
Abstract
Background
The extent to which plasma biomarkers of Alzheimer's disease and neurodegeneration capture domain-specific cognitive performance across diverse populations remains unclear.
Objective
To determine whether plasma phosphorylated tau181 (p-tau181) and neurofilament light (NfL) are independently associated with cognitive domains, and whether associations differ across non-Hispanic White (NHW), non-Hispanic Black (NHB), and Hispanic participants.
Methods
We analyzed 3023 community-dwelling older adults from the Health and Aging Brain Study–Health Disparities (38.4% NHW, 22.6% NHW, 38.9% Hispanic). We used linear regressions to test associations between plasma biomarkers and cognitive domains (memory, executive function, processing speed, language), adjusting for age, sex, education, and apolipoprotein ε4 carriership. We fit models including both p-tau181 and NfL to assess their independent associations, evaluate biomarker × racial/ethnic interactions, and test p-tau181 × NfL interactions within each racial/ethnic group.
Results
Among NHW participants, higher p-tau181 and NfL were associated with poorer memory, executive function, processing speed, and language. In NHB participants, p-tau181 was associated with memory, showed attenuated associations for language, and demonstrated similar associations with executive function and processing speed as observed in NHW participants. In Hispanic participants, p-tau181 was associated with memory and processing speed but was nonsignificant for executive function and language, and NfL showed significant but attenuated associations across all domains. Higher p-tau181 and NfL were jointly associated with slower processing speed only in NHW and NHB participants.
Conclusions
Plasma p-tau181 and NfL were associated with multiple cognitive domains, with the strongest effects in NHW participants and attenuated associations in NHB and Hispanic individuals.
Keywords
Introduction
Alzheimer's disease (AD) and AD-related dementias (ADRD) represent major public health challenges, with AD ranking as the sixth leading cause of death in the United States and dementia prevalence expected to rise substantially over the coming decades. 1 Despite recently approved amyloid clearing therapeutics2–5 and an approved blood test for monitoring AD, 6 there is still an urgent need to improve early identification of individuals at risk and to clarify biological pathways underlying disease development. Traditionally, diagnosis and staging of AD have relied on invasive cerebrospinal fluid (CSF) assays and costly positron emission tomography (PET) imaging. 7 Recent revisions to the Alzheimer's Association criteria have further emphasized a biological definition of AD, centering on the AT(N) framework that characterizes amyloid-β deposition (A), tau neurofibrillary pathology (T), and neurodegeneration (N). 8 This framework has since been expanded to incorporate markers of inflammation (I), notably glial fibrillary acidic protein (GFAP).8,9 Corresponding plasma biomarkers including Aβ42/40 ratio (A), phosphorylated tau species (p-tau181, p-tau217, p-tau231) (T), neurofilament light chain (NfL) (N), and GFAP (I) are now measurable in blood, providing scalable, non-invasive alternatives to CSF or PET.10–12
Phosphorylated tau species are the most specific indicators of AD pathology, with p-tau217 and p-tau181 showing strong diagnostic performance in recent head-to-head comparisons and meta-analyses.13,14 NfL, by contrast, is a nonspecific marker of neuroaxonal injury that rises with aging, traumatic brain injury, and multiple neurodegenerative diseases.15,16 Elevated NfL has been linked to structural brain changes, white matter hyperintensity burden, and incident dementia in large community cohorts.17–19 Together, these markers represent distinct but overlapping biological pathways including proteinopathy, neurodegeneration, and neuroinflammation that may differentially contribute to cognitive impairments across different domains.
Despite rapid advances in the biological staging,8,20 tracking the preclinical/prodromal stages of pathology accumulation prior to cognitive impairments, 21 and advancements in blood-based biomarkers, several critical gaps remain. First, the majority of biomarker validation studies have been conducted in populations of predominantly European ancestry, raising questions about generalizability to diverse populations.22,23 Second, although biomarkers such as p-tau181 and p-tau217 are established correlates of global cognition, disease progression, and dementia less is known about their specificity to individual cognitive domains. 24 Early evidence suggests that p-tau may be particularly linked to memory performance, consistent with its role in AD pathology, whereas NfL appears to capture more generalized age-related decline.25,26 However, most studies related to blood-based biomarkers to date have been limited by small sample sizes, cross-sectional designs, or reliance on global composite measures of cognition rather than harmonized domain-specific scores.10,27–33
Clarifying whether blood-based biomarkers are independently associated with domain-specific cognitive performance, 34 and whether these associations differ across racially and ethnically diverse populations, is critical for advancing both mechanistic understanding and clinical translation. Moreover, evaluating biomarkers jointly is necessary to determine whether disease-specific markers such as p-tau181 retain associations with cognition when considered alongside nonspecific indicators like NfL. Using the Health and Aging Brain Study–Health Disparities (HABS-HD) cohort, we sought to determine whether plasma blood-based biomarkers (p-tau181, NfL, total tau, and Aβ42/40 ratio) were associated with cognitive domain-specific associations (memory, executive function, language, and processing speed) cross-sectionally. We hypothesized p-tau181, a marker of AD pathology, would be specifically associated with memory performance, the domain most closely linked to AD. In contrast, we expected higher NfL concentrations, a nonspecific marker of age-related neurodegeneration, to be broadly associated with poorer performance across multiple domains. Importantly, we evaluated whether the association of p-tau181 with cognitive domain outcomes was independent of NfL, and whether these biomarker–cognition relationships were consistent across Non-Hispanic White (NHW), Non-Hispanic Black (NHB), and Hispanic participants.
Methods
Study design
The Health and Aging Brain Study – Health Disparities (HABS-HD), previously known as the Health & Aging Brain Among Latino Elders (HABLE) study, is an ongoing longitudinal cohort based at the University of North Texas Health Science Center's Institute for Translational Research. 35 HABS-HD represents the largest community-based study of AD that includes NHW, NHB, and Hispanic participants. The study was specifically designed to evaluate the AT(N) framework (amyloid, tau, and neurodegeneration) while addressing racial and ethnic disparities in AD research. 35 Participants undergo extensive assessments, including structured demographic and health questionnaires, comprehensive neuropsychological testing, blood draws for laboratory and biomarker analyses, brain magnetic resonance imaging (MRI), amyloid and tau PET, and functional examinations. 35 All HABS-HD protocols were approved by the University of North Texas Health Science Center Institutional Review Board, and all participants provided written informed consent in accordance with the Declaration of Helsinki. This study used data release 7 from HABS-HD.
Demographics
Participants responded to demographic questionnaires reported their age at the visit, gender/sex (male or female as option), years of formal education, and ethnicity with their options being white, black, or Hispanic (referred to as NHW, NHB, Hispanic).
APOE genotyping
Participant's had their DNA extracted and analyzed using TaqMan SNP Genotyping Assays for rs429158 and rs7412, in conjunction with the GTXpress Master Mix (ThermoFisher Scientific) to determine their apolipoprotein E (APOE) genotype.35,36 For analyses, participants were classified into two groups: carriers of at least one ε4 allele (ε4+) and non-carriers (ε4–). Although APOE ε4 exerts a strong dose-dependent effect on AD risk in NHW and European population,37,38 the binary grouping increases statistical power and interpretability.
Blood-based biomarkers related to ATN framework
Fasting blood samples were collected, processed, and stored following established international guidelines. Assay preparation was carried out using the automated StarPlus system (Hamilton Robotics, Reno, NV). Concentrations of plasma biomarkers including the p-tau181, NfL, total tau, and Aβ42/40 ratio were quantified with the highly sensitive single molecule array (Simoa) technology on the HD-X platform (Quanterix, Billerica, MA), following the manufacturer's specifications. For all assays, the coefficients of variation did not exceed 5%. Several studies have documented the use of these blood based biomarkers and their methods in HABS-HD.39–43
Plasma biomarker quantification and harmonization
Plasma biomarkers were quantified using multiple assay releases (releases 2, 3, 5, and 7) which all used the same Quanterix platform. The analytes of interest included p-tau181, NfL, total tau, Aβ40, and Aβ42. To optimize sample size while maintaining reproducibility, biomarker concentrations were harmonized by averaging all available release-specific values per participant. This approach assumes that small inter-release differences reflect random technical variation rather than systematic. A quality-control comparison of this averaging approach versus selecting the most recent non-missing measurement demonstrated high inter-method correlations but small mean differences for select biomarkers (e.g., Aβ42, total tau), consistent with minor calibration shifts across assay lots. Because these differences were directionally consistent and of small magnitude, averaged values were retained for all primary analyses. Moreover, the results were identical for the proposed analyses when using the most recent non-missing measurement. Extreme outliers exceeding five standard deviations (±5 SD) from the mean were coded as missing prior to analyses. To facilitate comparability across analytes, each biomarker was standardized using z-scores, and the Aβ42/40 ratio was computed and similarly z-transformed. 44 Analyses were restricted to complete cases with non-missing values for all four biomarkers.
Cognitive diagnosis and consensus review
Cognitive status was categorized as cognitively unimpaired (normal), mild cognitive impairment (MCI), or dementia using a structured diagnostic algorithm followed by expert consensus review. The HABS-HD procedures for cognitive diagnosis have been described previously and follow established diagnostic standards.45,46 Briefly, all participants meeting algorithmic criteria for mild cognitive impairment (MCI) or dementia, along with a 10% random sample of those classified as cognitively unimpaired, underwent consensus evaluations. Each case was independently reviewed by a multidisciplinary panel of dementia specialists, medical professionals, and study clinicians, integrating neuropsychological performance, clinical history, and supporting documentation.
Neuropsychological exams and harmonized cognitive domain scores
All neuropsychological measures were first reviewed for plausibility and completeness. Unavailable data were recoded to account for missingness. Following previous HABS-HD methodology, 47 domain-specific assessments included the Spanish–English Verbal Learning Test immediate recall and delayed recall trials, Logical Memory Story A and Story B from the Wechsler Memory Scale, Digit Span forward and backward, the Digit Symbol Substitution subtest, Trail Making Test Parts A and B, animal naming fluency, and phonemic (F-A-S) fluency. For each individual neuropsychological cognitive test, raw scores were (except the Trail Making times) converted to a z-score across the full analytic sample. Because shorter completion times on the Trail Making Test indicate better performance, we first standardized the Part A and Part B completion times to z-scores and then inverted them by multiplying by −1, so that a higher z-score reflects better cognitive function. We derived four harmonized composite scores by averaging the relevant z-scores for each participant. The episodic memory composite combined the z-scores of verbal learning immediate recall, verbal learning delayed recall, Logical Memory Story A, and Logical Memory Story B. The executive function composite averaged the z-scores of Digit Span and Digit Symbol Substitution. The language composite comprised z-scores from the animal naming and phonemic fluency tests. The processing speed composite averaged the inverted Trail Making Part A and Part B z-scores.
Statistical analyses
All analyses were conducted on the outlier-cleaned, complete-case dataset.
Overview
Descriptive statistics were used to summarize demographic and clinical characteristics; no formal group comparisons were performed for sample characteristics.
Group differences in biomarkers and cognition
Group differences in plasma biomarkers and cognitive domain scores were evaluated using analysis of covariance (ANCOVA). For biomarker models, race/ethnicity and cognitive status were entered as fixed factors, adjusting for age, sex, years of education, and APOE ε4 carrier status. Post hoc pairwise comparisons were performed using Tukey's honest significant difference correction. Adjusted mean ± SE z-scores were extracted from the models for visualization of race/ethnic differences in memory, executive function, language, and processing speed.
Biomarker associations with cognitive domains
Multivariable linear regression models were estimated separately for each plasma biomarker (p-tau181, NfL, total tau, and Aβ42/40 ratio) as predictors of domain-specific cognitive z-scores (memory, executive function, language, and processing speed). All models were adjusted for age, sex, education, and APOE ε4 carrier status. β coefficients (standardized given that the biomarkers and cognitive domains are both z-scored) and 95% confidence intervals (CI) were reported.
Combined p-tau181 + NfL models (overall sample)
To evaluate the independent and joint effects of amyloid/tau and neuronal injury, combined models including both p-tau181 and NfL were fit for each cognitive domain, controlling for age, sex, education, and APOE ε4. Nested model comparisons using ANOVA F-tests were conducted to compare single- versus dual-biomarker models to quantify improvement in model fit when both biomarkers were included. A sensitivity analyses was run in the combined models and were repeated with additional adjustment for clinical cognitive status (normal, MCI, dementia) to assess whether the independent effects of p-tau181 and NfL were robust.
P-tau181* race/ethnicity and NfL* race/ethnicity interaction models
To test whether associations between plasma biomarkers and cognition differed by race/ethnicity, we estimated interaction models that included two interaction terms (p-tau181 × racial ethnicity and NfL × racial ethnicity). Models were adjusted for age, sex, education, and APOE ε4 status.
Racial ethnic stratified models for combined p-tau181 + NfL models
Stratified regression models were conducted within each racial/ethnic group (NHW, NHB, Hispanic), including both p-tau181 and NfL as simultaneous predictors of domain-specific cognitive performance. All models were adjusted for age, sex, education, and APOE ε4 carrier status. Race-stratified models were repeated with further adjustment for clinical cognitive status to determine whether biomarker–cognition associations within each racial ethnic group were independent of cognitive impairment severity.
Racial ethnic stratified models for interactions of p-tau181 + NfL models
Racial ethnic-stratified linear regression models were estimated to examine whether associations between p-tau181 and cognitive performance varied as a function of NfL. Separate models were fit for memory, executive function, processing speed, and language, including a p-tau181 × NfL interaction term and adjusting for age, sex, education, and APOE ε4 status. A negative interaction indicated that higher levels of one biomarker strengthened the adverse association of the other with cognition. Sensitivity analyses further adjusted for clinical cognitive diagnosis to assess robustness.
Statistical analyses were conducted in R v4.2.3. Two-sided p-values < 0.05 were considered statistically significant.
Results
Sample characteristics
Table 1 summarizes demographic, genetic, clinical, and biomarker characteristics of the HABS-HD cohort stratified by racial/ethnic group. The analytic sample included 3023 participants from HABS-HD (1162 NHW, 684 NHB, 1177 Hispanic) with a mean age of 65.4 years (SD = 8.8). NHB participants were significantly older on average (69.0 ± 8.8 years) than NHB and Hispanic participants (both 63.2 ± 8.0 years). Females comprised 62.2% of the cohort. APOE ε4 carriage was most prevalent among NHB participants (40.5%) and least common among Hispanic participants (19.8%). Clinical cognitive status also varied: normal cognition was most frequent among NHB participants (80.6%) and least among NHB participants (62.1%), who exhibited the highest proportions of MCI (30.4%) and dementia (7.5%).
Participant characteristics stratified by racial ethnicity in HABS-HD.
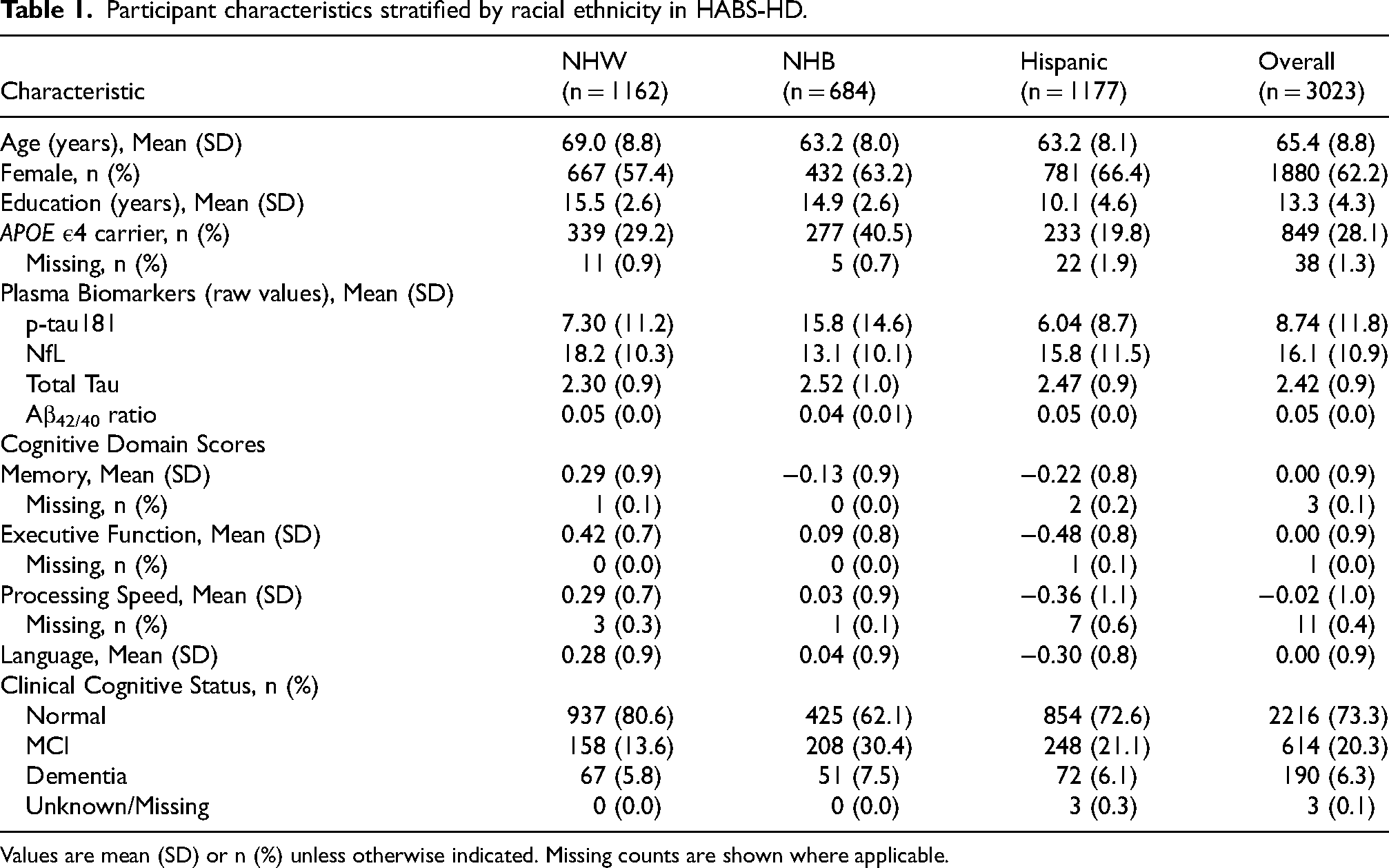
Values are mean (SD) or n (%) unless otherwise indicated. Missing counts are shown where applicable.
Racial/ethnic differences in plasma biomarkers
As shown in Figure 1A, plasma biomarker concentrations varied significantly by race/ethnicity after adjustment for age, sex, education, and APOE ε4 status. NHB participants exhibited the highest p-tau181 and NfL levels, followed by Hispanic and NHW participants (all p < 0.001). In contrast, NHW participants showed significantly higher plasma Aβ42/40 ratios than NHB and Hispanic adults (p < 0.001). Total tau concentrations were modestly higher in NHB and Hispanic, compared with NHW participants (p < 0.01). Across the sample, plasma p-tau181, NfL, and total tau increased progressively from cognitively normal to MCI and dementia groups (Figure 1B). Adjusted ANCOVA models indicated significant stepwise elevations for each biomarker across clinical groups (all p < 0.001). In contrast, Aβ42/40 ratios declined with worsening cognitive status (p < 0.01), reflecting increasing amyloid pathology. Adjusted cognitive domain analyses (Figure 1C) revealed that NHB and Hispanic participants performed significantly lower than NHW participants across almost all domains (memory, executive function, language, and processing speed; all p < 0.001). The cognitive domains findings mimic visual representations of any cognitive impairment versus normal cognition across racial ethnic groups with NHB having the highest prevalence of impairment (Supplemental Figure 1). Among NHW participants, 19.3% were cognitively impaired compared with 37.9% of NHB and 27.2% of Hispanic participants. Conversely, the proportion classified as cognitively normal was highest in the NHW group (80.7%), followed by Hispanic (72.8%) and NHB (62.1%) participants (Supplemental Figure 1).
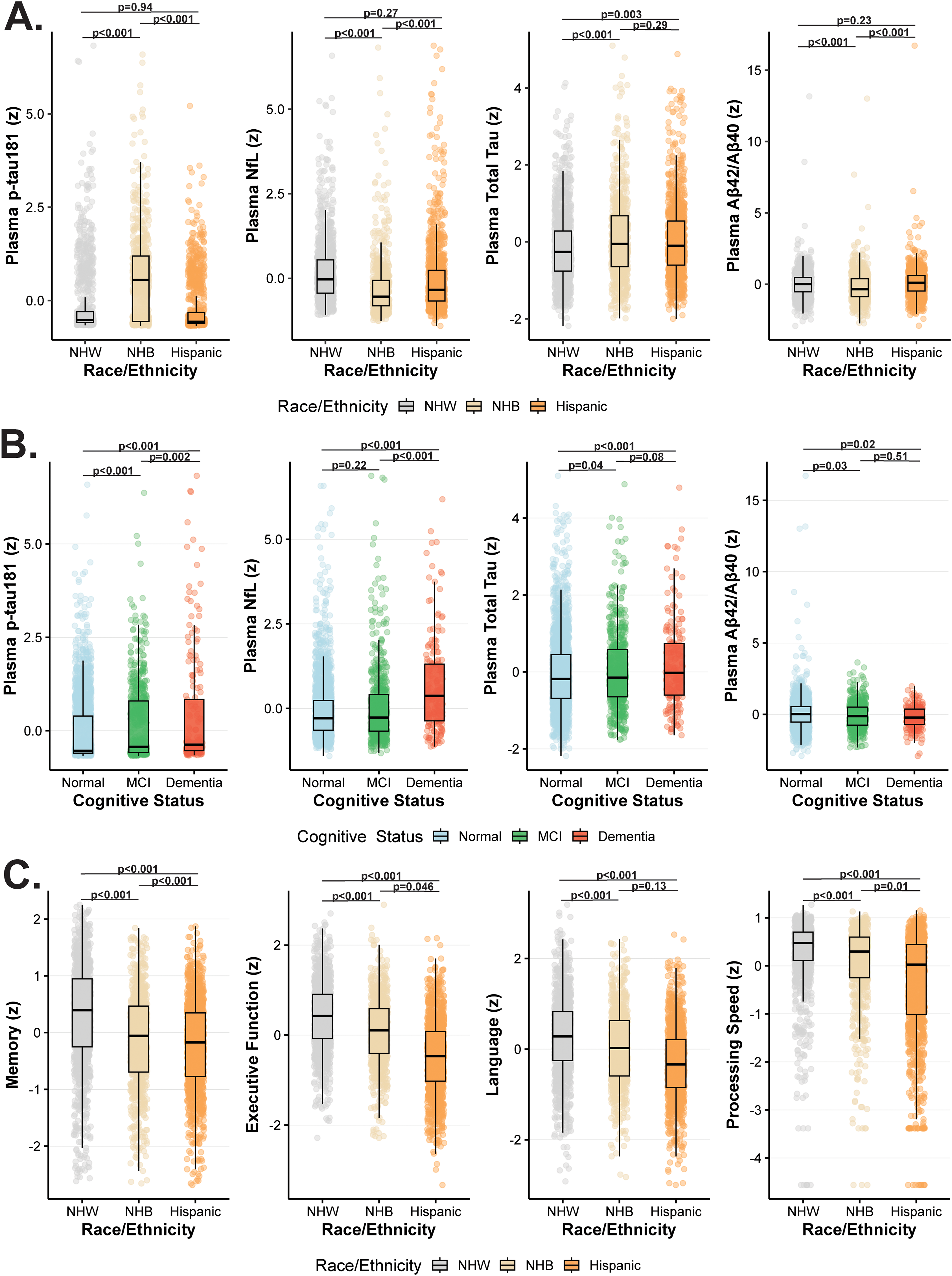
Racial/ethnic and cognitive differences in plasma biomarkers and cognitive performance. (A) Plasma biomarker levels (z scores) for p-tau181, NfL, total tau, and Aβ42/40 ratio are shown across racial/ethnic groups (Non-Hispanic White [NHW], Non-Hispanic Black [NHB], and Hispanic). (B) Plasma biomarker levels (z scores) are displayed across clinical cognitive groups (normal cognition, mild cognitive impairment [MCI], and dementia). p-tau181, NfL, and total tau increased progressively with worsening cognitive status, whereas Aβ42/40 ratios declined. (C) Adjusted mean (±SE) cognitive domain z scores for memory, executive function, language, and processing speed are shown by race/ethnicity. Models were adjusted for age, sex, education, and APOE ε4 status. The adjusted p values from ANCOVA models indicate significant group differences for each panel. p-tau181: plasma phosphorylated tau 181; NfL: neurofilament light chain; Aβ42/40: amyloid-β 42 to 40 ratio; NHW: Non-Hispanic White; NHB: Non-Hispanic Black.
Cross-sectional associations between plasma biomarkers and cognitive domains
Across all models, plasma biomarkers reflecting amyloid/tau- and neurodegeneration-related pathology were significantly associated with worse cognitive performance, whereas higher amyloid-related ratios were linked to better cognition across domains (Figure 2A-D). In fully adjusted linear regression models (adjusting for age, sex, education, race/ethnicity, and APOE ε4 carrier status), higher plasma p-tau181 concentrations were significantly associated with poorer performance across all cognitive domains (Figure 2A), including memory (β = −0.17, 95% CI −0.20 to −0.14; p < 0.01), executive function (β = −0.07, 95% CI −0.10 to −0.05; p < 0.01), processing speed (β = −0.09, 95% CI −0.12 to −0.06; p < 0.01), and language (β = −0.08, 95% CI −0.11 to −0.05; p < 0.01). Similarly, elevated plasma NfL was consistently associated with worse performance in memory (β = −0.12, 95% CI −0.15 to −0.09; p < 0.01), executive function (β = −0.10, 95% CI −0.13 to −0.08; p < 0.01), processing speed (β = −0.15, 95% CI −0.18 to −0.12; p < 0.01), and language (β = −0.12, 95% CI −0.15 to −0.09; p < 0.01) (Figure 2B). Total tau showed similar associations across all domains (Figure 2C). Higher plasma Aβ42/40 ratios were associated with better cognitive performance across all domains (Figure 2D).
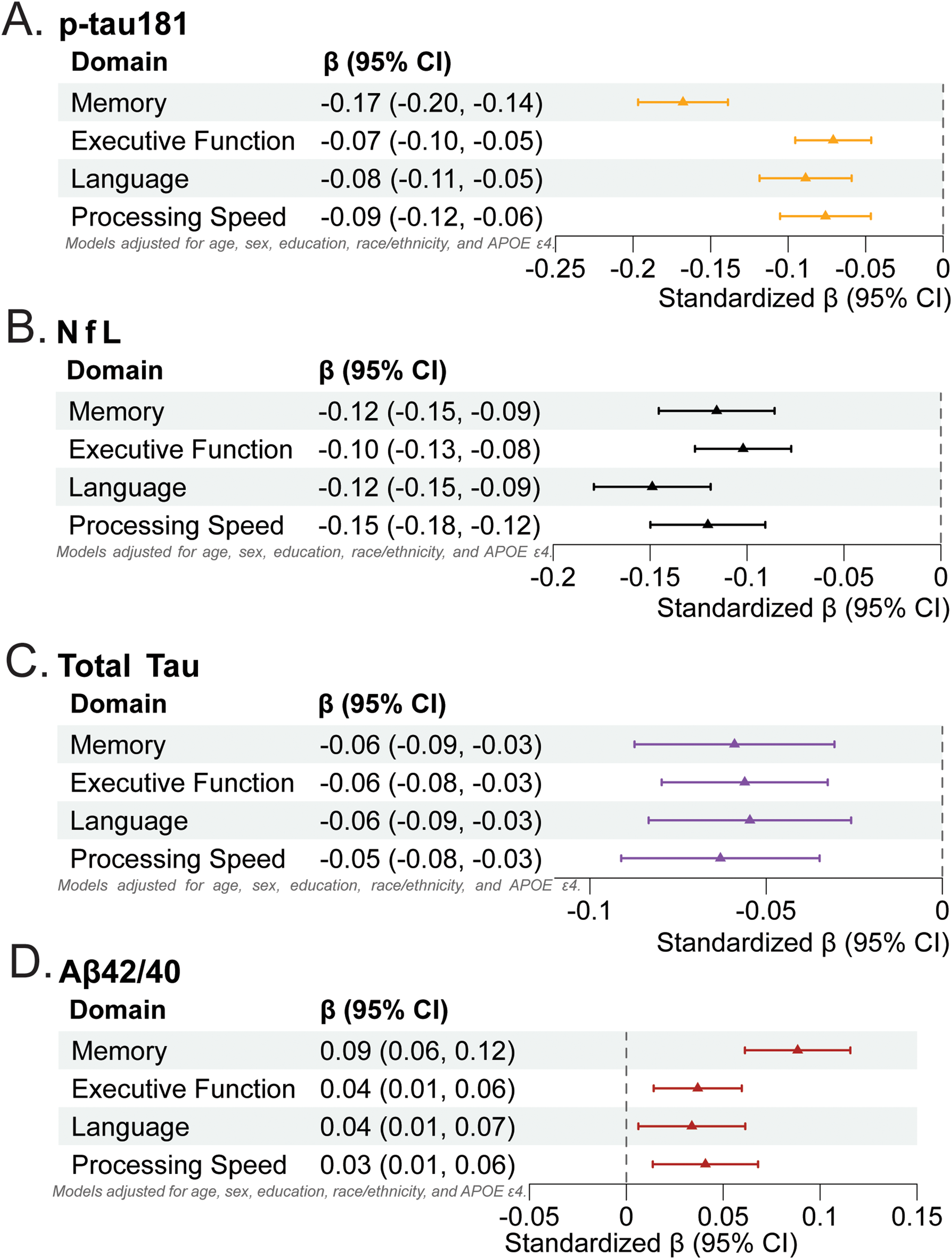
Cross-sectional associations between plasma biomarkers and cognitive domains. Each panel displays standardized β coefficients (95% CIs) from linear regression models examining associations between individual plasma biomarkers and harmonized cognitive domain z-scores. Standardized β coefficients (95% CIs) denote the magnitude and direction of association; negative coefficients indicate poorer cognitive performance with higher biomarker concentrations, with the exception of positive coefficients for Aβ42/40 reflect better cognition with higher ratios. All models were adjusted for age, sex, education, race/ethnicity, and APOE ε4 carrier status. (A) p-tau181; (B) Neurofilament light (NfL); (C) Total Tau; (D) Aβ42/40 ratio. Aβ: amyloid-β; NfL: neurofilament light; CI: confidence interval.
Independent associations of p-tau181 and NfL with cognitive domains
In models including both biomarkers, higher plasma p-tau181 and NfL concentrations were independently associated with poorer cognitive performance across multiple domains (Figure 3). P-tau181 was significantly associated with worse memory (β = −0.15, 95% CI −0.18 to −0.13; p < 0.001), executive function (β = −0.06, 95% CI −0.08 to −0.03; p < 0.001), processing speed (β = −0.07, 95% CI −0.10 to −0.04; p < 0.001), and language (β = −0.06, 95% CI −0.09 to −0.03; p < 0.001). Similarly, higher plasma NfL was associated with poorer performance in memory (β = −0.09, 95% CI −0.12 to −0.06; p < 0.001), executive function (β = −0.09, 95% CI −0.12 to −0.07; p < 0.001), processing speed (β = −0.14, 95% CI −0.17 to −0.11; p < 0.001), and language (β = −0.11, 95% CI −0.14 to −0.08; p < 0.001).
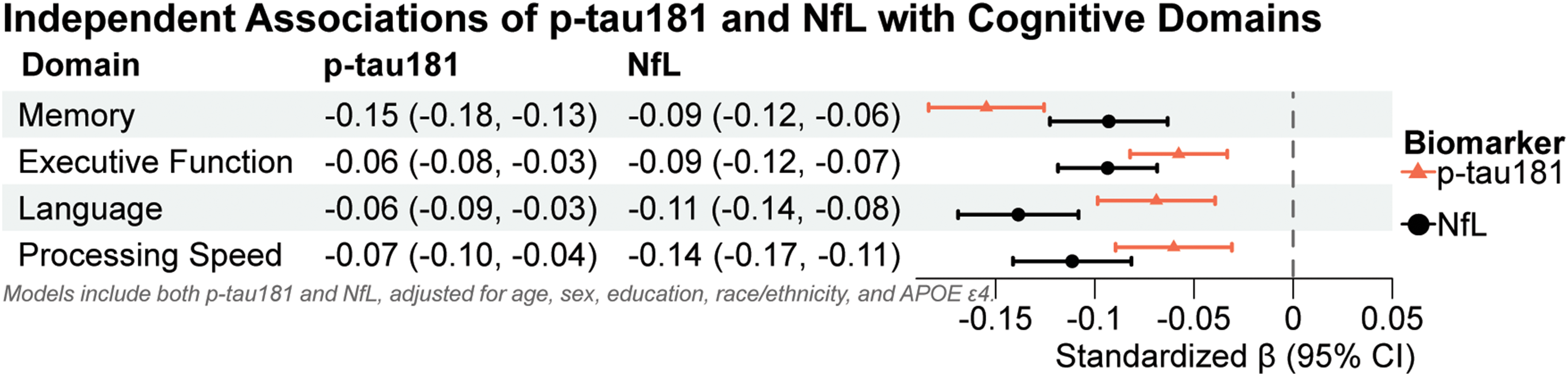
Independent associations of p-tau181 and NfL with cognitive domains. Forest plot showing standardized β estimates (95% CIs) for p-tau181 and NfL from linear regression models adjusted for age, sex, education, race/ethnicity, and APOE ε4. Both biomarkers were entered simultaneously to assess their independent associations with cognitive performance across domains. Triangles represent p-tau181; circles represent NfL. Negative β values indicate poorer cognitive performance with higher biomarker levels.
To further evaluate the incremental contribution of NfL beyond p-tau181, nested model comparisons were performed (Supplemental Table 1). Adding NfL significantly improved model fit for all cognitive domains (all p < 0.001), with the greatest improvement observed for processing speed (F = 80.06) followed by executive function (F = 53.82), language (F = 53.45), and memory (F = 37.74). Model comparisons indicate that while both biomarkers are related to cognitive performance, NfL explains additional, independent variance beyond p-tau181.
When additionally controlling for clinical cognitive diagnosis, plasma p-tau181 remained associated with memory and executive function while NfL remained significantly associated with cognitive domains previously highlighted though the effect sizes were modestly attenuated (Supplemental Table 2).
Racial ethnic × biomarker interaction models
In models including both plasma p-tau181 and NfL with race/ethnicity interaction terms, significant biomarker × race effects were observed across cognitive domains, indicating racial/ethnic modification of biomarker–cognition associations (Supplemental Table 3). Interaction terms revealed that the negative association between p-tau181 and memory was significantly attenuated among NHB participants (p-tau181 × NHB: β = 0.08, 95% CI 0.01 to 0.15; p = 0.02), while the p-tau181 × Hispanic interaction was significant for language (β = 0.09, 95% CI 0.01 to 0.16; p = 0.03). For NfL, the NHB × NfL interaction reached significance for memory (β = 0.09, 95% CI 0.01 to 0.16; p = 0.03), and the Hispanic × NfL interaction was significant for memory (β = 0.12, 95% CI 0.05 to 0.18; p < 0.01), executive function (β = 0.06, 95% CI 0.01 to 0.12; p = 0.01), and language (β = 0.11, 95% CI 0.05 to 0.17; p < 0.01).
Independent associations of p-tau181 and NfL with cognitive domain z-scores across racial/ethnic groups
Figure 4 summarizes race-stratified associations between plasma p-tau181 and domain-specific cognitive performance after controlling for NfL, adjusted for age, sex, education, and APOE ε4 status. The associations between p-tau181 were more strongly associated with lower cognition across all domains with effects greater than those observed in the NHB and Hispanic groups. Among NHW participants, both p-tau181 (β = −0.19, 95% CI −0.24 to −0.14; p < 0.01) and NfL (β = −0.16, 95% CI −0.21 to −0.10; p < 0.01) showed strong and uniform negative associations across all cognitive domains. In NHB participants, p-tau181 was significantly associated with memory (β = −0.12, 95% CI −0.16 to −0.07; p < 0.01), executive function (β = −0.05, 95% CI −0.10 to −0.01; p = 0.01), and processing speed (β = −0.06, 95% CI −0.11 to −0.01; p = 0.01), but not language (p = 0.25). NfL demonstrated its strongest association with processing speed (β = −0.20, 95% CI −0.26 to −0.13; p < 0.01) and remained significant for executive function and language. Among Hispanic participants, p-tau181 was associated with poorer memory (β = −0.14, 95% CI −0.20 to −0.09; p < 0.01) and processing speed (β = −0.09, 95% CI −0.15 to −0.02; p = 0.01), whereas NfL exhibited consistent negative associations across all domains (all p < 0.01).
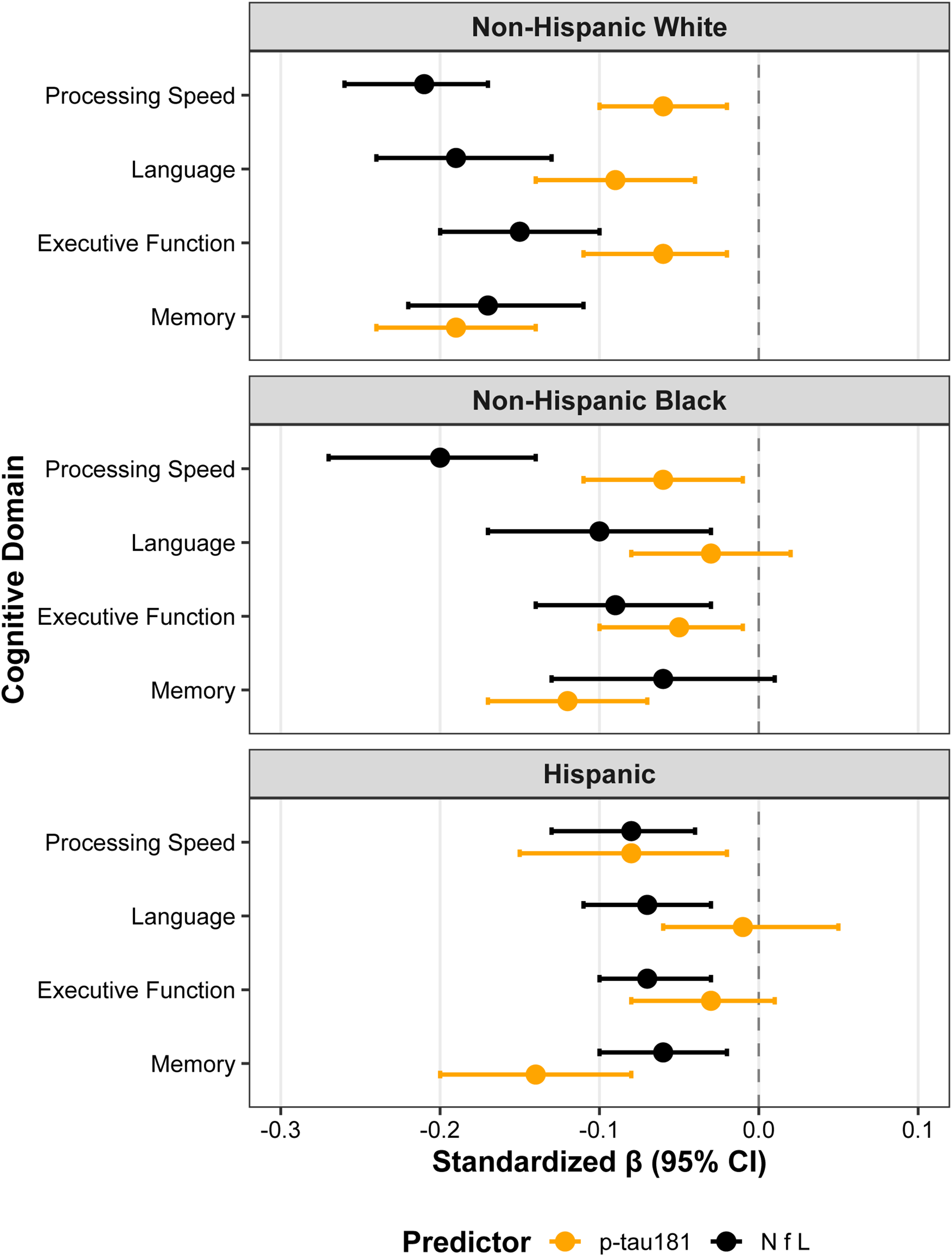
Independent associations of p-tau181 and NfL with cognitive domain z-scores across racial/ethnic groups. Forest plots display standardized β estimates (95% CIs) for p-tau181 (orange) and NfL (black) from race-stratified linear regression models adjusted for age, sex, education, and APOE ε4 carrier status. Panels represent (top) Non-Hispanic White, (middle) Non-Hispanic Black, and (bottom) Hispanic participants. Both biomarkers were entered simultaneously to assess their independent associations with cognitive performance across memory, executive function, processing speed, and language domains. Negative β coefficients indicate worse cognitive performance with higher biomarker concentrations.
To evaluate the robustness of associations between plasma biomarkers and cognitive performance, sensitivity analyses were conducted with additional adjustment for clinical cognitive status (Supplemental Table 4). Among NHW participants, higher p-tau181 concentrations remained significantly associated with poorer memory performance (β = −0.10, 95% CI −0.14 to −0.06; p < 0.01), while associations with executive function, processing speed, and language were attenuated and no longer significant. NfL, however, retained significant associations with executive function (β = −0.07, 95% CI −0.11 to −0.03; p < 0.01), processing speed (β = −0.10, 95% CI −0.14 to −0.06; p < 0.01), and language (β = −0.10, 95% CI −0.15 to −0.05; p < 0.01). Among NHB participants, p-tau181 remained significantly associated with memory (β = −0.11, 95% CI −0.15 to −0.07; p < 0.01) and processing speed (β = −0.04, 95% CI −0.08 to 0.00; p = 0.04), while NfL was associated with executive function (β = −0.05, 95% CI −0.10 to 0.00; p = 0.03) and processing speed (β = −0.15, 95% CI −0.20 to −0.10; p < 0.01), with a marginal association for language (p = 0.05). Among Hispanic participants, p-tau181 remained associated only with poorer memory (β = −0.09, 95% CI −0.13 to −0.04; p < 0.01), while NfL exhibited weaker but consistent associations with executive function (β = −0.04, 95% CI −0.07 to −0.01; p = 0.01), processing speed (β = −0.04, 95% CI −0.09 to 0.00; p = 0.04), and language (β = −0.04, 95% CI −0.08 to −0.01; p = 0.02).
Race/ethnicity-stratified interactions between plasma p-tau181 and NfL on cognition
Across racial/ethnic groups, there was a consistent trend toward synergistic effects of p-tau181 and NfL on cognitive performance, particularly for processing speed (Figure 5A-C, Supplemental Table 5). Among NHW participants, the p-tau181 × NfL interaction was significantly associated with slower processing speed (β = −0.03, 95% CI −0.06 to −0.00; p = 0.04), whereas associations for other cognitive domains were nonsignificant. Similar interaction patterns were observed among NHB participants for processing speed (β = −0.05, 95% CI −0.09 to −0.01; p = 0.01) but not for any other cognitive domains. No significant interactions were detected among Hispanic participants across any domain. In sensitivity analyses controlling for clinical cognitive status, these significant interaction effects between p-tau181 and NfL were attenuated (Supplemental Table 6).
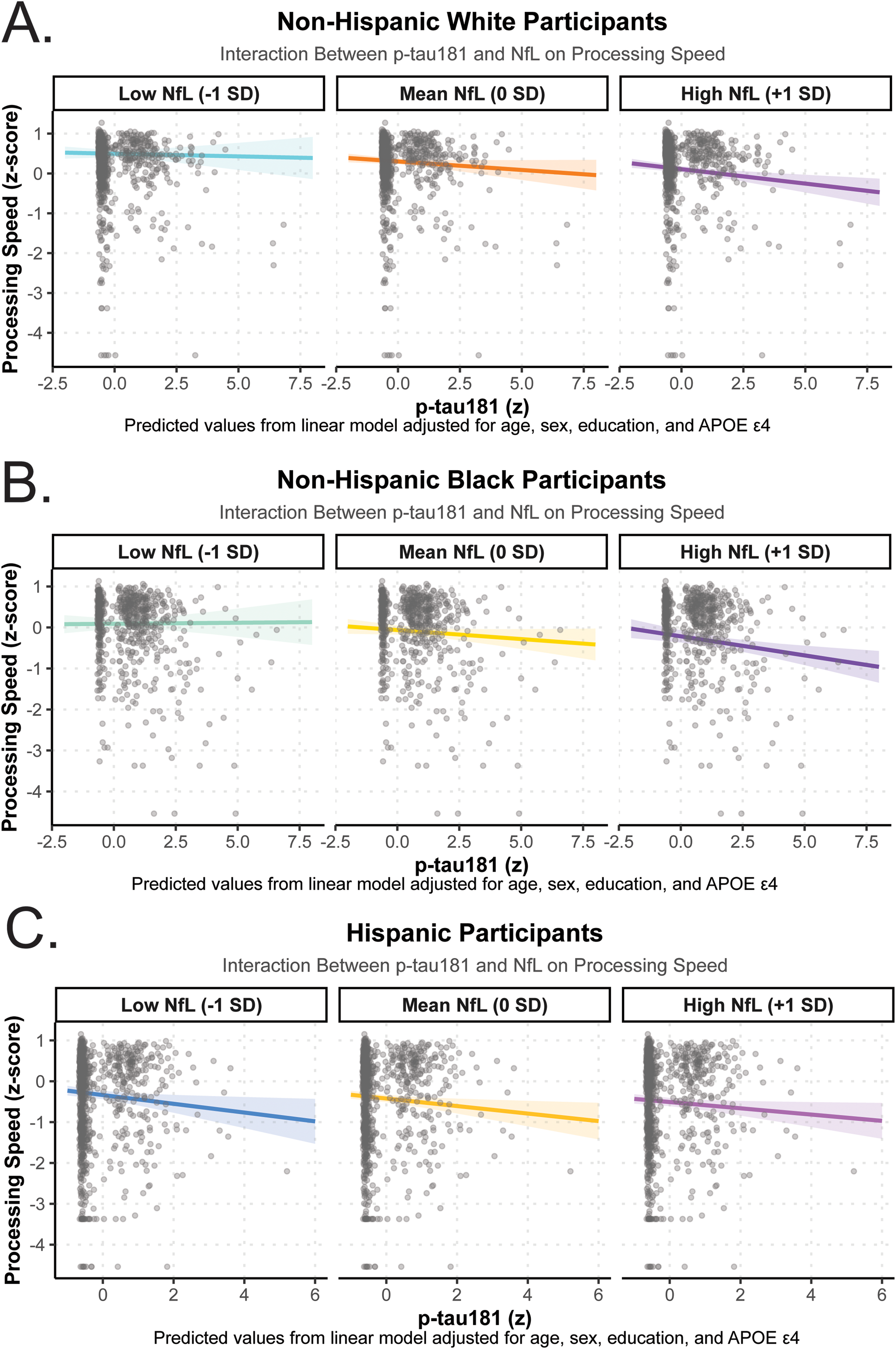
Race/ethnicity-stratified interactions between plasma p-tau181 and NfL on processing speed. Panels depict predicted processing speed z-scores by p-tau181 levels across low (−1 SD), mean (0 SD), and high (+1 SD) NfL concentrations among (A) Non-Hispanic White, (B) Non-Hispanic Black, and (C) Hispanic participants. Each line represents the estimated slope of p-tau181 at a given NfL level from linear regression models adjusted for age, sex, education, and APOE ε4 carrier status. Shaded regions indicate 95% confidence intervals. Negative slopes indicate poorer processing speed performance with higher p-tau181 concentrations, and steeper declines at higher NfL levels suggest a synergistic effect of axonal and tau-related neurodegeneration on cognitive slowing.
Discussion
In this large, racially and ethnically diverse cohort of older adults, plasma biomarkers reflecting amyloid/tau pathology and neuroaxonal injury demonstrated strong, domain-general associations with cognitive performance. Both p-tau181 and NfL were robustly associated with poorer memory, executive function, language, and processing speed performance, supporting their sensitivity to global neurodegenerative processes. When modeled together, each biomarker retained significant and independent associations with cognition, indicating that p-tau181 and NfL capture overlapping yet biologically distinct aspects of disease burden—amyloid and tau-mediated neurofibrillary degeneration and axonal injury, respectively. These findings reinforce the clinical value of combining amyloid/tau- and NfL-based measures to capture complementary facets of neurodegeneration rather than relying on a single marker of pathology. Importantly, the magnitude and pattern of associations differed by racial ethnic backgrounds, with attenuated p-tau181 cognitive relationships observed among NHB participants, while NfL effects were broadly preserved across groups. Lastly, synergistic interactions between p-tau181 and NfL, most pronounced for processing speed, suggest that concurrent amyloid/tau accumulation and axonal injury may exacerbate cognitive decline through convergent neurodegenerative pathways. Collectively, these findings highlight the interdependence of amyloid/tau and neuroaxonal injury in shaping cognitive outcomes and underscore the importance of examining biomarker cognitive relationships within racially and ethnically diverse populations.
In our analytical sample, NHB and Hispanic participants exhibited greater cognitive impairment, as reflected by higher prevalences of MCI and dementia compared with NHW participants. Cognitive domain specific scores followed a similar distribution, with NHB and Hispanic participants demonstrating lower performance across all domains relative to the NHW participants. Importantly, plasma AT(N) biomarkers were significantly associated with cognitive status, particularly p-tau181. Consistent with prior studies, elevated p-tau181 levels were observed among individuals with MCI and dementia, aligning with its well-established association with tau pathology and AD pathology progression.48–51 NfL concentrations showed a comparable pattern of increase across cognitive strata as previous studies52,53; however, in our cohort, NfL levels were similar between cognitively normal and MCI participants, rising notably only among those with dementia. Overall, these findings largely corroborate prior evidence linking plasma biomarkers with cognitive status, while also highlighting racial and ethnic disparities in both biomarker profiles and cognitive performance.
When examined individually, all plasma biomarkers were significantly associated with poorer cognitive performance across all domains. The effect sizes for NfL and total tau were comparable across domains, indicating similar magnitudes of association with global cognitive decline. In contrast, both p-tau181 and the Aβ42/40 ratio exhibited the strongest associations with memory, while their effects on executive function, processing speed, and language were of similar magnitude. These findings are particularly important as they reinforce that AD-specific pathology, as indexed by plasma p-tau181 and Aβ42/40, is most strongly linked to episodic memory impairment, supporting their discriminative ability to map molecular pathology onto cognitive phenotypes.54,55 The consistent effects of NfL across all domains underscore its role as a nonspecific marker of neurodegeneration rather than a domain-specific biomarker correlate.19,56 This pattern aligns with prior studies demonstrating that plasma NfL reflects widespread axonal injury and is associated with multi-domain cognitive impairment and global impairment.19,56–59
Consistent with our study objective, p-tau181 and NfL retained independent associations with cognition when included simultaneously in the statistical model. Notably, p-tau181 demonstrated a stronger association with memory than NfL. In contrast, NfL showed more robust associations with language and processing speed, domains often linked to diffuse white matter damage and global neurodegenerative processes.58–61 Although plasma p-tau181 is often interpreted as a marker of downstream tau pathology, its elevation in cognitively unimpaired individuals likely reflects early amyloid-driven phosphorylation events rather than extensive neurofibrillary tangle formation.62,63 This aligns with emerging evidence that plasma p-tau181 elevations precede overt tau deposition and may represent an early stage of AD-related pathophysiology.26,50 These findings reinforce the clinical value of combining tau- and NfL-based measures to capture complementary facets of neurodegeneration rather than relying on a single marker of pathology. 64
The independent associations of plasma p-tau181 and NfL with cognitive domain performance across racial and ethnic groups highlight both shared and distinct biological pathways linking neurodegeneration to cognitive aging. Although our study was not designed to elucidate underlying pathological mechanisms, heterogeneity of associations is consistent with reports that biomarker cognition relationships vary by disease clinical stage,65,66 sex, 67 pathology accumulation stages such as amyloid burden, 68 but there have been less investigations across racial ethnic groups.69,70 Among NHW participants, both p-tau181 and NfL exhibited robust and uniform associations across all cognitive domains, suggesting that AD-specific and nonspecific neurodegenerative processes act in parallel to influence global cognition. NHW participants were predominantly cognitively normal which emphasizes that these biomarkers are associated with cognition in the preclinical and prodromal phases of AD. 71 In contrast, among NHB and Hispanic participants, p-tau181 demonstrated more domain-specific associations, most notably with memory and processing speed while NfL remained consistently associated with multiple domains, particularly processing speed.
These racial-ethnic-stratified differences underscore the complexity of interpreting plasma biomarkers across diverse populations. The stronger and more diffuse associations observed among NHW participants may reflect a closer coupling between molecular pathology and clinical expression of cognitive impairment, whereas attenuated or domain-restricted effects in NHB and Hispanic groups could arise from multiple sources. Biological factors such as differential comorbid vascular or inflammatory burden, 72 variation in APOE ε4 frequency, 73 or distinct resilience mechanisms may modulate the clinical translation of biomarker burden. 74 Sociocultural contributors including educational quality, lifelong cognitive engagement, and social determinants of health also influence the cognitive expression of neuropathology, potentially decoupling biomarker load from observed performance in historically underrepresented groups. 75
The sensitivity analyses adjusting for clinical cognitive status further clarified these relationships. For NHW participants, the attenuation of p-tau181 associations outside the memory domain after adjustment suggests that tau-related pathology exerts its primary cognitive effects through memory impairment, consistent with prior imaging and autopsy studies linking medial temporal tau accumulation to episodic memory decline. 76 In contrast, NfL retained significant associations across executive, processing speed, and language domains, reinforcing its role as a global marker of neuroaxonal injury that extends beyond AD-specific processes.19,56–59 Among NHB and Hispanic participants, the persistence of memory- and processing speed- related associations with p-tau181 and NfL after adjustment indicates that these biomarkers capture complementary yet distinct aspects of cognitive vulnerability within each population.
The observed p-tau181 × NfL interactions for processing speed in NHW and NHB participants suggest a potential synergistic effect of concurrent tau pathology and axonal degeneration on subcortical-cortical network efficiency.77,78 Processing speed is often sensitive to diffuse white matter and network-level dysfunction, and its impairment in the presence of elevated levels of both biomarkers may signal accelerated disease progression or overlapping pathogenic cascades.79,80 Worse cognition when p-tau181 and NfL are simultaneously elevated suggests converging mechanisms of neuronal injury and tau phosphorylation that jointly accelerate neurocognitive impairment. 54 Among NHW participants, the p-tau181 × NfL interaction was significantly associated with slower processing speed, whereas in NHB participants, similar patterns emerged with slightly larger effect estimates, despite overall attenuation after adjustment for clinical cognitive status. These patterns may reflect differential vulnerability to combined tau- and axonal-related pathology across groups, potentially influenced by vascular comorbidities or systemic inflammation—pathways that warrant further investigation in diverse aging populations. 25 No significant interactions were observed among Hispanic participants, which may reflect demographic or health-related heterogeneity and differential distribution of underlying neuropathology. Taken together, these results underscore the importance of examining biomarker interactions, not solely their independent associations, when characterizing neurodegenerative risk. While p-tau181 and NfL each represent distinct biological pathways, their interplay may provide a more comprehensive measure of cumulative neuronal injury burden. The attenuation of interaction effects after adjusting for cognitive diagnosis further suggests that these biomarker relationships are tied to cognitive stage status. Future longitudinal work should examine whether combined elevations of p-tau181 and NfL predict accelerated decline or progression to dementia across racial and ethnic groups, and whether these relationships are modified by vascular, metabolic, or social determinants of health.
This study has several notable strengths. First, it leverages a large, racially and ethnically diverse cohort of older adults, allowing for robust examination of plasma biomarker–cognition relationships across groups that have been historically underrepresented in AD research. This study extended beyond global cognition by examining domain-specific performance, allowing clearer differentiation between AD–specific pathology (p-tau181, Aβ42/40) and nonspecific neurodegenerative processes (NfL, total tau). We also simultaneous modeled p-tau181 and NfL to evaluate their independent and joint contributions to cognitive performance offering insights into overlapping yet distinct biological pathways underlying cognitive decline. Lastly, the integration of racial ethnic-stratified analyses provides critical insights into how biomarker–cognition associations may differ across populations, supporting the broader goal of advancing biomarker equity and generalizability in precision dementia research.
Several limitations should also be noted. First, the analyses were cross-sectional, which precludes causal inference about temporal relationships between changes in biomarkers and subsequent cognitive decline. Second, there was a limited sample size for GFAP; therefore, we could not investigate this biomarker. Similarly, measures of brain pathology such as white matter hyperintensities were not incorporated, limiting our ability to evaluate potential mediating pathways between vascular injury, plasma biomarkers, and cognition. Lastly, we did not investigate p-tau217 and the p-tau217/Aβ42 ratio, preventing us from comparing the performance of these next-generation tau biomarkers relative to p-tau181.
To our knowledge, this represents one of the largest investigations of plasma biomarkers and cognition conducted to date, and the first of this scale to evaluate multiple cognitive domains within a racially and ethnically diverse cohort including NHW, NHB, and Hispanic participants. Future work should build on these findings by incorporating longitudinal follow-up to determine whether p-tau181, in combination with p-tau217 and other emerging tau markers, predicts differential trajectories of decline across specific domains, clarifying whether tau biomarkers serve as both diagnostic indicators and prognostic tools for monitoring domain-specific outcomes over time. The inclusion of blood-based markers of neuroinflammation such as GFAP, which has been linked to executive and processing speed deficits, will refine mechanistic interpretation, while the integration of structural imaging markers such as white matter hyperintensities may elucidate vascular contributions and potential mediating pathways between plasma biomarkers and cognition. It will also be essential to examine how social and structural determinants of health modify biomarker–cognition relationships across underrepresented populations, advancing equity in AD research. In parallel, analyses leveraging the available amyloid PET data within HABS-HD will be critical for evaluating concordance between plasma- and imaging-based amyloid measures and determining whether p-tau181–cognition associations differ by amyloid PET positivity. Such work could clarify whether plasma tau reflects amyloid-dependent or independent neurodegenerative processes and how p-tau181 and NfL jointly capture the interplay between tau- and neurodegeneration-related pathways. These findings underscore the complementary mechanistic and prognostic value of plasma p-tau181 and NfL, supporting their combined use to enhance early-stage detection and risk stratification of AD across diverse populations.
Supplemental Material
sj-docx-1-alz-10.1177_13872877251415367 - Supplemental material for Independent associations of phosphorylated tau181 and neurofilament light with cognitive outcomes in the Health and Aging Brain Study–Health Disparities (HABS-HD)
Supplemental material, sj-docx-1-alz-10.1177_13872877251415367 for Independent associations of phosphorylated tau181 and neurofilament light with cognitive outcomes in the Health and Aging Brain Study–Health Disparities (HABS-HD) by Mohammad Housini, Joey Annette Contreras, Cellas A. Hayes and in Journal of Alzheimer's Disease
Footnotes
Acknowledgements
The authors thank HABS-HD study participants for commitment to contribute to the ongoing research study.
Ethical considerations
All HABS-HD protocols were approved by the University of North Texas Health Science Center Institutional Review Board in accordance with the Declaration of Helsinki.
Consent to participate
All HABS-HD participants provided informed consent to participate in the study.
Consent for publication
Not applicable
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The HABS-HD Study is funded by the National Institute on Aging of the National Institutes of Health under Award Numbers R01AG054073 and R01AG058533, P41EB015922 and U19AG078109. MH is funded by the Health Enhancement Scientific Program (HESP) under U19AG024904. CAH is funded by Burroughs Wellcome Fund Postdoctoral Enrichment Program (PDEP) 1267001, and HESP U19AG078109.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.