
Abstract
Background
The retrosplenial cortex (RSC) is a cortical area that functions as a key component of the core network of brain regions involved in cognitive functions such as episodic memory, navigation, and planning. The RSC is capable of theta rhythm generation and, like the hippocampus, could be compromised in neurological diseases such as Alzheimer's disease (AD). Importantly, detecting early changes in RSC of transgenic animals could be translated into non-invasive biomarkers that can detect preclinical stages of AD and related dementias.
Objective
Our aim in the present study was to evaluate molecular and functional alterations in the RSC of very young 3xTg-AD mice (1-month-old).
Methods
Immunohistochemistry, anterograde viral tracer using an adeno-associated virus, western blotting, and electrophysiology were all carried out.
Results
Our results show significant accumulation of intracellular amyloid-β (Aβ) and hyperphosphorylated tau (pTau) in principal neurons from 1-month-old 3xTg-AD mice, which correlates with GSK3β activation and tau phosphorylation at serine 396. Coincidentally, oscillatory activity from the RSC is altered in the young 3xTg-AD mice. Specifically, we found that theta frequency is significantly higher in the transgenic animals.
Conclusions
In summary, our results suggest that the early accumulation of intracellular Aβ may affect the excitability of the RSC network, possibly due to changes in pTau resulting from GSK3β activation.
Introduction
Robust oscillations are detected at the theta frequency band (2–12 Hz) in the hippocampus.1–4 It is widely accepted that the synchronized firing of hippocampal pyramidal neurons contributes to endogenous theta rhythm generation.4–6 Similarly, in the retrosplenial cortex (RSC), the high correlation between neocortical neurons’ firing and theta power activity suggests that theta rhythm is generated locally, and the synchronized firing of neocortical pyramidal neurons is meant to be responsible for cortical theta activity generation and maintenance. 7 Both the hippocampus and the RSC are considered key nodes of a brain network that underpins cognitive functions such as navigation and memory.8–10 Indeed, it is known that theta rhythm is important for these brain functions.11,12 Considering that alterations in the generation of brain oscillation are involved in the pathophysiology of cognitive problems observed in patients with a variety of brain diseases,13,14 the cognitive deficits in Alzheimer's disease (AD) patients have been correlated with alterations in RSC activity and hippocampal theta rhythms.14,15 Similarly, altered hippocampal theta activity has been detected in AD transgenic mouse models.16–19 We recently reported that hippocampal theta activity is altered in triple transgenic AD mice (3xTg-AD) at 30 days of age. 19 Specifically, we found, for the first time, that hippocampal theta frequency increases in the 30-day-old 3xTg-AD mice. 19 In addition, we showed that hyperphosphorylated tau (pTau) at sites Ser396 and Thr231(major AD histopathological markers), which are located close to the microtubule domain region (MDr), coexists within hippocampal pyramidal neurons. 19 In J20-APP mice, we also reported altered theta activity that coincides with pyramidal cells exhibiting accumulation of β-CTF, a precursor of Aβ. 18 Altogether, our findings suggest that very early accumulation of intracellular Aβ and pTau could impact the excitability of the hippocampal circuit. Considering that Aβ and pTau pathology is propagated to interconnected circuits, and that there is a high hippocampal-RSC structural and functional connectivity,20,21 it is likely that the RSC network could be affected during early AD pathology. Surprisingly, the molecular and functional state of RSC pyramidal cells has never been evaluated in very young AD transgenic mice and compared to those in the hippocampus. To address this issue, we characterized the molecular and functional changes in RSC pyramidal cells of 30-day-old 3xTg-AD mice.
Our results revealed that Aβ and pTau, two major AD histopathological markers, coexist within pyramidal neurons from the hippocampus and RSC, which correlated with the activation of GSK3β and alteration in RSC network activity.
Methods
Animals
30-day-old male 3xTg-AD mice and age-matched non-transgenic (Non-Tg) mice (strain B6129SF2/J; Jackson Labs.) were housed in groups of 2–4 animals and maintained under a 12:12 light cycle with water and food available ad libitum. In 3xTg-AD mice, amyloid-beta precursor protein (APPSwe) and microtubule-associated protein tau (tauP30IL) transgenes are integrated at the same locus and are under the control of the mouse Thy1.2 promoter. Experimental procedures were approved by the Bioethics Committee of the Institute of Neurobiology, UNAM. The genotype used by each animal was confirmed by PCR (Jackson Labs. Protocol Standard PCR, at the Proteogenomic Unit of INB).
Electrophysiology
Animals were anesthetized with urethane (1.5 g / kg, i.p) and subsequently mounted on a stereotaxic apparatus for the placement of the recording electrode. The tip of the electrode reached the following coordinates (with respect to Bregma and dura): AP = -0.2 mm, ML = 0.4 mm, and DV = 0.1 mm. Local field potentials were recorded using a borosilicate electrode filled with aCSF (2–6 MΩ). Signals were recorded with a differential AC amplifier (A-M Systems), filtered (high pass 1.5 Hz, low pass 10 KHz). Using custom MATLAB software, field traces were further filtered and analyzed using the Fourier transform. Power spectra were calculated using the multitaper method (Chronux toolbox, i.e., using seven tapers). 22 Integrated theta-band power was calculated in 5-s bins over a period of 2–10 min, and the mean spectrum was taken as the grand mean of each animal. Theta power was measured in µV2/Hz.
Immunohistochemistry
Additional male mice were anesthetized with pentobarbital and transcardially perfused (PBS followed by 4% PFA). Brains were stored in fixative for 24 h at 4°C, then in a sucrose solution (30% in PBS) for three days at 4°C; next, they were frozen using dimethylbutane and stored at −80°C. Brains were sliced coronally at 40 μm using a freezing microtome for light microscopy, or a vibrating microtome at 50 μm for fluorescence microscopy, and free-floating sections were stored in a preservative solution (3:3:4 glycerol: ethylene glycol: PBS) at −20°C, in preparation for immunohistochemistry (IHC). Washes in PBS preceded all steps except primary antibody addition. All steps were performed at room temperature unless otherwise specified. For immunofluorescence, sections were incubated for 2 h with PBS containing 1% normal goat serum, 0.25% Triton X-100, and 0.45% gelatin. Sections were incubated overnight at 4°C with one of the following antibodies: pS396 (1:250 dilution) or BAM10 (1:500 dilution) (Thermo Fisher and Sigma-Aldrich, respectively). Sections were incubated for 2 h with secondary antibody (Alexa 488-596 conjugated goat anti-rabbit/mouse at; 1:1000 dilution and HRP goat anti-rabbit/mouse 1:1000). Sections were then mounted on glass slides with Vectashield Mounting Medium, Vector Labs. Fluorescence was visualized using an epifluorescence microscope (Axioplan2, Zeiss). The RSC areas were visualized with a 20× objective. A high magnification photomicrograph was obtained with a 40× objective.
Western blotting
For western blot analysis, total protein from brain (hippocampus) was obtained by lysis buffer, containing: 50 mM Trizma base, 150 mM NaCl, 1% Triton, 0.5% SDS, 1 mM NaP2O7 and protease inhibitors (Complete Protease Inhibitor Cocktail Tablets; Roche; Mannheim, Germany). A total of 15–20 µg of total protein was loaded onto a 10% SDS-PAGE, separated, and transferred onto Immobilon-P transfer membranes (Millipore). The following antibodies and dilutions were used: Tau5 (monoclonal antibody (mAb) against total Tau, Abcam, 1:2000); pS396 (mAb against phosphorylated Tau, ThermoFisher, 1:1000) and anti-pGSK3β (phospho S9, Abcam, 1:2000). Blots were developed using ECL-Plus (GE Healthcare) or SuperSignal West Femto (Pierce). Results were quantified with ImageJ software and normalized to the total tau protein signal for pS396 and for GSK3β (S9).
Data analysis
All data were compared statistically using GraphPad Prism 5 Windows, version 5.01 software. In all tests, values of p < 0.05 were considered significant unless otherwise specified. Bar graphs show experimental mean, with dispersion bars indicating standard error of the mean unless otherwise specified.
Results
Hippocampal pyramidal cells from young 3xTg-AD mice exhibit intracellular Aβ and pTau accumulation
Immunohistochemical analysis showed the presence of intracellular Aβ accumulation, as revealed by the presence of BAM10 immunostaining, in subicular-pyramidal neurons from p30 3xTg-AD samples (Figure 1A). The observed of Aβ in pyramidal cells was also present in the hippocampal CA1 area of 3xTg-AD samples (Figure 1B). Within those two areas, well-defined pyramidal cells were detected by Aβ labeling (Figure 1A, B, arrowheads). Non-detected levels of Aβ were observed in the family-related Non-Tg group (Figure 1C). Aiming to explore if the Aβ accumulation in the hippocampus of 30-day-old 3xTg-AD mice was related to an increase in pTau at the repeat-microtubule domain sites (MDr), we evaluated the presence of tau phospho-serine 396 labeling. As previously shown, 19 we found an increase of phosphorylated tau protein at serine 396 in the pyramidal cells from the CA1 and subiculum area of the young 3xTg-AD group (Figure 1D). When we compared the total amount of labeled neurons per mm2, the number of principal cells comprising Aβ (mean = 67.22 cells/mm2) was statistically higher than the number of neurons comprising Ser396 (mean = 42.55 cells/mm2, BAM10 versus pSer396, p = 0.027, Wilcoxon test, Figure 1E). Additionally, we identified a direct connection between hippocampal CA1-Sub and RSC in mice that was made using an anterograde viral tracer using an adeno-associated virus to induce GFP expression (AAV-GFP). 9 The anatomical location of the injection (CA1-Sub) and projection (RSC) sites were identified and registered [Allen Mouse Brain Connectivity Atlas (http://connectivity.brain-map.org/projection)]. Representative images of the strong GFP expression at the CA1-Sub injection site are shown in coronal sections (Figure 1F, G). As expected, incoming GFP-labeled projections into the ipsilateral RSC are clearly visible (Figure 1F, G).
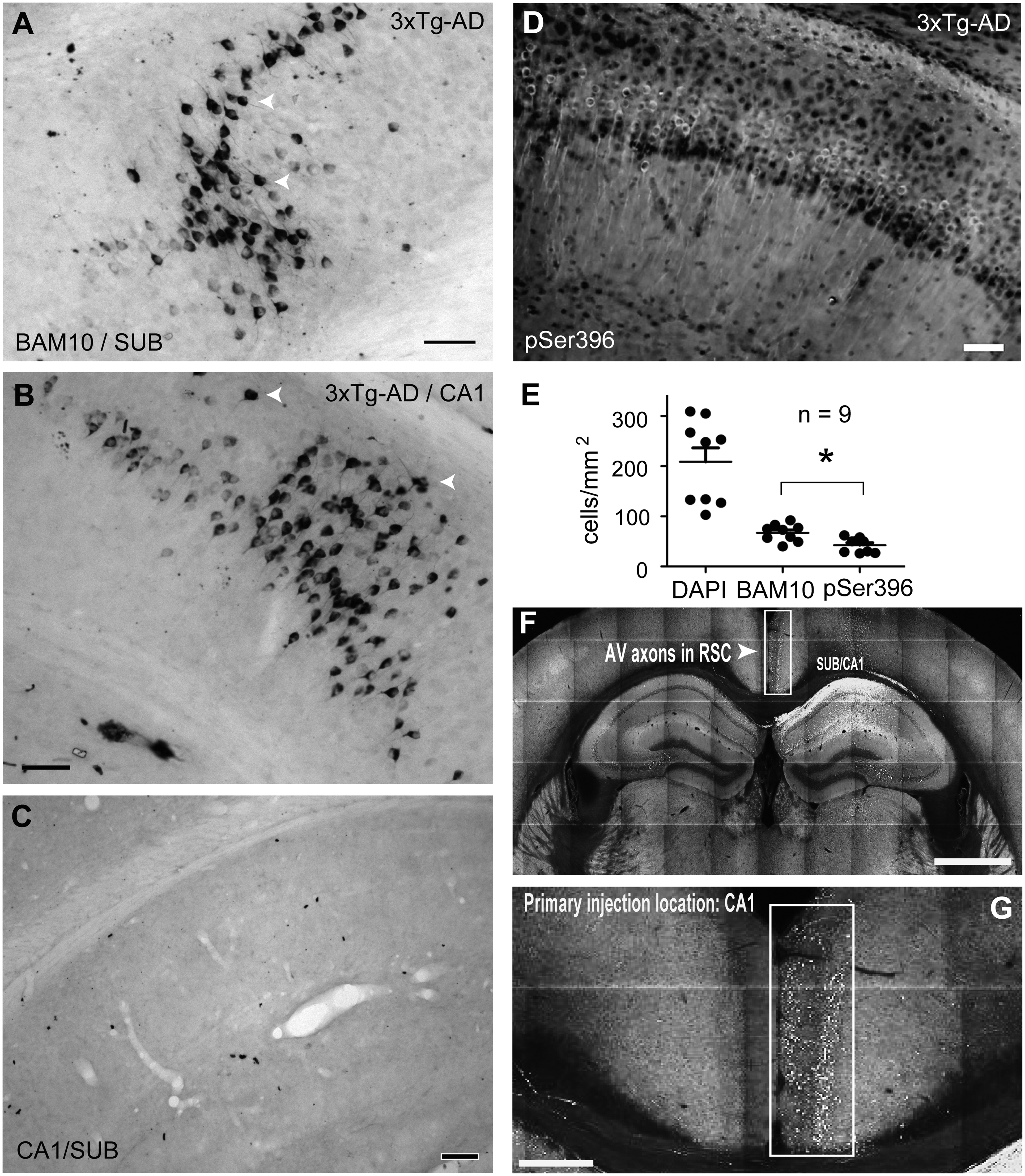
Taken together, the immunohistochemical studies revealed that CA1-Sub hippocampal principal cells from young 3xTg-AD mice, which establish long-range connections with the RSC, are characterized by Aβ accumulation and abnormal tau phosphorylation.
RSC cells from young 3xTg-AD mice exhibit intracellular Aβ accumulation
To explore the possibility that RSC from very young 3xTg-AD mice reflects similar molecular alterations to the ones observed in the hippocampus, we evaluated the presence of Aβ in pyramidal cells from RSC (Figure 2A). As expected, the RSC from Non-Tg samples showed low-detectable traces of extracellular and intracellular Aβ accumulation (Figure 2B). In contrast, immunohistochemical detection with BAM10 revealed the existence of intracellular Aβ primarily at layer V pyramidal neurons in the 30-day-old 3xTg-AD mice (Figure 2C, see magnification in 2C’). The presence of somatic Aβ outside layer V was barely detected in the 3xTg-AD samples (Figure 2D). Again, the presence of Aβ in pyramidal cells continued along layer V of 3xTg-AD independent samples (Figure 2D, E, see magnification and white arrows in 2D’ and 2E’). Fewer cells from layer VI comprising Aβ were detected (Figure 2E). When we compared the relative Aβ fluorescence, 3xTg-AD group exhibited a significantly higher fluorescence than the Non-Tg group (p = 0.0002, Mann-Whitney U test, n = 4, Figure 2F). Summarizing, the RSC from very young 3xTg-AD mice showed intracellular Aβ accumulation.
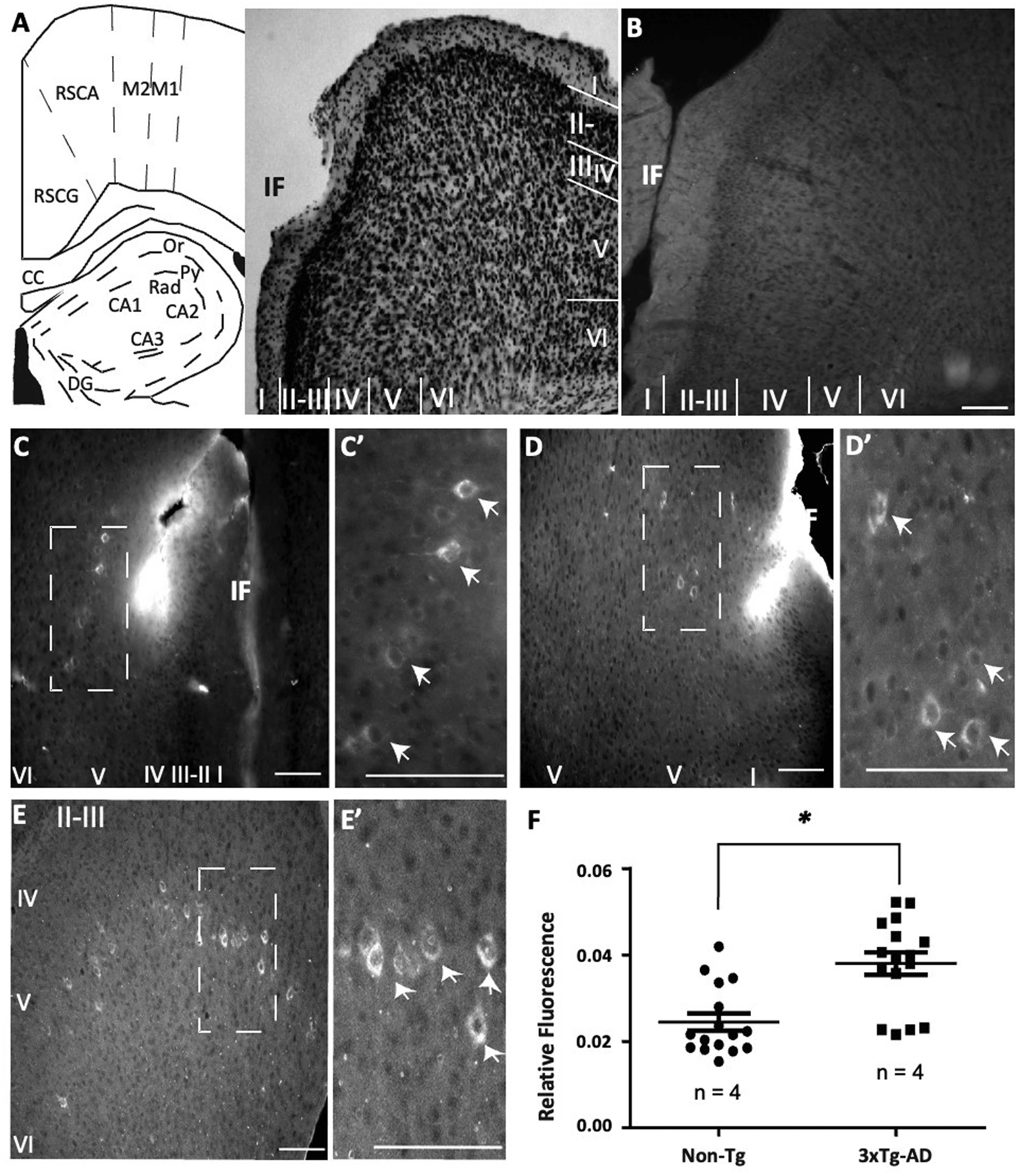
RSC cells from young 3xTg-AD mice exhibit intracellular pTau accumulation
Further pursuing the molecular characterization of RSC cells from very young 3xTg-AD mice (Figure 3A), we evaluated the potential changes in pTau levels. In accordance with our previous findings in the hippocampus, RSC from Non-Tg samples showed low-detectable traces of pTau (Figure 3B). In contrast, the pyramidal cells from 3xTg-AD RSC-layer V exhibited increased levels of phosphorylated tau at MDr site S396 (Figure 3C). Cells from all the layers of the RSC (I to VI) were characterized by the presence of pTau (Figure C, see magnification in C’). Increases in pTau were not only detected in somata but also in all the dendritic compartments of RSC cells (Figure 3Dto E’). When we quantified the relative fluorescence of pTau, the 3xTg-AD group showed a significant increase when directly compared to the Non-Tg group (p = 0.0001, Mann-Whitney U test, n = 4, Figure 3F).
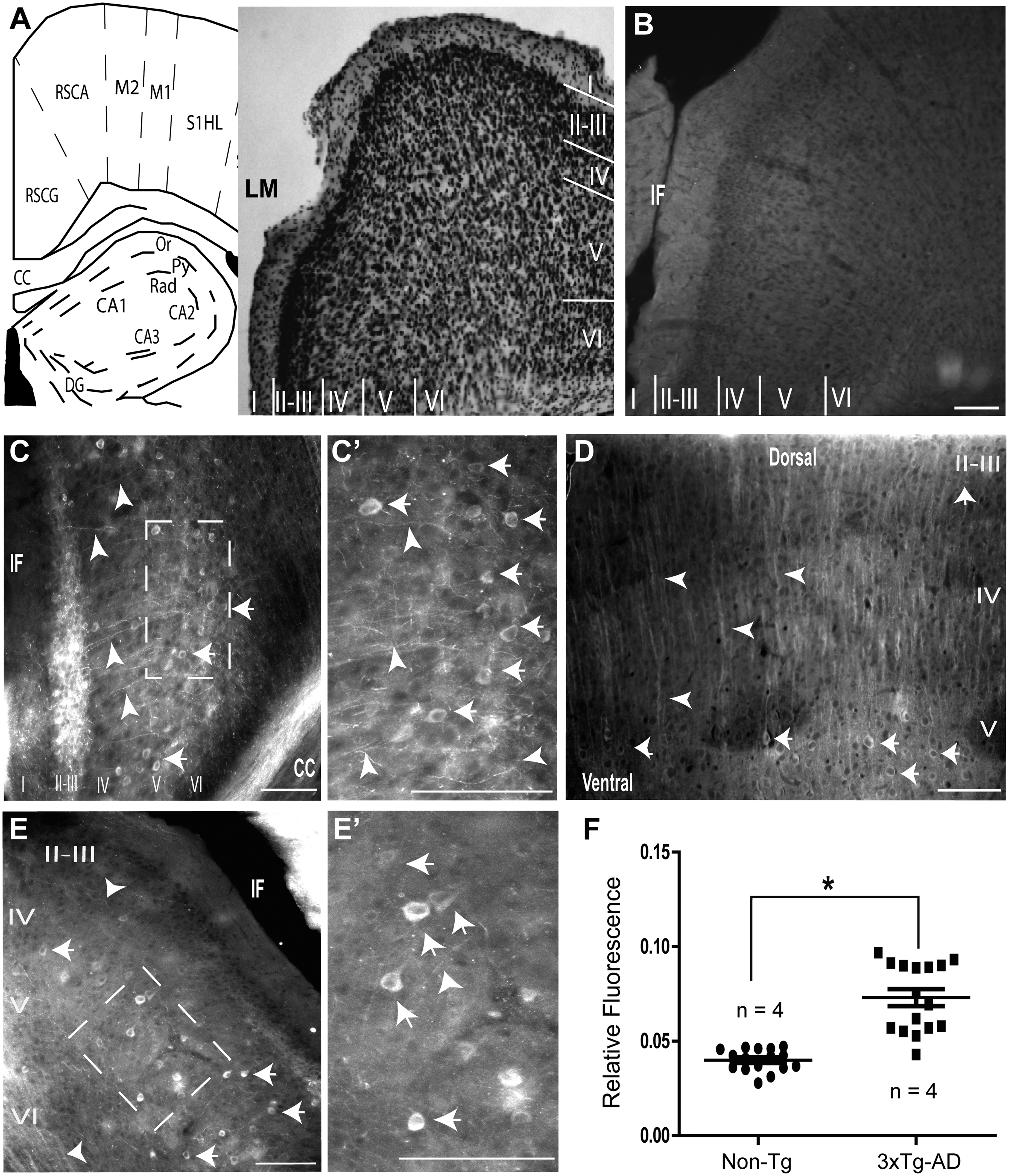
In sum, like hippocampal principal cells from young 3xTg-AD mice, the principal cells from RSC, as well as other cortical cells, were characterized by an increase in pTau.
Network activity at theta band frequency is altered in RSC of young 3xTg-AD mice
So far, we have shown that RSC exhibits similar neuropathological changes to those of the hippocampus in 3xTg-AD mice. That is, Aβ and pTau coexist within the RSC pyramidal neurons in the young 3xTgAD mice. Altogether, our findings suggested that, as occurs in the hippocampus, 19 very early accumulation of intracellular proteins could impact the excitability of the RSC network. To address this question, we studied the intrinsic spontaneously generated theta oscillations in the RSC of the 30-day-old 3xTg-AD mouse model. In line with previous reports, 18 population activity from Non-Tg animals showed rhythmic theta activity in the RSC (Figure 4A). Filtered traces at theta band frequency (2–12 Hz) further confirmed the presence of rhythmic RSC network activity (Figure 4B). In contrast, RSC network activity recorded from the young 3xTg-AD mice displayed altered rhythmic activity (Figure 4A, B). The spectrograms revealed a stable power theta frequency in the Non-Tg group (4.54 ± 0.48 Hz, Figure 4C, D); this frequency increased in the young 3xTg-AD mice (6.24 ± 0.44 Hz; Mann-Whitney test, p = 0.04, Figure 4C, D). Importantly, no significant differences were observed in the relative theta power between Non-Tg and 3xTg-AD groups (Non-Tg: 0.278 × 10−7 ± 0.05 µV2 and 3xTg-AD: 0.251 × 10−7 ± 0.09 µV2, Figure 4E).
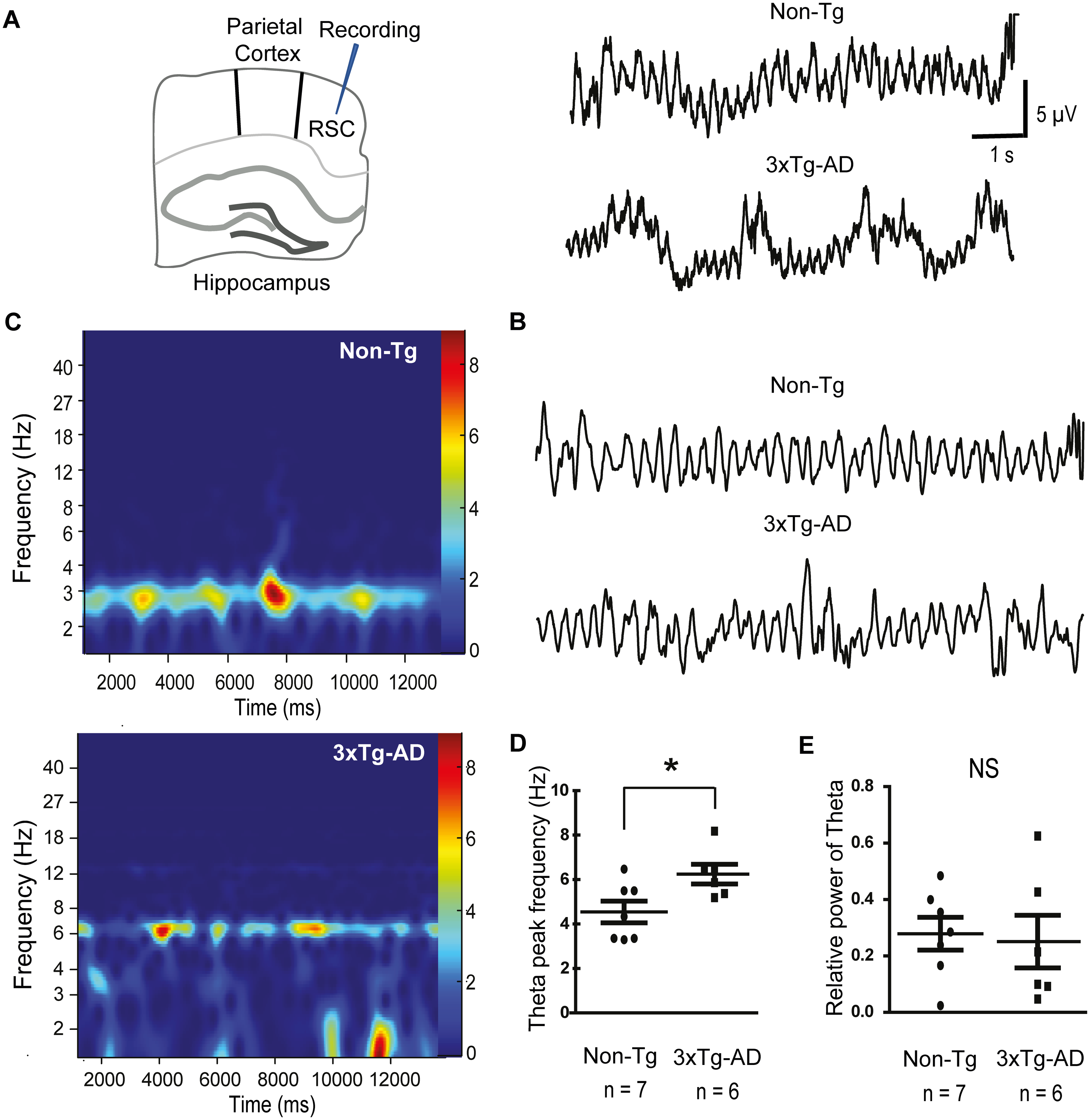
Alterations in GSK3β contribute to tau hyperphosphorylation
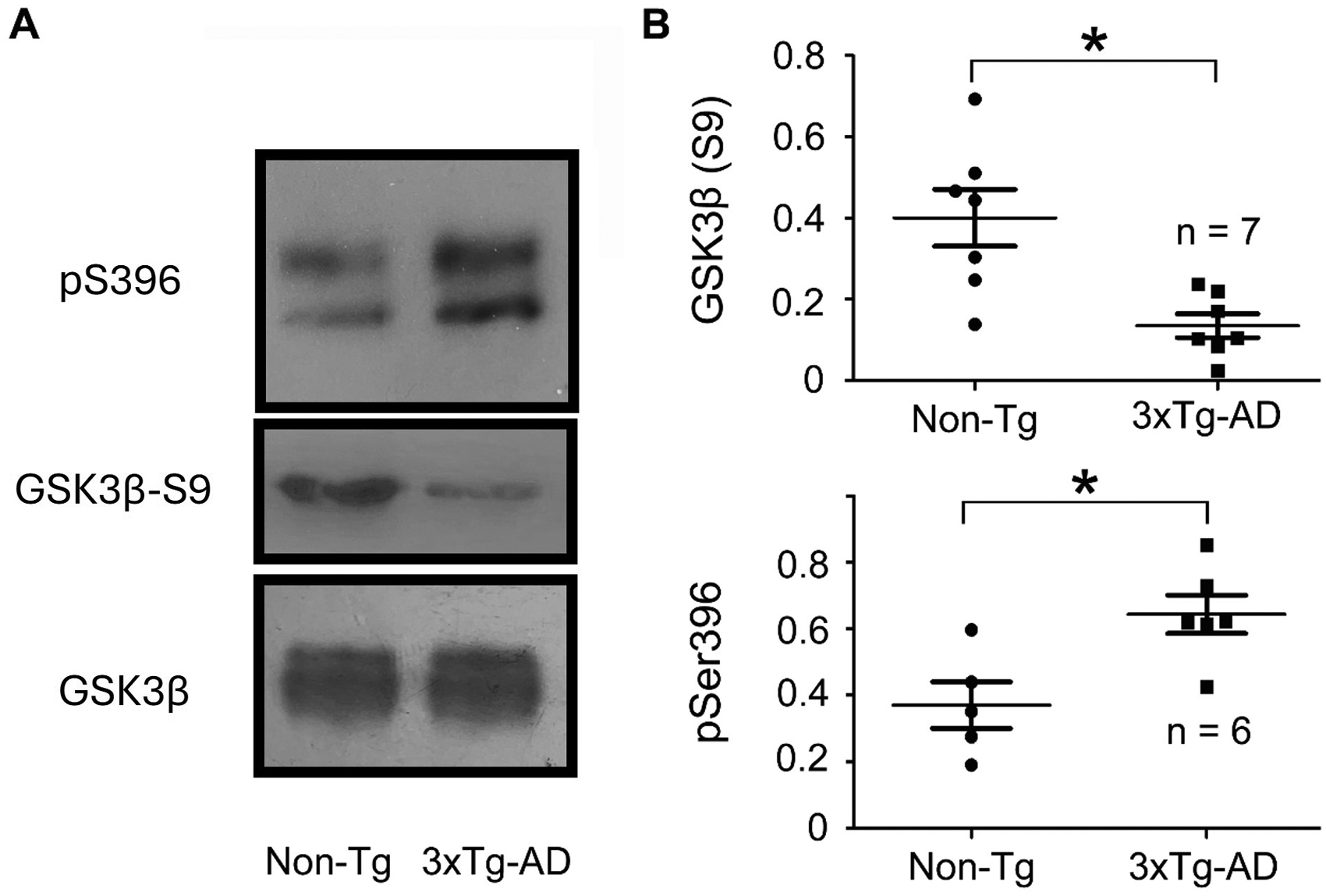
Considering that GSK3β plays a major role in Aβ-induced tau hyperphosphorylation. 23 Here, we evaluated the levels of GSK3β phosphorylation at Ser9 as a reporter of its activity.24,25 Western blot analysis of total protein revealed that GSK3β phosphorylation is altered in p30 3xTg-AD animals (Figure 5A). We found reduced phosphorylation of GSK3β at Ser9 in p30 3xTg-AD animals (Figure 5A), with no detectable changes in the total amount of GSK3β protein (Figure 5A). Statistical analysis further confirmed our findings. The quantification of the phosphorylation levels of GSK3β at Ser9, measured as the ratio of phosphorylated GSK3β at Ser9 over total GSK3β, showed a significant reduction compared to the family related Non-Tg group (Non-Tg = 0.4003 and 3xTg-AD = 0.1343, α= 0.05, p = 0.004, df = 12, Mann-Whitney U test, Figure 5B). Likewise, the ratio of phosphorylated tau over total tau was significantly higher in the 3xTg-AD samples when directly compared to Non-Tg samples (>15%, α = 0.05, p = 0.01, df = 6, Mann-Whitney U test, Figure 5A, B).
Discussion
In the present study, we have assessed the biochemical and functional changes of pyramidal neurons in very young 3xTg-AD mice (P30). Despite the wide consensus that Aβ deposition in a variety of AD transgenic mice, including 3xTg-AD, is detected after several months of age,26,27 this study shows the first evidence of intracellular Aβ accumulation in pyramidal cells as early as 30 days of age (Figures 1 and 2), which does not seem to be related to any cognitive compromise. 28 This last observation is similar to the absence of cognitive impairment recently examined in p30 J20-APP transgenic mice. 29 Thus, this observation is in line with the clinical finding of cerebral Aβ aggregation two or three decades before the onset of detectable cognitive symptoms in AD. 30 Despite some controversy as to where Aβ accumulation arises in the human brain, 31 there is evidence that deposition begins in cortical regions and then in the hippocampus.26,32 In our study, Aβ accumulation seems to be stronger in the hippocampal pyramidal cells when compared to the neocortical pyramidal neurons from the RSC (Figures 1 and 2). This finding further supports the concept that hippocampal pyramidal cells are challenged since very early stages of AD development.
The Aβ cascade hypothesis postulates that pTau is a downstream target of Aβ.33,34 In this regard, we and others have reported that Aβ is responsible for increasing tau phosphorylation close to the MDr sites.34,35 In agreement with these reports, here we observed that pyramidal cells from young 3xTg-AD mice displayed both Aβ accumulation and increased pTau close to the MDr sites (Figure 1). We think that at the time point of our evaluation (P30), this pathological relationship is still developing, since we found a higher number of cells exhibiting Aβ than pTau accumulation (Figure 1), which is consistent with previous reports,34,36,37 indicating that Aβ and pTau are mechanistically and chronologically linked since the very early stages of the pathophysiological process.
Many reports have shown that synaptic dysfunction is an early event that precedes the presence of fibrillar lesions, Aβ plaques, and neurofibrillary tangles. 38 Here, we demonstrate that non-fibrillar lesions (intracellular Aβ and pTau) coexist with altered brain activity (Figure 4). Similarly, the occurrence of intraneuronal Aβ and pTau immunoreactivity in CA1 pyramidal neurons has been correlated with impairments in synaptic plasticity (LTP/ LTD),27,39 and recently, with altered theta activity. 19
Anatomical and molecular connections between the hippocampus and RSC
In line with recently published data, 10 results showed the existence of a long-range connection between the CA1-Sub and RSC (Figure 1). Another key connection involves reciprocal pathways between the RSC and the prefrontal cortex, providing an indirect route for hippocampal influence on the dorsolateral prefrontal cortex and vice versa, potentially modulating executive functions. Additionally, these circuits integrate visual and somatosensory feedback through reciprocal connections with the dorsal superior temporal sulcus and the anterior thalamic nuclei. 40 Due to this connection that carries hippocampal-related information, 8 it is being suggested that RSC is highly involved in hippocampus-dependent functions such as episodic memory, navigation, and future planning.8,21,41
Both hippocampal and RSC pyramidal neurons were characterized by intracellular accumulation of Aβ and pTau proteins (Figures 2 and 3). However, in contrast to the hippocampus, the RSC pyramidal neurons showed lower levels of Aβ when compared to pTau (Figures 2 and 3). These data suggest that increased pTau within the RSC can respond to increased Aβ in both the hippocampus and the RSC. Despite their origins, the presence of both AD-related proteins within the pyramidal cells of RSC suggests functional alterations in this network.
Considering that pyramidal cells are essential for theta activity,4,5,42 we found that this network activity in the RSC increases in frequency as early as thirty days of age in the 3xTg-AD mouse model (Figure 4). It is likely that the alterations in theta rhythm in the RSC can reflect alterations not only within this network but also in its direct functional connection with the hippocampus, since we have recently reported that the frequency of hippocampal theta is also significantly increased in very young 3xTg-AD mice.19,28
Accordingly, the 3xTgAD model at seven months of age exhibited increased power and frequency of cortical oscillatory activity. 43 In line with this, we recently published a meta-analysis examining molecular and electrophysiological changes in AD-transgenic mouse models across different ages. Our analysis revealed that Aβ pathology is associated with hyperexcitability in older animals, whereas pTau is linked to hypoexcitability. Notably, the Aβ-driven phenotype tends to dominate over the pTau phenotype as the disease progresses. 44
Although there is no consensus about the sequence of pathological changes occurring at the different hippocampal subfields during AD, human studies have revealed prominent atrophy of the subicular complex at early stages of AD, 45 which coincides with our data of early Aβ and pTau accumulation in the subiculum of 3xTg-AD mice, but also with a functionally-related projection area, namely the RSC. Altogether, these findings support a functional interplay between the RSC and hippocampus in AD pathology. Nevertheless, the relevance and dynamics of these connections in the human brain remain to be fully elucidated and warrant further investigation.
Involvement of GSK3β in the alterations of brain network activity
Included in the hypothesis of the Aβ cascade, the active form of GSK3β has been widely associated with the hyperphosphorylation of tau protein.23,46–48 Specifically, GSK3β has been associated with increases in tau phosphorylation close to the MDr sites such as Thr231, Ser235, Ser396, and Ser404.46,47 Further supporting this hypothesis, scientific documents have reported increased GSK3β expression in the brains of AD patients and several AD models. 49 GSK3β was one of the first identified substrates of the oncogenic kinase AKT. 50 Phosphorylation at Ser9 inhibits GSK3β activity via the formation of an autoinhibitory pseudosubstrate. 49 In the absence of phosphorylation at Ser9, GSK3β exists in an activated state, directly promoting tau phosphorylation and APP cleavage. 49 GSK3β phosphorylates tau sequentially at Ser404 and then Ser396. 51 Importantly, phosphorylated tau at Ser 404/396 has been associated with impaired LTD.35,52 In the 3xTgAD model, we previously identified an association between phosphorylated tau at Ser396/404 and disrupted hippocampal theta oscillations, accompanied by reduced hippocampal excitability. 19 Extending these findings, the present study reveals similar molecular and electrophysiological alterations in the RSC, a region with reciprocal connectivity to the hippocampus (Figure 4). Notably, pTau burden in the RSC exceeded Aβ burden, suggesting that increased pTau in the RSC may be driven by Aβ accumulation in both the hippocampus and RSC. Collectively, these data indicate that pTau likely impairs hippocampal synaptic function and network dynamics, with effects propagating to interconnected neocortical regions, ultimately contributing to behavioral deficits.34,35,53
Here, we showed that GSK3β could be responsible for increasing the levels of pTau at MDr sites in very young 3xTg-AD mice (Figure 5). Mechanistically, it was demonstrated that tau phosphorylation controls the interaction of tau with the postsynaptic PSD95-Fyn-NMDA receptor complex, leading to changes in synaptic plasticity.34,35 Therefore, an abnormal increase in GSK3β activity, leading to increased pTau, helps explain changes in brain network activity (Figure 5). Consistent with the above data, it was reported that Aβ alters hippocampal-entorhinal cortex population activity both in vivo and in vitro by activating the PI3 K/GSK3β pathway.24,54 Further supporting the pathological role of this kinase, it was reported that pharmacological and non-pharmacological inhibition of GSK3β prevented an Aβ-induced pathological effect in the population activity of the hippocampal-entorhinal cortex24,54 and the olfactory bulb. 25 Altogether, these molecular alterations likely underlie activity-dependent changes in synaptic strength that are essential for cognitive processing. 55 Importantly, our findings indicate that this pathological mechanism extends beyond the hippocampus to neocortical regions with strong functional connectivity, specifically the RSC.
In sum, our results show that at very early stages (P30), the CA1/Subicular/RSC circuit excitability from 3xTg-AD mice is already affected by the presence of Aβ protein in principal cells. It is likely that accumulation of Aβ results in pTau accumulation throughout GSK3β activation. It is also likely that Aβ and pTau could induce synergistic pathological processes, constituting important effectors of very early-onset familial AD.
Footnotes
Acknowledgements
We thank Jessica Gonzalez-Norris for proofreading. The authors acknowledge Sofía Díaz-Cintra and Azucena Aguilar-Vázquez for technical support and critical comments. We thank the following facilities from INB-UNAM: Proteogenomic Facility Unit, Behavioral Analysis Core Facility, and Vivarium Facility.
This manuscript is part of the requirements for obtaining a doctoral degree at the Posgrado en Ciencias Biológicas, UNAM, by Carlos A. Garcia-Carlos.
ORCID iDs
Ethical considerations
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the National Council of Humanities, Sciences, and Technologies CONAHCYT (grant numbers 269021 and 319863) and supported by the Dirección General de Asuntos del Personal Académico (DGAPA). Carlos Antonio García-Carlos was awarded by CONAHCYT (fellowship number 1082520), México. Dr Mondragón-Rodríguez was awarded a Cátedra position by CONAHCYT, México.
Declaration of conflicting interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Dr Perry is the Editor-in-Chief of JAD but was not involved in the peer-review process nor had access to any information regarding its peer-review. GP serves on Nervgen (Scientific Advisory Board) and Synaptogenix (Equity and Scientific Advisory Board).
The remaining authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.