
Abstract
Background:
Epiretinal membranes (ERMs) consist of glial and fibrocellular tissue, potentially arising from pathologic microgliosis, macrogliosis, and/or retinal pigment epithelium (RPE) cell migration. The clinical relevance of ERMs in people with multiple sclerosis (PwMS) remains largely unstudied.
Objectives:
To investigate ERM prevalence and their associations with retinal layer thicknesses and disability measures in PwMS.
Methods:
PwMS exhibiting ERMs (ERM+) were matched 1:1 with PwMS without ERMs (ERM−) by optic neuritis (ON) history, multiple sclerosis (MS) subtype, age, sex, and race. Disability assessments included high-contrast/low-contrast letter acuity (HC-/LCLA), Expanded Disability Status Scale (EDSS), walking speed test (WST), manual dexterity test (MDT), and processing speed test (PST) scores.
Results:
Among 1264 participants screened (1120 PwMS and 144 healthy controls [HCs]), ERMs were observed in 206 PwMS (18.4%) and 17 HCs (11.8%). ERM+ PwMS exhibited higher EDSS (1.0 ± 0.3, p = 0.002), longer WST (0.8 ± 0.2 seconds, p < 0.001), longer MDT (2.2 ± 0.5 seconds, p < 0.001), lower PST (−3.0 ± 1.4, p = 0.08), lower LCLA (−4.2 ± 1.1 letters, p < 0.001), and lower ganglion cell-inner plexiform layer (−1.8 ± 0.8 µm, p = 0.03), inner nuclear layer (−0.8 ± 0.3 µm, p = 0.01), outer nuclear layer (−1.6 ± 0.6 µm, p = 0.01), and RPE (−0.5 ± 0.1 µm, p < 0.001) thicknesses. Higher ERM stage correlated with greater disability.
Conclusion:
ERMs may identify a distinct MS phenotype, potentially characterized by pathological microglial and/or macroglial activity. ERMs may be a marker of similar global processes throughout the central nervous system that potentially contribute to greater overall disability.
Introduction
Multiple sclerosis (MS) is an autoimmune, demyelinating central nervous system (CNS) disorder, in which inflammation is initially driven mostly by pathological adaptive immune cells. 1 However, microglia and astrocytes also play pivotal roles in neuroinflammation and neurodegeneration in MS through spatially and temporally dynamic interactions. 2 Activated microglia are not confined to focal demyelinating lesions, but are also observed in normal-appearing white and gray matter, suggesting their involvement in early MS pathogenesis.3,4 Microglia initiate and propagate inflammation by presenting antigens to T cells, releasing pro-inflammatory cytokines, while also facilitating repair through phagocytosis.5,6 Furthermore, activated microglia induce reactive astrocytosis, further compromising the blood–brain barrier, promoting leukocyte infiltration, demyelination and axonal injury. 7
Chronic active compartmentalized inflammation (smoldering neuroinflammation) within the CNS is recognized as a key contributor to MS progression. 8 Chronic activation of CNS-resident immune cells, including microglia and autoreactive B cells, is thought to drive diffuse neurodegenerative processes in MS.9,10 In addition, bioenergetic disturbances, such as mitochondrial dysfunction 11 and oxidative stress, 12 exacerbate axonal and neuronal injury and degeneration. While acute lesion activity may be dominated by infiltrating macrophages and T cells, chronic glial activation may drive progressive CNS damage. 8
Retinal microglia share similarities with brain microglia. 13 In people with multiple sclerosis (PwMS), ganglion cell-inner plexiform layer (GCIPL) thickness correlates with global disability measures and brain volumes, suggesting that retinal layer thicknesses reflect global neurodegenerative processes.14,15 Moreover, inner nuclear layer (INL) and outer nuclear layer (ONL) changes are also observed and may be clinically relevant.16–18
Müller glia reside in the INL and have radial processes extending to all retinal neuronal layers to regulate fluid and ionic homeostasis.19,20 Under pathologic conditions, Müller cells may fail to clear excess fluid, thereby contributing to capillary leakage and edema.20,21 Similar to microglia, Müller cells can migrate in response to retinal injury.22,23 This response includes extending their radial processes to the vitreous surface, where they promote the formation of fibrotic connective tissue known as epiretinal membranes (ERMs).24,25
ERMs are fibrocellular proliferations on the internal limiting membrane (ILM), consisting of myofibroblast-like cells, macrophages, fibrocytes, fibrous astrocytes, and retinal pigment epithelial (RPE) cells. 25 Previous studies have reported that ERMs are present in 7%–11% of the healthy individuals.26,27 ERMs can be idiopathic or secondary to retinal vascular diseases, diabetes, trauma, or inflammatory conditions. 25 The pathogenesis of ERM formation involves microglial migration, residual vitreous cortex retention, and ILM avulsion defects, thereby providing complementary insights into how cellular and cytokine-mediated responses converge to produce this retinal phenomenon. 25
ERMs are readily appreciable on optical coherence tomography (OCT) scans, but they remain understudied in MS. In PwMS without other retinal diseases, ERMs might serve as a marker of Müller gliosis due to retinal inflammation and neurodegeneration. We hypothesize that MS may cause secondary ERMs via migration of activated microglia and/or macroglia (i.e. Müller cells). This might reflect similar processes occurring throughout the CNS, identifying greater susceptibility for disability accumulation. We aimed to determine ERM prevalence in PwMS and their relationships with global disability measures, serum neurofilament light chain (sNfL) levels, and retinal layer thicknesses.
Methods
Participants
We retrospectively screened MS participants and healthy controls (HCs) from an ongoing observational study at the Department of Neurology at Johns Hopkins University (JHU). The study was approved by the institutional review board of JHU and adhered to the tenets of the Declaration of Helsinki. Written informed consent was obtained from all study participants.
Inclusion criteria for MS participants were as follows: (1) age between 18 and 75 years and (2) an MS diagnosis in accordance with the 2017 revised McDonald diagnostic criteria. 28 MS participants with known non-MS-related neurological or ophthalmological disorders (e.g. uveitis, trauma, and retinal tears) were excluded. All participants underwent a combination of clinical and paraclinical evaluations for every documented episode of optic neuritis (ON). Eyes with ON within 6 months before OCT acquisition were excluded due to transient peripapillary retinal nerve fiber layer (pRNFL) swelling.
Disability measures
MS disability was assessed by the Expanded Disability Status Scale (EDSS) and multiple sclerosis performance test (MSPT: 25-foot walking speed test [WST], manual dexterity test [MDT] of the dominant and non-dominant hands, and processing speed test [PST]). 29
As standard care at the JHU MS center, all patients are recommended to undergo annual ophthalmological assessments with a general ophthalmologist or neuro-ophthalmologist. All participants underwent monocular and binocular visual acuity assessments using their habitual corrective lenses, beginning with low-contrast letter acuity (LCLA) using 2.5% contrast Sloan letter charts at a distance of 2 m, followed by high-contrast letter acuity (HCLA) testing with ETDRS charts at 4 m. All charts were mounted on a retro-illuminated light box (Precision Vision, LaSalle, Illinois, USA) in a darkened room, with the number of letters correctly identified (range: 0–70 letters) recorded.
Optical coherence tomography
OCT scans were acquired using Cirrus HD-OCT (Carl Zeiss, Dublin, CA, USA; Model 5000). Each scan underwent rigorous quality control in accordance with the OSCAR-IB criteria 30 and study reporting is consistent with APOSTEL 2.0 recommendations. 31 Scans that did not meet all OSCAR-IB criteria were excluded. All scans were manually reviewed by experienced physicians and technicians to ensure scan quality and segmentation accuracy. The identification and grading of ERM pathology was conducted exclusively by clinicians with relevant expertise.
pRNFL thicknesses were determined using the device-incorporated segmentation of scans acquired using the Optic Disc Cube 200 × 200 protocol. 32 Macular scans were acquired with the 512 × 128 macular cube protocol. 32 Retinal layer thicknesses in the macular region were calculated within a 5-mm-diameter annulus sparing the fovea, using a validated JHU-developed, openly available segmentation algorithm. 33
We modified an existing ERM classification system by adding an additional Stage 0, thereby expanding it from four to five stages. 34 Stage 0 ERMs were minimally visible (spanning less than three B scans, see Supplemental Methods for more details); Stage 1 ERMs appeared thin, with a preserved foveal depression; Stage 2 ERMs showed features of Stage 1 ERMs, along with loss of the foveal depression and widening of the ONL; Stage 3 ERMs were defined by the presence of continuous ectopic inner foveal layer (EIFL) spanning the entire foveal area, while the retinal layers remained distinguishable; Stage 4 ERMs were thick, along with having EIFL and distorted retinal layers (Figure 1).
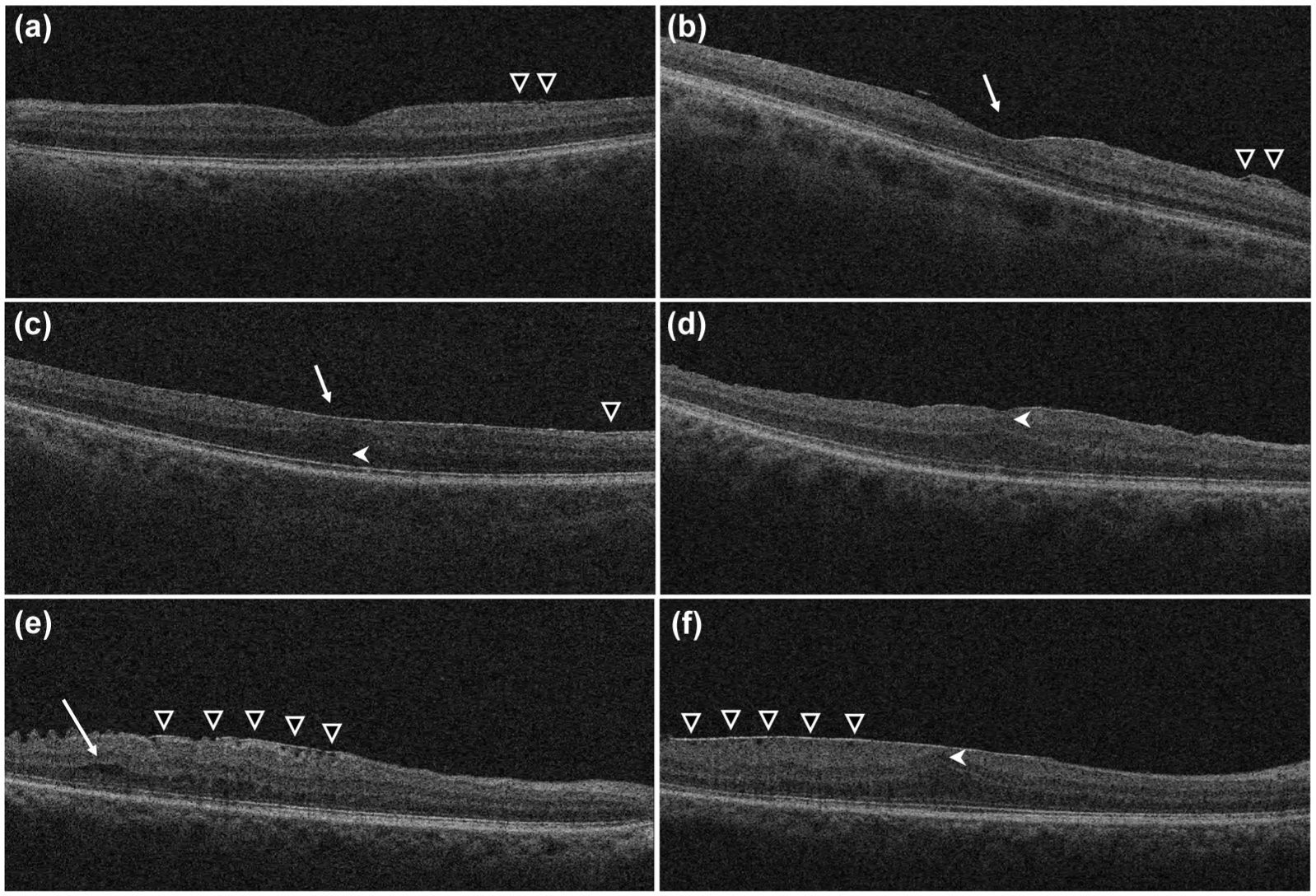
OCT scans illustrating the staging of ERMs. (a) Stage 0: Minimally visible ERMs (white triangle); (b) Stage 1: Thin ERMs (white triangle) with preserved foveal depression (white arrow); (c) Stage 2: Thin ERMs (white triangle) with loss of the foveal depression (white arrow) and widening of the outer nuclear layer (white arrowhead); (d) Stage 3: ERMs with continuous ectopic inner foveal layers extending across the entire foveal area (white arrowhead), without distortion of the retinal layers; (e and f) Stage 4: Thick ERMs (white triangle) along with ectopic inner foveal layers (white arrowhead) and distorted retinal layers (white arrow). e and f are from the same patient’s eye in different locations on the retina.
To assess the inter-rater reliability of ERM identification and staging, three experienced raters each evaluated an independent set of 85 OCT scans, which were not included in the main analyses. An initial set of 30 scans was used for standardization and training, with the remaining 55 scans used for the final agreement analyses. Inter-rater agreement for ERM identification (presence versus absence) was assessed using Fleiss’ Kappa and showed strong agreement (κ = 0.88, p < 0.001). Inter-rater agreement for ERM severity staging (Stages 0–4) was evaluated using Kendall’s Coefficient of Concordance (Kendall’s W) to account for the ordinal classification. This analysis also indicated relatively strong agreement among raters (W = 0.71, p < 0.001).
Serum neurofilament light chain measures
sNfL levels were measured using the Quanterix Simoa® NF-light assay or the Siemens Atellica® Solution platform. To ensure consistency, sNfL values obtained from the Siemens platform were converted to equivalent Quanterix values. 35 An age-adjusted sNfL reference database was established from 160 HCs using generalized additive models for location, scale, and shape methods, and sNfL values from PwMS were transformed into age-adjusted Z-scores accordingly. 36
Statistical analysis
We performed 1:1 matching between PwMS with ERMs (ERM+) and PwMS without ERMs (ERM−) according to the known history of prior ON, MS subtype, age, sex, and race. For ON history matching, we specifically considered ON history that occurred in the ERM-affected eye(s) of PwMS in the ERM+ group due to the possibility, albeit low, that the presence of ERMs might be related to prior ON. Furthermore, participants with a surgical history of ERM peeling were explicitly excluded from the ERM− group.
Comparisons of disability measures and sNfL age-adjusted Z-scores between the ERM+ and ERM− groups were performed using Student’s t-test or Wilcoxon rank sum test, as appropriate. OCT-derived measures were compared between groups using linear mixed-effect models (LMMs), accounting for within-subject, inter-eye correlations. The marginal and conditional coefficients of determination were calculated with pseudo-R-squared. 37 Since ON matching was based on ERM and ON history in the same eye, some eyes in the ERM+ group had a history of ON without exhibiting ERMs, leading to a higher number of ON eyes in the ERM+ group, as compared to the ERM− group. Therefore, we conducted two sensitivity analyses: one adjusting for ON history (independent of ERM presence) and another completely excluding ON eyes without ERMs in the ERM+ group.
The comparisons of OCT-derived measures, disability measures, and sNfL age-adjusted Z-scores across different ERM stages were performed using LMMs (with ERM stages classified at the eye level) or generalized linear models (with ERM stages classified based on the higher stage between eyes), as appropriate. An interaction term between age and ERM stage was planned to be included if exploratory analyses of nested models indicated statistical significance. If no significant interaction was found, age was instead to be included as a fixed effect without interaction with ERM stages. Post hoc pairwise comparisons between ERM stages were performed using estimated marginal means adjusted for age, with p-values corrected for multiple comparisons from Tukey’s honestly significant difference test.
All analyses were conducted using R statistical software (version 4.4.2), with statistical significance defined as a two-tailed p value < 0.05.
Results
Of the 1120 PwMS and 144 HCs screened (Figure 2 and Supplemental Table 1), ERMs were detected in 206 PwMS (18.4%) (ERM+ group) and 17 HCs (11.8%). Retinal structural and visual functional measure comparisons between healthy eyes with or without ERMs were described in Supplemental Table 2. Among the 206 ERM+ MS participants (412 eyes), ERMs were present in 270 eyes (65.5%): 104 eyes (38.5%) at Stage 0, 128 eyes (47.4%) at Stage 1, 31 eyes (11.5%) at Stage 2, 6 eyes (2.2%) at Stage 3, and 1 eye (0.4%) at Stage 4. A matched cohort of 206 PwMS without ERMs in both eyes (ERM− group) was selected based on prior ON history, MS subtype, age, sex, and race. The ERM+ group contained 67 ON eyes with ERMs and 25 ON eyes without ERMs. Since ON matching was based on ERM and prior ON history within the same eye, the ERM− group was matched according to the 67 ON eyes with ERMs in the ERM+ group, resulting in 67 ON eyes in this ERM− group. There were no differences between the ERM+ and ERM− groups in disease duration, disease-modifying therapy (DMT) usage, or the proportion of individuals receiving high-efficacy versus low-intermediate efficacy DMTs (Table 1). While sphingosine-1-phosphate receptor modulators may affect retinal layer thicknesses, the number of participants receiving these agents was very low and balanced between the ERM+ (n = 5) and ERM− (n = 7) groups.
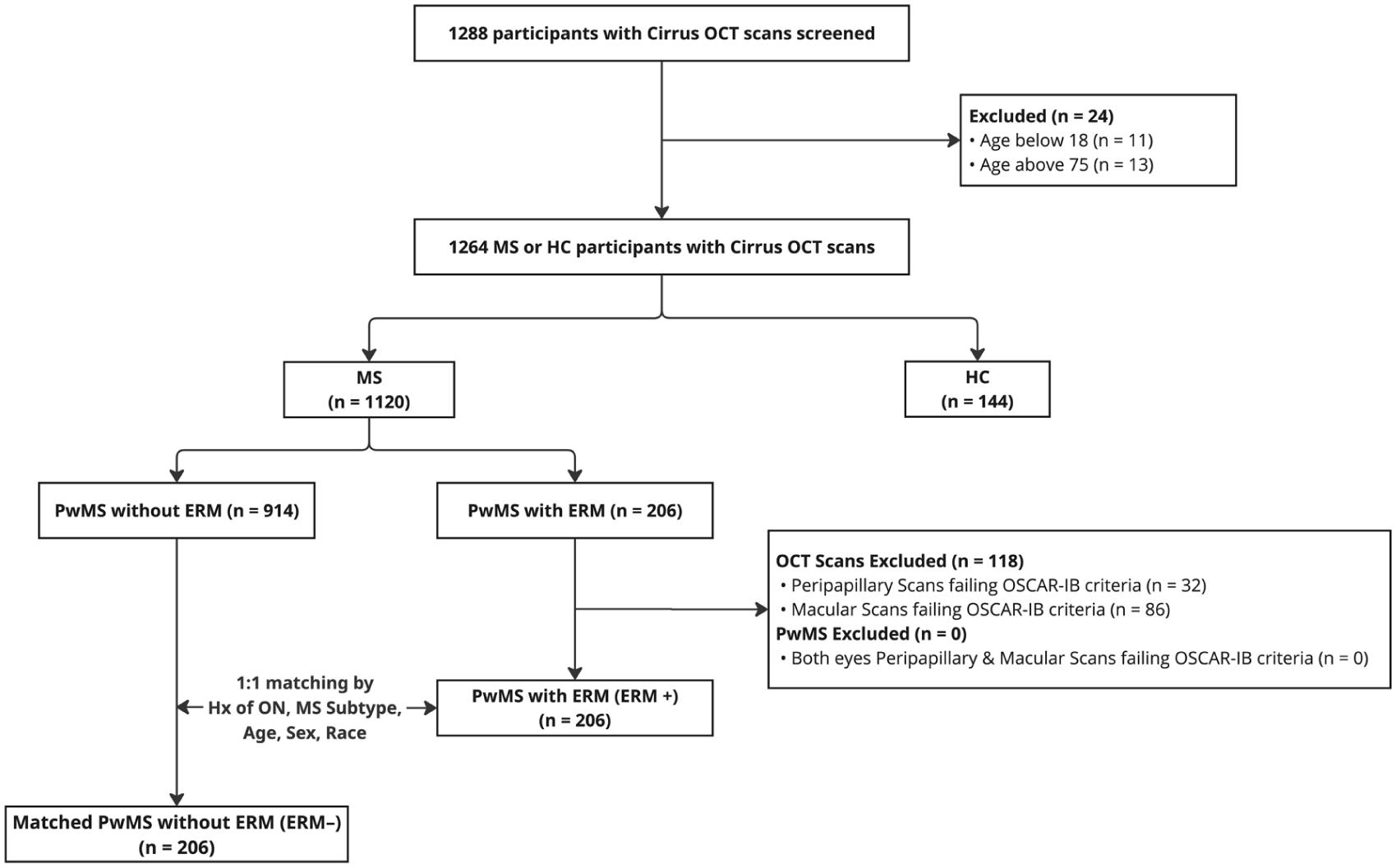
Consort chart regarding selection of participants.
Demographics and clinical characteristics of PwMS with and without ERMs.
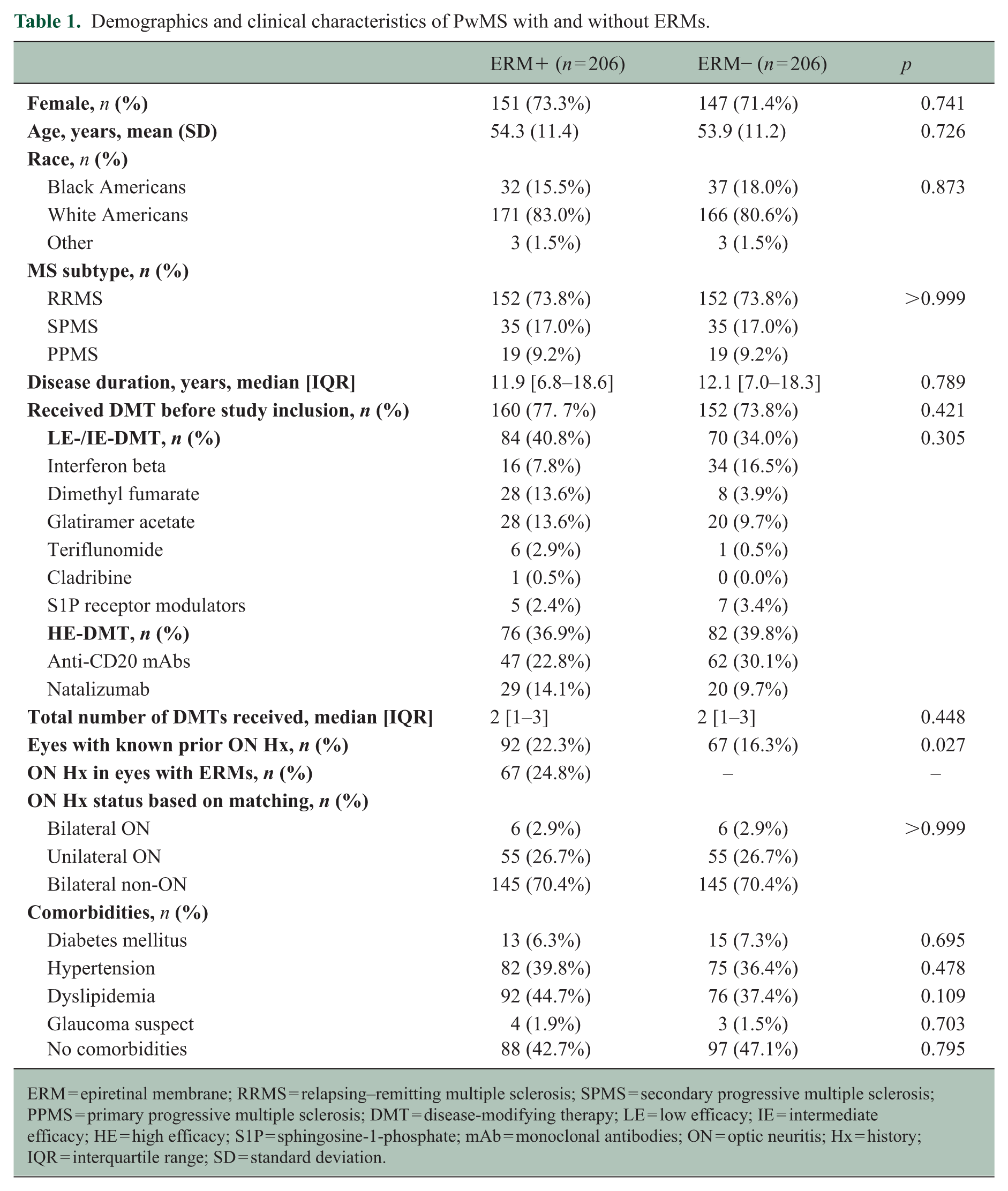
ERM = epiretinal membrane; RRMS = relapsing–remitting multiple sclerosis; SPMS = secondary progressive multiple sclerosis; PPMS = primary progressive multiple sclerosis; DMT = disease-modifying therapy; LE = low efficacy; IE = intermediate efficacy; HE = high efficacy; S1P = sphingosine-1-phosphate; mAb = monoclonal antibodies; ON = optic neuritis; Hx = history; IQR = interquartile range; SD = standard deviation.
Relationships between ERMs and disability measures in PwMS
The presence of ERMs was associated with worse performance across numerous disability assessments (Table 2). The ERM+ group had higher EDSS scores than the ERM− group (1.0 point higher; p = 0.002), required more time to complete the WST (0.8 seconds longer; p < 0.001) and MDT (2.2 seconds longer; p < 0.001), as well as demonstrated lower PST performance (3 points lower, p = 0.081). Due to the positively skewed distributions of the WST and MDT times, log-transformed analyses were performed, which also confirmed these findings. In addition, ERM+ PwMS exhibited higher age-adjusted sNfL Z-scores, as compared to the ERM− group (p = 0.042).
Group comparisons of disability measures and sNfL.
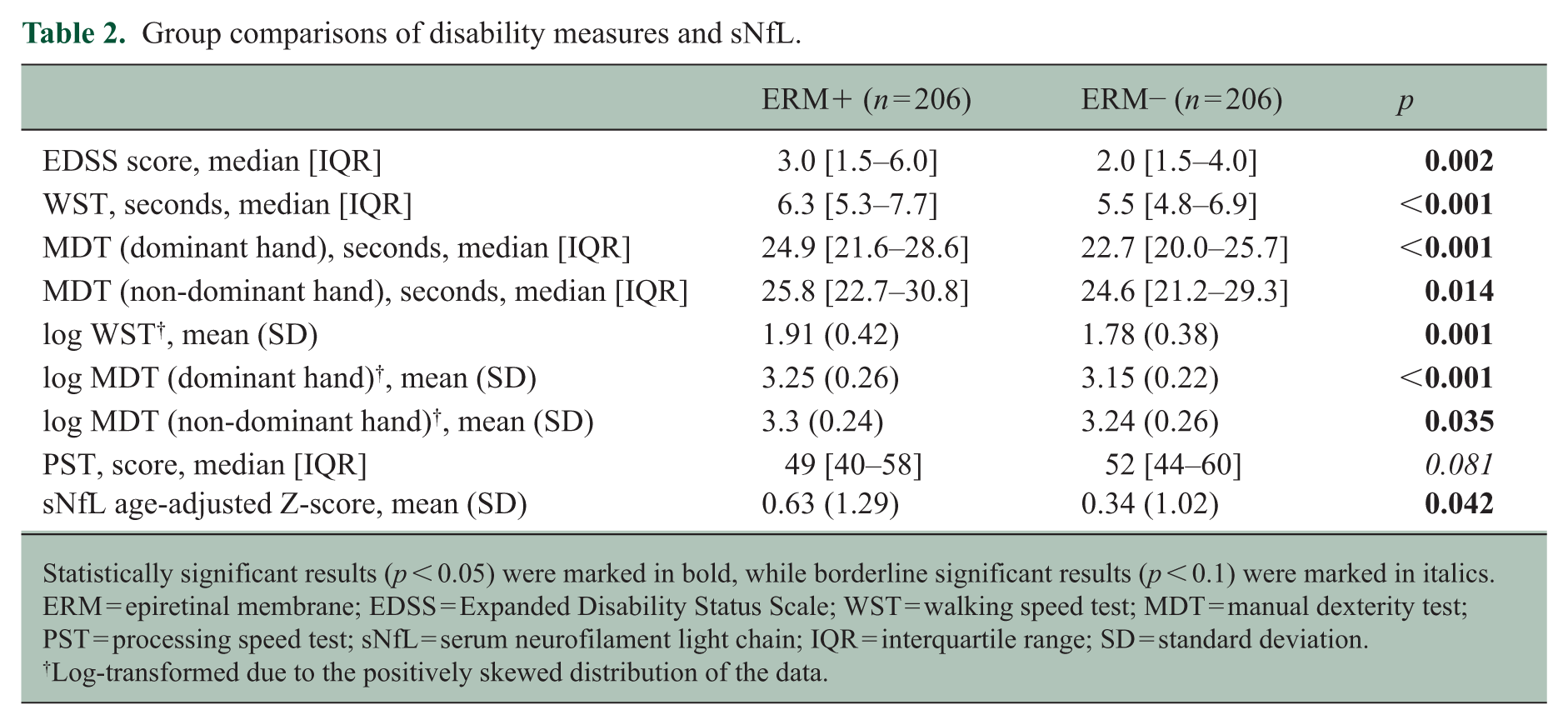
Statistically significant results (p < 0.05) were marked in bold, while borderline significant results (p < 0.1) were marked in italics. ERM = epiretinal membrane; EDSS = Expanded Disability Status Scale; WST = walking speed test; MDT = manual dexterity test; PST = processing speed test; sNfL = serum neurofilament light chain; IQR = interquartile range; SD = standard deviation.
Log-transformed due to the positively skewed distribution of the data.
Visual function was also lower in the ERM+ group across all testing paradigms. Compared with the ERM− group, binocular performance in HCLA and LCLA was lower by 1.8 and 4.3 letters, respectively, in the ERM+ cohort. Similarly, monocular HCLA and LCLA scores were lower in the ERM+ relative to the ERM− group (Table 3).
Comparison of visual function measures between eyes in the ERM+ and ERM− groups.
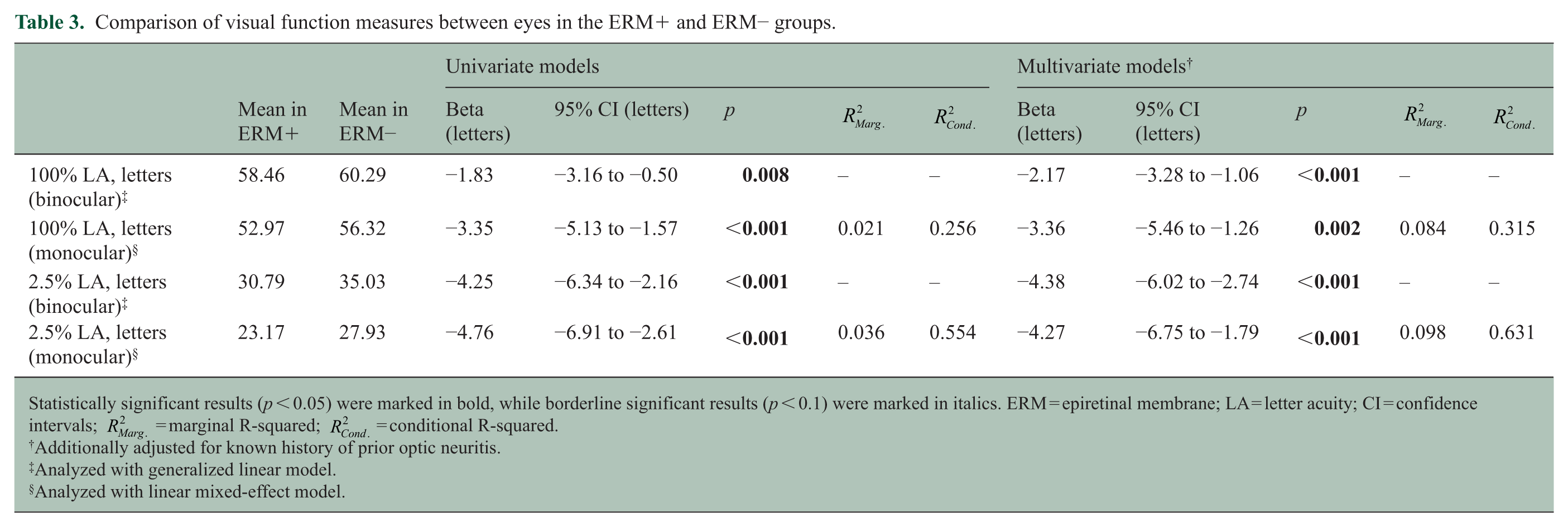
Statistically significant results (p < 0.05) were marked in bold, while borderline significant results (p < 0.1) were marked in italics. ERM = epiretinal membrane; LA = letter acuity; CI = confidence intervals;
Additionally adjusted for known history of prior optic neuritis.
Analyzed with generalized linear model.
Analyzed with linear mixed-effect model.
Relationships between ERMs and OCT retinal layer thicknesses in MS eyes
The average pRNFL thickness did not differ between the ERM+ and ERM− groups (p = 0.385), although regionally, temporal quadrant pRNFL (tRNFL) thickness was 3.0 μm higher in the ERM+ relative to the ERM− group (p = 0.018). In the macular region, MS eyes in the ERM+ group exhibited lower GCIPL (−1.8 μm; p = 0.026), INL (−0.8 μm; p = 0.008), ONL (−1.6 μm; p = 0.010), and RPE (−0.3 μm; p < 0.001) thicknesses, as compared to MS in the ERM− cohort (Table 4).
Comparison of retinal layer thicknesses in eyes between the ERM+ and ERM− groups.
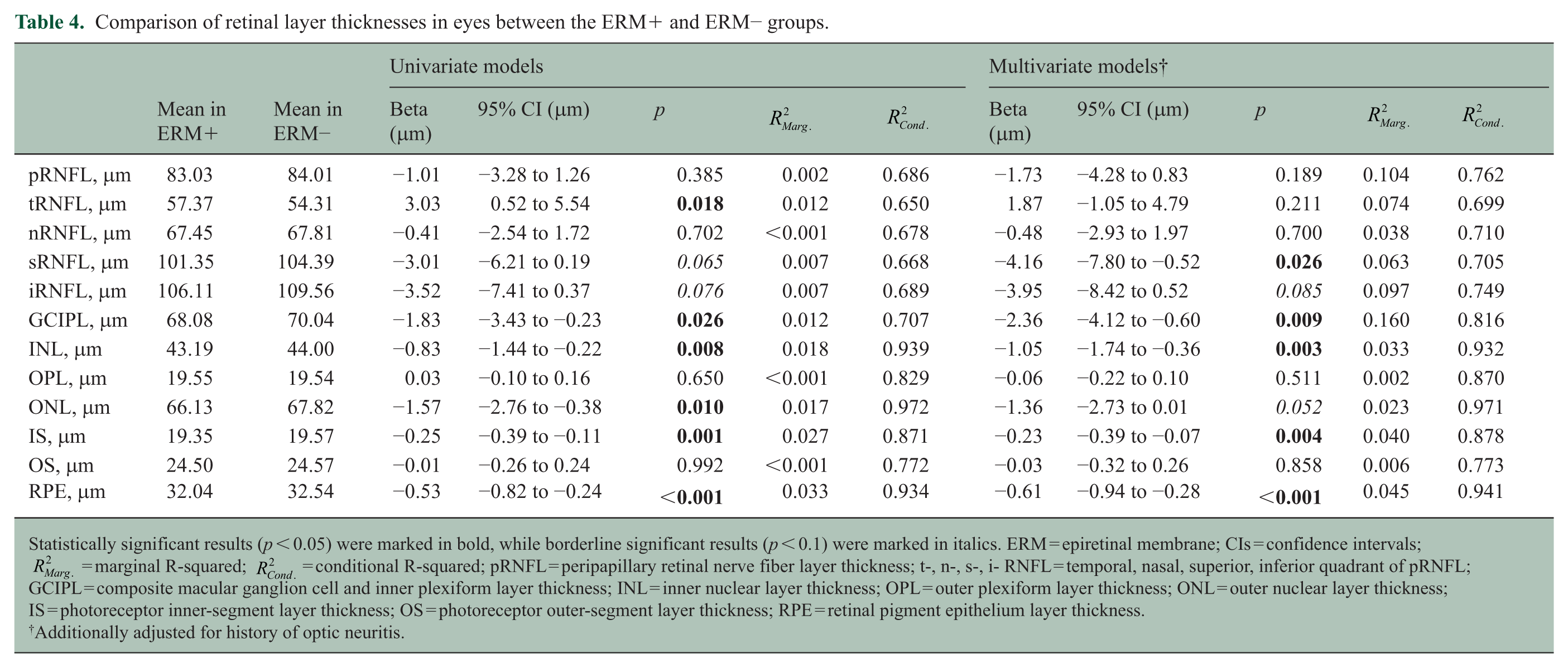
Statistically significant results (p < 0.05) were marked in bold, while borderline significant results (p < 0.1) were marked in italics. ERM = epiretinal membrane; CIs = confidence intervals;
Additionally adjusted for history of optic neuritis.
As described earlier, matching of prior ON was based on both ERMs and prior ON in the same eye. This introduced an imbalance in the number of ON eyes between the groups. Accordingly, two sensitivity analyses were conducted. In models also adjusting for prior ON history, eyes in the ERM+ group continued to show lower GCIPL, INL, ONL, and RPE thicknesses, along with lower HCLA and LCLA. However, no differences were observed in tRNFL thicknesses when adjusting for ON history (Tables 3 and 4). Furthermore, when excluding the 25 ON eyes without ERMs in the ERM+ group, the aforementioned findings remained similar (Supplemental Table 3).
To further investigate whether the effects of ERMs may be independent of MS-driven neurodegeneration, we evaluated a subgroup of eyes with prior ON, with and without ERMs (67 ERM+ ON eyes vs. 67 ERM− ON eyes). Within this subgroup, ERM+ ON eyes still exhibited worse visual acuity and lower GCIPL thicknesses, suggesting that ERMs might independently compound the structural and functional decline caused by optic neuropathy (Supplemental Table 4).
Sensitivity analyses accounting for comorbidities
Some PwMS in the studied cohort had known comorbidities (Table 1). Despite these comorbidities being well-controlled and without evidence of retinal damage related to these comorbidities, we conducted additional sensitivity analyses excluding all PwMS with any comorbidity to eliminate potential contributions to secondary ERM formation. After exclusion, the ERM+ group still demonstrated greater global disability (worse EDSS, WST, MDT, HCLA, LCLA), higher sNfL age-adjusted Z-scores, as well as lower GCIPL, INL, and RPE thicknesses, as compared to the ERM− group, although ONL thickness differences were no longer detected (Supplemental Table 5).
Associations of ERM severity with OCT and disability measurements
We further investigated the association of ERM severity with MS disability outcomes and OCT-derived measures. Considering that a primary mechanism of ERM formation may be related to aging, with higher-stage ERMs being more prevalent in older participants in this cohort (Supplemental Figure 1), interaction analyses were performed to first determine whether interaction terms need to be included in statistical models (Table 5). Of all the OCT and disability measurements, interaction effects were only significant for monocular HCLA (ηρ2 = 0.039, p = 0.026), EDSS (ηρ2 = 0.041, p = 0.019), and WST (ηρ2 = 0.053, p = 0.009) scores.
Interaction analyses of ERM stages with age in ERM+ PwMS.
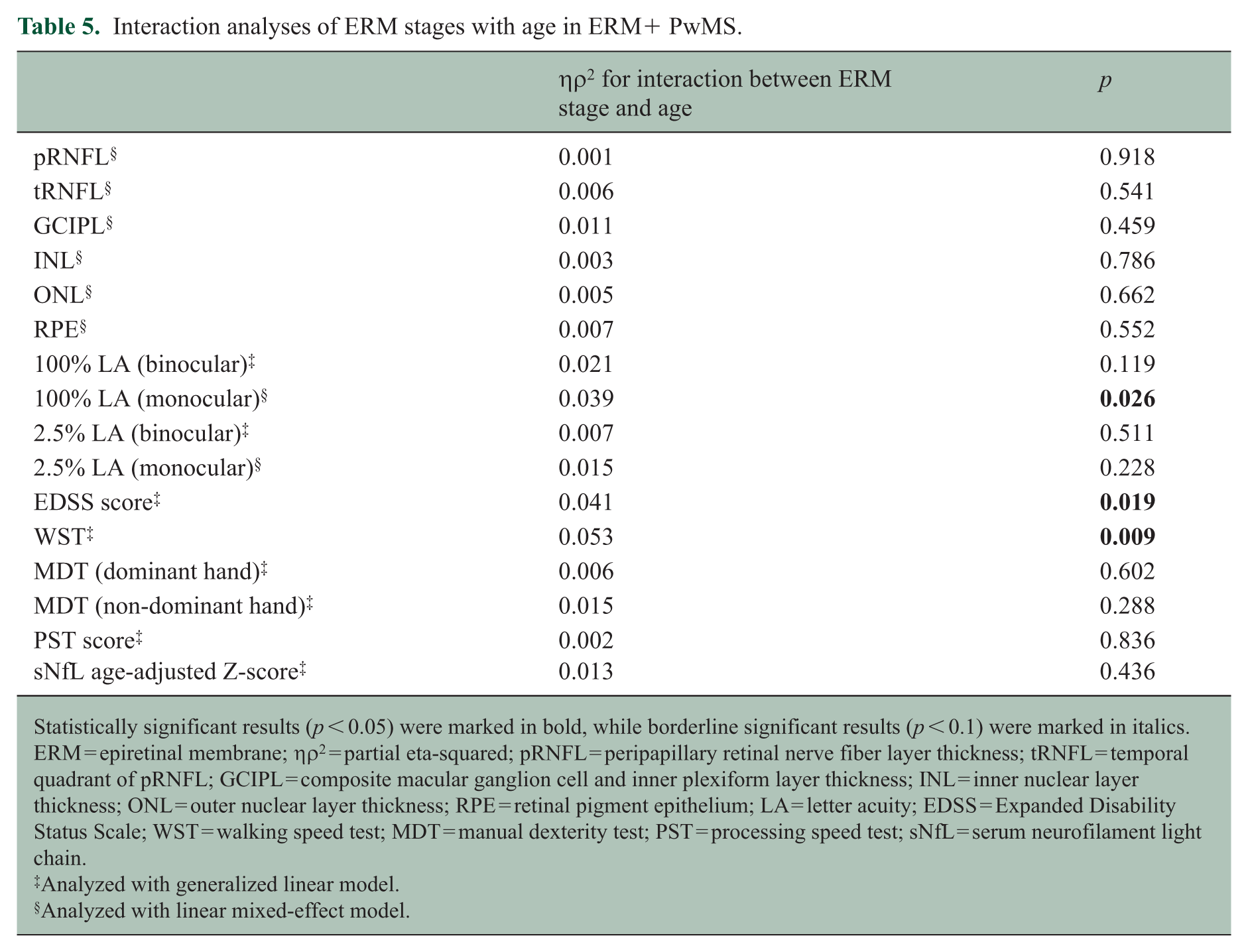
Statistically significant results (p < 0.05) were marked in bold, while borderline significant results (p < 0.1) were marked in italics. ERM = epiretinal membrane; ηρ2 = partial eta-squared; pRNFL = peripapillary retinal nerve fiber layer thickness; tRNFL = temporal quadrant of pRNFL; GCIPL = composite macular ganglion cell and inner plexiform layer thickness; INL = inner nuclear layer thickness; ONL = outer nuclear layer thickness; RPE = retinal pigment epithelium; LA = letter acuity; EDSS = Expanded Disability Status Scale; WST = walking speed test; MDT = manual dexterity test; PST = processing speed test; sNfL = serum neurofilament light chain.
Analyzed with generalized linear model.
Analyzed with linear mixed-effect model.
In the ERM+ group, higher ERM stages were associated with greater global disability (higher EDSS, WST, and MDT scores). However, PST and sNfL age-adjusted Z-scores did not differ by ERM severity (Figure 3). Visual acuity was comparable between Stage 0 and Stage 1 ERMs, whereas eyes with at least Stage 2 ERMs demonstrated worse visual acuity than either Stage 0 or Stage 1 (Figure 3). Eyes with Stage 1 ERMs had lower INL, ONL, and RPE thicknesses as compared to Stage 0 ERMs. However, no differences were observed in any of the retinal layer thicknesses between eyes with Stage 1 and those with at least Stage 2 ERMs (Supplemental Figure 2).
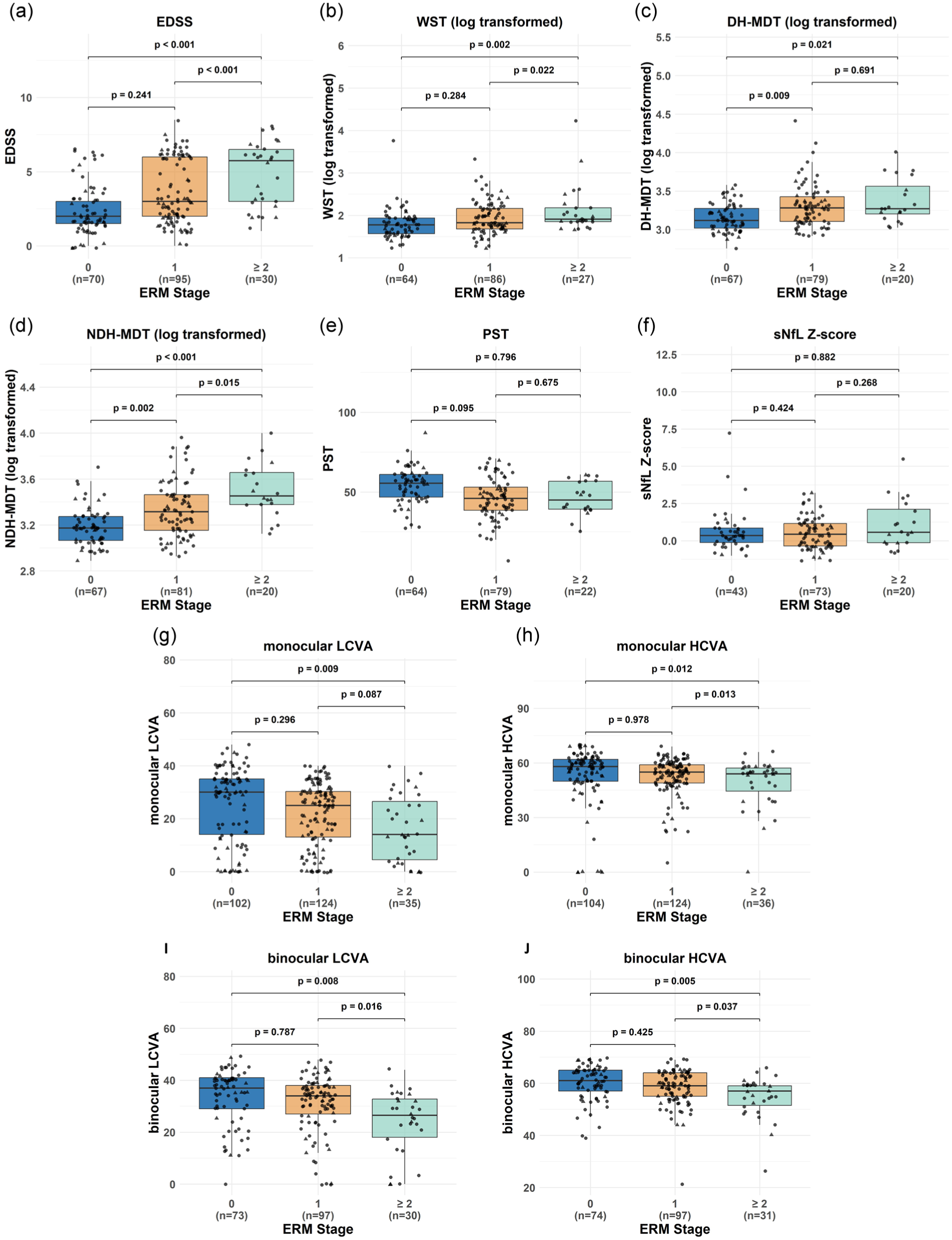
Comparisons of disability measures by ERM stages.
Discussion
In this cross-sectional study, we observed a higher ERM prevalence in PwMS without other identifiable etiologies for ERMs, relative to HCs, suggesting that MS might contribute to secondary ERM formation. PwMS exhibiting ERMs demonstrated greater global disability, including impaired motor function, reduced cognitive function, diminished visual acuity, and elevated sNfL levels, as compared to those without ERMs. Furthermore, GCIPL, INL, ONL, and RPE thicknesses were lower in the eyes of PwMS with ERMs, independent of prior ON history. Higher ERM severity correlated with greater disability, poorer visual outcomes, and lower retinal layer thicknesses, independent of age. These findings indicate that ERMs in PwMS may serve as a potential phenotypic marker of increased disease severity, although the underlying mechanisms remain to be determined.
The higher ERM prevalence in this MS cohort, as compared to HCs, and exceeding prevalence rates reported in older populations from large epidemiological studies, 27 suggests that MS pathophysiology might directly contribute to ERM formation. This is particularly striking given that age is a major contributing factor to idiopathic ERM formation, yet this younger MS cohort exhibited a higher prevalence of ERMs. These observations suggest that the chronic inflammatory and neurodegenerative processes operative in MS may contribute to ERM development.
The blood–retinal barrier (BRB) relies on endothelial tight junctions to maintain homeostasis. Our findings might support that BRB disruption in MS compromises tight junction integrity. One possible explanation could be that retinal inflammation may release cellular distress signals, disrupting the BRB and potentially activating Müller glia. 38 This might explain why ERMs correlate with greater disability in PwMS. Furthermore, our results show reductions in INL and ONL thicknesses in eyes with ERMs. One possible mechanism is the disruption of the glial support in the retina, prompting displaced Müller cells to form scaffolding processes that support the migration of other retinal cells. 39
As expected, PwMS with ERMs demonstrated worse visual acuity as compared to the ERM− group. ERMs may inherently cause visual impairment directly through mechanical traction and structural distortion of the macula, independent of MS-related optic neuropathy. Therefore, the reduced visual acuity observed in the ERM+ group may, at least in part, represent a direct localized consequence of the ERMs. However, the robust associations between ERMs and numerous global MS disability and sNfL measures, combined with the higher prevalence of ERMs observed in PwMS, raise the possibility that ERMs may represent more than localized pathology, potentially indicating widespread glial activation throughout the CNS. In MS, microgliosis and macrogliosis within the brain and spinal cord contribute to neuroinflammation and neurodegeneration.3,7,40 These glial responses involve similar molecular mechanisms as those observed in ERM formation, including cellular migration and proliferation. 24 ERMs might, therefore, reflect the propensity for exaggerated glial reactivity throughout the CNS. This is supported by our finding of more severe global disability and elevated sNfL levels in PwMS with ERMs. The observation that PwMS demonstrating ERMs exhibited higher sNfL levels suggests a possible link between localized retinal pathology and more generalized neuroaxonal injury. One possible explanation could be that ERMs reflect a structural endpoint of chronic retinal inflammation, primarily mediated by microgliosis and/or astrogliosis. These glial responses could occur in parallel with CNS inflammatory processes that contribute to axonal damage. Just as reactive gliosis in brain lesions can transition from a protective response to a maladaptive one that hinders repair, the glial reactions underlying ERMs in PwMS might follow similar pathological trajectories. 8 Therefore, given both the increased prevalence of ERMs and their strong associations with global disability, ERMs in MS might represent a phenotypic manifestation of dysregulated glial responses that are driving MS pathogenesis.
Müller glial cells can migrate across retinal layers in response to injury, potentially initiating cellular differentiation and neurogenesis.22,23 Therefore, our findings may reflect this migratory response, possibly driven by upregulation of proteins involved in intracellular trafficking, such as glial fibrillary acidic proteins (GFAPs). This raises the question of whether the observed ERMs could instead represent protective migratory cells involved in promoting retinal regeneration, maintaining retinal homeostasis, and digesting cellular debris, among other functions.
We also observed greater GCIPL and RPE, in addition to INL and ONL, thickness reductions in the eyes of PwMS with ERMs. GCIPL thinning suggests a potential link between ERMs and accelerated neurodegenerative processes. ERM-related mechanical traction, chronic inflammation, or glial activation may impose additional retinal stress, exacerbating neuronal loss. RPE thinning might reflect the same fibrogenic and inflammatory mechanisms implicated in idiopathic ERM formation. 41 Interestingly, despite higher numbers of ON eyes in the ERM+ versus ERM− group (92 vs. 67), rather than the expected temporal quadrant predominant pRNFL thinning following ON, 42 our findings demonstrated the opposite pattern. This counterintuitive finding can likely be attributed to the mechanical effects of ERM-related traction causing apparent tRNFL thickening through structural distortion.
The strengths of this study include rigorous matching of PwMS with and without ERMs, minimizing potential confounding factors that could contribute to ERM formation. In addition, we included a broad range of disability measures to evaluate widespread MS disease burden. However, our study has several limitations. First, the lack of GFAP levels limited our ability to explore the potential contribution of astrogliosis to ERM development. Having GFAP data would have allowed us to probe glial activation and reactivity, which may play a crucial role in ERM formation in the context of MS-related inflammation. Second, early ERMs may be dynamic in nature, and their transient appearance could result in under-detection, potentially leading to an underestimation of their true prevalence in MS. On the contrary, since previous studies on ERM prevalence in asymptomatic subjects among the general population are not designed to detect subtle ERMs, our inclusion of minimally visible ERMs (Stage 0) may have contributed to overestimation. Nevertheless, these opposing factors may have partially offset each other. Third, an inherent challenge in this study is that ERMs can disrupt the inner retinal layers, leading to segmentation algorithm failures. In this study, 32 peripapillary and 86 macular scans were excluded due to failing quality control criteria. This relatively high exclusion rate might have disproportionately removed severe ERM cases from our analyses, potentially introducing a selection bias that may have influenced the observed results. Fourth, the cross-sectional design of this study limits our ability to establish a temporal relationship between ERMs and MS disease pathology. Finally, considering that glial pathology is thought to be more relevant in progressive MS, one could hypothesize that ERMs may be more prevalent in progressive forms than in relapsing forms. However, the proportion of PwMS with progressive disease in this cohort was low, precluding any meaningful investigation of this hypothesis.
To conclude, our findings suggest ERMs are more common among PwMS than previously thought, raising the possibility that MS could be a secondary cause of ERMs. Along these lines, ERMs in PwMS appear to be a marker of an MS phenotype characterized by greater retinal neurodegeneration, but more relevantly higher levels of global neurological disability, including greater motor, cognitive, and visual impairment. Collectively, our findings raise the possibility that ERMs in PwMS may reflect broader disease processes extending beyond the retina, possibly linked to global CNS neuroinflammation. Further longitudinal studies are needed to validate whether ERMs can serve as meaningful biomarkers of disease severity in MS. In particular, evaluating patients at onset or nadir following acute ON episodes could help determine whether active inflammation directly drives secondary ERM formation. Such insights could enhance our understanding of MS pathophysiology and offer a foundation for more personalized monitoring and treatment strategies.
Supplemental Material
sj-docx-1-msj-10.1177_13524585261446834 – Supplemental material for Association of epiretinal membranes with disability in people with multiple sclerosis
Supplemental material, sj-docx-1-msj-10.1177_13524585261446834 for Association of epiretinal membranes with disability in people with multiple sclerosis by Ting-Yi Lin, Anna Bacchetti, Brenna McCormack, Hussein Moussa, Jonathan D Krett, Omar Ezzedin, Gabriel Otero-Duran, Nicole Pellegrini, Madeline Inserra, Scott D Newsome, Bardia Nourbakhsh, Kathryn C Fitzgerald, Elias S Sotirchos, Peter A Calabresi, Amir H Kashani and Shiv Saidha in Multiple Sclerosis Journal
Supplemental Material
sj-docx-2-msj-10.1177_13524585261446834 – Supplemental material for Association of epiretinal membranes with disability in people with multiple sclerosis
Supplemental material, sj-docx-2-msj-10.1177_13524585261446834 for Association of epiretinal membranes with disability in people with multiple sclerosis by Ting-Yi Lin, Anna Bacchetti, Brenna McCormack, Hussein Moussa, Jonathan D Krett, Omar Ezzedin, Gabriel Otero-Duran, Nicole Pellegrini, Madeline Inserra, Scott D Newsome, Bardia Nourbakhsh, Kathryn C Fitzgerald, Elias S Sotirchos, Peter A Calabresi, Amir H Kashani and Shiv Saidha in Multiple Sclerosis Journal
Supplemental Material
sj-docx-3-msj-10.1177_13524585261446834 – Supplemental material for Association of epiretinal membranes with disability in people with multiple sclerosis
Supplemental material, sj-docx-3-msj-10.1177_13524585261446834 for Association of epiretinal membranes with disability in people with multiple sclerosis by Ting-Yi Lin, Anna Bacchetti, Brenna McCormack, Hussein Moussa, Jonathan D Krett, Omar Ezzedin, Gabriel Otero-Duran, Nicole Pellegrini, Madeline Inserra, Scott D Newsome, Bardia Nourbakhsh, Kathryn C Fitzgerald, Elias S Sotirchos, Peter A Calabresi, Amir H Kashani and Shiv Saidha in Multiple Sclerosis Journal
Supplemental Material
sj-tiff-1-msj-10.1177_13524585261446834 – Supplemental material for Association of epiretinal membranes with disability in people with multiple sclerosis
Supplemental material, sj-tiff-1-msj-10.1177_13524585261446834 for Association of epiretinal membranes with disability in people with multiple sclerosis by Ting-Yi Lin, Anna Bacchetti, Brenna McCormack, Hussein Moussa, Jonathan D Krett, Omar Ezzedin, Gabriel Otero-Duran, Nicole Pellegrini, Madeline Inserra, Scott D Newsome, Bardia Nourbakhsh, Kathryn C Fitzgerald, Elias S Sotirchos, Peter A Calabresi, Amir H Kashani and Shiv Saidha in Multiple Sclerosis Journal
Supplemental Material
sj-tiff-2-msj-10.1177_13524585261446834 – Supplemental material for Association of epiretinal membranes with disability in people with multiple sclerosis
Supplemental material, sj-tiff-2-msj-10.1177_13524585261446834 for Association of epiretinal membranes with disability in people with multiple sclerosis by Ting-Yi Lin, Anna Bacchetti, Brenna McCormack, Hussein Moussa, Jonathan D Krett, Omar Ezzedin, Gabriel Otero-Duran, Nicole Pellegrini, Madeline Inserra, Scott D Newsome, Bardia Nourbakhsh, Kathryn C Fitzgerald, Elias S Sotirchos, Peter A Calabresi, Amir H Kashani and Shiv Saidha in Multiple Sclerosis Journal
Footnotes
Author Contributions
S.S., P.A.C., and T.-Y.L. conceptualized the study. T.-Y.L., A.B., B.M., H.M., and O.E. performed OCT quality control and segmentation. N.P. took part in the project administration. T.-Y.L., A.B., B.M., G.O.-D., N.P., and M.I. contributed to clinical and OCT data acquisition. S.S., P.A.C., J.D.K., S.D.N., B.N., and E.S.S. provided clinical data. T.-Y.L. analyzed the data, conducted the analyses, and wrote the original draft. S.S., T.-Y.L., K.C.F., E.S.S., P.A.C., and A.H.K. contributed to data interpretation. All authors reviewed and edited the final manuscript. S.S., P.A.C., and T.-Y.L. had access to raw data and final responsibility for the decision to submit for publication.
Declaration of Conflicting Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: T.-Y.L. reports no relevant disclosures. A.B. reports no relevant disclosures. B.M. reports no relevant disclosures. H.M. reports no relevant disclosures. J.D.K. reports no relevant disclosures. O.E. reports no relevant disclosures. G.O.-D. reports no relevant disclosures. N.P. reports no relevant disclosures. M.I. reports no relevant disclosures. S.D.N. has received consulting fees for scientific advisory boards from Biogen, Genentech, Inc., Bristol Myers Squibb, Novartis, and TG Therapeutics; study lead PI for a Roche clinical trial program; received research funding (paid directly to institution) from Biogen, Lundbeck, Roche, Genentech, Inc., Sanofi, National MS Society, The Stiff Person Syndrome Research Foundation, Department of Defense, and Patient-Centered Outcomes Research Institute. B.N. has been funded by the National Multiple Sclerosis Society, the National Institutes of Health, the US Department of Defense, PCORI, Genentech, and Axsome Therapeutics. He has received consulting honoraria from TG Therapeutics and Alkermes. K.C.F. reports no relevant disclosures. E.S.S. has consulted for Alexion, Amgen, TG Therapeutics, and Roche/Genentech, is the site principal investigator for studies funded by Alexion, Roche/Genentech, UCB, and Ad Scientiam, and is principal investigator for an investigator-initiated study funded by Astoria Biologica. P.A.C. has received consulting fees from Lilly, Idorsia, and Novartis and is PI on grants to Johns Hopkins from Genentech and the Myelin Repair Foundation. A.H.K. has received Grants from Carl Zeiss Meditec, Regenxbio, Aspen Biosciences, and Alcon and has consulted for Carl Zeiss Meditec. S.S. has received consulting fees from Medical Logix for the development of CME programs in neurology and has served on scientific advisory boards for Biogen, EMD Serono, Novartis, Genentech Corporation, Amgen, Horizon Therapeutics, Clene Pharmaceuticals, Sanofi and ReWind Therapeutics. He has performed consulting for Novartis, Genentech Corporation, JuneBrain LLC, Innocare Pharma, Kiniksa Pharmaceuticals, Lapix Therapeutics, and Setpoint Medical. He has received equity compensation from JuneBrain LLC and Lapix Therapeutics for consulting. He is the PI of investigator-initiated studies funded by Genentech Corporation, Biogen, TG Therapeutics, and Novartis. He previously received support from the Race to Erase MS foundation. He was also the site investigator of trials sponsored by MedDay Pharmaceuticals and Clene Pharmaceuticals, and is the site investigator of trials sponsored by Novartis, as well as Lapix Therapeutics.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by National Institute of Health (R01NS082347 to S.S. and P.A.C.; K23NS117883 to E.S.S.), National Multiple Sclerosis Society (RG-1606-08768 and RG-1907-34405 to S.S.; RG-1904-33834 to E.S.S.), and Boone Pickens Professorship in Ophthalmology (A.H.K.). The sponsor or funding organization had no role in the design or conduct of this research.
ORCID iDs
Data Availability Satement
Written requests for access to the data reported in this paper will be considered by the corresponding author and a decision made about the appropriateness of the use of the data. If deemed appropriate, a data-sharing agreement will be established before providing access to a fully de-identified version of the data set. The corresponding author had full access to all the data in the study and was responsible for the final decision to submit the manuscript for publication.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.