
Abstract
Background
Insulin-like growth factor-1 (IGF-1) promotes neurogenesis, cell survival, and glial function, making it a promising candidate therapy in Alzheimer's disease (AD).
Objective
Long arginine 3-IGF-1 (LR3-IGF-1) is a potent IGF-1 analogue. We sought to determine whether intranasal (IN) LR3 treatment would delay cognitive decline and pathology in 5XFAD mice.
Methods
Wildtype and 5XFAD male mice were treated for 7 months (3–10 months of age), with IN LR3-IGF-1 or IN Vehicle (Veh) (n = 19–27 mice/group). Behavior, memory, and brain imaging were assessed at 8–9 months of age and tissues collected at 10 months. A comprehensive amyloid-β (Aβ) profile and other pathologic features were conducted and supportive in vitro stimulation studies in BV-2 microglial cells were also performed.
Results
In male 5XFAD mice, IN LR3-IGF-1 treatment improved body composition, but did not significantly alter cognitive symptoms, as assessed by multiple assays. In cortex, LR3 treatment improved some facets of pathology, including a reduction in filamentous plaques, and increase in inert plaques, corresponding with a reduction in low molecular weight Aβ oligomers. In vitro, uptake of Aβ1–42 peptide by BV2 cells was enhanced by LR3-IGF-1, which was also found to promote gene pathways implicated in actin remodeling and endocytosis.
Conclusions
LR3 promotes favorable effects on Aβ plaque remodeling in cortex of male 5XFAD mice but fails to preserve aspects of behavior or memory. While these data do not support LR3 as a monotherapy per se, they do warrant further investigation into its potential for combinatorial formulations aimed at targeting the complexity of AD.
Introduction
The incidence of late-onset Alzheimer's disease (AD) roughly doubles every five years in adults aged 65 years and older. 1 Traditional pharmacologic approaches to AD therapy, such as cholinesterase inhibition and N-methyl-D-aspartate (NMDA) antagonism, offer modest temporary improvements in cognitive function but do not prevent neurodegeneration. 2 More recently, anti-amyloid-β (Aβ) monoclonal antibody therapies targeting a suite of different epitopes and Aβ species have variably been shown to reduce brain Aβ levels—even proteotoxic aggregates in the case of newer monoclonal antibodies (mAbs)—but have shown uncertain efficacy in slowing cognitive decline in moderate to severe AD. 3
A significant challenge in implementing promising strategies for AD remains effectively targeting delivery to the brain. To this end, intranasal (IN) administration has been leveraged as an approach to more effectively direct agents to the central nervous system (CNS) while also minimizing peripheral exposure. Indeed, a great deal of interest has centered around the promise of IN insulin to confer cognitive benefits based on animal models and small pilot studies, which led to a large clinical trial in mild cognitive impairment (MCI) and AD (Study of Nasal Insulin in the Fight Against Forgetfulness; SNIFF; NCT01767909). Unfortunately, despite early promise, this study failed to meet its primary endpoints, which may have been in part attributed to the device used. 4 This raises the possibility that early presymptomatic iterations of the SNIFF trial incorporating new formulations and alternatives to insulin may be needed. To this end, we were intrigued with the therapeutic potential of a related, but distinct ligand, IGF-1, as an IN candidate for AD therapy.
Interest in the therapeutic potential of peripheral IGF-1 administration in AD dates back nearly two decades, with studies producing conflicting outcomes.5–9 Interestingly, indirectly augmenting brain IGF-1 signaling via subcutaneous administration of growth hormone releasing hormone was found to improve executive function and verbal memory in MCI patients, 10 but elevating serum growth factors (i.e., growth hormone (GH) and IGF-1) is not a viable strategy due to many well-documented side effects, including risk for malignancy. 11
The role of IGF-1 in brain aging and neurodegenerative diseases is complex and controversial, and this topic has been extensively reviewed elsewhere. 12 Mouse models of reduced GH/IGF-1 activity are typically long-lived,13,14 yet lower circulating IGF-1 levels in the GH/IGF-1-deficient Ames mouse are contrasted with elevated hippocampal expression of IGF-1 in this model, without evidence of cognitive decline. 15 Moreover, dwarf mice are typically characterized by reduced inflammation and improved insulin sensitivity, which are both linked to better cognitive and overall health. On the other hand, isolated IGF-1 deficiency leads to impairments in neurovascular coupling,16,17 adaptive cerebral arterial remodeling, 18 mitochondrial function in astrocytes, and visuospatial memory. 19 In contrast, central administration of IGF-1 can unambiguously produce favorable effects on features of normal brain aging in rodents,20–23 and we have previously demonstrated beneficial effects on cognition conferred by elevating brain IGF-1 levels in aged mice via transgenic central igf1 overexpression or IN IGF-1 treatment. 24
IN IGF-1 has been shown to bypass the blood-brain barrier to activate brain IGF-1 receptor signaling without the need to directly expose the peripheral circulation to pharmacologic doses of IGF-1. 25 This approach has previously been shown to be protective in murine models of Huntington's disease, 26 ischemic and hemorrhagic stroke,27–29 and neuroinflammation,30,31 yet no attempts to intranasally stimulate brain IGF-1 signaling as long-term prophylaxis against cognitive decline or neurodegeneration in AD models have been described. Thus, we aimed to test the hypothesis that intermittent stimulation of the CNS with a potent IGF-1 analogue (Long-R3-IGF-1) via IN delivery32,33 in a mouse model of cerebral amyloidosis could be harnessed to elicit therapeutic benefits while minimizing potential risks to the periphery.
Methods
Animals
A 5XFAD mouse colony was established by mating male 5XFAD mice (MMRRC #034848) with female C57BL/6J mice (Jax stock#000664). Male wild-type and 5XFAD mice, respectively, were group-housed (4–5 animals per cage) and randomly assigned to treatment or control groups. An overview of the experimental design is shown in Figure 1A. Mice were housed under a standard photoperiod (14:10 light/dark cycle) at 22°C and provided standard rodent chow (Purina 5001) and water ad libitum. All experiments were approved by the Einstein Institutional Animal Care and Use Committee (IACUC) and cared for according to IACUC standards.
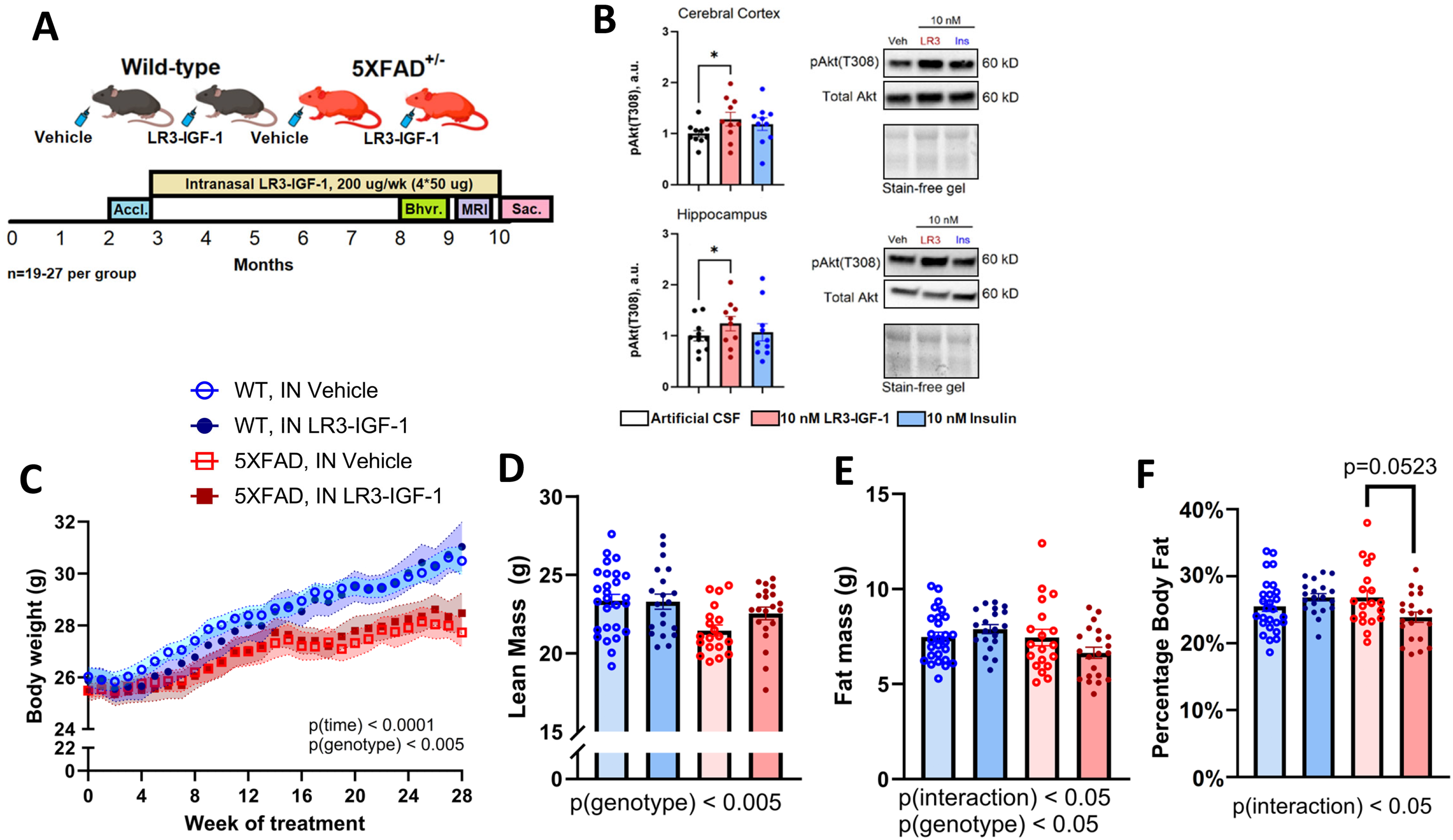
Experimental design, ex vivo LR3-IGF-1 stimulation, and body composition with long-term LR3-IGF-1 treatment. (A) Experimental design for longitudinal intranasal (IN) treatments. Male C57BL/6J WT or 5XFAD mice were acclimated at 10 weeks of age, then treated intranasally with Vehicle or LR3-IGF-1 (4 × 50 µg weekly) from 3 months to 10 months of age. Behavior and memory were assessed from 8–9 months of age, and MRI imaging was acquired at 9 months of age (made in Biorender). (B) Ex vivo stimulation of cerebral cortex and hippocampus of wild-type 8-month-old male C57BL/6J mice (n = 10) with aCSF (vehicle), 10 nM LR3-IGF-1, or 10 nM insulin for 2 min. Phospho-Akt(T308) expression increased in both cortex and hippocampus treated with 10 nM LR3-IGF-1 relative to control. Image of the strain-free gel is included to confirm equal loading. (C) Body weightchange over the study duration. Mean weight change from 3–10 months was +3.9 g (WT-Veh), + 4.6 g (WT-LR3), + 2.4 g (5X-Veh), and +2.6 g (5X-LR3), with significant weight change over time (p < 0.0001) modified by genotype (p < 0.005). (D-F) Lean mass (p < 0.005) and fat mass (p < 0.05) were modified in 5XFAD mice compared with WT, with a trend toward reduced percentage body fat (fat mass / total body weight) in IN LR3-IGF-1 treated 5XFAD mice compared with those treated with IN Vehicle (p = 0.0523). Data are shown as mean ± S.E.M. *p < 0.05.
Intranasal treatments
In the longitudinal IN study, male WT and 5XFAD mice were assigned to receive either IN saline vehicle (WT-Veh, 5X-Veh) or IN Long R3-IGF-1 (IN LR3-IGF-1; Repligen, LONG®R3 IGF-I, 10-1010-5) (WT-LR3, 5X-LR3). Stock LR3 was adjusted from pH 3 to 6 immediately prior to IN administration, which we confirmed had no effect on pAkt activation dynamics in BV2 cells (Supplemental Figure 1). Animals were acclimated to IN treatment for 3 weeks, as described. 34 Beginning at 3 months of age, mice were treated four times per week with 4 × 5 µL IN Vehicle or 4 × 5 µL IN LR3-IGF-1, alternating between each naris (2.5 µg/µL; 50 µg total dose per treatment) until the mice were 10 months of age, and sacrificed 24 h after their final treatment for tissue collection.
Body weight and body composition
For longitudinal IN studies, body weight was recorded every two weeks and body composition for total fat and lean mass at 8 months of age via EchoMRI body composition analyzer (EchoMRI; Houston, TX).
Assessment of behavior and cognition
At 8 and 9 months of age, mice were evaluated by a series of behavioral and cognitive assessments. For all tests, the operator was blinded to group assignment by a collaborator, who de-blinded the operator only after all data were collected. To evaluate visuospatial memory, the Y-Maze and Box Maze tests were used as described.35,36 In the Y-Maze test, mice were placed in a maze with three identical arms and permitted to roam for 5 min. Each entry into a new arm was recorded, and an alternation was defined as sequential entry into three distinct arms without repetition. Percent alternation was calculated as [total alternations/(total entries – 2)]*100%. In the box maze test, mice were placed in a well-lit arena with eight possible exits, only one of which allows for escape to a dark enclosure which served as a safe harbor. 35 Each mouse was subjected to one training period and four consecutive trials with 2 min intervals between training and each trial, with a maximum of 10 min to locate the true escape route per trial. Latency to escape the main arena to the dark enclosure was measured, as was the number of false escapes (defined as entry of the mouse into a false exit).
To evaluate anxiety-like behavior and locomotor activity, the Zero Maze and Open Field tests, respectively, were used, while depressive-like behavior was determined via forced swim test. 37 The zero maze comprises an elevated, ring-shaped platform, half of which is walled off on both sides (closed arms), the other half lacking walls (open arms). For testing, mice are placed in one of the closed arms and allowed to roam freely for minutes, and the percentage of the elapsed time spent in the open arm was recorded. For the open field test, mice were placed in the corner of an enclosed square arena and allowed to roam for 9 min. A smaller, central square was defined as the central zone of the field. An aerial camera and companion software (Bioseb, Inc) were used to track the movements of each mouse, including average velocity, track length, activity level, and time spent in the central zone. In the forced swim test, the mouse was transferred to a large glass beaker containing clean tap water at 25°C. The mouse was gently placed in the water and given 2 min of lag time prior to the beginning of the assay to allow for acclimation. Mouse movement patterns were then closely observed for 4 min to determine the percentage immobility. Behaviors classified as Mobile include climbing or moving against the walls of the cylinder, active swimming with or without propulsion through the water, and vertical motions (e.g., diving). Behavior classified as Immobile and thus indicating depressive-like tendencies include intermittent treading water in place or idle floating. 38 Finally, to determine whether IN treatments impacted olfaction, mice were acclimated to a vanilla-scented wafer cookie over two days, then fasted overnight and placed in a clean cage with 1 cm bedding. 39 The wafer cookie was buried at a random location in the cage, and the time to obtain the buried wafer was recorded.
Magnetic resonance imaging (MRI)
All MRI data were acquired in a 9.4T Varian Direct Drive system (Agilent Technologies, Santa Clara, CA, USA). Mice were anesthetized with 1.25% for acquisition of diffusion tensor imaging (DTI) 40 which included fractional anisotropy (FA) and related readouts. FA is derived from DTI, which maps the diffusion of water molecules in the brain and is sensitive to microstructural changes in the brain that correlate with cognitive decline. Axial diffusivity was also derived from DTI and is considered a surrogate marker of axonal damage. Cerebral blood flow images were acquired using arterial spin labeling methods. 41 Brain water longitudinal relaxation time (T1) was estimated using inversion recovery with echoplanar readout. Volumetric analysis was performed by registering the Dorr mouse brain atlas 42 to T2-weighted FSE images. T2* is sensitive to susceptibility differences, it correlates with iron concentration in tissue. T2* is also influenced by myelin, calcium concentration and blood. The shortening of T2* is a good surrogate for increased iron content. These analyses were accomplished with a pipeline specifically designed for this project.
Ex vivo brain parenchyma stimulation
Eight-month-old male C57BL/6 mice (n = 10) were fasted for 3 h, then anesthetized with 2% isoflurane gas and sacrificed for brain collection. The prefrontal cortex and hippocampus were freshly dissected, methodically minced with a sharp scalpel into ∼1 mm cubed pieces, and immediately transferred to artificial cerebrospinal fluid (aCSF, Harvard Apparatus, 59-7316) at 37°C in 8-well plates. Tissues were equilibrated for 10 min and then treated with aCSF vehicle, 10 nM LR3-IGF-1, or 10 nM insulin (Sigma) for 2 min at 37°C in 5% CO2, as previously described. 43 Tissues were then immediately washed and homogenized with ice-cold RIPA buffer for protein extraction and downstream applications.
Autophagic flux
Freshly collected prefrontal cortex was dissected, minced, and transferred to DMEM supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin warmed to 37°C. Minced tissue was incubated for 2 h at 37°C in 5% CO2 with or without autophagy inhibitors (NH4Cl [20 mM]; leupeptin [200 μM]). The ratio of p62 expression plus/minus inhibitors was determined for each animal, and p62 flux was normalized against mean flux in wild-type IN vehicle-treated mice as described. 44
Immunohistochemistry
Saline-perfused brains were hemisected along the sagittal midline and the right hemisphere was transferred to 4% paraformaldehyde and fixed, cryoprotected via 30% sucrose, embedded in M-1 embedding matrix (Epredia, #1310), and stored at −80°C. Free-floating 30 µm sagittal sections were subjected to standard immunofluorescence or diaminobenzidine (DAB) staining techniques. DAB sections were digitized with P250 high-capacity slide scanner for analysis in Panoramic Caseviewer software. Immunofluorescence images were obtained with a Leica SP8 confocal microscope. Cell counts, plaque analysis, morphological assessments, and fluorescent intensity analysis were performed in Fiji/ImageJ. For triple staining, antibodies targeting microglia (Iba1) and Aβ (D54D2) were combined with methoxy-X04 (MeX04) staining, and cover-slipped. For BV-2 cell immunocytochemistry, coverslip-grown cells were fixed, permeabilized with 0.1% Triton X-100, incubated with appropriate antibodies, inverted onto glass slides, and mounted.
Amyloid plaque morphological analysis
Plaque size distribution in cerebral cortex and hippocampus were determined in 4 × 500 × 500 µm fields (3–4 sections used per animal) with Caseviewer and exported to ImageJ for analysis. After defining the pixel/µm ratio, plaques were manually encircled to obtain both plaque count and area, from which plaque diameter was derived. Per section averages were obtained by taking the mean field value of plaque count and size and sections averaged to obtain biological replicates. For size-distribution analysis, plaques >20 µm were considered, and split into 20–40, 41–60, 61–80, and >80 µm bins.
To evaluate plaque morphology, 2–3 fields per region of interest (cerebral cortex, hippocampus) per section (2–3 per animal) were randomly selected and imaged with a Leica SP8 confocal microscope to detect MeX04, Iba1, and Aβ (D54D2) fluorescence, respectively. Histologic classification of plaques was performed on two-channel fields (D54D2, MeX04) after blinding the evaluator to treatment group. The presence of a dense-core deposit staining positive for MeX04 defined a plaque as either Compact or Inert; if a halo of D54D2 + plaque was present, the plaque was classified as Compact, whereas isolated MeX04 + cores were classified as Inert. D54D2 + plaques without a dense-core deposit were classified as Filamentous. Thus, plaques were categorized as Filamentous (D54D2 high, MeX04 low/null), Compact (D54D2 high, MeX04 high), or Inert (D54D2 low/null, MeX04 high), as previously described.45,46 The colocalized fraction of Iba1 + microglia to D54D2 + plaques was determined by dividing the number of microglia in direct contact with plaques by the total number of microglia per field.
BV-2 cell culture and Aβ1-42 uptake
To evaluate uptake of Aβ1-42 peptide, BV-2 cells were seeded in poly-D-lysine (PDL) (Alkali Scientific) chamber slides were pre-treated for 3 h with 50 nM LR3-IGF-1 or vehicle in the presence or absence of 50 µM LY294002 (CST, 9901S). Cells were then treated with 200 nM HiLyte Fluor™ 488 Aβ1-42 (AnaSpec, AS-60479-01) for 1 h, processed, and visualized at 20× magnification, and the percentage fluorescent area and number of Aβ-positive puncta per square millimeter were calculated In Fiji/ImageJ.
RNA extraction and gene expression
RNA was isolated from tissues and cells using TRIzol® Reagent (TRIzol, Life Technologies), cDNA synthesized (Bio-Rad iScript cDNA synthesis kit), and RT-qPCR reactions performed with Applied Biosystems PowerTrack SYBR Green Master Mix on a Bio-Rad CFX384 RT-qPCR thermocycler in triplicate. The delta-delta Ct (2–ΔΔCt) method was used to determine relative gene expression normalized to cyclophilin A (ppia). Primer sequences are available in Supplemental Material. For bulk RNAseq, RNA was isolated from BV-2 cells treated with 50 nM LR3-IGF-1, 50 nM Insulin or vehicle control, in the presence or absence of 1 µM oligomeric human Aβ1-42 peptide (Sigma Aldrich, PP69) (n = 4 per group) for 4 or 24 h. Only the analysis of the LR3-IGF-1 versus LR3-IGF-1+ Aβ1-42 peptide comparison at 4 h is presented here, but all transcriptomic data are deposited in SRA for open access. RNA quality control, library preparation and sequencing were performed by Novogene. Groupwise analysis of differentially expressed genes were performed with DESeeq2 and an FDR < 0.05. GSEA analysis was conducted in R with the fgsea package (v1.20.0) using KEGG and Gene Ontology databases.
Amyloid ELISA
To quantify levels of Aβ40 and Aβ42 levels in plasma and tissues, Invitrogen Human Aβ40 (KHB3481) and Aβ42 (KHB3441) ELISA Kits were used according to the manufacturer instructions. To extract soluble and insoluble forms of Aβ, a protocol modified from Huang et al. was used. 47 Amyloid concentrations were reported as ng/mg total protein.
Protein extraction and western blotting
Protein expression in tissues or cells were determined via western blotting, as described previously. 24 Representative blots are included in main figures and whole uncropped blots are found in the Supplemental Material. Following overnight incubation with primary antibody, membranes were washed and incubated for 1 h in goat anti-rabbit IgG HRP-linked secondary antibody (1:5000, #7074), exposed to Clarity Western enhanced chemiluminescence (ECL) substrate (Bio-Rad, Hercules, CA) and visualized with a Bio-Rad Chemidoc MP imager to first-pixel saturation. Densitometric analysis was performed with Image Lab software (Bio-Rad, Hercules, CA).
Statistics
Cross-sectional data were analyzed using one-way or two-way ANOVA (genotype×treatment), while longitudinal measures were assessed with two-way ANOVA with repeated measures on time. Planned contrasts were performed as appropriate. When a significant main effect was detected, planned two-group contrasts with Šidák correction were applied to the following comparisons: WT-Veh versus WT-LR3, WT-Veh versus 5X-Veh, and 5X-Veh versus 5X-LR3. Normality of distribution was routinely assessed via the Shipiro-Wilk procedure. When data were not normally distributed, they were log-transformed to ensure normality, which was verified using Q-Q plots. For two-group comparisons, unpaired t-tests were run when the normality criterion was met, and Welch's test was used for unequal variances. All analyses were performed using Prism software. Values are reported as means ± standard error (SE). A p ≤ 0.05 was considered statistically significant.
Results
Pharmacodynamic profile of LR3-IGF-1
We first confirmed the ability of LR3-IGF-1 peptide to stimulate canonical IGF-1R signaling ex vivo (Figure 1B). Indeed, stimulation of cerebral cortex and hippocampus tissue from WT 8-month-old male mice with 10 nM LR3-IGF-1 was confirmed to augment pAkt(T308) levels in excised tissues versus vehicle or insulin-stimulated conditions (Figure 1B).
Phenotypic effects of intranasal LR3-IGF-1 treatment in 5XFAD and Wt mice
Weight gain among groups was similar during the first three months of treatment (∼6 months of age), after which a plateau in weight gain was observed in 5XFAD mice compared with WT (Figure 1C, n = 19–27 per group, p(genotype) < 0.005). Moreover, at 8 months of age, a significant genotype effect was observed for lean mass (Figure 1D; p < 0.005) and a significant genotype×treatment interaction (Figure 1E; p < 0.05) was observed for fat mass, but no significant differences were observed among groups. However, 5X-LR3 treated mice tended to have a reduction in percent body fat as compared to 5X-Veh mice (Figure 1F; p = 0.0523).
Effect of LR3-IGF-1 treatment on signaling and gene expression in vivo
We next assessed effects of long-term LR3-IGF-1 treatment on canonical signaling pathways in cerebral cortex. Longitudinal IN LR3-IGF-1 did not suppress IGF-1R expression, nor were there any significant differences in IGF-1R levels between WT and 5XFAD mice (Figure 2A). Furthermore, autophagic flux in prefrontal cortex was not altered with IN LR3-IGF-1 treatment (Supplemental Figure 2). Immunoblotting against a variety of markers of canonical IGF-1R signaling after 7 months of treatment revealed no difference in p-p70S6K(T389) (Figure 2B), but Akt(Thr308) phosphorylation was numerically increased in 5X-LR3 mice compared with WT-LR3, and an overall genotype×treatment interaction effect was also observed (Figure 2C, p < 0.05). Meanwhile, phosphorylation of GSK3β(Ser9), and ERK1/2(Thr202/Tyr204) were not significantly different among groups (Figure 2D,E), but phospho-4E-BP1(Thr37/46) levels were elevated in 5XFAD mice relative to WT, irrespective of treatment (Figure 2F, p < 0.05). Moreover, a four-fold increase in igf1 gene expression was observed in cerebral cortex of 5XFAD mice compared with WT (Supplemental Figure 3A, p < 0.0001) as were expression of trem2, iba1, il1b, tnfa, and il6 transcripts, but no significant genotype×treatment effects were observed (Supplemental Figure 3).
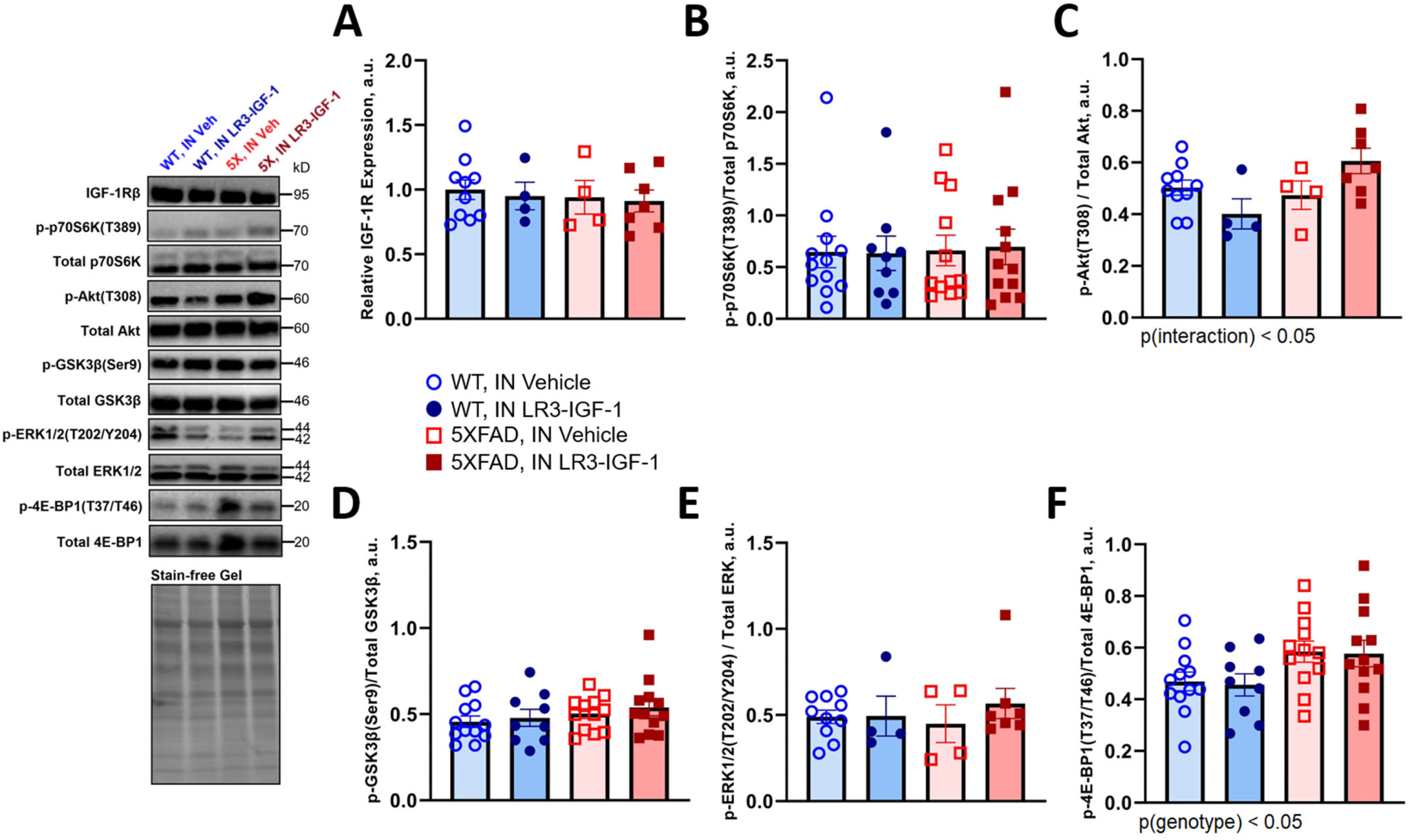
Protein markers of canonical IGF-1 signaling in WT and 5XFAD mice following long-term IN vehicle or LR3 treatment. (A) IGF-1R expression was unaltered with longitudinal LR3-IGF-1 treatment and was not different between 10-month-old male WT and 5XFAD heterozygous mice (n = 4–10 per group). (B-F) Immunoblotting for total and phosphorylated proteins downstream of IGF-1R was used to assess changes in basal signaling following 7 months of treatment, 24 h after the final intranasal dose. Basal phosphorylation of p70S6 K (n = 9–12), GSK3β (n = 9–12), and ERK1/2 (n = 4–10) were not different among treatment groups. A significant interaction was observed for phosphorylation of Akt on Threonine 308 (p < 0.05), but no significant differences were observed among planned comparisons (n = 4–10). Phosphorylation of 4E-BP1 on Threonine 37 and 46 was modified by 5XFAD genotype but was not different among groups (n = 9–12). Image of the strain-free gel is included to confirm equal loading. Data are shown as mean ± S.E.M.
Amyloid-β burden and plaque morphology
We next characterized the effect of LR3 treatment on amyloid pathology. Aβ plaque load was robust in both 5X-Veh and 5X-LR3 mouse brain, with undetectable levels in WT mice (Figure 3A). In cortex, a trend toward fewer 41–60 µm plaques was observed (Figure 3B, p = 0.0614) whereas plaque diameter was unchanged in hippocampus with IN LR3-IGF-1 treatment (Figure 3C). To further evaluate the impact of IN LR3-IGF-1 treatment on plaque morphology and microglial colocalization to plaques, Aβ and Iba1 stained sections were complemented with MeX04 to delineate dense plaque cores (Figure 3D). Although the number of MeX04-positive puncta was unchanged with IN LR3-IGF-1 (Supplemental Figure 4), the integrated Aβ fluorescent intensity was significantly reduced in 5X-LR3 cerebral cortex (Figure 3G, p < 0.05). In cerebral cortex, IN LR3-IGF-1 treatment also reduced the percentage of filamentous plaques (D54D2-positive, MeX04-low) (p < 0.05) and increased the percentage of inert plaques (D54D2-low, MeX04-positive) (p < 0.01; Figure 3F); similar trends were also observed in hippocampus but were not significant (Figure 3E).
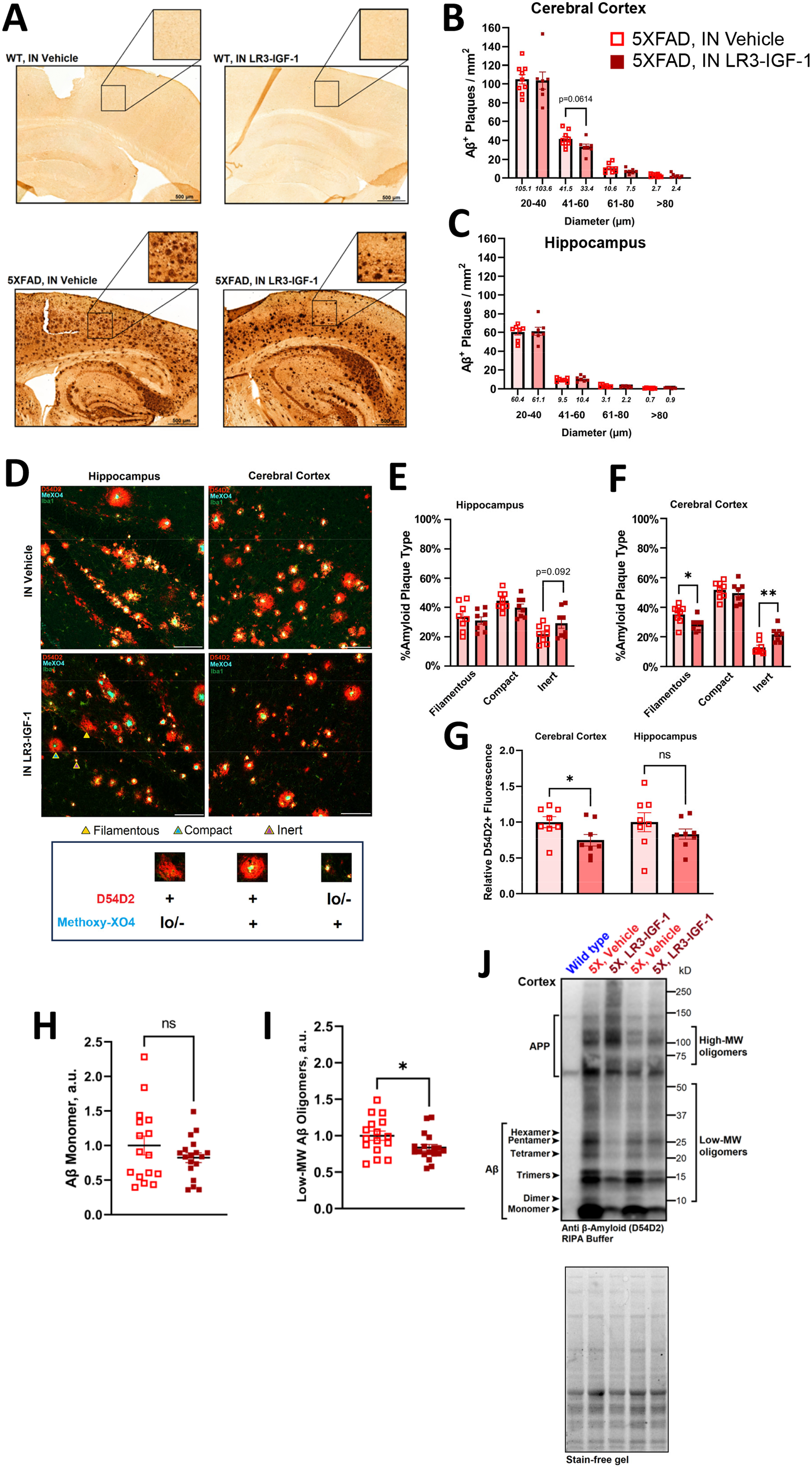
Aβ phenotype in WT and 5XFAD mice following long-term IN vehicle or LR3 treatment. (A) Diaminobenzidine (DAB) stain of Aβ (1:500, D54D2) in sagittal sections obtained from intranasally treated mice (n = 4 per WT group, n = 7–8 per 5XFAD group). Plaques are undetectable in WT mice but are abundant in both 5X-Veh and 5X-LR3 mice. Scale bar = 100 µm. (B,C) Size distribution analysis of D54D2 + plaques. A trend toward reduced plaques in the 40–60 um range was seen in cerebral cortex with IN LR3-IGF-1 treatment (p = 0.0614). (D) Triple stain of D54D2+ Aβ, Iba1 (microglia marker), and methoxy-X04 to identify dense plaque cores, n = 8 per group. Representative filamentous (D54D2+,MeX04 lo/-), compact (D54D2+,MeX04+), and inert (D54D2 lo/-,MeX04+) plaques are indicated with arrows. 20× magnification, scale bar = 100 µm. (E) (E-F) Percentage of each plaque type in cerebral cortex and hippocampus. The percentage of filamentous plaques is diminished (35% versus 29%, p < 0.05) and of inert plaques is increased (13% versus 22%, p < 0.01) in cerebral cortex with IN LR3-IGF-1 treatment, with similar, but non-significant, changes observed in the distribution of plaque types in hippocampus. (G) Normalized D54D2 + fluorescence in 5X-Veh compared with 5X-LR3. Cortical D54D2 + fluorescence is reduced in IN LR3-IGF-1 treated 5XFAD mice (1.0 versus 0.62, p < 0.01) but is not significantly different in hippocampus (1.0 versus 0.77, p = 0.170). (H-J) Representative immunoblot and quantification for RIPA-soluble Aβ (D54D2) in cerebral cortex. Image of the strain-free gel is included to confirm equal loading. Whereas only APP bands can be seen in WT control, diffuse D54D2 bands are seen in 5XFAD lysate, representing multimers and increasingly large aggregates. Low-molecular weight oligomers are observable between 10–50 kD, with higher-molecular weight oligomers in the >50 kD range. No significant change was observed in normalized Aβ monomer expression (cortex, 1.0 versus 0.82, p = 0.267; hippocampus, 1.0 versus 0.96, p = 0.642), but low molecular-weight Aβ oligomers were reduced in cerebral cortex (1.0 versus 0.84, p < 0.05). N = 16–18 per 5XFAD group. Data are shown as mean ± S.E.M. *p < 0.05.
5XFAD mice expressed notable levels of Aβ on western blot, relative to WT, along with diffuse D54D2 + banding indicative of Aβ multimers and larger aggregates (Figure 3J). In cerebral cortex (Figure 3H) and hippocampus (Supplemental Figure 4E), Sβ monomer protein expression was not modified with IN LR3-IGF-1 treatment. However, the burden of low-molecular weight (10–50 kD) Aβ oligomers was significantly reduced in cerebral cortex in 5X-LR3 mice (Figure 3I; p < 0.05). However, TBS-soluble and GuHCl-soluble fractions obtained from cortex and hippocampus showed no significant changes in Aβ42 concentrations, nor were plasma levels of Aβ40 and Aβ42 altered with IN LR3-IGF-1 treatment (Supplemental Figure 4).
Behavior and visuospatial memory performance
A series of behavioral and memory assays were performed in WT and 5XFAD groups. Visuospatial memory performance, as measured by percentage alternations in Y-maze, did not detect any deficit in 5XFAD groups, compared to WT, nor was any improvement observed with IN LR3-IGF-1 treatment in either WT or 5XFAD mice (Figure 4A). We further assessed visuospatial memory via Box Maze, where overall escape latency was initially impaired in 5XFAD mice (p(genotype) < 0.0001), but these groups still appeared to demonstrate rapid learning regardless of treatment, as demonstrated by convergence with controls at trial 2 onward (Figure 4B). Meanwhile, no significant interactions were observed in the Open Field Test (Figure 4C, D) or Zero Maze test (Figure 4E). In the Porsolt Forced Swim test, a significant genotype (p < 0.005) and genotype×treat interaction (p < 0.05) were observed, with 5X-LR3 tending to be more mobile than 5X-Veh (Figure 4F, p = 0.0788). Importantly, IN LR3-IGF-1 treatment did not impair olfaction, as indicated by latency to detect a familiar scent on which the animals were trained (Figure 4G).
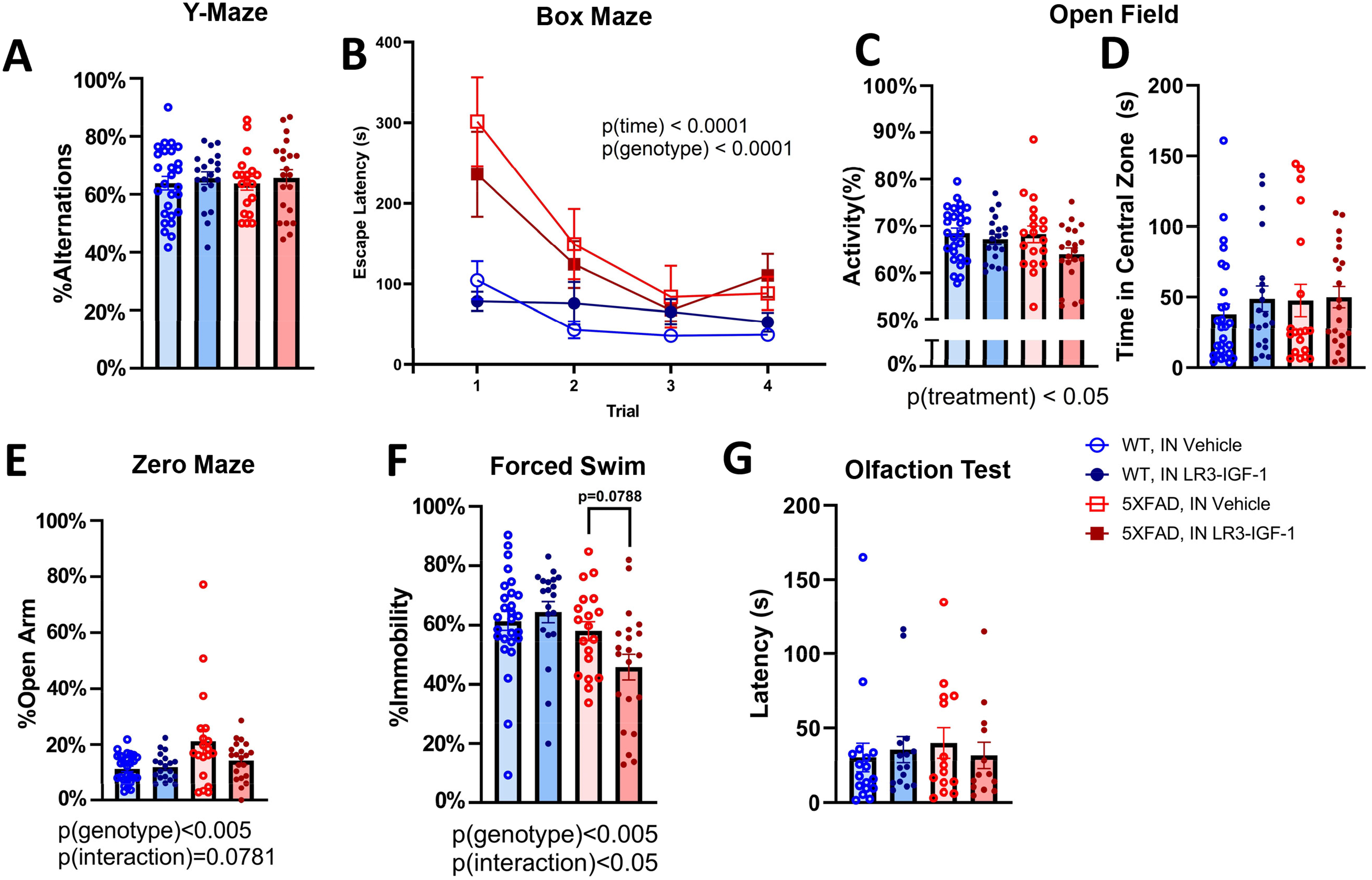
Behavioral and memory performance following long-term IN vehicle or LR3 treatment. A series of behavioral and cognitive assays were performed in intranasally treated mice between 8–9 months of age, 5–6 months into intranasal treatment. (A) Y-maze did not detect any cognitive deficits in 5XFAD mice relative to WT nor was there any effect of treatment. (B) During the box maze, 5XFAD mice take significantly longer than WT mice to escape the arena (p < 0.0001), but all groups reduce latency as trials progress (p < 0.0001). (C-D) Treatment (p < 0.05), but not genotype, led to altered activity in the open field test, but all groups spend a comparable amount of time in the central zone of the open field. (E) 5XFAD mice spend more time in the open arm of the zero maze relative to WT (p < 0.005), but the percentage of time spent in the open arm is not significantly different among groups. (F) 5XFAD mice are less immobile in the forced swim test than WT (p < 0.005), with 5X-LR3 mice tending to show reduced immobility relative to vehicle-treated groups (p = 0.0788). (G) Latency to detect an olfactory stimulus is unchanged with intranasal LR3-IGF-1 treatment and does not differ among groups. (F) n = 19–27 for all assays except box maze (n = 13–17). Data are shown as mean ± S.E.M.
Structural MRI, cerebral blood flow, and neuronal markers
MRI was performed in a subset of mice to non-invasively assess effects on features of brain health. Neither total brain volume nor cerebral blood flow differed with genotype or treatment (Figure 5A,B). However, T2* relaxation time was significantly reduced in 5X-Veh mice relative to WT in the cortical frontal lobe (p < 0.05), but not in 5X-LR3 mice (Figure 5C). In hippocampus, IN LR3-IGF-1 tended to increase T2* time in WT, but decreased T2* time in 5XFAD mice (genotype×treatment = p < 0.05) (Figure 5D). In the parieto-temporal lobe of cortex, FA was significantly diminished in 5XFAD mice compared with WT (p < 0.005), whereas axial diffusivity was unchanged (Figure 5E, F). In hippocampus, fractional anisotropy was similarly reduced in 5XFAD mice (p < 0.0005) (Figure 5G), as was axial diffusivity (p < 0.005), but in no instance were these parameters impacted by treatment. Moreover, immunostaining of neuronal markers (NeuN + cells) revealed no difference in cortical neuronal density or PSD95 protein levels among groups (Supplemental Figure 6).
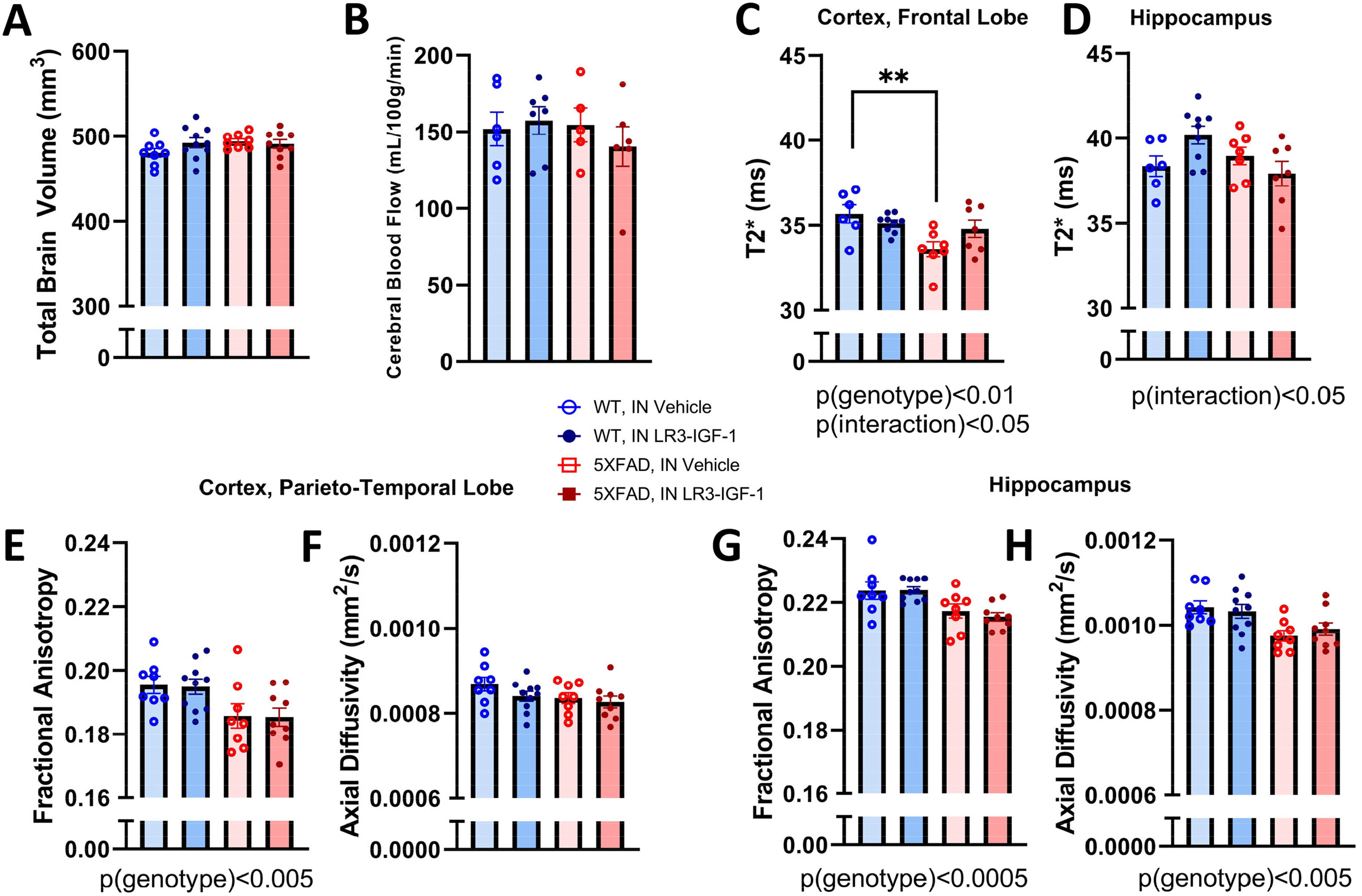
Structural MRI and cerebral blood flow. (A,B) Neither total brain volume nor cerebral blood flow significantly varied among groups. (C) In the cortical frontal lobe, a significant interaction was observed (p < 0.05) for T2*-weighted relaxation time and was found to decrease in 5X-Veh mice relative to WT (p < 0.01). (D) In hippocampus, a significant interaction (p(interaction) < 0.05) for T2* relaxation time was observed, but there were no differences among groups. (E,F) In the cortical parieto-temporal lobe, fractional anisotropy was lower in 5XFAD mice relative to WT (p < 0.005), but axial diffusivity was unaltered. (G,H) In hippocampus, fractional anisotropy and axial diffusivity were both reduced by 5XFAD genotype (p < 0.005). n = 5–10 per group. Data are shown as mean ± S.E.M. **p < 0.01.
BV-2 cells internalize Aβ and activate endophagocytic pathways in response to LR3-IGF-1
In order to determine the possible connection between LR3 treatment and microglia in 5XFAD mice, we assessed the effects on BV-2 cell characteristics on microglial Aβ uptake in vitro (Figure 6). Pre-treatment of BV2 cells with 50 nM LR3-IGF-1 for 3 h augmented Aβ fluorescence without changing the number of Aβ-positive intracellular puncta in BV-2 cells (Figure 6A-C, p < 0.05). Moreover, RNAseq analysis of BV-2 cells treated for 4 h with 1 µM oligomeric Aβ in the presence of 50 nM LR3-IGF-1 (AR) elicited a transcriptomic profile distinct from that of oligomeric Aβ alone (A) (Figure 6D-F). Furthermore, pdcd4, ypel3, and klhl24 genes were significantly downregulated, and gm12033 upregulated, in AR relative to A (Figure 6E and Supplemental Table 2). KEGG analysis further revelated key pathways enriched in the AR group, including those involved with regulation of the actin cytoskeleton, FCγR-mediated phagocytosis, endocytosis, junctional maintenance and cellular adhesion, and responses to bacterial infection (Figure 6F and Supplemental Table 1). In contrast, pathways involved in Huntington's disease, Parkinson's disease, and AD were negatively enriched in the AR group.
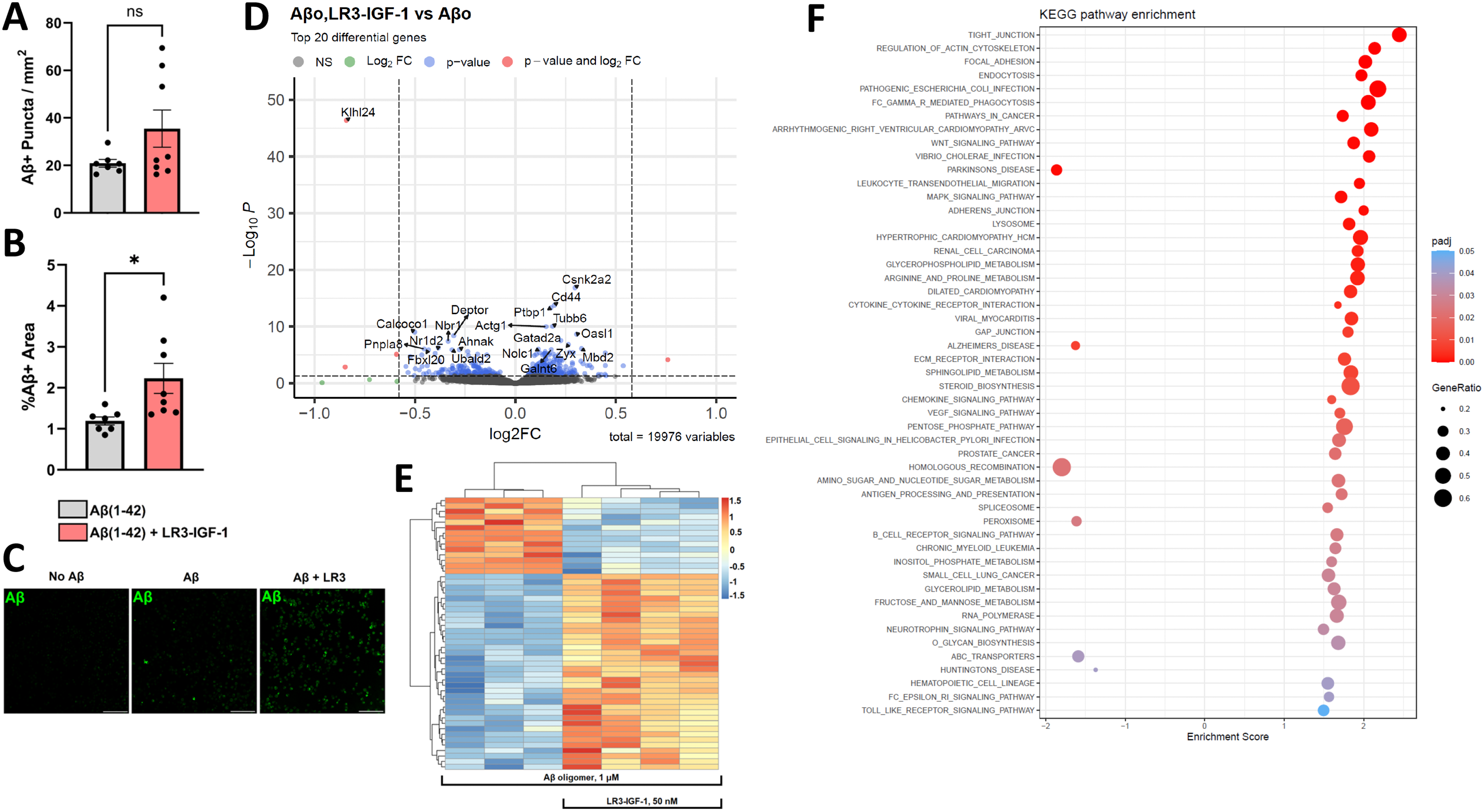
LR3-IGF-1 stimulation of Aβ1–42 peptide uptake and transcriptional changes in BV-2 cells. (A-C) BV-2 cells were incubated with 200 nM HiLyte Fluor™ 488 Aβ1−42 for 1 h with or without pre-treatment for 3 h with 50 nM LR3-IGF-1 (n = 7–8 per condition). LR3-IGF-1 pre-treatment did not significantly increase the number of puncta but did increase the %Aβ-fluorescent area (p < 0.05). Scale bar = 100 µm. (D-F) Bulk transcriptomic analysis of BV-2 cells treated for 4 h with 1 µm oligomeric Aβ (A) or LR3-IGF-1 + Aβ (AR) (n = 4 biological replicates per condition. (E) Volcano plot comparing AR versus A. The top 20 differentially expressed genes are shown. (F) Heatmap of AR versus A with hierarchical clustering of top 50 genes. (G) Normalized enrichment scores for top KEGG pathways positively or negatively enriched in AR versus A. Bar graphs and lines show results as mean ± S.E.M. *p < 0.05.
Discussion
Here we show that longitudinal IN LR3-IGF-1 treatment of male 5XFAD mice from 3 to 10 months of age was able to induce favorable remodeling of plaques in cerebral cortex, as evidenced by the relative abundance of filamentous plaques and increased fraction of inert plaques in cerebral cortex. The abundance of dense-core plaques is constant between preclinical and late-stage AD in human patients, but a shift from geometrically irregular, diffuse plaques to spherical fibrillar plaques is seen with disease progression. 46 5XFAD mice in this study showed a hyperabundance of compacted, dense-core plaques, as indicated by MeX04 positivity of the central core, regardless of IN LR3-IGF-1 treatment. However, the depletion of loosely aggregated D54D2+ Aβ forming the outer corona of spherical plaques seen with LR3-IGF-1 treatment explains both the reduction in 41–60 µm plaques as well as the increased fraction of inert plaques. However, IN LR3-IGF-1 was unable to preserve or improve aspects of behavior or visuospatial memory, which may in part be attributed to the lack of robust deficits observed in 5XFAD for some assays. Moreover, the lack of observable therapeutic efficacy with regards to amyloid load or morphology with LR3 treatment in 5XFAD hippocampus was also evident across several assays, though the tendency for preserved T2*-weighted relaxation time in the cortical frontal lobe with IN LR3-IGF-1 treatment in 5XFAD mice is consistent with plaque remodeling in this region.
Constitutive hyperactivation of microglial mTOR via Tsc1 knockout results in a similar reduction in filamentous plaques and increase in inert plaques due to increased microglial activation and TREM2 expression. 45 Microglia play a critical role in remodeling plaques and phagocytosing Aβ and cellular debris, while depletion of microglia via CSF1R inhibition in 5XFAD mice shifts plaques toward a more diffuse, less compacted state. 48 It has been proposed that the dense-core plaques seen in late-stage AD are granulomatous structures formed by chronically-activated microglia, that internalize and aggregate oligomeric and fibrillar Aβ and exocytose the aggregates that go on to form plaques. 49 In this study, low-molecular weight Aβ oligomers were decreased in the cerebral cortex of IN LR3-IGF-1 treated 5XFAD mice, in conjunction with persistently activated trem2 and iba1 expression and elevated il1b. Together, these findings suggest that longitudinal IN LR3-IGF-1 treatment results in activation of brain-resident immune cells and enhanced Aβ compaction into dense-core, inert plaques.
IGF-1 modulates phagocytic activity of astrocytes via PI3K/Akt signaling, but its effect on microglial phagocytosis has not been well characterized. 47 In this study, we have demonstrated that LR3-IGF-1 similarly induces Aβ1-42 uptake in BV-2 microglial cells. Neither the quantitative burden of Aβ40 and Aβ42 peptides in 5XFAD brain, nor plasma Aβ levels, were altered with IN LR3-IGF-1 treatment. This observation is consistent with an effect of treatment primarily on plaque remodeling rather than an effect on Aβ clearance or export to the peripheral circulation. Consistent with this observation, LR3-IGF-1 triggers BV-2 cells to upregulate transcriptional pathways involved in actin cytoskeletal remodeling, phagocytosis, and endocytosis in the presence of Aβ oligomers.
Brain IGF-1 resistance is a feature of terminal AD, 50 though it is unclear whether this constitutes a compensatory response to combat hyperactivation of mTOR and preserve autophagy and amyloid clearance, 51 or if disturbances in insulin/IGF-1 signaling in AD contribute to pathology and disease progression. 52 Indeed, others have shown that either whole-body IGF-1R haploinsufficiency confers protection against cognitive decline in APP/PS1 mice and induces hyperaggregation of Aβ with consequent reduction in Aβ plaque area in 12–13-month-old mice, 53 while neuronal IGF-1R deletion protects against Aβ toxicity and preserves cognition in APP/PS1 mice, 54 favoring the interpretation that early suppression of insulin/IGF-1 signaling is a protective response to stave off amyloid proteotoxicity. In contrast, isolated IGF-1 deficiency in brain has been clearly linked to deleterious effects by many labs, while we and others have also reported a relative deficiency of IGF-1 and insulin in aged rat CSF, 43 while raising central IGF-1 levels confers benefits to the brain of aging male mice. 24 In addition, the notable ability of LR3-IGF-1 to modulate aspects of amyloid plaque in this study coupled with extensive, positive signals from intranasal insulin trials 55 suggests that efforts toward exploring and optimizing the therapeutic window and provisional strategies of one or more of these growth factors, alone, or in combination with other emerging therapies, should continue to be pursued.
While IN LR3-IGF-1 was able to induce the favorable remodeling of plaques in cerebral cortex, its effects were uniformly more pronounced in cortex compared with hippocampus, which may partially explain why memory benefits were absent. We cannot explain why effects are seen only in cortex but speculate that this could be due to several possibilities. For instance, hippocampus may be inherently more refractory to treatment than cortex in this model, while pharmacokinetic limitations of LR3-IGF-1 to adequately reach and penetrate hippocampus, may have also played a role. Indeed, owing to its inability to be bound by IGFBPs, LR3-IGF-1 has increased potency in vitro but its distribution and half-life may be altered in vivo. Along these lines, we cannot rule out that more aggressive dosing (amount and/or frequency) could potentially overcome these limitations. Alternatively, while IN insulin trials in APP/PSEN mice proved efficacious, the 5XFAD model used here is notoriously more aggressive and difficult to treat. To develop a comprehensive understanding of the potential utility of IN LR3-IGF-1 in AD prophylaxis or treatment, future studies will need to evaluate its efficacy with additional dosing schedules using both sexes, and in other amyloid and tauopathy models of AD.
In summary, IN LR3-IGF-1 shifts Aβ plaque abundance away from large, filamentous plaques to smaller, inert plaques, and reduces soluble low-molecular weight Aβ oligomers in cerebral cortex, but these effects were less pronounced in hippocampus of 5XFAD male mice. This corresponded with a lack of obvious benefit with LR3 treatment on several indices of cognition, and thus do not support LR3 as an effective candidate for monotherapy in AD. Nevertheless, further research into combinatorial strategies that include intranasally-administered IGF-1 and its analogues along with an array of other intriguing ligands, such as BDNF, VGF, insulin, and Klotho, could cooperatively form the basis of precision therapies to target the multiple, complex facets contributing to the instigation, progression, and manifestation of AD and related dementias.
Supplemental Material
sj-docx-1-alz-10.1177_13872877241299056 - Supplemental material for Intranasal long R3 insulin-like growth factor-1 treatment promotes amyloid plaque remodeling in cerebral cortex but fails to preserve cognitive function in male 5XFAD mice
Supplemental material, sj-docx-1-alz-10.1177_13872877241299056 for Intranasal long R3 insulin-like growth factor-1 treatment promotes amyloid plaque remodeling in cerebral cortex but fails to preserve cognitive function in male 5XFAD mice by Matthew G Engel, Sushma Narayan, Min-Hui Cui, Craig A Branch, Xusheng Zhang, Samuel E Gandy, Michelle Ehrlich and Derek M Huffman in Journal of Alzheimer's Disease
Footnotes
Acknowledgments
We would like to recognize technical assistance provided by Hillary Guzik, M.Sc., and Vera DesMarais, PhD We also thank Kamalakar Ambadipudi, Hongqian Liang, and Zhiping Wu for technical assistance.
Author contributions
Matthew G Engel (Conceptualization; Data curation; Formal analysis; Investigation; Methodology; Project administration; Validation; Visualization; Writing – original draft; Writing – review & editing); Sushma Narayan (Investigation; Methodology; Writing – review & editing); Min-Hui Cui (Data curation; Formal analysis; Investigation; Validation; Visualization; Writing – original draft); Craig A Branch (Formal analysis; Investigation; Methodology; Supervision; Writing – original draft); Xusheng Zhang (Formal analysis; Methodology; Writing – original draft); Samuel E Gandy (Conceptualization; Funding acquisition; Methodology; Supervision; Writing – original draft); Michelle Ehrlich (Conceptualization; Methodology; Supervision; Writing – review & editing); Derek Huffman (Conceptualization; Data curation; Formal analysis; Funding acquisition; Investigation; Methodology; Project administration; Supervision; Visualization; Writing – original draft; Writing – review & editing).
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by NIH grants R56 AG066431 and P30AG038072, and the American Federation for Aging Research to D.M.H. M.G.E. is supported by a T32 Training Grant (T32 GM007288) and S.N. is supported by T32 Training Grant (T32 AG023475). This work was also supported by the Analytical Imaging Facility at Albert Einstein College of Medicine under the NCI Cancer Center Support Grant P30CA013330 and Shared Instrument Grants 1S10OD019961-01 and 1210OD023591-01.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
Data are freely available from the corresponding author upon reasonable request. Transcriptomic data are deposited in SRA.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.