stimulation (cTBS). TBS was delivered over the motor “hot spot” of the FDI of the dominant hand through a Magstim Rapid 2 stimulator. The AMT was defined as the minimum stimulation intensity required to evoke a liminal MEP from the FDI during voluntary contraction (about 200 µV in 50% of 10 trials).
Stimulation intensity was 80% of AMT. The TBS protocol consisted of 200 bursts repeated at a theta frequency of 5 Hz, each burst composed of three stimuli at 50 Hz, for a total of 600 stimuli (40 seconds).
The effect of cTBS on corticospinal excitability was quantified by measuring the amplitude of MEPs evoked in the dominant FDI by a constant intensity TMS pulse given over the contralateral motor cortex. Stimulation intensity was set to induce a stable MEP of approximately 1 mV peak-to-peak amplitude in the relaxed FDI of the dominant hand at baseline and remained unchanged until end of recordings. A total of 36 MEPs were collected before cTBS (baseline) and at two different time points (0 and 15 minutes) after the end of TBS. MEP’s amplitudes were then averaged at each time point and normalized to the mean baseline amplitude.
MRI acquisition and analysis
A 3-Tesla (3T) MRI scan consisted of dual-echo proton density, FLAIR, T2-weighted spin-echo images, and pre-contrast and post-contrast T1-weighted spin-echo images. All images were acquired in the axial orientation with 3-mm-thick contiguous slices. The presence of gadolinium-enhancing (active MS; 0.2 mL/kg e.v.) lesions was assessed by a neuroradiologist who was unaware of the patient’s clinical details. 6
Extracellular recordings in hippocampal CA1
Preparation of hippocampal slices was performed in accordance with the European Communities Council Directive (86/609/EEC). Vibratome-cut parasagittal slices (400 µm) were prepared, incubated for 1 hour, and then transferred to a recording chamber submerged in a continuously flowing artificial CSF (30°C, 2–3 mL/minute) gassed with 95% O2 and 5% CO2. The composition of the control solution was (in mM) as follows: 126 NaCl, 2.5 KCl, 1.2 MgCl2, 1.2 NaH2PO4, 2.4 CaCl2, 11 glucose, and 25 NaHCO3. Field excitatory post-synaptic potentials (fEPSPs) were evoked by stimulation of the Schaffer collateral/commissural pathway with a concentric bipolar stimulating electrode. For slices in which the presynaptic fiber volley was distinguishable, input–output relations were examined by plotting the initial slope of the fEPSP against the amplitude of the presynaptic fiber volley. LTP was induced by conventional TBS applied to the Schaffer collateral-CA1 synapses TBS (4 trains of 5 pulses at a frequency of 100 Hz, with an intertrain interval of 200 ms). Data were expressed as mean ± standard error (SE) and assessed for significance using the Student’s t test.
Data analysis
Kolmogorov–Smirnov test was used to test for normality distribution of data. Between-group differences were evaluated using the Student’s t test or Mann–Whitney test as appropriate. The effect of each TMS protocol was evaluated using repeated measures analysis of variance (ANOVA). The relationship between RANTES and TMS protocols was evaluated calculating Spearman’s correlation coefficients. Data, expressed as mean ± standard deviation (SD), were considered significant for p < 0.05.
Results
RANTES in the CSF of MS patients and of controls
Mann–Whitney test revealed that RANTES expression in the CSF was higher in MS patients (mean ± SD = 825.94 ± 412.67) compared to controls (mean ± SD = 70.39 ± 39.51; p < 0.001), according to previous reports showing that RANTES increases during neuroinflammatory insults.17–20
In all, 57 patients (54 RRMS and 3 PPMS) showed active inflammatory brain lesions measured using gadolinium (Gd)-enhanced brain MRI or clinical signs indicative of an acute relapse (active MS). The remaining 35 patients did not show any neuroradiological or clinical signs/symptoms indicative of ongoing brain inflammation (non-active MS). Mean RANTES CSF levels were impressively higher in active than in non-active MS patients and in control subjects (p < 0.01; Figure 1(a)), consistently with the conclusion that disease activity is associated with altered RANTES metabolism in MS brains. Accordingly, in MS patients, we also found a correlation between RANTES CSF levels and those of the classical pro-inflammatory cytokine IL-1β (rs = 0.35, p < 0.001; Figure 1(b)). Conversely, RANTES did not differ comparing RRMS to PPMS patients (p = 0.22).
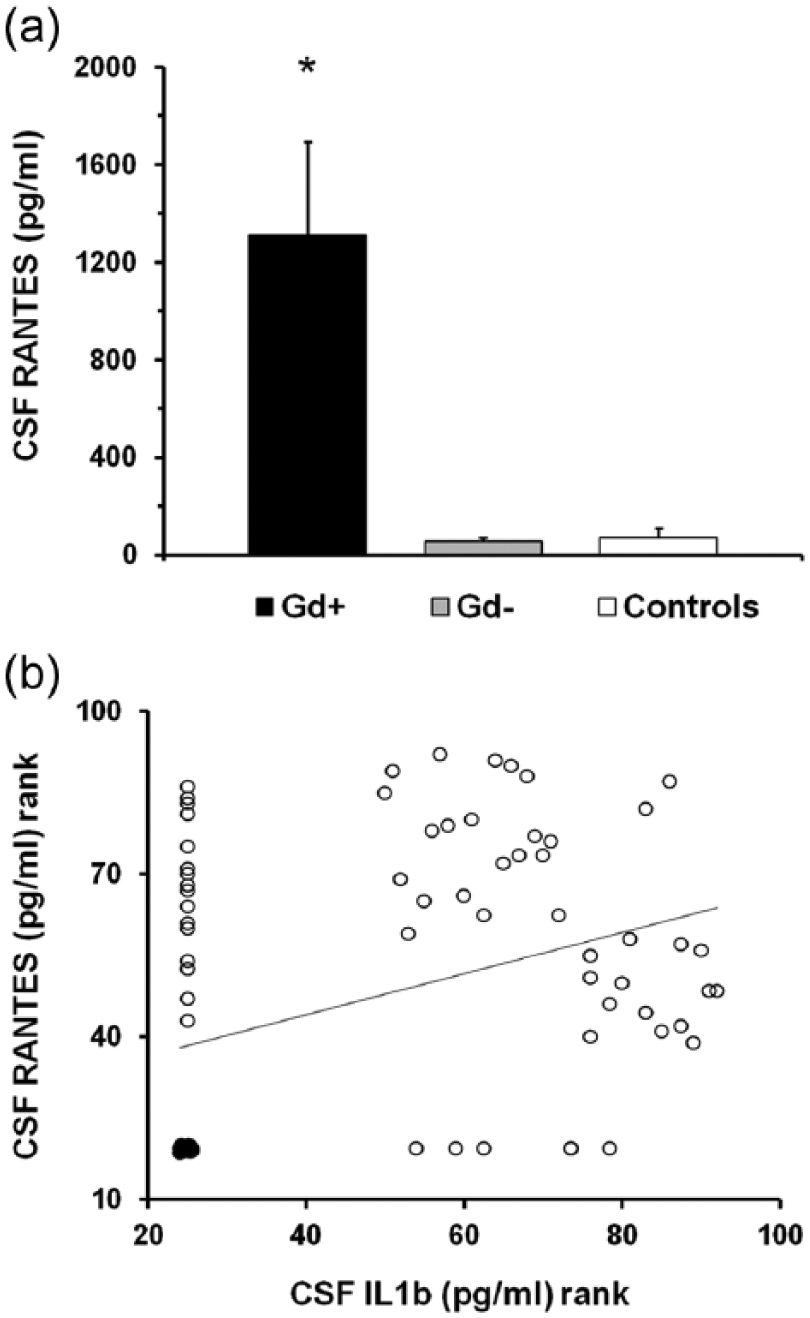
RANTES levels in the CSF of MS patients are associated with disease activity. (a) CSF RANTES levels are strongly increased in MS patients showing contrast enhancing brain lesions or clinical signs indicative of an acute relapse (active MS) compared to patients without MRI and clinical signs of disease activity (non-active MS) and controls. (b) CSF RANTES levels correlate with CSF levels of IL-1β (rs = 0.35, p < 0.001), a pro-inflammatory cytokine involved in MS pathogenesis. Ranked values are displayed in (b).
RANTES in the regulation of neuronal excitability in MS patients
Exploring intracortical motor circuits through ppTMS showed a significant effect of ISI for SICI (ISI = 2 ms, F = 5.46, p < 0.05), ICF (ISI = 10 ms, F = 3.54, p < 0.05), SICF (ISI = 1.5 ms, F = 2.86, p < 0.05; ISI = 2.7 ms, F = 3.08, p < 0.05; ISI = 4.5 ms, F = 2.76, p < 0.05), and LICI (ISI = 100 ms, F = 374.85, p < 0.001; Figure 2(a)).
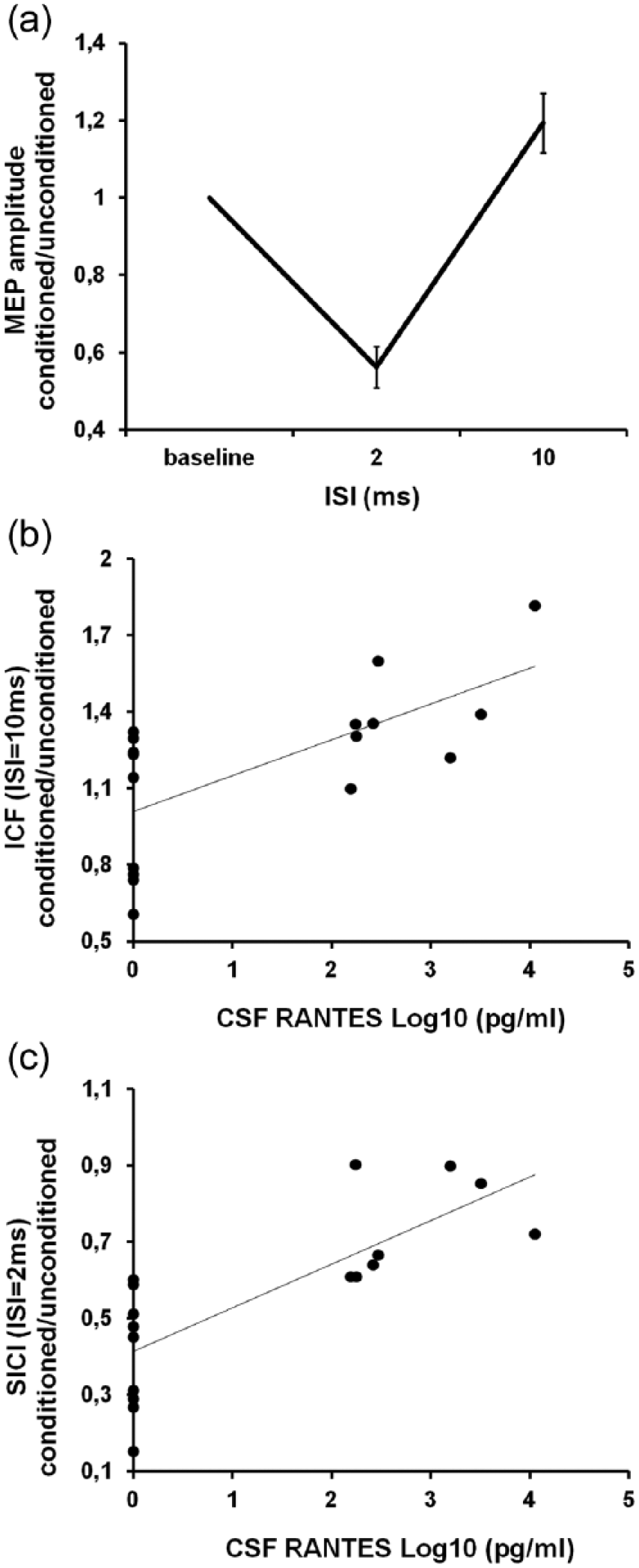
Intracortical facilitation and short-interval intracortical inhibition correlate with the levels of RANTES in the CSF of RRMS patients. (a) During paired-pulse TMS, the cortical motor output is inhibited at 2 ms ISI (short-interval intracortical inhibition (SICI); p < 0.05), while it is facilitated at an ISI of 10 ms (intracortical facilitation (ICF); p < 0.05). Both (b) ICF (rs = 0.55, p < 0.01) and (c) SICI (rs = 0.81, p < 0.001) correlate with RANTES CSF levels in our patients. To better represent the relationship between RANTES and ppTMS parameters, RANTES pg/mL concentration has been transformed into its base-10 logarithm.
Correlation analysis showed that higher levels of RANTES in the CSF of MS patients were associated with increased ICF (rs = 0.55, p < 0.01; Figure 2(b)) and decreased SICI (rs = 0.81, p < 0.001; Figure 2(c)), suggesting that RANTES increases synaptic excitability in MS patients. In contrast, CSF levels of RANTES did not correlate with SICF (rs = 0.30, p = 0.30 for ISI = 1.5 ms; rs = 0.27, p = 0.27 for ISI = 2.7 ms; rs = 0.33, p = 0.41 for ISI = 4.5 ms) nor LICI (rs = −0.06, p = 0.41).
RANTES in the control of TBS-induced LTP-like plasticity in MS
Repeated measures ANOVA showed a significant effect of time (F = 5.09, p = 0.02). MEP amplitude significantly increased 15 minutes after TBS (mean ± SE post/pre MEP amplitude = 1.64 ± 0.19; F = 10.84, p < 0.05; Figure 3(a)). The association between RANTES and TBS after-effects was explored through Spearman’s correlation analysis and showed no significant correlations between MEP amplitude changes 0 and 15′ after TBS and RANTES CSF levels of MS patients (Figure 3(b)).
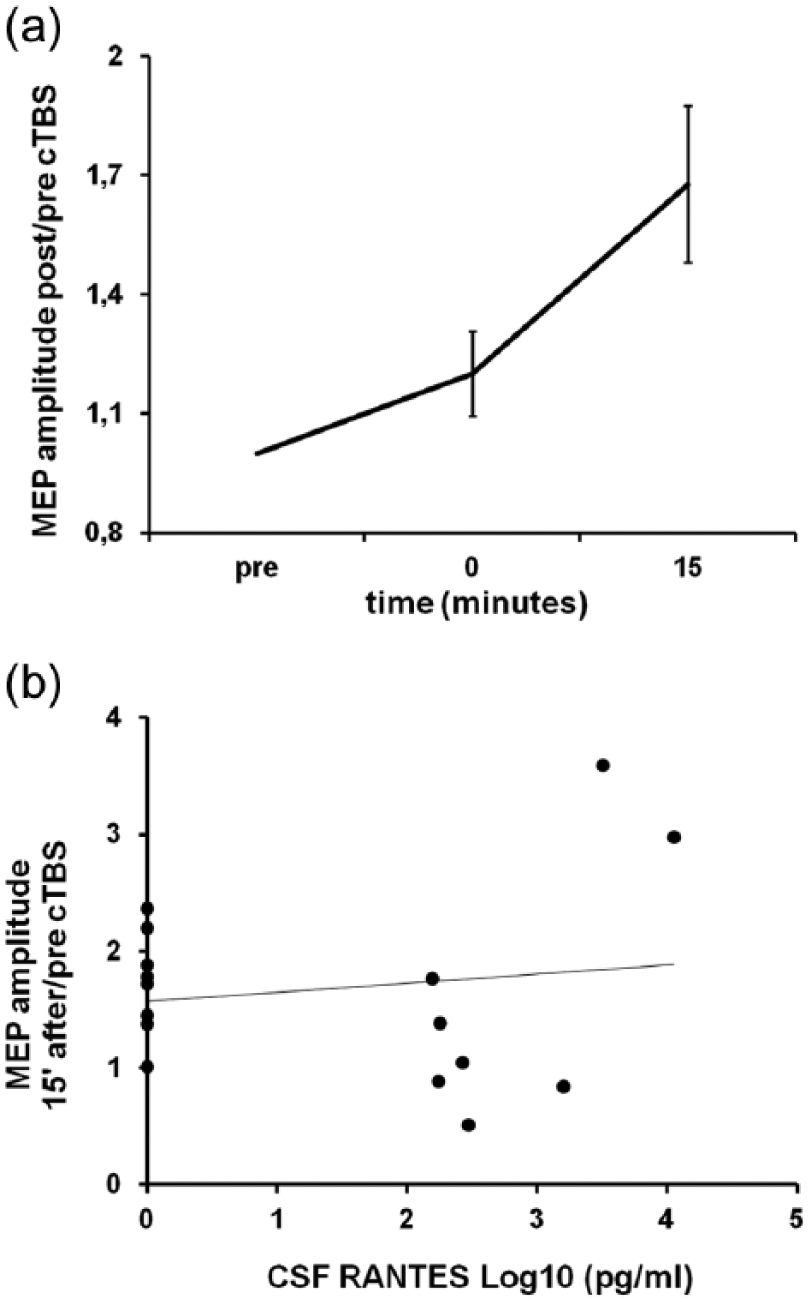
cTBS-induced LTP-like effect does not correlate with the levels of RANTES in the CSF of RRMS patients. (a) cTBS induces a long-lasting facilitation of the motor cortical output (LTP-like effect; p < 0.05). (b) No correlation between RANTES CSF levels and TBS-induced LTP was found in our patients. To better represent the relationship between RANTES and ppTMS parameters, RANTES pg/mL concentration has been transformed into its base-10 logarithm.
RANTES in the control of synaptic transmission and LTP in brain slices
The TMS data are compatible with the idea that RANTES modulates synaptic transmission by enhancing glutamate-mediated excitability but not LTP. Thus, we next sought to test the effect of RANTES on synaptic transmission and LTP in mouse hippocampal slices. A scatter plot relating the initial slope of the fEPSP to the size of the presynaptic fiber volley showed that slices incubated with RANTES (30 nM for 1 hour) displayed enhanced input–output relationship at CA1 hippocampal synapses (p < 0.05; Figure 4(a)). Next, to determine whether synaptic plasticity is modulated by RANTES, we studied hippocampal LTP induced by TBS. CA1-LTP was always comparable to vehicle-treated slices following pre-incubation with RANTES. Indeed, the magnitude of potentiation measured between 50 and 60 minutes after TBS was similar in RANTES-treated (150% ± 8%, n = 6) compared with control slices (156% ± 11%, n = 6, p > 0.05; Figure 4(b)).
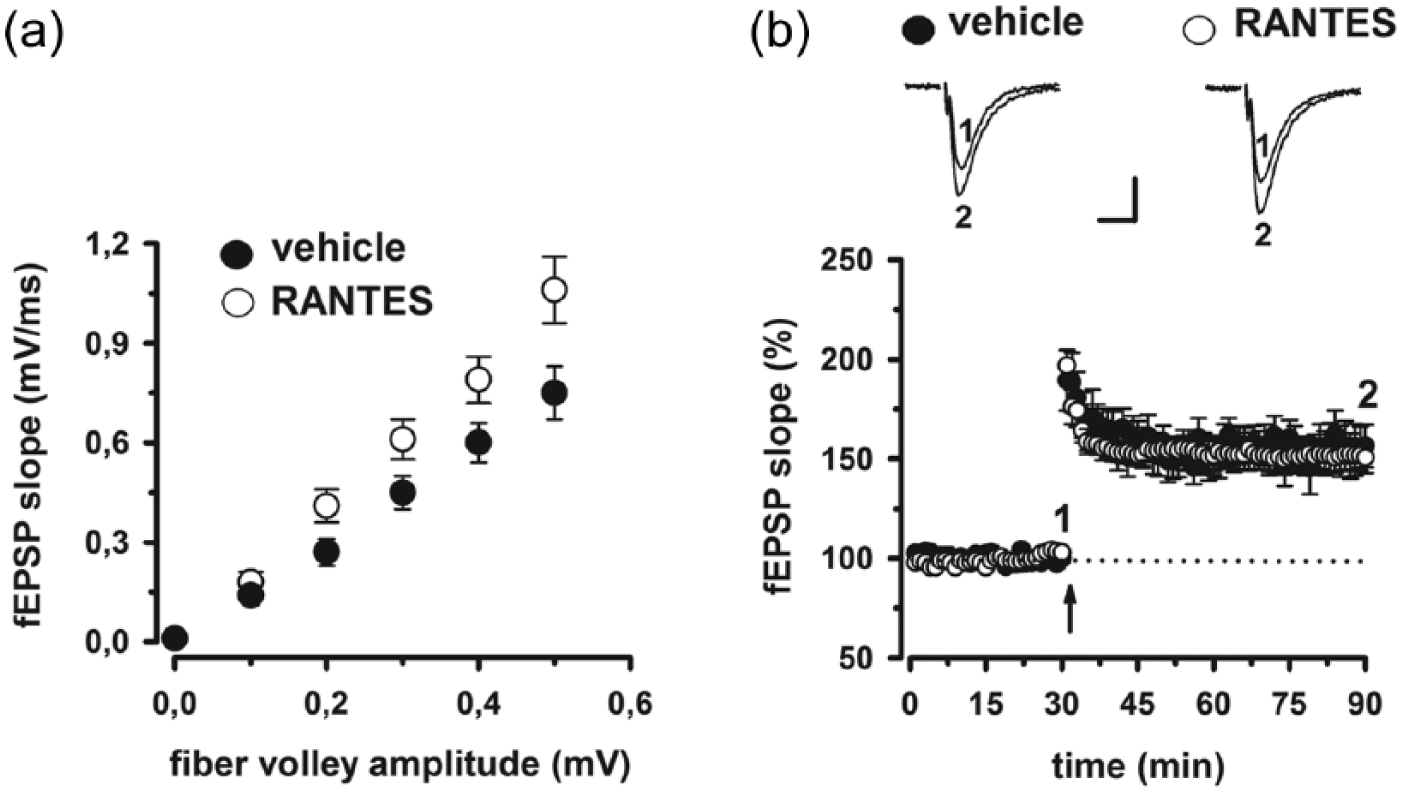
Effect of RANTES on hippocampal synaptic function. (a) Input–output relationship is shown as plots of the fEPSP slopes against the corresponding presynaptic fiber volley amplitudes (p < 0.05 at 0.3, 0.4, and 0.5 mV). Each treatment group is the average ± SE of at least six recordings performed on separate slices. Significant differences between treatment groups were detected. (b) Time plot of fEPSP slope responses showing that pre-incubation with RANTES has no effect on the magnitude of CA1-LTP compared to vehicle-treated slices. Representative fEPSP recordings from time points 1 and 2 are shown for each condition. Calibration bars: 0.5 mV, 10 ms.
Discussion
RANTES increases during MS disease activity.17–20 Here, we report that higher levels of RANTES in the CSF of MS subjects were associated with increased motor cortex excitability as assessed by ppTMS protocols at both 2 and 10 ms ISIs. At ISI of 10 ms and 80% AMT intensity, a CS stimulus usually induces excitatory effects on motor cortex excitability, which are believed to be mediated by glutamate receptors. 24 On the contrary, a CS at 2 ms and 80% AMT usually induces inhibitory effects on motor cortical excitability which are thought to be mediated by GABA-A receptor synaptic inhibition, as suggested by pharmacological studies. 29 However, at this ISI, SICI does not result from a pure inhibition, and enhancement of glutamatergic transmission reduces the degree of synaptic inhibition recorded at this ISI, as observed in our study. 28
Accordingly, drugs that reduce facilitation, such as glutamate receptor antagonists, lead to an apparent increase in SICI.23,24
Our in vitro study provided direct evidence that RANTES increases synaptic excitability by enhancing glutamatergic transmission. Glutamatergic nerve endings in the human neocortex seem to possess all three RANTES receptor subtypes, CCR1, CCR3, and CCR511, and previous in vitro studies showed that RANTES may have a dual role on glutamatergic transmission. It can potentiate the basal release of glutamate and can also inhibit its depolarization-evoked release. 12 RANTES inhibits glutamate release from cultured rat hippocampal neurons, 11 from human and mouse cortical nerve ending terminals, 12 and from mouse spinal cord terminals. 13 However, during experimental autoimmune encephalomyelitis (EAE), the rodent model of MS, which is associated to an early reduction in the KCl-evoked glutamate release in the cortex, the CCL5-mediated inhibition of glutamate exocytosis observed in control mice turns to facilitation. 13
Thus, we hypothesize that RANTES may enhance glutamate-mediated synaptic transmission in the cortex of MS patients. In support of this hypothesis, we found that application of RANTES in mouse hippocampal slices was able to increase basal synaptic transmission. We also explored whether CNS RANTES expression was associated with expression of synaptic plasticity effects in MS patients. However, no correlation emerged between RANTES CSF levels and TBS-induced synaptic plasticity and, consistently, no effect on TBS-induced LTP was observed after bath application of RANTES in hippocampal slices.
As in previous reports, continuous TBS in our patients induced an LTP-like effect. Indeed, even though this TBS protocol is mainly reported to induce a long-term depression (LTD)-like effect in the motor cortex of healthy subjects, it also showed the ability to induce LTP sometimes, both in healthy subjects 30 and in MS patients.1,5 In particular, an LTP-like response to the continuous TBS protocol was observed in RRMS patients but not in healthy controls, nor in patients affected by the progressive form of MS. 1
RANTES has been proposed to be neuroprotective. 31 Adding RANTES to the culture medium slowed the rate of apoptotic death of hippocampal neurons after removal of their glial cell feeder layer. 11 RANTES, indeed, can induce the expression of genes involved in neuronal survival, neurite outgrowth, and synaptogenesis. 16 However, genetic polymorphisms regulating the expression of RANTES or of its receptor, the CCR5, have been associated with MS progression and severity, suggesting that RANTES expression is associated with increased disease severity. 32 Genetic polymorphisms resulting in lower expression of RANTES (i.e. the single nucleotide A to G substitution in position 403 in RANTES gene promoter) or of its receptor the CCR5 (A to G substitution in position 303 of the CCR5 promoter sequence, 32 base pair deletion (X32) in the coding region of CCR5 gene) were associated with lower indices of MS severity. 32 The X32 mutation has also been associated to the risk of developing MS by a number of studies, with controversial results (for a review, see Ghorban et al. 33 ).
Our study reports for the first time an association between the expression of RANTES in the CNS and cortical excitability in MS patients, providing further support to the idea that RANTES modulates glutamate transmission in this disorder. Accordingly, here, we observed that application of this chemokine in hippocampal slices enhanced input–output curves for basal excitatory transmission, which also reflects glutamate release, with no effect on synaptic plasticity. These reports enrich the scenario of the neuron-glia interaction in MS and are in agreement with the general idea that chemokines, acting on their specific receptors, constitute a major communication system in the CNS, in concert with neurotransmitters and neuropeptides. 34
Footnotes
Conflict of interest
Dr Diego Centonze acted as an Advisory Board member of Merck Serono, Teva, Bayer Schering, Biogen Idec, Novartis, Almirall, and GW Pharmaceuticals and received funding for traveling and honoraria for speaking or consultation fees from Merck Serono, Teva, Novartis, Bayer Schering, Sanofi-Aventis, Biogen Idec, and Almirall. He is the principal investigator in clinical trials for Novartis, Merck Serono, Teva, Bayer Schering, Sanofi-Aventis, Biogen Idec, and Roche. The other authors declare that they have no competing interests.
Funding
The present investigation was funded by a grant from the Fondazione Italiana Sclerosi Multipla (Progetto Speciale FISM, 2012/S/2) to DC.