
Abstract
Background
Increasing numbers of studies indicate that the pathophysiological progression of Alzheimer's disease (AD) could be accelerated by chronic cerebral hypoperfusion (CCH). These findings suggest that cerebrovascular dysfunction may induce cognitive impairment in AD by expediting neurodegeneration. Dendrobium nobile Lindl. alkaloids (DNLA), the primary active components of Dendrobium nobile Lindl., have been shown to enhance cognitive function and exhibit neuroprotective effects in AD animal models.
Objective
However, the impact of DNLA on AD with CCH is still elusive.
Methods
In this study, we explored the therapeutic potential and underlying mechanisms of DNLA using a novel AD plus CCH mouse model.
Results
Our results demonstrate that DNLA significantly improved cerebral blood flow, attenuated motor and cognitive decline, reduced amyloid-β deposition, mitigated neuroinflammation, and alleviated neural oxidative stress in 12-month-old AD mice with CCH.
Conclusions
Our study suggests that DNLA exerts multiple neuroprotective effects, effectively preserving motor and cognitive function in AD with CCH mice of 12 months. Thus, DNLA represents a promising therapeutic candidate for the prevention and treatment of AD plus CCH.
Keywords
Introduction
Alzheimer's disease (AD), a progressive neurodegenerative disorder pathologically characterized by amyloid-β (Aβ) plaques and neurofibrillary tangles, is projected to affect approximately 131.5 million individuals globally by 2050. 1 Emerging evidence highlights chronic cerebral hypoperfusion (CCH) as a critical contributor to AD progression, exacerbating both pathological hallmarks and cognitive decline.2–4 However, no effective therapies currently exist for AD, particularly in cases complicated by CCH.
Our recent studies have shown that the therapies, such as Galanthamine, Edaravone, and plasmalogen, targeting multiple mechanisms, including anti-inflammatory effects and oxidative stress reduction, are effective in improving cognitive deficits of the novel AD + CCH mouse model.5–9 However, such previous therapies have their disadvantages, such as non-oral administration, cumulative side effects, high expense, and limited accessibility. Consequently, the development of a safer, orally administered alternative for chronic use, lower cost, and readily available is of paramount importance.
Dendrobium nobile, a valuable epiphytic orchid species, has been traditionally used in China for centuries as a medicinal herb and functional health supplement. 10 It is composed of various bioactive compounds, including alkaloids and polysaccharides. 11 These components exhibit broad pharmacological properties such as antioxidant, immunomodulatory, anti-aging, hypoglycemic, and hypolipidemic effects. 12 Through these mechanisms, it contributes to the treatment of various diseases, including tumors, diabetes, hyperlipidemia, and age-related neurodegenerative disorders. 12
Notably, Dendrobium nobile Lindl. alkaloids (DNLA), the primary bioactive components of Dendrobium nobile, have demonstrated therapeutic potential in AD models, ameliorating cognitive deficits via multiple mechanisms including Aβ clearance, anti-inflammatory effects, oxidative stress reduction, and synaptic plasticity enhancement.10–12 However, the efficacy and underlying mechanisms of DNLA in AD with CCH remain unexplored.
Our research group recently developed a novel AD with CCH model using APP23 transgenic mice expressing human Swedish mutant amyloid precursor protein.5–9 This model effectively mimics the CCH progression, behavioral impairments, and neuropathological features observed in clinical AD patients with comorbid cerebrovascular dysfunction.13,14 Therefore, in the present study, we aim to investigate the potential effects and mechanisms of DNLA on this AD plus CCH mouse model.
Methods
Experimental model and drug treatment
All animal experiments were conducted in accordance with the guidelines of the Animal Committee of Guizhou Medical University (Approval No. GMU#2303239), the ARRIVE guidelines (https://www. nc3rs.org.uk), and institutional regulations for the Care and Use of Laboratory Animals. APP23 transgenic mice, a well-established AD model, were used in this study. These mice overexpress the human APP gene containing the Swedish mutation under the control of the Thy1 promoter and were derived from B6, D2-TgN (Thy1-APPSwe) strain.15–17
Sample size estimation was based on preliminary data and statistical power requirements. 18 To detect a significant intergroup difference in cerebral blood flow (CBF) reduction (α = 0.05, power = 80%) between the vehicle group and the DNLA group, eight mice per group was required. According for an anticipated 30% attrition rate by 12 months (prior unpublished data), we initially assigned fifteen 4-month-old male mice to each experimental group (total N = 45). And 15 of all 45 mice were excluded due to the exclusion criteria: perioperative mortality during CCH surgery (n = 10), failure to exhibit post-CCH CBF reduction (n = 5). Consequently, the final 12-month cohorts include: APP23 group (APP23 + sham surgery, n = 10), CCH group (APP23 + CCH, n = 10), and DNLA group (APP23 + CCH + DNLA, n = 10).
CCH was surgically induced using ameroid constrictors (0.75 mm internal diameter; Research Instruments NW, Lebanon, OR, USA). 8 At 4 months of age, mice in the CCH and DNLA treatment groups underwent bilateral common carotid arteries (BCCAs) occlusion via constrictor placement following cervical incision. Mice in the APP23 + CCH + DNLA group received daily oral gavage of DNLA (40 mg/kg) for 8 months until sacrifice, based on established protocols.10–12 Sham surgery was performed in control (APP23) mice.
In detail, all mice were housed under controlled environmental conditions (constant temperature, humidity, and light) for at least one week before surgery for acclimatization. Mice underwent a 6-h fasting period (with ad libitum water access) immediately before surgical procedures and then all mice were randomly assigned to experimental groups before surgical procedures.
During surgery, 4-month-old mice were anesthetized via inhalation of a nitrous oxide/oxygen/isoflurane mixture (69%:30%:1%, respectively) in an induction chamber. Surgical anesthesia was confirmed by absence of pedal withdrawal reflex. Following cervical incision, ameroid constrictors (0.75 mm ID) were bilaterally implanted on the common carotid arteries.
After surgery, mice were maintained in a thermoregulated recovery chamber (37°C) until fully ambulatory. Animals were then individually housed under standard conditions with ad libitum access to food and water. For postoperative analgesia, buprenorphine (0.05 mg/kg, 0.015 mg/ml) was administered via intramuscular injection.
Preparation of Dendrobium nobile Lindl. alkaloids
The high-purity DNLA (molecular weight: 263.27, CAS: 2115-91-5) used in this study were obtained from Chengdu Herbpurify Co., Ltd (Batch: S-023, Chengdu, China), with a total alkaloid purity of 99.87%. The principal bioactive constituents were quantitatively characterized as Dendrobine (C16H25O2N, 98.62%), Nobilonine (C17H27O3N, 0.55%), and Dendrobine-N-oxide (C16H25O3N, 0.81%). For administration, DNLA was dissolved in sterile distilled water containing 0.5% Tween 80 to achieve a working concentration of 2 mg/ml, as previously reported. 10
Cerebral blood flow test
CBF was assessed in 12-month-old mice prior to sacrifice using a laser-Doppler flowmeter (FLO-C1, Omegawave, Tokyo, Japan). Under anesthesia (69% nitrous oxide/30% oxygen/1% isoflurane via inhalation chamber), the probe was stereotaxically positioned perpendicular to the skull (1 mm posterior and 2.5 mm lateral to bregma). Following loss of pedal withdrawal reflex, five consecutive CBF measurements were obtained and averaged for analysis.
Behavioral test
Motor coordination and balance were evaluated using an accelerating Rotarod apparatus (MK670; Muromachi Kikai Co., Tokyo, Japan) as previously reported.5,19 The speed progressively increased from 4 to 40 rpm over 5 min. The latency duration (seconds) until trial completion or mice fell was recorded for each trial. We repeated the Rotarod test five times with an interval of five minutes between each trial.
Working memory was evaluated in 12-month-old mice using an 8-arm radial maze following established protocols.5,6,20,21 Briefly, mice experienced food restriction to maintain 10% body weight reduction and Ad libitum water access throughout testing. As for acquisition phase, a 5-day maze adaptation period was conducted 1 trial/day. During the acquisition phase, mice can explore the 8-arm maze for five minutes. For each formal trial, mice can freely select arms until either the consumption of all 4 baited pellets or 5 min is reached. Working memory errors are quantified as re-entries into previously visited baited arms.
Tissue preparation and immunohistochemistry
At 12 months of age, all mice were deeply anesthetized with an intraperitoneal injection of pentobarbital (40 mg/kg) and transcardially perfused with 20 mL ice-cold PBS followed by 20 mL 4% paraformaldehyde in 0.1 M phosphate buffer. After overnight post-fixation in 4% PFA at 4°C, brains were sectioned coronally at 50 µm thickness using a vibrating microtome (LEICA VT1000S). For immunohistochemical staining, free-floating sections were first treated with 0.6% periodic acid to quench endogenous peroxidase activity, then blocked with 5% BSA containing 0.1% Triton X-100 in PBS (pH 7.4) to prevent nonspecific binding. Sections were incubated overnight with primary antibodies against Aβ oligomer (A11, 1:200; Thermo Fisher), Aβ40 (BA27, 1:100; Wako), GFAP (ab53554, 1:100; Abcam), Iba-1 (NCNP24, 1:200; Wako), IL-1β (AF-401-NA, 1:200; R&D Systems), TNF-α (AF-410-NA, 1:300; R&D Systems), 4-HNE (MHN-020P, 1:100; JaICA), and 8-OHdG (MOG-100P, 1:100; JaICA), with negative control sections processed without primary antibodies. Immunoreactivity was visualized using HRP-conjugated secondary antibodies followed by amplification with Vectastain ABC kit (PK-6104) and development with DAB substrate.
Detection and semi-quantitative analysis
Immunohistochemical analysis was performed using digitized sections captured with an Olympus BX-51 microscope (Olympus Optical Co., Tokyo, Japan). For each brain, three coronal sections at distinct rostrocaudal levels relative to bregma (1.0 mm, 0.5 mm, and 0 mm) were analyzed, with six randomly selected fields captured per section (a total of 18 measurements per animal). Quantitative assessments were conducted using ImageJ software (NIH, Bethesda, USA), with specific analytical approaches tailored to each target: cortical and hippocampal Aβ oligomer, IL-1β, TNF-α, 4-HNE, and 8-OHdG immunoreactivity were quantified by measuring mean pixel intensity of DAB staining; Aβ40-positive deposits were evaluated as the percentage area occupied by immunopositive signals in both cortical regions and thalamic vasculature; while GFAP and Iba-1 immunoreactivity were assessed by enumerating positively stained cells in the regions of interests.
Statistical analysis
To ensure objective assessment, all data collection and analyses were performed by investigators blinded to treatment group allocation. Quantitative data are expressed as mean ± standard deviation (SD). Intergroup comparisons were analyzed by one-way analysis of variance (ANOVA) followed by Tukey-Kramer post hoc testing, with statistical significance defined as p < 0.05.
Results
CBF was improved by DNLA in AD mice with CCH
Laser Doppler measurements revealed a significant reduction in CBF of 12-month-old APP23 + CCH mice compared to age-matched APP23 controls (Figure 1, ##p < 0.01 versus APP23). Notably, chronic DNLA treatment partially restored CBF in CCH mice at 12 months, demonstrating a significant improvement in blood perfusion (Figure 1, *p < 0.05 versus APP23 + CCH).
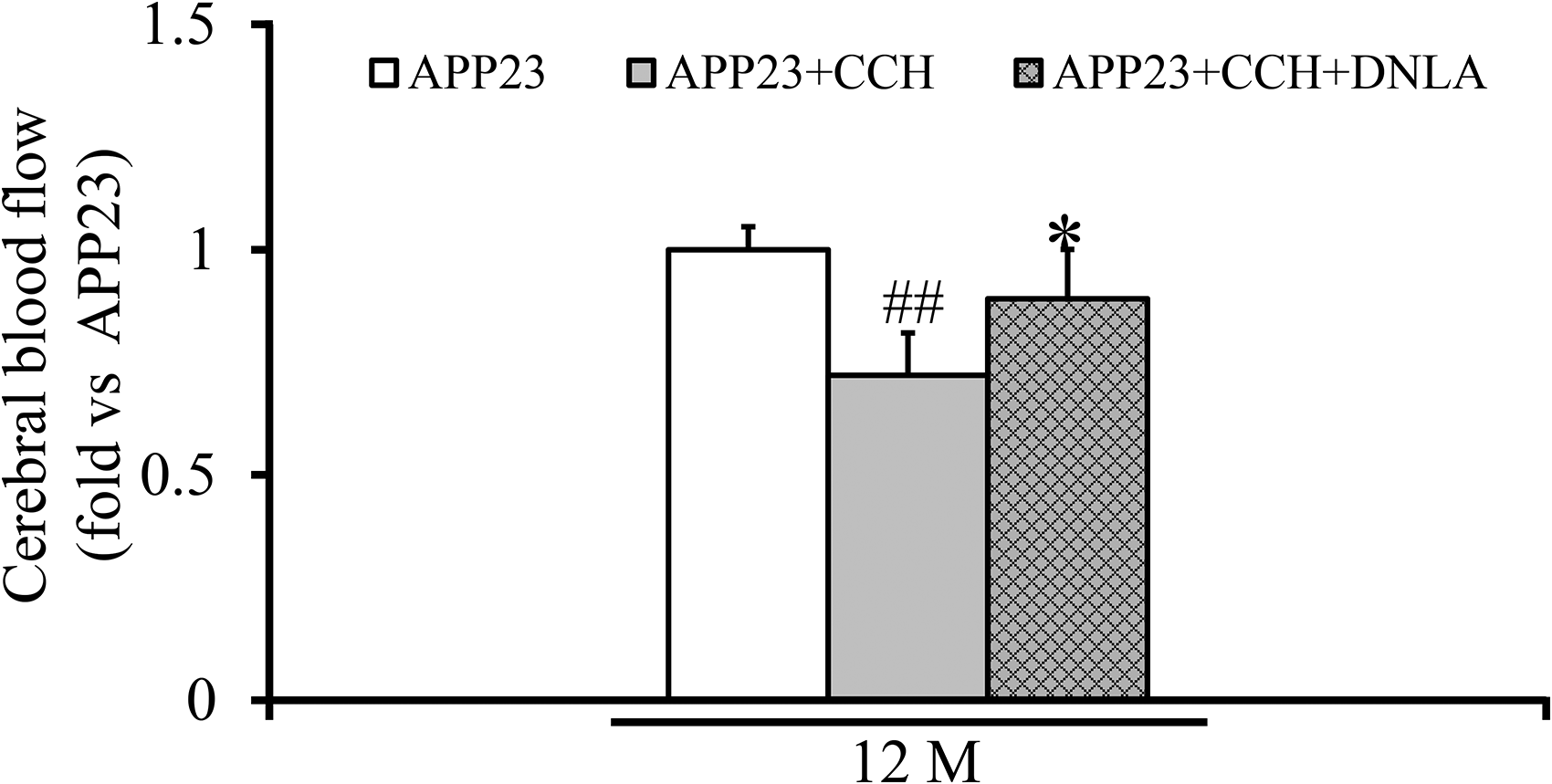
Dendrobium nobile Lindl. alkaloids (DNLA) ameliorate the reduction of cerebral blood flow (CBF) in APP23 mice with chronic cerebral hypoperfusion (CCH) at 12 months (M) of age. Temporal profiles of CBF in mice at 12 months from APP23, APP23 + CCH, and APP23 + CCH + DNLA mice. The CBF values are displayed as fold versus APP23. Data are presented as mean ± SD. Statistical comparisons are tested by one-way ANOVA followed by a Tukey-Kramer posthoc comparison (##p < 0.01 versus APP23, *p < 0.05 versus APP23 + CCH).
Motor and cognitive deficits were ameliorated by DNLA in AD mice with CCH
The Rotarod test demonstrated significantly impaired motor coordination in 12-month-old APP23 + CCH mice, evidenced by reduced latency times across all five trial blocks compared to APP23 controls (Figure 2A, #p < 0.05 versus APP23). DNLA treatment substantially rescued this deficit in APP23 + CCH mice, showing significantly prolonged latency in three trial blocks (Figure 2A, *p < 0.05 versus APP23 + CCH), indicating DNLA's protective effect against CCH-induced motor dysfunction in APP23 mice at 12 months.
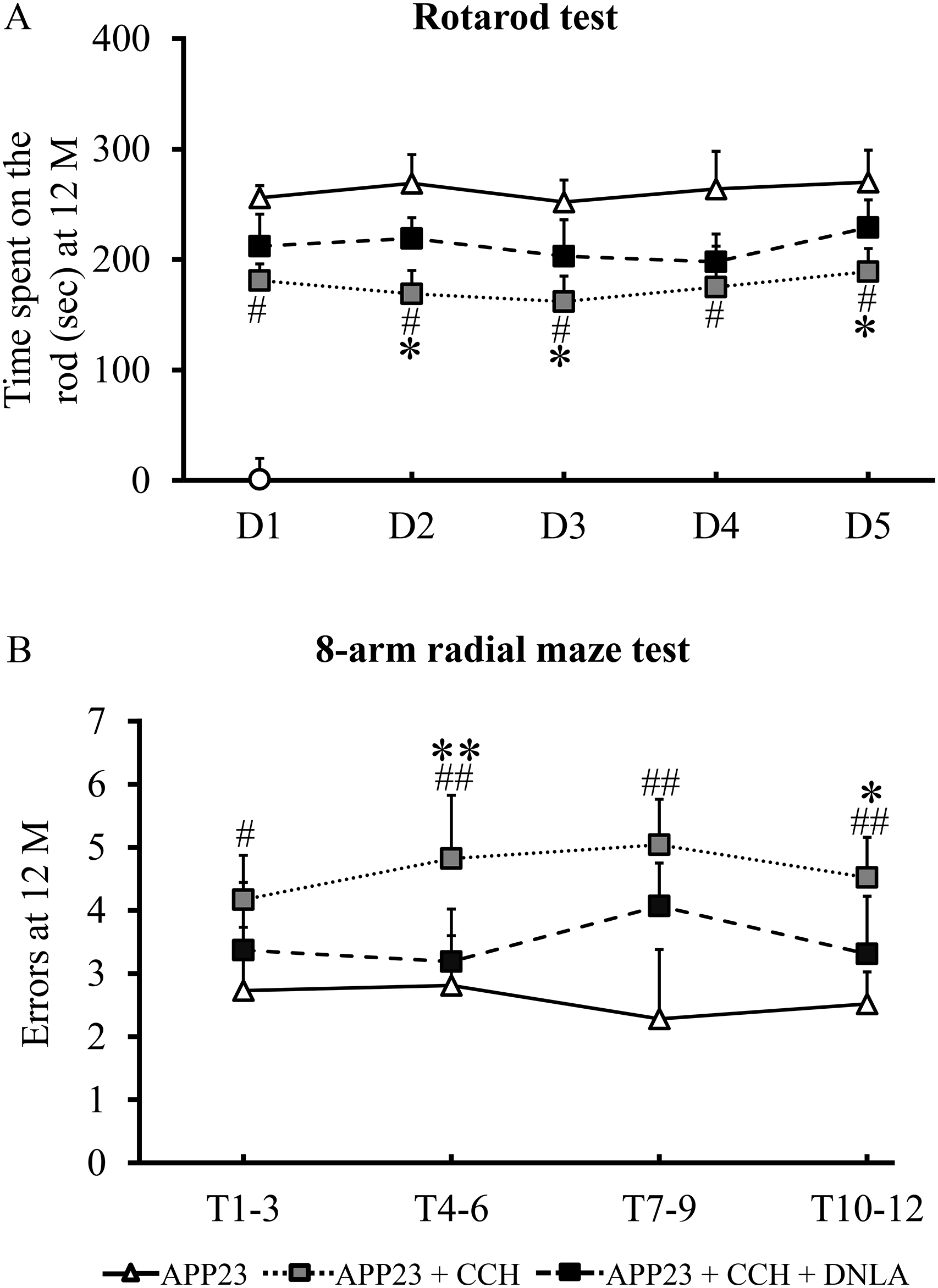
Dendrobium nobile Lindl. alkaloids (DNLA) improve cerebral chronic hypoperfusion (CCH)-induced motor and memory deficits in APP23 mice at 12 months (M) of age. Mean latency time indicates motor capacity in the Rotarod test. Significant shorter mean latency time is in APP23 + CCH mice than that in APP23 mice at 12 months (A). The mean number of re-entry choices indicates working memory capacity in the 8-arm test. Significantly more errors are in APP23 + CCH mice than in APP23 mice at 12 months (B). DNLA intervention significantly attenuated motor and memory deficits in APP23 + CCH mice at 12 months (A and B). D1- D5 means five consecutive trial days (Day 1, Day 2, Day 3, Day 4, and Day 5) and a trial each day (A). T1-T12 represents 12 trials in 12 consecutive trial days and a trial each day (Trial 1-Trial 12) (B). T1-3 is the mean value of trial 1-trial 3. The implication of T4-6, T7-9, and T10-12 is the same as that represented in T1-3 (B). Data are mean ± SD. Statistical comparisons are tested by one-way ANOVA followed by a Tukey-Kramer posthoc comparison (#p < 0.05 versus APP23, ##p < 0.01 versus APP23; *p < 0.05 versus APP23 + CCH, **p < 0.01 versus APP23 + CCH).
The 8-arm radial maze test revealed significant working memory deficits in 12-month-old APP23 + CCH mice, as evidenced by increased revisiting errors compared to APP23 controls (Figure 2B, #p < 0.05 versus APP23, ##p < 0.01 versus APP23). DNLA treatment effectively attenuated this impairment, with DNLA-treated CCH mice committing significantly fewer errors than untreated CCH mice at 12 months (Figure 2B, *p < 0.05 versus APP23 + CCH, **p < 0.01 versus APP23 + CCH), demonstrating DNLA's ability to rescue CCH-induced working memory decline.
DNLA reduced Aβ oligomer and Aβ40 deposition in AD plus CCH mice
Immunohistochemical analysis revealed prominent Aβ oligomer expression in both cellular membranes and cytoplasm throughout cortical and hippocampal regions (Figure 3A). Quantitative assessment demonstrated significantly elevated Aβ oligomer levels in 12-month-old APP23 + CCH mice compared to APP23 controls (Figure 3B, ##p < 0.01 versus APP23). Notably, DNLA treatment substantially reduced Aβ oligomer accumulation in CCH mice at 12 months (Figure 3B, **p < 0.01 versus APP23 + CCH), indicating its potent inhibitory effect on CCH-induced Aβ pathology.
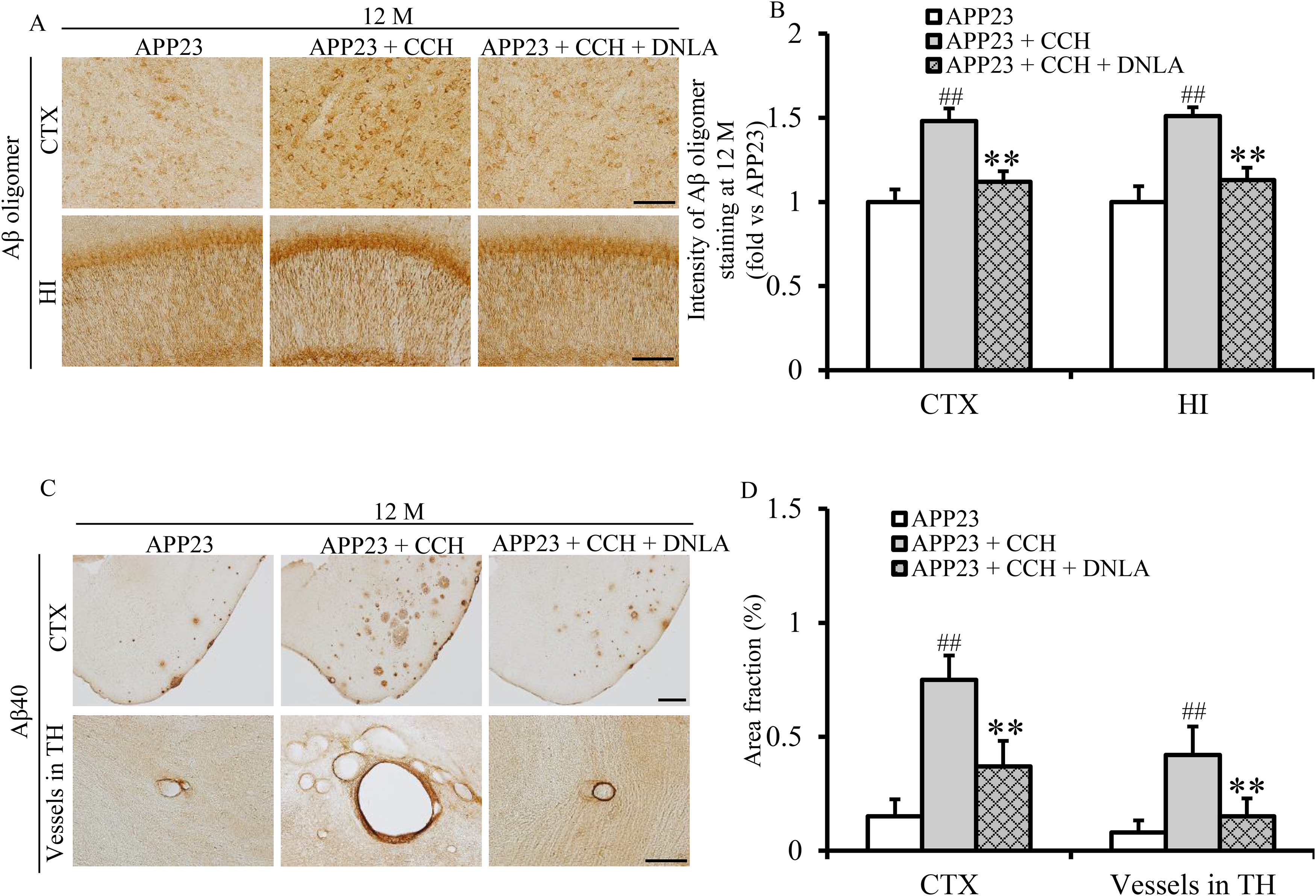
Dendrobium nobile Lindl. alkaloids (DNLA) attenuate cerebral chronic hypoperfusion (CCH)-induced massive amyloid-β (Aβ) oligomer and 40 burdens in APP23 mice at 12 months (M) of age. Representative photomicrographs of Aβ oligomer immunohistochemistry staining (A) and quantitative analysis of Aβ oligomer-positive pixel intensity ratios (B) in the cortex (CTX) and hippocampus (HI) at 12 M. Representative photomicrographs of Aβ40 immunohistochemistry staining (C) and quantitative analysis of Aβ40 area fraction (D) in the CTX and vessels of thalamus (TH) at 12 M. Data are indicated as mean ± SD. Statistical comparisons are tested by one-way ANOVA followed by a Tukey-Kramer posthoc comparison (##p < 0.01 versus APP23; **p < 0.01 versus APP23 + CCH. Scale bar = 50 µm).
Immunohistochemical analysis identified Aβ40 deposits in both cortical parenchyma and thalamic vasculature in APP23 mice (Figure 3C). CCH-treated mice showed significantly exacerbated deposition of Aβ40 in cortex and thalamic vasculature at 12 months (Figure 3D, ##p < 0.01 versus APP23). DNLA treatment effectively attenuated this pathology, demonstrating a marked reduction in Aβ40 burden (Figure 3D, **p < 0.01 versus APP23 + CCH), particularly in cortical and thalamic vascular compartments at 12 months. These findings suggest DNLA potently suppresses CCH-accelerated Aβ40 aggregation in multiple brain regions.
DNLA attenuated neuroinflammation in AD mice with CCH
Immunohistochemical analysis revealed prominent GFAP+ astrocytes, Iba-1+ microglia, IL-1β, and TNF-α expression in cortical and hippocampal regions across all 12-month-old APP23 mouse groups (Figure 4A, C, E, G). Quantitative assessment demonstrated significant increases in GFAP+ astrocytes and Iba-1+ microglia in CCH-treated mice compared to APP23 controls (Figure 4B and 4D, ##p < 0.01 versus APP23), with DNLA treatment effectively reducing both glial cell populations (Figure 4B and 4D, **p < 0.01 versus APP23 + CCH). Similarly, IL-1β and TNF-α expression levels were markedly elevated in CCH mice (Figure 4F and 4H, #p < 0.05 versus APP23, ##p < 0.01 versus APP23) but significantly attenuated by DNLA administration (Figure 4F and 4H, *p < 0.05 versus APP23 + CCH). These findings collectively demonstrate DNLA's potent anti-neuroinflammatory effects in mitigating CCH-induced glial activation and cytokine production in APP23 mice with 12 months.

Dendrobium nobile Lindl. alkaloids (DNLA) mitigate cerebral chronic hypoperfusion (CCH)-induced overexpression of GFAP-positive astrocytes, Iba-1-positive phagocytes, and cytokines (IL-1β and TNF-α) of APP23 mice at 12 months (M) old. Representative photomicrographs of GFAP (A), Iba-1 (C), IL-1β (E), and TNF-α (G) immunohistochemistry staining in the cortex (CTX) and hippocampus (HI) at 12 M. Quantitative analysis of the number of GFAP-positive astrocytes (B) and Iba-1-positive phagocytes (D), and the pixel intensity ratios of IL-1β (F) as well as TNF-α (H)-positive signals in the CTX and HI at 12 M. Data are presented as mean ± SD. Statistical comparisons are tested by one-way ANOVA followed by a Tukey-Kramer posthoc comparison (#p < 0.05 versus APP23, ##p < 0.01 versus APP23; *p < 0.05 versus APP23 + CCH, **p < 0.01 versus APP23 + CCH. Scale bar = 50 µm).
DNLA reduced neural oxidative stress in AD mice with CCH
Immunohistochemical analysis revealed distinct expression of oxidative damage markers in 12-month-old APP23 mice, with 4-hydroxynonenal (4-HNE, lipid peroxidation marker) and 8-hydroxy-2'-deoxyguanosine (8-OHdG, DNA oxidation marker) prominently localized in cortical and hippocampal regions across all groups (Figure 5A, C). Quantitative evaluation demonstrated significantly elevated 4-HNE and 8-OHdG levels in CCH-treated mice compared to APP23 controls at 12 months (Figure 5B and 5D, #p < 0.05 versus APP23, ##p < 0.01 versus APP23). Notably, DNLA treatment substantially reduced oxidative damage, showing significantly lower 4-HNE and 8-OHdG levels versus untreated CCH mice at 12 months (Figure 5B and 5D, *p < 0.05 versus APP23 + CCH). These results demonstrate DNLA's potent antioxidant capacity to mitigate CCH-induced lipid and nucleic acid peroxidation in the AD mouse model.
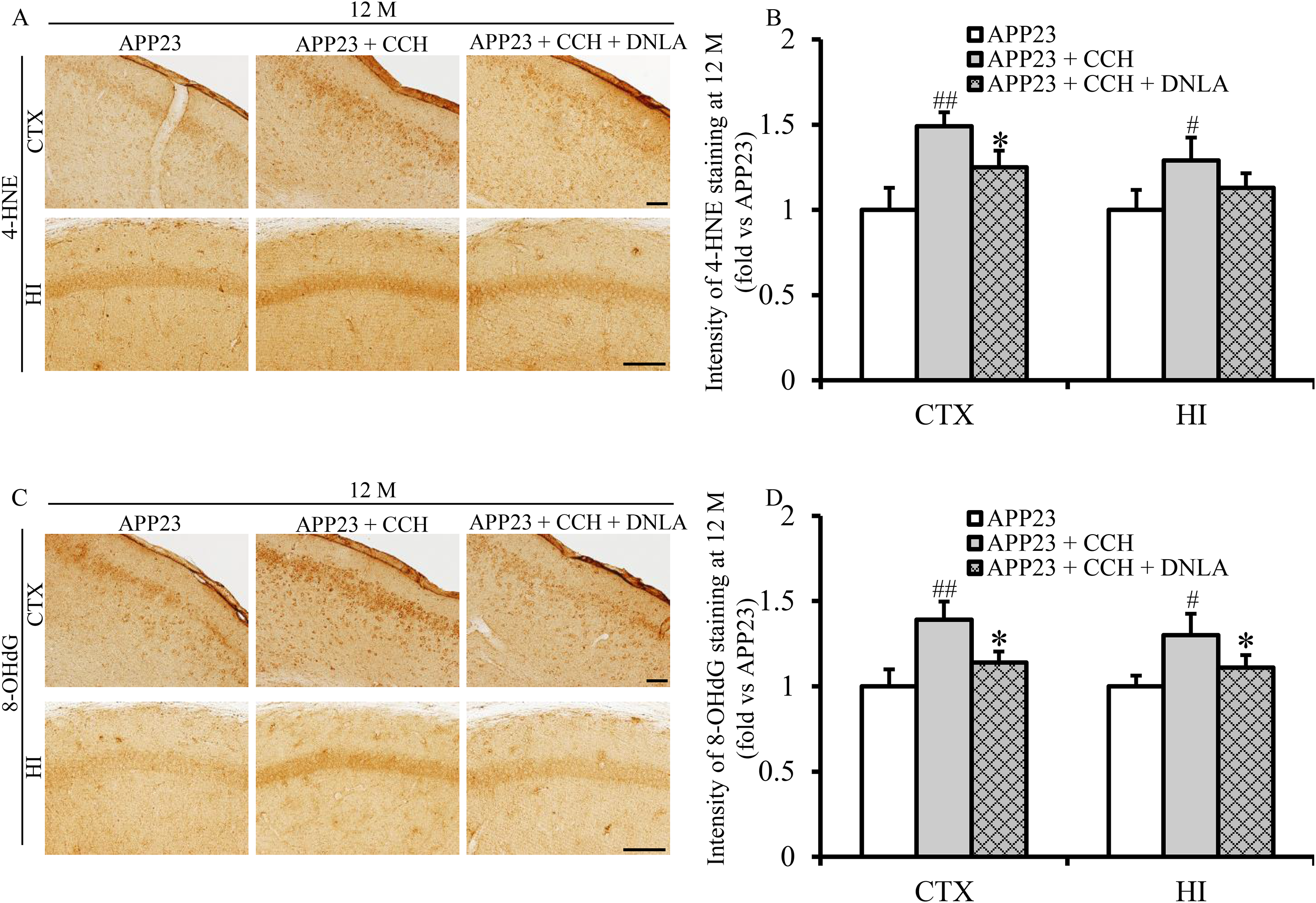
Dendrobium nobile Lindl. alkaloids (DNLA) decrease cerebral chronic hypoperfusion (CCH)-induced overexpression of 4-HNE and 8-OHdG of APP23 mice at 12 months (M) old. Representative photomicrographs of 4-HNE (A) and 8-OHdG (C) immunohistochemistry staining in the cortex (CTX) and hippocampus (HI) at 12 M. Quantitative analysis of the pixel intensity ratios of 4-HNE (B) and 8-OHdG (D)-positive signals in the CTX and HI at 12 M. Data are presented as mean ± SD. Statistical comparisons are tested by one-way ANOVA followed by a Tukey-Kramer posthoc comparison (#p < 0.05 versus APP23, ##p < 0.01 versus APP23; *p < 0.05 versus APP23 + CCH. Scale bar = 50 µm).
Discussion
CCH is commonly observed in older adults with AD. 2 Previous studies indicate that among dementia patients more than 75 years old, AD is present in 69% of cases, 22 and within this AD group, nearly 90% also exhibit signs of cerebrovascular disease. 23 CCH is thought to play a critical role in both initiating and accelerating cerebrovascular disease. 3 Therefore, CCH could be a bridge between cerebrovascular disease and AD. Potential mechanisms include promoting Aβ overproduction and deposition, 5 impairing Aβ clearance, 24 triggering neuroinflammation, 25 and exacerbating oxidative stress. 6 Moreover, CCH may not only initiate but also accelerate the AD pathogenesis (vascular hypothesis). Consequently, investigating mechanisms and developing treatments that target the complicated pathophysiology of CCH + AD is of critical importance. Recent insights indicate that therapies targeting only a single protein or pathway are ineffective against AD with CCH. 26 It is therefore essential to develop novel therapies capable of simultaneously targeting multiple key pathways involved in their shared pathogenesis.
DNLA, one main active component of Dendrobium nobile, was demonstrated to improve cognitive decline in AD experimental animal models by multiple pathways in terms of inhibition of Aβ deposits, reduction of neuroinflammation, and decrease of neural oxidative stress.27–30 However, the effects of DNLA in AD with CCH remain elusive. Our study provides the first evidence that DNLA exerts comprehensive neuroprotective effects in a novel AD with CCH mouse model, as evidenced by: (1) significant improvement in CBF (Figure 1), (2) mitigation of both motor and cognitive deficits (Figure 2), (3) reduction of Aβ oligomer and Aβ40 accumulation (Figure 3), (4) suppression of neuroinflammatory responses (Figure 4), and (5) alleviation of oxidative stress markers (Figure 5). These results position DNLA as a promising multi-target therapeutic candidate for AD complicated by cerebrovascular dysfunction, which indicates DNLA could have potential for the prevention and treatment of patients with AD plus CCH.
The CBF of 12-month-old CCH-treated AD mice significantly decreases, which is related to a great deal of Aβ40 deposition in vessels.5,6,13 In this study, DNLA remarkably improved the CBF decrease in 12-month-old CCH-treated AD (Figure 1), partially resulting from the inhibition of Aβ40 deposition accelerated by CCH (Figure 3C, D). And the improvement of CBF by DNLA could be associated with inhibited Aβ deposits, reduced oxidative stress, and decreased neuroinflammation. 6 Additionally, DNLA improved cognitive decline in AD mice, whereas the effect of DNLA on CCH-induced motor and cognitive deficits in AD animals is elusive. In the current behavioral assessments, we first observed that DNLA treatment markedly ameliorated motor and cognitive impairments in APP23 mice with CCH at 12 months of age. This improvement may be attributed to enhanced CBF, reduced levels of Aβ oligomers and Aβ40, suppression of neuroinflammation, and reduced neural oxidative stress (Figures 1, 3, 4, and 5).
Our increasing evidence suggests that CCH induces Aβ oligomer and Aβ40 accumulation in the cerebral parenchyma of 12-month-old APP23 mice, contributing to increased neuroinflammation and neural oxidative stress (Figure 3).5,6,8,13 DNLA administration has been proven to delay pathophysiological progression of Aβ in AD animal models.27–30 Consistent with these findings, our study revealed that eight-month DNLA administration significantly reduced CCH-induced Aβ oligomer and Aβ40 deposition in the cortex and hippocampus of 12-month-old APP23 mice (Figure 3), which indicates that the remarkable motor and cognitive deficits induced by CCH in 12-month-old APP23 mice were significantly improved, partly due to the inhibition of Aβ pathophysiological pathways. But specific mechanisms of the Aβ deposit decrease are still unclear. It is possibly due to inhibited neuroinflammation and oxidative stress (Figures 4 and 5) because abnormal neuroinflammation and oxidative stress can dramatically accelerate Aβ deposition.31–34 Moreover, some papers reported that DNLA decreased Aβ deposition via modulating amyloid precursor protein processing by suppressing γ-secretase activity in both in vitro and in vivo AD models.10,30 Furthermore, DNLA alleviated cerebral amyloid angiopathy in the thalamus by reducing Aβ40 deposition in arterioles and improving vascular remodeling (Figure 3C, D), suggesting improved Aβ clearance following treatment.
Neuroinflammation is involved in the pathophysiological progression of AD with CCH.5–8 Macrophages and microglia serve as key regulators in CCH-associated neuroinflammation by releasing neurotoxic cytokines and activating coagulation and complement cascades.35–37 The accumulation and activation of these inflammatory factors further induce immune reactions and act on the degeneration of neurons, which causes cognitive decline and dementia. 33 Previous papers showed that DNLA not only inhibited lipopolysaccharide (LPS)-induced microglial activation in in vitro AD models but also attenuated neuro-inflammation in AD mice and LPS-induced mice. This effect is mediated through the inhibition of glial cell activation and neurotoxic cytokine expression, likely via suppression of toll-like receptor 4 endocytosis and nuclear factor-Kb (NF-kB) signaling.38,39 Our present study revealed that DNLA treatment significantly reduced the number of Iba-1-positive microglia and GFAP-positive astrocytes, as well as the expression of pro-inflammatory cytokines (IL-1β and TNF-α), in the cortex (CTX) and hippocampus (HI) of 12-month-old CCH-treated APP23 mice (Figure 4). These results demonstrate that DNLA effectively attenuates CCH-induced neuroinflammation in this AD model.
Oxidative stress is another critical factor in the AD plus CCH pathogenesis. Recent studies have confirmed DNLA's ability to reduce oxidative stress in Aβ25−35-induced PC12 cells and liver injury animal models.40–42 This is particularly relevant as cell membranes, composed largely of oxidation-sensitive lipids, are especially vulnerable to oxidative stress damage. The peroxidation of lipids is often observed in AD with CCH diseases.5–7 Extensive evidence supports DNLA's potent antioxidant properties, including its capacity to scavenge reactive oxygen species (ROS) in various disease models.10,43,44 Our current findings demonstrate that DNLA treatment significantly suppressed lipid peroxidation in 12-month-old APP23 mice with CCH, particularly by reducing levels of 4-hydroxynonenal (4-HNE), a terminal product of lipid peroxidation, in the cortex. This protective effect on membrane lipids likely contributes to cellular homeostasis by preserving membrane stability and permeability in the CCH-affected brain. Furthermore, we observed that DNLA administration markedly decreased 8-hydroxy-2'-deoxyguanosine (8-OHdG), a biomarker of oxidative DNA damage typically elevated in cells undergoing ROS-induced degeneration. 45 This finding aligns with previous reports of elevated 8-OHdG expression in AD with CCH models.8,14,37 The significant reduction of 8-OHdG in both cortex and hippocampus of DNLA-treated mice demonstrates its protective effect against nucleic acid oxidation in this combined pathology.
At the end of the discussion, we would like to clarify the limitations of the present study. 1) As for behavioral tests, we only conducted rotarod and 8-arm experiments for testing the motor ability and working memory of mice. The limited number of behavioral tests is insufficient to fully reflect the cognitive changes of experimental mice. More indexes reflecting cognitive abilities, such as episodic, spatial memory, social memory, and cognitive flexibility, need to be tested by multiple experiments in our future research. 2) More potential therapeutic targets of DNLA require further exploration, such as the tau protein pathway, Aβ dynamics (production and clearance), the neurovascular units, and disturbances in cerebral glycolysis and metabolism. 3) Based on the present findings, further experiments are required to investigate the deeper and more precise underlying mechanisms, such as the activation or inhibition of astrocytes and microglia (M1 or M2). 4) The therapeutic benefits of DNLA observed in this mouse model should be further tested in larger animal species, such as non-human primates or pigs that exhibit better neuroanatomical and physiological similarity to humans than rodents. Such studies will be critical for a more accurate translation of these preclinical findings.
Conclusions
In summary, the present study first reports that DNLA improved motor and cognitive declines in APP23 plus CCH mice at 12 months via CBF amelioration, inhibition of Aβ oligomer and Aβ40 expression, neuro-inflammation reduction, and decrease of neural oxidative stress. Together, DNLA improves motor and cognitive deficits via multiple neuroprotective pathways in AD plus CCH mice and could be a potential candidate in the therapy for AD plus CCH patients.
Footnotes
Acknowledgements
We thank all co-authors for their essential contributions to the design and execution of experiments, data analysis, and critical review and editing of the manuscript. And we also thank Dr Yan Zhu (The Emergency Center of the First Hospital of Qiqihar City, China) for her excellent technical assistance with respect to the Nissl staining experiment during manuscript revision. Their collaborative efforts were fundamental to the completion of this study.
Ethical considerations
All animal experiments were conducted in accordance with the guidelines of the Animal Committee of Guizhou Medical University (Approval No. GMU#2303239), the ARRIVE guidelines (![]() . nc3rs.org.uk), and institutional regulations for the Care and Use of Laboratory Animals. All efforts were made to minimize animal suffering and reduce the number of animals used.
. nc3rs.org.uk), and institutional regulations for the Care and Use of Laboratory Animals. All efforts were made to minimize animal suffering and reduce the number of animals used.
Consent to participate
Not applicable
Consent for publication
Not applicable
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study received funding support from the following sources: Science and Technology Planning Project of Guizhou Province (grant number: ZK[2024] general project 183), the Science and Technology Foundation of Guizhou Provincial Health Commission (grant number: gzwkj2023-274), and High-level Talents of Guizhou Medical University Scientific Research Start-up Fund Project (grant number: Xiao Bo He J Zi [2022] No. 006).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The data supporting the findings of this study are available on request from the corresponding author.