
Abstract
Background:
CD59, a broadly expressed glycosylphosphatidylinositol-anchored protein, is the principal cell inhibitor of complement membrane attack on cells. In the demyelinating disorders, multiple sclerosis (MS) and neuromyelitis optica spectrum disorder (NMOSD), elevated complement protein levels, including soluble CD59 (sCD59), were reported in cerebrospinal fluid (CSF).
Objectives:
We compared sCD59 levels in CSF and matched plasma in controls and patients with MS, NMOSD and clinically isolated syndrome (CIS) and investigated the source of CSF sCD59 and whether it was microparticle associated.
Methods:
sCD59 was quantified using enzyme-linked immunosorbent assay (ELISA; Hycult; HK374-02). Patient and control CSF was subjected to western blotting to characterise anti-CD59-reactive materials. CD59 was localised by immunostaining and in situ hybridisation.
Results:
CSF sCD59 levels were double those in plasma (CSF, 30.2 ng/mL; plasma, 16.3 ng/mL). Plasma but not CSF sCD59 levels differentiated MS from NMOSD, MS from CIS and NMOSD/CIS from controls. Elimination of microparticles confirmed that CSF sCD59 was not membrane anchored.
Conclusion:
CSF levels of sCD59 are not a biomarker of demyelinating diseases. High levels of sCD59 in CSF relative to plasma suggest an intrathecal source; CD59 expression in brain parenchyma was low, but expression was strong on choroid plexus (CP) epithelium, immediately adjacent the CSF, suggesting that this is the likely source.
Keywords
Introduction
The complement system is a central component of innate immunity responsible for recognition and killing of bacteria by lysis and promoting phagocytosis through opsonisation. The main source of most complement proteins is the liver; however, other cells, including fibroblasts, epithelial and endothelial cells, glia and neurons, also produce complement proteins. 1 Activation of complement via the classical, lectin or alternative pathway leads to the formation of C3 convertase complexes that cleave C3 to C3a and C3b. C3b is the nidus for the C5 convertase that cleaves C5 to C5a and C5b, the latter initiating formation of the membrane attack complex (MAC) responsible for osmotic cell lysis. CD59, an ~18- to 20-kDa glycosylphosphatidylinositol (GPI)-anchored protein, regulates formation of MAC by preventing recruitment of C9. 2 Little is known about expression of CD59 in the brain; in one study, neurones were reported to lack CD59, and expression of CD59 on glia was increased in inflamed brain. 3 The blood–brain barrier (BBB) restricts entry of molecules into the central nervous system (CNS), providing a natural defence against circulating toxic or infectious agents. 4 The choroid plexus (CP) plays a crucial role in the production of cerebrospinal fluid (CSF) and may be a site of immune activation; for example, in a mouse model of human systemic lupus erythematosus (SLE), dense deposits of immunoglobulin G (IgG) and C3 were found in the CP. 5 More recently, the role of CP in cellular immunology became more evident with the demonstration that CP was responsible for controlling and development of immune responses in the CNS. 6 Recent immunohistochemical (IHC) data on human CP showed deposition of complement proteins: C1q, C3d and C9 in stromal concretions and IgG in epithelial cells, suggesting significant involvement of CP in clearance of complement-tagged immune complexes. 7 Multiple sclerosis (MS), neuromyelitis optica spectrum disorder (NMOSD) and clinically isolated syndrome (CIS) are autoimmune demyelinating brain disorders in which dysregulation of complement has been noted in CSF, plasma and brain.8–15 It was recently reported that soluble CD59 (sCD59) is increased in CSF in MS and NMOSD. 16 This finding prompted us to further investigate levels of sCD59 in CSF and plasma from MS, NMOSD and CIS cases and healthy controls with the aim of gaining insight into underlying pathology. sCD59 was measured using a commercial enzyme-linked immunosorbent assay (ELISA), and the nature of sCD59 in CSF, microparticle associated or free, was explored. To discover the source of sCD59 in CSF, we analysed brain CD59 expression by IHC.
Materials and methods
All chemicals, except where otherwise stated, were obtained from either Fisher Scientific UK (Loughborough, Leicestershire, UK) or Sigma–Aldrich (Gillingham, Dorset, UK) and were of analytical grade.
Subjects
CSF and plasma samples
CSF and plasma samples from 50 MS patients, 11 NMOSD patients, 17 CIS patients and 34 controls and matched ethylenediaminetetraacetic acid (EDTA) plasma samples from 41 MS patients, 10 NMOSD patients, 14 CIS patients and 26 controls were obtained from the Welsh Neuroscience Research Tissue Bank, Institute of Neurosciences and Mental Health, Cardiff University. Informed consent was obtained from all patients, and ethical approval was gained from South East Wales Ethics Committee (ref. no. 05/WSE03/111). Diagnosis for MS cases was made according to established criteria; 17 all MS cases had clinically definite disease and had exhibited a relapsing or progressive disease course for which comprehensive follow-up clinical data were available. In addition, all MS cases had undergone diagnostic and follow-up magnetic resonance imaging (MRI) as part of routine clinical care. All patients with NMOSD were reviewed, and diagnosis confirmed according to established criteria 18 by a neurologist with experience in the diagnosis and management of neuroinflammatory disorders including NMOSD. All NMOSD cases had undergone diagnostic and follow-up MRI as part of routine clinical care. AQP4-Ab and MOG-Ab status was assessed at the Clinical Neuroimmunology Service, John Radcliffe Hospital Trust, Oxford, United Kingdom.19,20 Demographics and clinical data are summarised in Table 1.
Demographics and clinical data of patients included in the study. Control cohort comprised patients with Benign (Idiopathic) Intracranial Hypertension (BIH).
MS: multiple sclerosis; NMOSD: neuromyelitis optica spectrum disorder; CIS: clinically isolated syndrome; SD: standard deviation; CSF: cerebrospinal fluid; EDSS: Expanded Disability Status Scale; PPMS: primary progressive multiple sclerosis; RRMS: relapsing–remitting multiple sclerosis; SPMS: secondary progressive multiple sclerosis; N/A: not applicable; MP: methylprednisolone; MM: mycophenolate mofetil; Az: azathioprine; Rx: Rituximab; ON: optic neuritis; ATM: acute transverse myelitis.
Autopsy tissue
MS (n = 5 cases) and non-neurological control (n = 7 cases) post-mortem tissue was provided by the UK Multiple Sclerosis Society Tissue Bank at Imperial College and the Oxford Brain Bank at Oxford University, with research ethical approval (South West Wales REC study 12/WA/0292). Sections of 6 µm thick were prepared from a single region-matched paraffin-embedded block per case of the anterior hippocampus at the level of the lateral geniculate nucleus. All sections assessed contained intact meninges and CP (of the inferior horn of the lateral ventricle) and were characterised in terms of white matter and grey matter demyelination, tissue and parenchymal microglial/ macrophage infiltration as described previously. 21 Case details are provided in Table 2.
Details of the cases used for IHC in this study. All cases were pathologically confirmed cases of progressive MS; controls were non-neurological disease (Ctrl). One tissue block, containing the anterior hippocampus, was sampled in each case. MS tissue blocks contained periventricular white matter lesions (p’vent WML), which were characterised as chronic active in terms of the extent of microglia/ macrophage infiltration, cortical grey matter lesions (GML – either affecting the subpia or crossing the leucocortical boundary) or normal tissue.
MS: multiple sclerosis; PMD: post-mortem delay; PPMS: primary progressive multiple sclerosis; SPMS: secondary progressive multiple sclerosis.
CD59 ELISA
The concentration of sCD59 in CSF and plasma samples was measured using a commercial ELISA (HK374-02; Hycult Biotech, Uden, The Netherlands). Levels of sCD59 in CSF, unfractionated and following high-speed centrifugation (21200 rcf for 5 minutes at room temperature (RT)) to remove microparticles, were compared in a subset of 23 samples.
Sodium dodecyl sulphate polyacrylamide gel electrophoresis and western blotting
Patient and control CSF was subjected to sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting to further characterise anti-CD59-reactive materials in CSF. Prior to analysis, the patient CSF samples (MS and NMOSD) were 10-fold concentrated using Vivaspin sample concentrators (cut-off size: 10 kDa; GE Healthcare, Amersham Place, Little Chalfont, UK). As a positive control, human erythrocyte membranes, prepared by lysing erythrocytes in deionised water, centrifuging and washing the membrane pellet in phosphate buffer saline (PBS), were solubilised in non-reducing sample buffer (0.1 M Tris, 10% w/v glycerol, 2% w/v SDS, bromophenol blue, pH 6.8 in deionised water). Undiluted plasma or CSF samples concentrated as above were mixed 1:5 with non-reducing sample buffer. All samples were resolved on 4%–20% gradient SDS-PAGE gels (456–1094; Bio-Rad, Watford, Hertfordshire, UK) and then electrophoretically transferred onto 0.45-µm nitrocellulose membrane (10600002; GE Healthcare, Amersham, UK) at a constant voltage of 90 V in 25 mM Tris, 19.2 mM Glycine buffer with 20% methanol. After transfer, non-specific sites on the membrane were blocked with 3% bovine serum albumin (BSA) in PBS for 1 hour at RT. All subsequent incubations were carried out for 1 hour at RT and followed by three washes with PBS containing 0.1% Tween-20 (PBS-T). Post-blocking, the membrane was incubated with the primary antibody, monoclonal anti-human CD59 (MEM43 at 1 µg/mL in PBS-T). After washing, the bound antibodies were revealed by incubation with donkey anti-mouse IgG-horseradish peroxidase (HRP) conjugate (715-035-151; Jackson ImmunoResearch, West Grove, PA, USA) at an optimised dilution of 1:2000 in PBS-T. The blot was developed with ECL (RPN 2106; GE Healthcare) and visualised by autoradiography.
Immunohistochemistry and analysis
Sections were de-waxed, rehydrated and subjected to heat-induced epitope retrieval in citraconic anhydride (0.05%) and then quenched in H2O2 and blocked with normal serum before overnight incubation with monoclonal (MEM43) or rabbit polyclonal anti-CD59 antibody. Bound primary antibody was detected using the appropriate biotinylated goat secondary antibody (Vector Labs, Peterborough, Cambridgeshire, UK) prior to incubating with the Avidin-biotin HRP complex (ABC Elite; Vector Labs). Staining was visualised using diaminobenzidine (ImmPACT DAB; Vector Labs), and all sections were counterstained with haematoxylin before dehydrating, clearing and mounting in DePex (Fisher Scientific). No-primary-antibody and irrelevant mouse or rabbit antibodies were included as staining controls, and all sections were immunostained in the same experimental run. Sections were viewed, and images were captured using a Zeiss Axio Imager II fitted with an AxioCam HR camera (Carl Zeiss Ltd, Cambridge, Cambridgeshire, UK). The relative intensity of CD59 immunoreactivity (mouse monoclonal antibody MEM43) in the CP, ependyma, meninges, vasculature and brain parenchyma was evaluated as follows: 0, absent; +, mild (very few stained cells or weakly stained structures); ++, moderate (30%–70% of cells immunopositive or structures showing intermediate immunopositivity); and +++, substantial (majority of cells or tissues are intensely immunopositive).
In situ hybridisation
In situ hybridisation (ISH) was performed using a 5′ fluorescein (FAM)-labelled 22mer antisense oligonucleotide containing locked nucleic acid (LNA) and 2′-O-methyl (2′-O-Me)-RNA moieties in a 1:2 ratio. The probe was provided by Eurogentec (Southampton, UK). Probes were as follows: antisense CD59 (NM_203329) FAM – GguAauGagAcaCgcAucAaaA and sense FAM – TuuTgaTgcGugTcuCauTacC (capitals indicate LNA and lowercase 2′-O-Me-RNA), designed following previously described protocols.21,22 Specifically bound probe was visualised with a peroxidase-conjugated goat anti-FAM (Vector Labs) and DAB. Sections were counterstained with haematoxylin before dehydrating, clearing and mounting.
Results
Levels of sCD59 in CSF and plasma
sCD59 was detected in all CSF and plasma samples tested; levels in CSF were approximately double those in matched plasma (Figure 1(a); p < 0.0001). There was no significant difference in CSF sCD59 levels between the various disease groups (Figure 1(b)); however, measurement of sCD59 in plasma differentiated MS from NMOSD (p < 0.005), MS from CIS (p < 0.003) and NMO/CIS from controls (p < 0.003; Figure 1(c)). Plasma sCD59 levels were, on average, around half that in matched CSF samples (mean sCD59 in CSF = 30.2 ng/mL, plasma = 16.3 ng/mL; n = 91). The observed ratios between CSF and plasma sCD59 levels (~2:1) were consistent across the groups (Figure 1(d)).
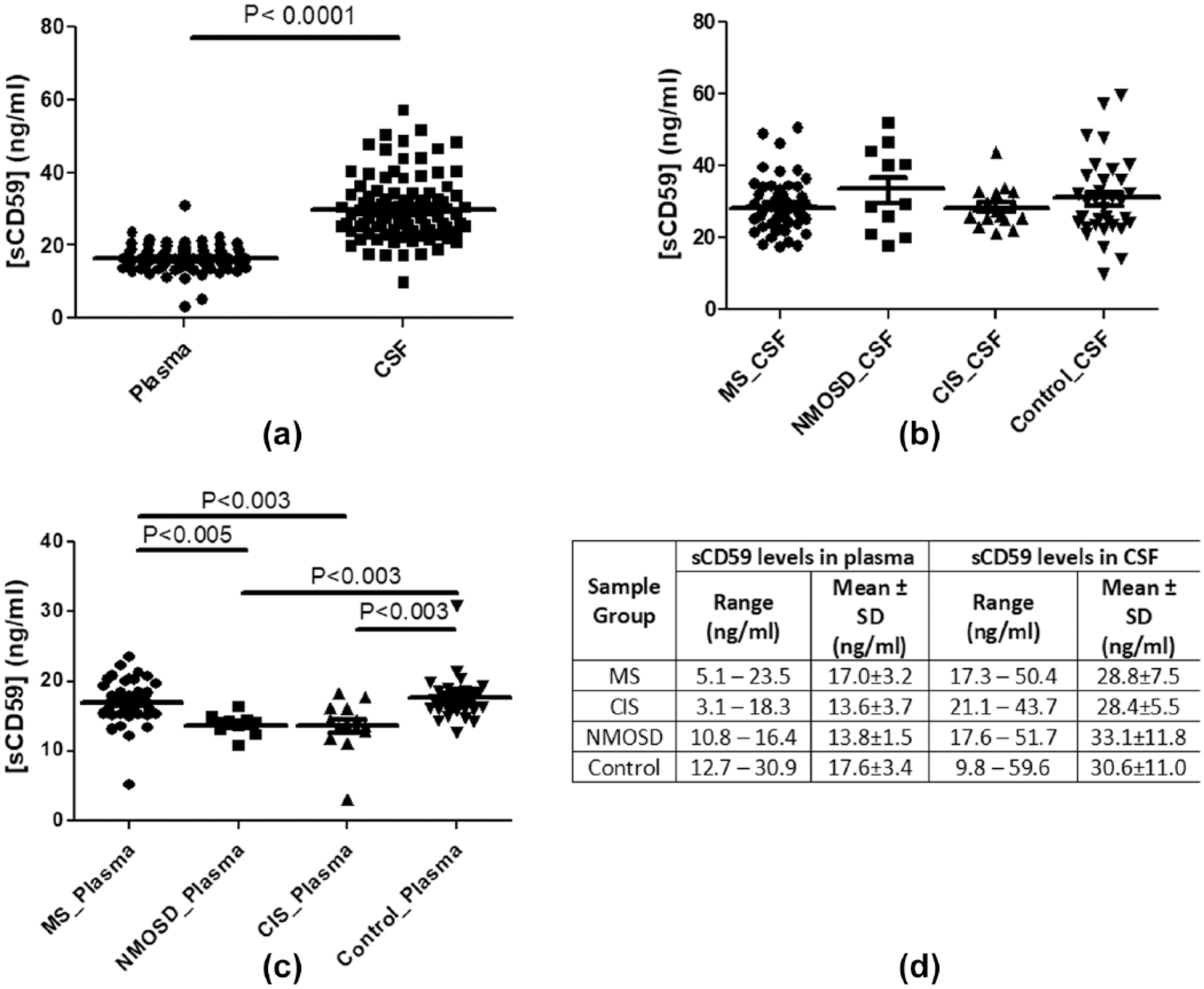
Measuring sCD59 in CSF and matched plasma samples: (a) sCD59 is measureable in all plasma and CSF samples tested and are ~2-fold lower in plasma compared to CSF; (b) CSF levels of sCD59 were not significantly different between the individual groups: MS, NMOSD, CIS or controls; (c) plasma levels of sCD59 in MS, NMOSD and CIS patients compared with normal controls differentiate MS from NMOSD (p < 0.005), MS from CIS and NMOSD/CIS from controls (p < 0.003); and (d) summary of the data.
The majority of the CD59 in CSF is not lipid-anchored to microparticles
sCD59 can exist in biological fluids either as an anchor-free soluble small molecule or as a GPI-anchored intact lipid/microparticle-associated molecule. 23 To test the nature of sCD59 found in CSF, those samples available in sufficient volume (23) were subjected to high-speed centrifugation to remove microparticles, and sCD59 concentration was measured prior to and after centrifugation. Levels of sCD59 in centrifuged CSF were not significantly different compared to uncentrifuged (mean concentration before centrifugation: 32.5 ng/mL; after centrifugation: 28.1 ng/mL), confirming that majority of the CD59 (>85%) in CSF is not anchored to microparticles (Figure 2). As an independent verification of this finding, CSF was passed through a 0.2-µM filter; sCD59 level was not significantly altered by filtration (data not included).
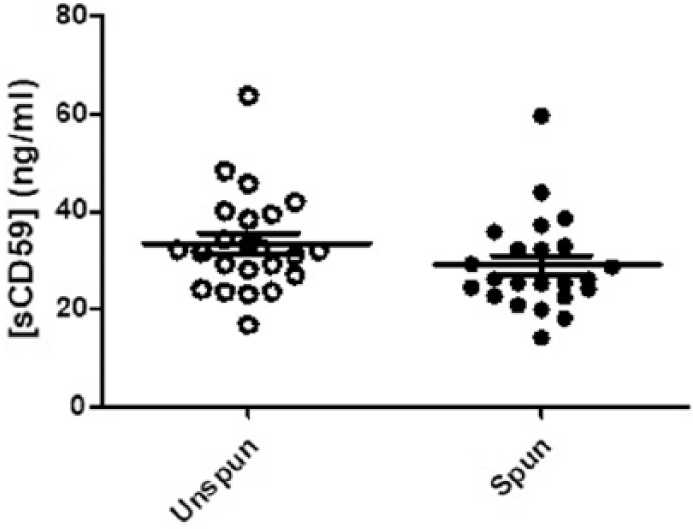
Measurement of sCD59 in CSF; samples from 23 individuals (MS = 9, NMOSD = 2, CIS = 4 and control = 8) were measured before (unspun) and after (spun) high-speed centrifugation; no significant difference seen in sCD59 levels between both groups (mean; unspun: 28.1 ng/mL, spun: 32.5 ng/mL).
sCD59 detected in patients CSF samples resembles cell-associated CD59
The nature of the sCD59 reactivity in MS, NMOSD and control CSF samples was tested by western blot analysis. In each sample tested, CSF sCD59 comprised a broad band of 19–23 kDa, slightly higher in molecular weight (MW) and more compact than the band obtained in erythrocyte lysates and resembling published data on nucleated cell-derived CD59 24 (Figure 3(a) and (b)).
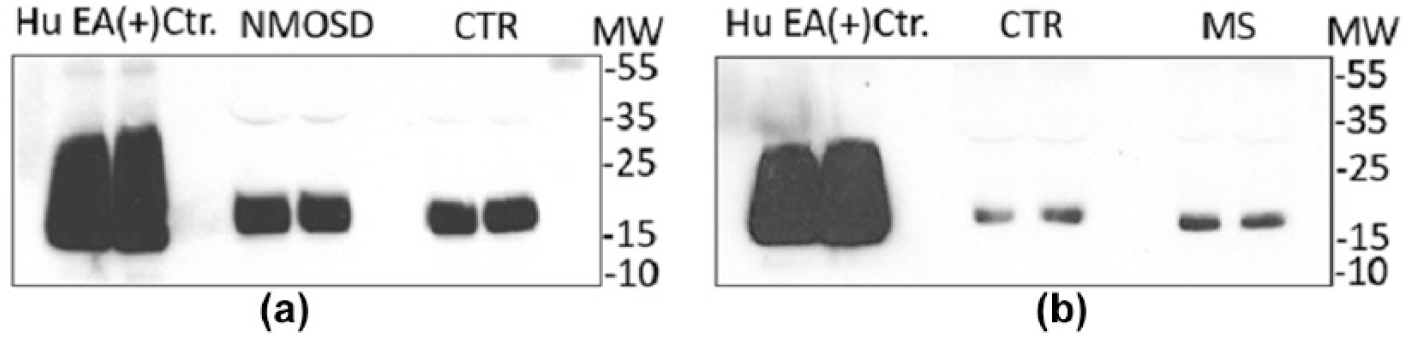
Detection of sCD59 in CSF patient samples using western blot; CSF samples were concentrated 10× (MS = 3, NMOSD = 3 and control = 6): (a) samples with higher concentrations of sCD59 as measured in the ELISA (NMOSD and control) and (b) samples with lower concentration of sCD59 in ELISA (MS and control). In all cases, 10-fold concentrated sample (25 µL per line) was resolved on 4%–20% gradient PAGE under non-reducing conditions and processed for western blot with MEM43 (monoclonal anti-CD59).
The CP and brain vasculature are enriched in CD59
Using a mouse monoclonal antibody to CD59, we demonstrated strong anti-CD59 immunoreactivity at the brain-CSF and BBBs of the human brain. Anti-CD59 immunoreactivity was comparable between MS (Figure 4(a) and (b)) and non-inflammatory control brains (Figure 4(c) and (d)) and was notable in the parenchymal vasculature (both arteries and veins, staining endothelium strongly and occasional luminal staining likely representing erythrocyte aggregates), meningeal vasculature, ependymal lining of the ventricles and the CP epithelium (Figure 4). In contrast to the moderate to substantial immunoreactivity of these structures, cell-specific staining within the hippocampus and parahippocampal grey and white matter was absent or weak. This pattern of CD59 expression was verified using a polyclonal anti-CD59 antibody raised in rabbit (Figure 4(k)). We did not see an association between the extent of anti-CD59 immunoreactivity with age, gender, MS type, extent of tissue block pathology or with topographically associated areas of white or grey matter pathology in this preliminary pathological analysis. To confirm that the observed CD59 staining represented local biosynthesis, we used ISH; epithelial cells of the CP and ventricle wall and endothelium of blood vessels expressed transcripts for CD59 (Figure 4(m)–(p)).
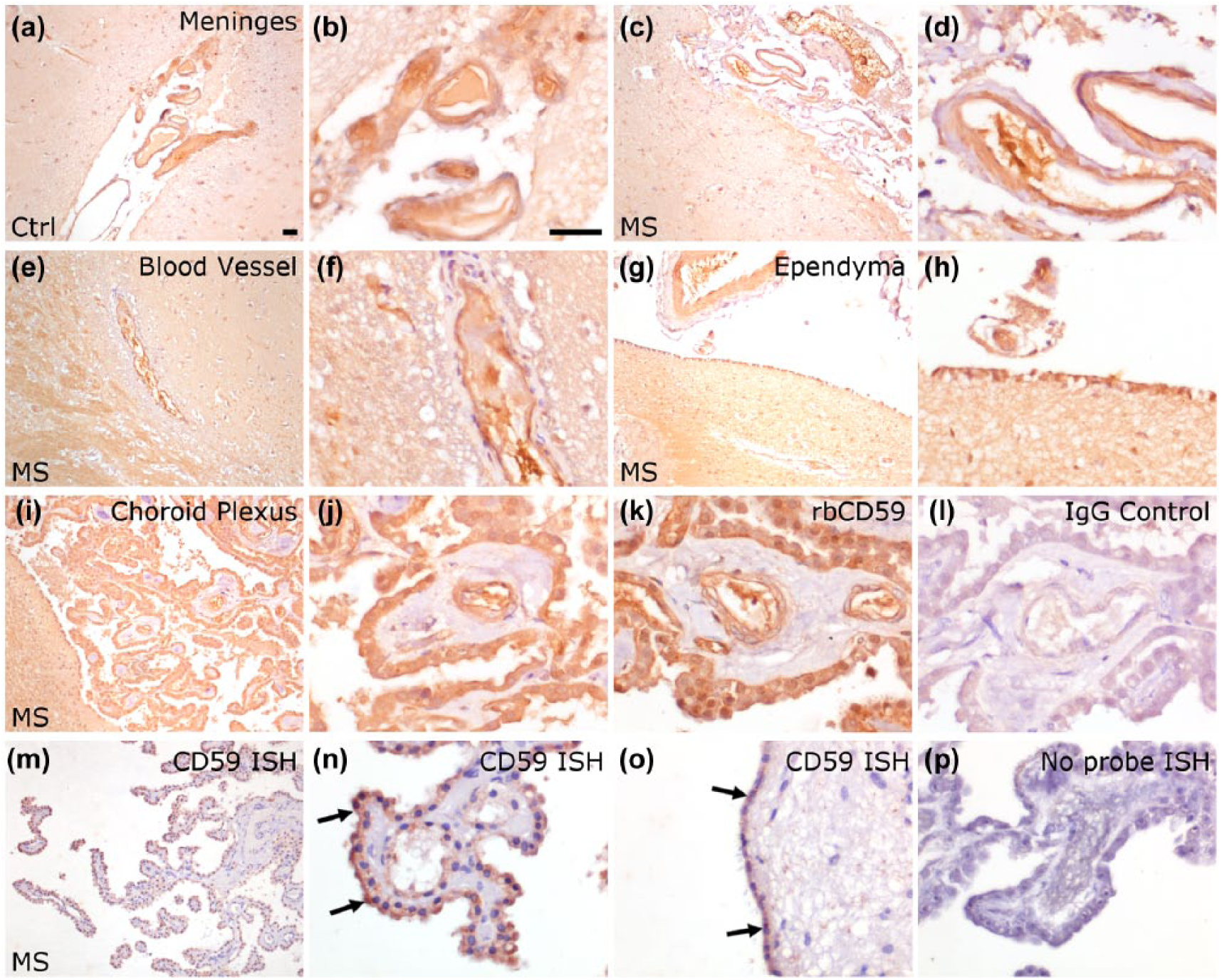
CD59 is expressed at vessel luminal surfaces and by cells lining the choroid plexus and ventricle wall. Immunohistochemistry of formalin-fixed paraffin-embedded MS and control hippocampus revealed strong staining with monoclonal anti-CD59 in the vasculature of the meninges (a–d), parenchymal vessels (e and f), cells of the ventricle wall (ependyma of the inferior horn of the lateral ventricle (g and h) and epithelial cells of the choroid plexus (i and j). An identical pattern of staining was seen with a polyclonal anti-CD59 antibody (k). Staining with an irrelevant mouse (l) or rabbit (not shown) IgG was negative. ISH confirmed that epithelial cells of the choroid and ventricle walls (m and o, arrows) and vessel endothelium (n) expressed CD59 transcript. No signal was seen in the absence of the CD59 antisense oligoprobe (p) or sense probe (not shown). All images captured with a 10× (a, c, e, g and i) or 40× (b, d, f, h, j, k and l) objective. Scale bar = 50 µm shown in (a) and (b).
Discussion
Measurement of complement proteins, regulators and activation products in CSF has been described in several studies and proposed as contributory biomarkers in demyelinating and degenerative neurological diseases. Levels of C3 and FH were reduced in Alzheimer’s disease CSF and correlated with disease staging. 21 The soluble terminal complex sC5b-9 was increased in CSF in both MS and NMOSD, more so in the latter, indicating more complement activation. 25 Soluble complement receptor 2 (sCR2) levels were increased in MS CSF and suggested to be of functional relevance, but its source was not explored. 26 A recent study reported that levels in CSF of the soluble form of the GPI-anchored cell surface complement regulator CD59 (sCD59) were significantly elevated in MS and NMOSD compared to non-inflammatory neurological disease controls provoked our interest. 16 We set out to replicate these findings in a larger sample and to explore the source of sCD59 in CSF.
Expression of complement regulators, including CD59, in the human brain has been the subject of previous work. CD59 expression in the normal, non-inflamed brain is very low with minimal expression on glia and negligible expression on neurones. 3 The only sites in the normal brain where CD59 is abundantly expressed are the vascular endothelium and the CP where CD59 is reported to decorate the apical surface of the ependymal cells. In the inflamed brain, expression of complement regulators, including CD59, is increased, although levels remain low compared to the periphery. 27 It is likely that this increase in regulator expression is a homeostatic response as levels of complement proteins are increased in the inflamed brain due either to intrathecal production or transudation of complement proteins across the leaky BBB. 28
We measured sCD59 levels in CSF from 50 MS, 11 NMOSD, and 17 CIS patients and 34 non-inflammatory controls. CSF sCD59 levels were ~30 ng/mL in the samples and were not different between the groups; in particular, levels were not higher in MS and/or NMO compared to controls in contrast to recent report. 16 In 91 of the available cases, matched plasma taken at the time of lumbar puncture was available; in these, we measured plasma sCD59 levels in the same assay.
In contrast to published work, 16 our results do not support the conclusion that CSF sCD59 levels are a powerful biomarker of NMOSD and MS. There was no significant difference between the disease groups in our study. The reasons for this failure to replicate are uncertain. Our cohort was overwhelmingly comprised of Caucasians with well-characterised disease followed up over a prolonged period; the published study was from Japan and thus was very different in terms of ethnicity. We used a commercial assay using validated anti-CD59 monoclonal antibodies and a recombinant protein standard; the published study used a different commercial CD59 ELISA assay (by MyBioSource, Inc. (San Diego, CA, USA), detection range; 31.25–2000 pg/mL) with no details of its architecture. Our measured values were approximately 100-fold higher than those in the previous report (~30 ng/mL, cf. ~300 pg/mL); of note, our ability to detect sCD59 by western blotting in 10-fold concentrated CSF supports our finding of higher levels.
CD59 is a GPI-anchored membrane-associated molecule, and its appearance in the fluid phase requires either shedding on lipid vesicles with an intact GPI anchor or enzymatic anchor cleavage and release as a soluble, unanchored molecule; sCD59 in urine has been shown to be anchor-free, likely derived by enzymatic cleavage from renal tubular cells.29–31 In order to investigate the nature of sCD59 in CSF, we performed high-speed centrifugation to remove microparticles and remeasured sCD59 in a subset of CSF samples. Measured levels were little affected by centrifugation, indicating that CSF sCD59 was anchor free; this was further confirmed by showing that passage of the few CSF samples available in sufficient quantities through a 0.22-µM filter did not reduce measured sCD59 levels. Characterisation of sCD59 by western blotting showed that it ran as a 19- to 23-kDa band closely resembling published blots of urine-derived CD59. 32
To explore the source of sCD59 in CSF, we stained human brain sections; as previously reported, CD59 staining in brain parenchyma was restricted to vessel endothelium and lumen. In sharp contrast, CP staining was abundant, co-localised with vessels and with ependyma, specialised glial cells responsible for production of the CSF. We suggest that enzymatic shedding from this surface represents the likely source of CSF sCD59; this suggestion is supported by ISH that demonstrates the presence of CD59 specific RNA transcript in the choroid epithelium and vascular endothelium.
We also measured sCD59 in matched plasma from the majority of cases and controls; plasma sCD59 was significantly lower in NMOSD and CIS samples compared with controls, although there was considerable overlap between groups. A surprising finding was that CSF levels of sCD59 were on average double those in matched plasma. A possible explanation of this is that although sCD59 might be generated in peripheral tissues, its small size when anchor free means that it is excreted first-pass through the kidney where it accumulates in urine; in CSF, sCD59 accumulates because the BBB is impermeant to molecules larger than 600 Da. 33 Whether the (relatively) high amounts of sCD59 in CSF are of any functional relevance remains to be demonstrated. In principle, sCD59 might contribute to homeostasis by dampening down complement activation in the CSF; however, in other biological fluids, sCD59 is a poor regulator because it associates with other proteins. 34
In conclusion, we show that a non-anchored soluble form of sCD59 is present in CSF in health and neuroinflammatory diseases at similar levels, that CSF levels of sCD59 are consistently higher than in matched plasmas and that the likely source of CSF sCD59 is the CP, a site of abundant expression in the normal and inflamed brain.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) declared receipt of the following financial support for the research, authorship and/or publication of this article: WMZ was supported by a PhD Studentship from the Life Sciences Research Network Wales.