
Abstract
Background
Habitual daytime napping is a significant aspect of many older adults’ sleep-wake cycle. Growing evidence links napping and cognition in the context of Alzheimer's disease (AD), yet little is known about how genetic risk influences this relationship.
Objective
This study investigates interactions between genetic risk for AD and napping on cognition in 1655 cognitively healthy middle-aged to older adults.
Methods
Cognition was assessed using a self-administered online battery and reduced to three variables: memory, visuospatial abilities and executive functions. Genetic risk was assessed with the presence of APOE ɛ4 allele and polygenic risk scores for AD (PRS; excluding APOE). ANCOVA assessed interactions.
Results
There was a significant interaction between APOE ɛ4 and napping duration on the memory component (F(2,1645) = 3.84, p = 0.022, ηp2 = 0.005). APOE ɛ4 carriers reporting long naps demonstrated better memory than non-carriers also reporting long naps. Among APOE ɛ4 carriers, those who napped for ≥ 1 h performed better than those reporting shorter naps. In a separate analysis, there was a significant interaction between PRS and napping (F(2653) = 3.44, p = 0.033, ηp2 = 0.010). Low PRS was related to better memory than high PRS among those who did not nap. Within the low PRS group, participants who did not nap outperformed those reporting short naps. Results remained significant after accounting for overnight sleep efficiency.
Conclusions
Findings suggest that the relationship between napping and memory may vary as a function of genetic risk for AD. Results could inform studies looking into personalized preventative treatment based on genetic profiles.
Introduction
Alzheimer's disease (AD) is the most common form of dementia, with increasing global incidence. 1 Unless preventive measures are developed, the prevalence is set to triple by 2050. 2 It can take between 20 and 30 years for AD to develop, with pathology beginning several years before any clinical symptoms. 3 Changes in cognitive function are a key feature, with memory problems typically being one of the early signs of cognitive decline. 4 In addition to age, which is the primary risk factor, genetic predisposition and lifestyle factors significantly contribute to the overall risk of developing late-onset AD.5,6 The present study examines these factors, focusing on napping, a common behavior among older adults, and investigates their interaction in influencing cognition.
The Apolipoprotein E (APOE) ε4 is the strongest known genetic risk factor for AD, with the global proportion of carriers who are not living with dementia estimated to be 23.9%. 7 The risk of developing AD increases in those with two ε4 alleles and the onset tends to be earlier. Specifically, homozygotes have 60% chance of developing AD by the age of 85. 8 The mechanisms underlying the relationship between APOE ε4 and AD are complex. ApoE has been shown to function in the clearance of inflammatory and pathogenic molecules, and APOE ε4 is associated with changes in pathological hallmarks in AD, including amyloid-β (Aβ), tau pathologies, neuroinflammation, synaptic dysfunction, and mitochondria/metabolism. 9 Differences in cognitive function between cognitively healthy APOE ɛ4 carriers and non-carriers have been proposed, with episodic memory and executive functions identified as specific domains where carriers tend to perform worse. 10 There is also emerging evidence suggesting an antagonistic pleiotropic effect of APOE ɛ4 on cognition, being beneficial early in life, but detrimental in older age. 11 AD polygenic risk scores (PRS), defined as an aggregation of multiple genetic risk variants to estimate a person's predisposition to the disease, 12 have also been used to study the relationship between genetic risk and cognition. In one study, both AD PRS, and APOE ɛ4 predicted the rate of cognitive change in cognitively healthy people. 13 In contrast, no significant associations were found between AD PRS and cognitive ability or age-related changes in another large study of cognitively healthy older adults. 14 Overall, inconsistencies in studies investigating genetic risk for AD have been suggested, highlighting the importance of considering variations in methodologies used to assess cognitive domains and differences in participants, including age range and where they sit in the progression to AD. 15
Amongst the various lifestyle factors, sleep patterns have been proposed to contribute to dementia risk. Sleep disturbances in older adults are associated with cognitive impairment and an increased risk of developing dementia.16–18 These disturbances are also closely related to genetically mediated biomarkers of AD, particularly Aβ. For instance, people who report short and long sleep duration, poorer sleep quality or longer sleep latency (time to fall asleep) are more likely to have higher levels of Aβ deposition in their brain.19–21 While findings on the role of genetics in the sleep-Aβ relationship are mixed, emerging evidence suggests that genetic predisposition can modulate the influence of non-genetic factors on dementia risk, for instance sleep. 22
Earlier research on sleep and cognition in older adults often excluded napping. However, the approach is shifting to encompass the full 24-h cycle to obtain a more comprehensive understanding of naturalistic sleep patterns. Daytime napping is a significant aspect of the older adult's sleep-wake cycle, given its higher prevalence in this population and its association with poorer nocturnal sleep, and altered circadian sleep-wake cycle.23–25 Napping is suggested to have a non-linear association with cognition. 26 Notably, short daytime naps have been associated with a lower risk of cognitive impairment and cognitive decline.27,28 In contrast, long and frequent daytime naps have been associated with poor cognitive performance and an increased risk of AD.29–31 Few studies have investigated daytime napping and genetic risk for AD specifically. Limited napping of up to 60 min has been suggested to have a protective effect against AD in APOE ɛ4 carriers, while longer naps increased the risk of AD. 32 A possible interaction of APOE genotype and napping frequency has also been suggested, with people at a higher genetic risk who occasionally nap showing a lower incidence of dementia. 33 To our knowledge, the relationship between daytime napping and AD PRS in cognitively healthy older adults has not yet been examined. However, higher AD PRS has been correlated with higher habitual daytime sleepiness in healthy young men. 34 This suggests a potential link between AD PRS and napping, given the strong association between daytime sleepiness and napping. 35
Despite significant recent advances in our understanding of AD and its pharmacological treatments, effective prevention remains paramount. This, combined with a rising popularity in genetic testing 36 highlights the need for a better understanding of modifiable factors that could play a key role in personalized interventions, particularly for at-risk groups such as APOE ɛ4 carriers. While existing literature suggests that napping may influence cognition, the extent to which genetic risk for AD influences this association remains unclear. Building on evidence that genetic risk may modify the effect of lifestyle factors on dementia, 22 investigating the interplay between napping and genetic risk for AD on cognitive abilities may provide valuable insights for the development of early lifestyle interventions.
The current study investigates the interactions of genetic risk for AD and daytime napping on the cognitive abilities of middle-aged to older adults. First, we examined whether genetic risk for AD (APOE ɛ4 and PRS) modified the relationship between napping and cognition. We then explored these results in the context of overnight sleep parameters.
Methods
Ethical approval was obtained from the Human Research Ethics committees of QIMR Berghofer (P2210).
Study sample
Data were sourced from the Prospective Imaging Study of Ageing: Genes, Brain and Behaviour (PISA). 37 PISA is a large cohort study of midlife and older Australians. The initial recruitment process involved leveraging extensive genetically informed cohorts previously recruited for genome-wide association studies (GWAS) and twin studies. Participants for the present study represent a subset of mid to late life adults without a diagnosis of dementia (see Table 1) who completed both online cognitive assessments and sleep questionnaires described below. Where twin pairs were included in the dataset, one twin was excluded at random.
Characteristics of study sample stratified by napping habits.
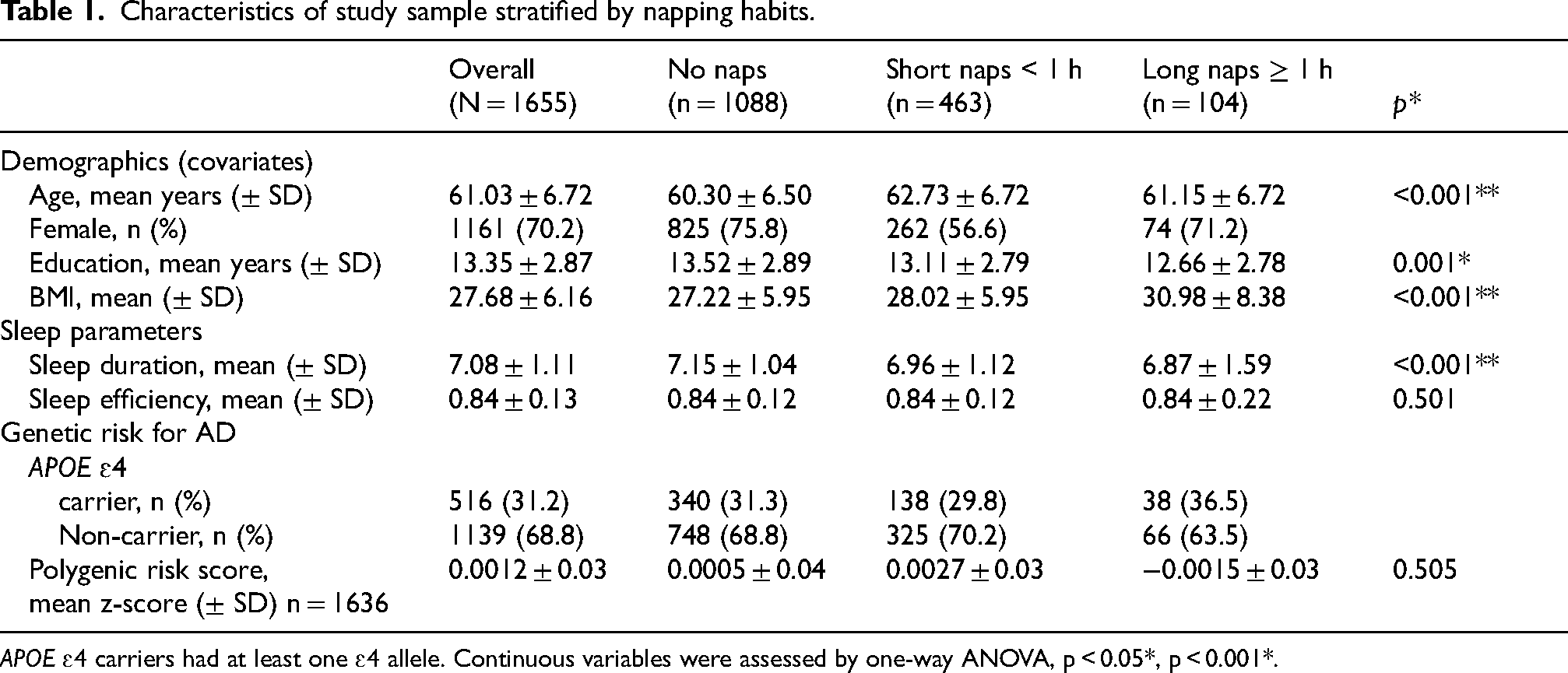
APOE ɛ4 carriers had at least one ɛ4 allele. Continuous variables were assessed by one-way ANOVA, p < 0.05*, p < 0.001*.
Demographic data, including date of birth, sex, body mass index (BMI) and years of education, was obtained via self-reported online questionnaires. BMI was calculated using self-reported height and weight. Age at the time of data acquisition was derived from dates of birth. Years of education was extrapolated from highest level of tertiary education.
Genetic data
Genome-wide genotyping of participants was performed using a range of genotyping arrays.38,39 Datasets were combined with strict quality control procedures and imputed to the Haplotype Reference Consortium (HRC) Release 1 reference panel. 40 APOE genotype was derived from SNPs rs429358 and rs7412, and was obtained from genome-wide SNP chip data, where either the APOE SNPs were directly genotyped on the array or imputed with a high degree of certainty, or directly genotyped using TaqMan SNP genotyping assays on an ABI Prism 7900HT and analyzed using SDS software (Applied Biosystems), as previously described. 41
PRS are the weighted sum of genetic risk alleles across the genome, with their weights derived from the results of GWAS. We calculated the PRS using GWAS results summary statistics for AD from Bellenguez et al. (2022), which is one of the largest AD GWAS meta analyses available including a large number of clinically defined AD cases that has been widely validated for PRS construction. 6 Single-nucleotide polymorphisms (SNPs) of low imputation quality (r2 < 0.6), low allele frequency (MAF<0.1), non-autosomal and strand ambiguous were excluded. SNPs within 500 kb distance at either side of the APOE locus were excluded to ensure the exclusion of the APOE-associated signal, as APOE would otherwise dominate the score, masking the contribution of other variants. The PRS was calculated using PLINK. 42 SNPs were clumped to account for LD using an r2 value of 0.1 and distance of 10000 kb (PLINK 1.90). Using the SNP p-value significance threshold p < 0.5 the AD PRS was calculated by multiplying the dosage score and effect size for each SNP, and these values were summed across all loci (PLINKv2). A linear regression was performed of PRS on 4 ancestry principal components derived from the GWAS data, and two variables for imputation batches, and the standardized residual used in the subsequent analysis. The PRS was split into quintiles where the lowest quintile was classed as low genetic risk and the highest quintile classed as high genetic risk.
Sleep assessment
To be included in the sample, participants were required to have both self-reported daytime napping duration and a completed Pittsburgh Sleep Quality Index questionnaire (PSQI). 43 Self-reported online questionnaires were used to assess sleep parameters. Habitual daytime napping was assessed using the question: “How many hours do you spend napping in a typical day?”. Answers could range from none to >6 h. Participants selected one of; none, less than 1 h, 1–2 h, 2–3 h, 3–4 h, 4–5 h, 5–6 h, or more than 6 h. Napping duration data was segmented in three categories: no napping, short nap (<1 h), long nap (≥)1 h). Only one participant reported napping > 3 h.
Sleep duration and efficiency were derived from the PSQI. 43 Due to the constraints of the PISA multi-choice questionnaire, participants rounded their answers to the nearest time approximation. Four participants who reported sleeping less than 4 h were recoded as 3.5 h and one participant reporting sleeping > 12 was recoded as 12.5 to allow for the continuous data in line with the pattern from multiple choices. The same approach was taken with sleep efficiency. Those reporting a wake-up time earlier than 5am were recoded as 4.30am (n = 75), and after 11am were recoded as 11:30am (n = 5). Participants reporting a bedtime earlier than 8pm were recoded as 7:30pm (n = 16), and those reporting a bedtime after 3am were recorded as 3:30am (n = 2). Self-reported sleep duration and efficiency (i.e., sleep duration divided by the duration between reported bedtime and wake time) were included in participant characteristics, T-test analyses, with sleep efficiency added as a covariate in the main model. A sensitivity analysis for the main findings was conducted without these participants to ensure that recoding did not affect the results.
Cognitive assessments
Cognition was assessed using the self-administered online battery Creyos, previously known as Cambridge Brain Sciences (https://creyos.com). The Creyos battery includes 12 neurocognitive tasks that evaluate cognitive domains such as memory, reasoning, verbal ability, and concentration (see Supplemental Table 1 for test descriptions). The battery took approximately 30 min to complete. Higher scores indicate better performance. Cognitive tests were z-scored. Participants with more than 50% data missing were excluded. A mean substitution approach was used for imputation of missing data in the remaining participants.
Statistical analyses
Using IMB SPSS v29, we conducted a Principal Component Analysis (PCA) to reduce the dimensionality of the Creyos cognitive data (z-scored derived from raw data for 12 subtests) and identify components to be used in subsequent analyses. A Varimax rotation was used to produce uncorrelated components and to maximize the variance of the squared loadings on each factor, with Kaiser normalization applied for better interpretability.44,45 Analyses of covariance (ANCOVA) were used to determine the main effects and interactions between self-reported napping duration (independent variable; IV) and genetic data (IV) on cognitive abilities (dependent variable; DV). APOE ɛ4 and PRS were investigated separately in the ANCOVA (i.e., 3 models for each genetic risk factor, see Tables 2 and 3 for details of each main models) to ensure that the strong effects typically observed with APOE would not mask any small contribution of PRS. Age, years of education, sex, and BMI were added as covariates. Sleep efficiency was also treated as a covariate in follow-up ANCOVA to account for the 24-h sleep cycle and as detailed in results. Additional ANCOVA were conducted on the components where interactions were observed to further explore the independent main effect of each predictor (i.e., IVs) on cognition. APOE was also added as a covariate in the ANCOVA with PRS as an IV in further analyses and as described in results. BMI was used as a covariate given its association with sleep apnea, and evidence showing that OSA is often underdiagnosed in older adults.46,47 Mean imputation for missing data was used for the BMI of 29 people and education of 7 people. Additional analyses were conducted on the DV components where interactions were observed. First, the independent main effect of each IVs on the DV were assessed via ANCOVA. Additive interactions between genetic risk for AD and napping were assessed using interaction contrasts (IC), computed from adjusted marginal means. The IC was defined as IC = (Y11 – Y10) – (Y01 – Y00), where Yab denotes the predicted mean memory score for genetic status a (0 = APOE ɛ4 non-carrier or low PRS, 1 = APOE ɛ4 carrier or high PRS) and napping category b. Positive IC values indicate that the combined effect of genetic risk and napping exceeds the sum of their independent effects.
Results from ANCOVA with APOE ɛ4 and napping duration.
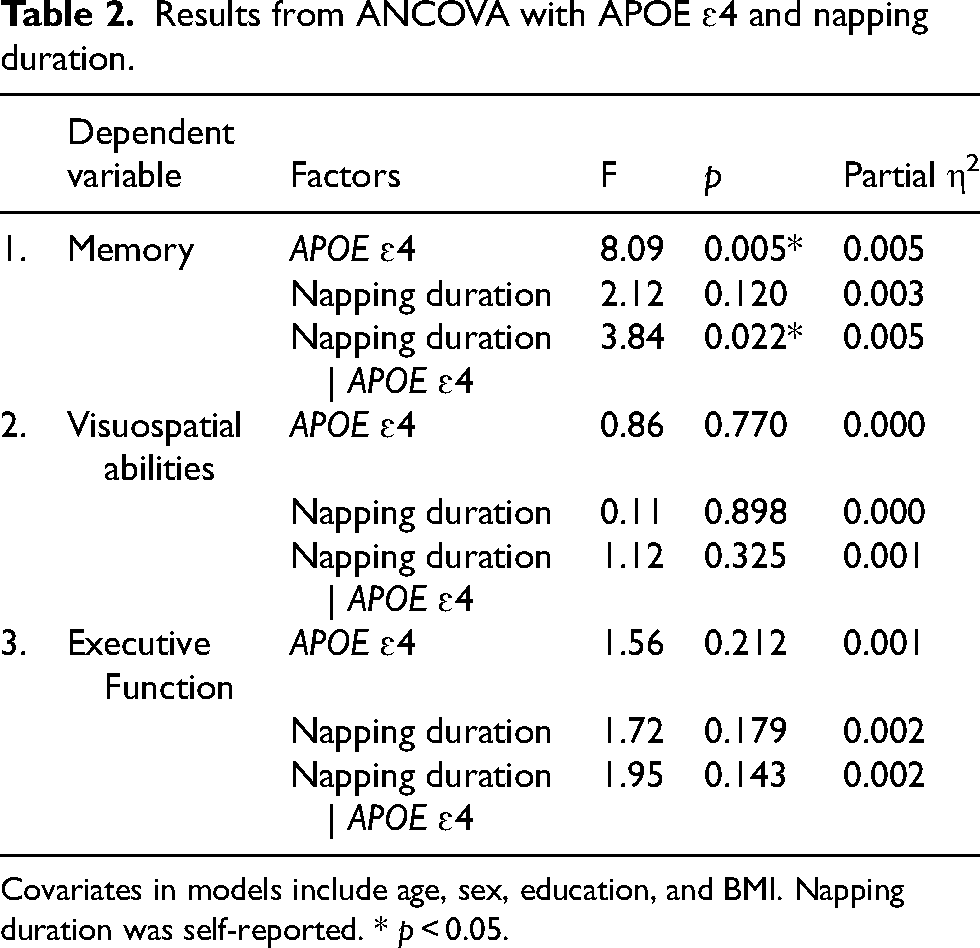
Covariates in models include age, sex, education, and BMI. Napping duration was self-reported. * p < 0.05.
Results from ANCOVA with AD PRS and napping duration.
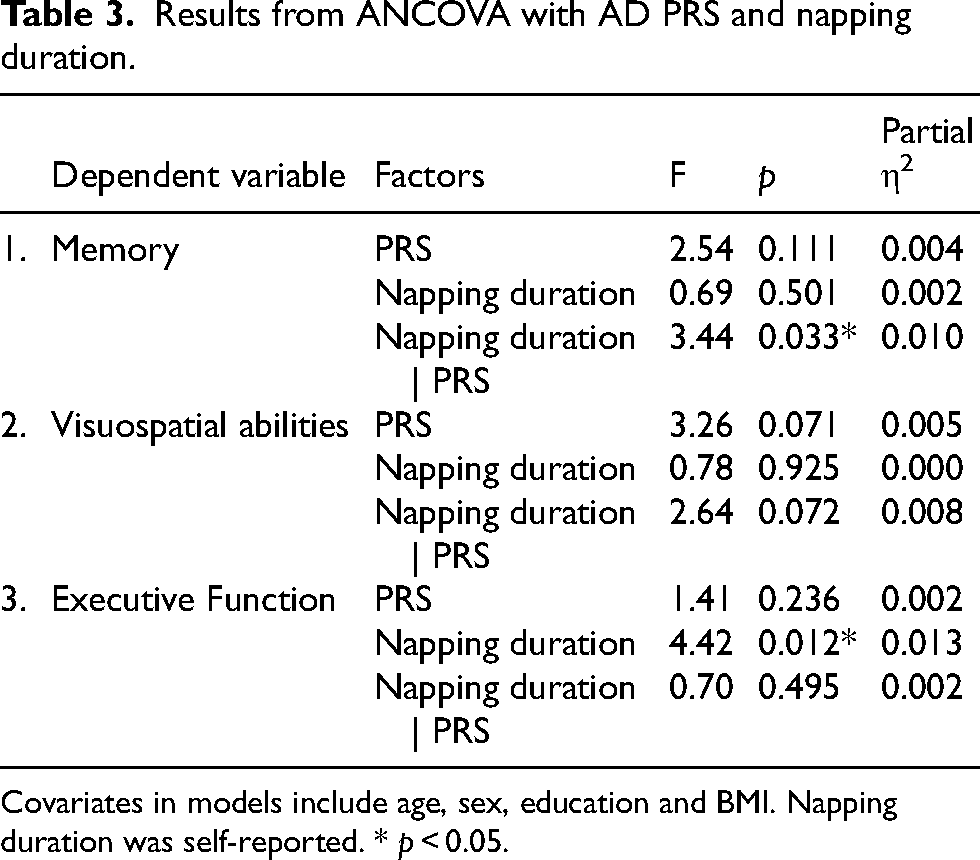
Covariates in models include age, sex, education and BMI. Napping duration was self-reported. * p < 0.05.
Bonferroni corrections for multiple comparisons were used for simple main effects analyses. Independent samples Mann-Whitney U Tests were used to investigate the difference between APOE ɛ4 carriers and non-carriers, and between low and high PRS, on the napping categories. Independent t-tests were used to investigate the differences in sleep duration and efficiency between APOE ɛ4 carriers and non-carriers, as well as between low and high PRS. Python 3.11.4 and R Studio version 2024.04.1 + 748 were used for figures.
Results
A total of 1655 participants aged 42 to 75 years old were included in the study, with a mean age of 61.03 years. Most participants were female (70.2%). The average BMI was 27.7, which is classified as overweight (Seidell, 1997). 8.2% of participants reported a diagnosis of obstructive sleep apnea. Nearly half of the participants reported having had sleep issues in the past 12 months (47%), with 13.9% of participants reporting using sleep medication. On average, participants reported sleeping 7.07 h per night, with a mean efficiency of 0.84. When sleep quality was calculated using the PSQI, over half of the participants (60.5%) were categorized as good sleepers. For genetic risk of dementia, 31.2% of participants were APOE ɛ4 carriers. Of these, 7.05% (i.e., 2.2% of the full sample) were homozygotes, with two copies of the APOE ɛ4 allele, while the rest were heterozygotes, with only one APOE ɛ4 allele. Demographics, sleep and genetic characteristics presented by categories of reported napping durations are summarized in Table 1.
In the PCA analysis, the spree plot and eigenvalues cut-off of one suggested three potential components, explaining a cumulative 44.2% of the variance (Supplemental Table 2). The Bartlett's Test of Sphericity was significant (p < 0.001) and Kaiser-Meyer-Olkin measure of sampling adequacy was of 0.867 indicating the data was suitable for factor analysis. Table 4 shows the Rotated Component Matrix with loadings. The three components (visuospatial, memory and executive functions) were labelled based on variables with loadings above 0.5 contributing to each component.
Results from PCA in rotated component matrix for cognitive tests.
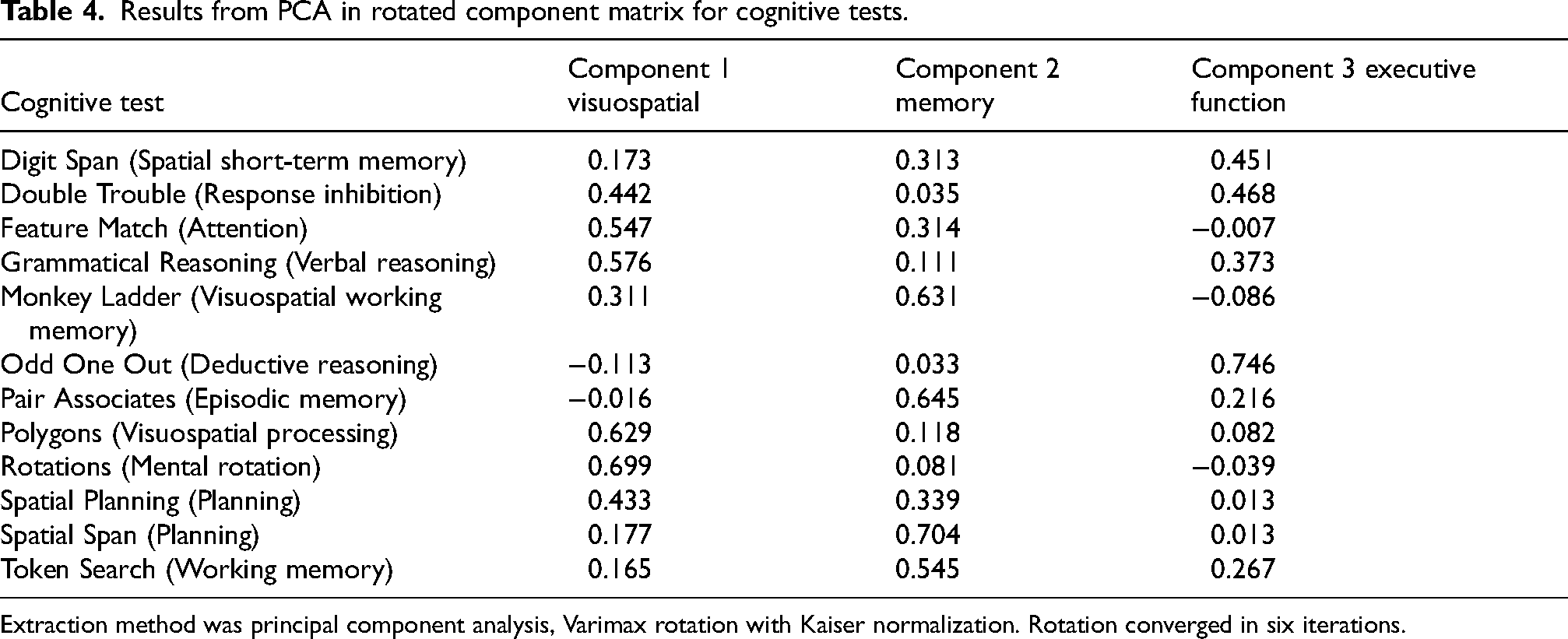
Extraction method was principal component analysis, Varimax rotation with Kaiser normalization. Rotation converged in six iterations.
Associations of napping and APOE ɛ4 with cognition
ANCOVAs with APOE ɛ4 carrier status and reported napping duration as factors and age, sex, BMI as covariates were computed for all three cognitive components (Table 2). There was a significant main effect of APOE ɛ4 (F(1,1645) = 8.09, p = 0.005, ηp2 = 0.005), and significant interaction with napping (F(2,1645) = 3.84, p = 0.022, ηp2 = 0.005) on the memory component (Figure 1(a)), but no significant main effect of napping (p = 0.120). Simple main effects analyses indicated that APOE ɛ4 carriers who were long nappers (M = 0.32, SE = 0.16) had higher memory performance (p = 0.003) than APOE ɛ4 non-carriers who napped for the same duration (M = -0.27, SE = 0.12). In APOE ɛ4 carriers, long nappers (M = 0.32, SE = 0.16) significantly outperformed (p = 0.048) short nappers (M = -0.08, SE = 0.08).
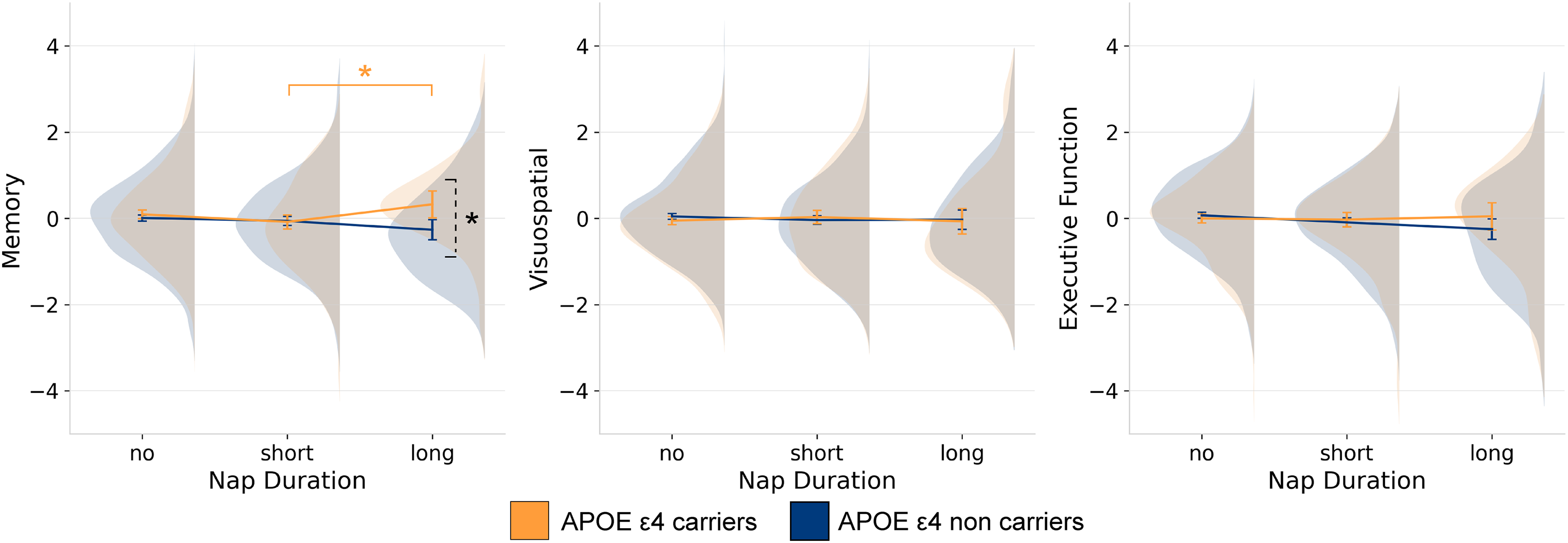
Interactions of APOE ɛ4 and napping duration on cognitive variables. Results from ANCOVA showing the interaction of APOE ɛ4 and self-reported habitual napping on each of the cognitive components resulting from the PCA analysis. Cognition is represented with marginalized means. The dashed bracket represents a significant difference between APOE ε4 groups, while the solid bracket indicates a significant difference within a group. *p ≤ 0.05, **p ≤ 0.001.
In contrast, results showed no significant main effects of APOE ɛ4 (p = 0.770), napping (p = 0.898) or interaction (p = 0.325) on the visuospatial component (Figure 1(b)). Similarly, there was no significant main effects of APOE ɛ4 (p = 0.212), napping (p = 0.179), or interaction (p = 0.143) on the executive function component (Figure 1(c)).
When subjective sleep efficiency was added to consider night time sleep in the model as a covariate, we also found a significant main effect of APOE ɛ4 (F(1,1644) = 7.40, p = 0.005, ηp2 = 0.005), and the interaction with napping (F(2,1644) = 3.78, p = 0.023, ηp2 = 0.005) on the memory component. Results for the other components remained non-significant.
Additionally, when factors were investigated alone neither napping (p = 0.150), APOE ɛ4 status (p = 0.079) alone had a significant main effect on the memory component, indicating that each predictor alone doesn’t show a clear influence on memory. The additive interaction assessing whether the combined effect of APOE and napping on memory was greater than the sum of individual effects showed significant positive additive interactions of long versus short naps (IC = 0.61, CI [0.19, 1.04]) and long versus no naps (IC = 0.50, CI [0.12, 0.89]). These findings suggest that the memory benefits are amplified in APOE ɛ4 carriers who take long naps.
Associations of napping and AD PRS with cognition
ANCOVAs with AD PRS (top and bottom quintiles) and reported napping duration as factors and age, sex, BMI as covariates were also computed for all three cognitive components (Table 3). There were no main effects of napping (p = 0.501) or AD PRS (p = 0.111) on the memory component. However, there was a significant interaction (Figure 2(a)) of PRS and napping duration (F(2653) = 3.44, p = 0.033, ηp2 = 0.010). Further analyses showed a significant primary effect within the no napping group (p = 0.007), where memory performance was higher in the low-risk (M = 0.12, SD = 0.06) than in the higher-risk group (M = -0.13, SD = 0.68). Additionally, we found that not napping (M = 0.12, SD =0.06) was significantly associated with better memory than short naps (M = -0.18, SD = 0.10), but only in the low-risk group (p = 0.049). Another analysis with PRS alone as an IV showed that PRS did not have a significant main effect (p = 0.051) on the memory component. The additive interaction analysis showed a significant positive additive interaction of short versus no naps (IC = 0.40, CI [0.08, 0.73]), indicating that those with high PRS gain more cognitive benefit from short naps than expected from the sum of their separate effects. Taken together, these findings suggest that short naps may be detrimental for people with low genetic risk for AD but may provide a relative memory benefit for those at higher polygenic risk.
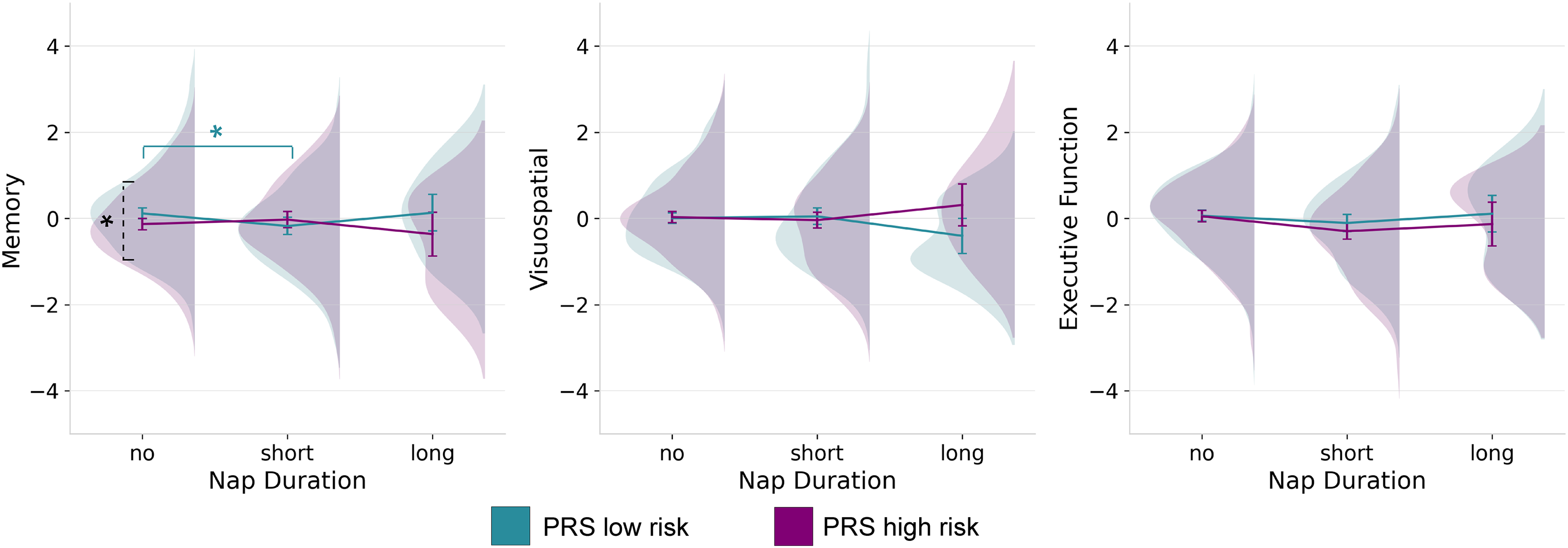
Interactions of PRS and napping duration on cognitive variables. Results from ANCOVA showing the interaction of PRS and self-reported habitual napping on each of the cognitive components resulting from the PCA analysis. Cognition is represented with marginalized means. The dashed bracket represents a significant difference between PRS groups, while the solid bracket indicates a significant difference within a group. *p ≤ 0.05, **p ≤ 0.001.
Napping had a main effect (F(2, 653) = 4.42, p = 0.012, ηp2 = 0.013) on the executive function component (Figure 2(c)); however, the main effect of AD PRS (p 0.236) and its interaction with napping (p = 0.495) were not significant. Main effects of napping (p = 0.925), AD PRS (p = 0.071), and interactions (p = 0.072) on the visuospatial component (Figure 2(b)) were not significant.
When adding sleep efficiency to the AD PRS model, the interaction of napping and AD PRS on the memory component (F(2, 652) = 3.46, p = 0.032, ηp2 = 0.011) and the main effect of napping on executive function (F(2, 652) = 4.44, p = 0.012, ηp2 = 0.013) remained significant. All other associations remained non-significant. In separate analyses, APOE ɛ4 was added as a covariate in the main model, but was not significant and did not change the significance of the interaction between PRS and napping (p = 0.033) on memory or the main effect of napping on executive function (p = 0.012).
Exploring the full sleep cycle
Independent samples Mann-Whitney U Tests investigating the difference between APOE ɛ4 carriers and non-carriers on the napping categories (p = 0.897) were not significant. Independent t-tests looking at differences in sleep duration (p = 0.520) and efficiency (p = 0.288) between APOE ɛ4 carriers and non-carriers were also not significant. Independent samples Mann-Whitney U Tests investigating the difference between low and high PRS on the napping categories (p = 0.526) were not significant. Independent t-tests showed that low and high AD PRS did not significantly have different sleep duration (p = 0.544) and efficiency (p = 0.353).
Discussion
Previous research has suggested a non-linear relationship between daytime napping and cognition, where longer naps have been associated with a higher risk of AD. 29 In contrast, short naps of less than 30 min were observed to reduce the risk of cognitive decline in older adults. 28 However, very few studies have investigated the influence of genetic risk of AD on the napping-cognition relationship. In this cross-sectional study, we investigated both APOE ɛ4 (as the strongest genetic risk factor for AD) and additional risk variants combined in a PRS, allowing for a more comprehensive examination of the AD risk spectrum. We found that the association of napping and memory is moderated by genetic risk.
APOE ɛ4 carriers who reported habitual long naps were more likely to have better memory scores compared to those taking shorter naps, and to non-carriers who reported long naps. This finding is in line with the antagonistic pleiotropy hypothesis, which suggests that APOE ɛ4 may have various effects on cognitive health throughout the lifespan, including being beneficial in midlife. 11 As such, the memory advantage observed for APOE ε4 carriers in the present study may be related to the younger age of our sample. Our findings are also consistent with a specific advantage in memory, which was previously observed in APOE ɛ4 carriers in midlife.48,49 The significant interaction between napping and APOE ɛ4 on memory did not change after accounting for overnight self-reported sleep efficiency, suggesting that long daytime sleep may potentially function as an enabler for APOE ɛ4 carriers to reap the memory benefits.
The interaction results were specific to the memory domain across the genetic risk spectrum, which may be due to episodic memory being particularly affected in the early stages of the disease. 50 While longer naps benefited APOE ɛ4 carriers, participants with a low PRS benefited from not napping compared to napping for short periods. These differences could be explained by factors related to the timing and mechanisms behind the genetic effects. APOE ε4 effects have been shown to occur early in the disease process at the amyloid deposition stage. Here, APOE ɛ4 carriers at a high genetic risk of AD are more likely to be in an early prodromal disease phase, when long naps could be beneficial to memory performance. In contrast, polygenic risk for AD, outside of APOE, may have more effect later in the disease process in the progression to tau pathology and be more linked to neuronal degeneration rather than AD-related amyloid pathology. 51 In the present study, those with low PRS are at the lowest genetic risk of AD later in life. As such, short napping may disturb the memory consolidation process in healthy sleep patterns.
Future directions and limitations
In this study, habitual daytime and nighttime sleep were assessed through self-report, which limits the reliability of the data due to potential recall bias. Additionally, self-report sleep limits the ability to explore more intricate details of sleep. As discussed earlier, no significant differences were observed between APOE ɛ4 carriers and non-carriers, or between low and high PRS groups, indicating similar sleep patterns. Nonetheless, napping may differentially influence the macro and micro-architecture of overnight sleep, or vice versa, depending on AD genetic profiles, which may in turn affect memory performance. Evidence points to an essential role of non-rapid eye movement (NREM) sleep spindles in memory consolidation,52–54 as well as a deficit in NREM spindles in neurodegenerative disorders. 55 A recent meta-analysis indicated that naps may involve spindle-related consolidation mechanisms similar to those observed during whole-night sleep. 52 Additionally, recent findings suggest that time spent in rapid-eye movement (REM) sleep is reduced in late middle-aged and older APOE ɛ4 carriers, likely due to early neurodegeneration in regions involved in REM. 56 Taken together, changes in sleep stages associated with AD may also vary as a function of genetic risk, and this relationship may also change over time. Replicating the current study using objective measures of sleep with a larger age range would assist in better understanding the mechanisms underlying our findings. This would also allow for further exploration of the possible antagonistic pleiotropy effect on cognition. Napping could be a marker of changes in sleep homeostasis at one point in time, but also a modifiable factor promoting independent memory benefits. Investigating intentional versus unintentional napping could also assist in testing this hypothesis.
We also acknowledge that data used in the study was analyzed retrospectively, which restricted the choice of categories for reported sleep and napping duration. For instance, narrower napping category ranges would allow to test for the possibility of non-linear patterns. The data-driven approach to derive the memory domain using PCA incorporates other domains, which may limit interpretation. Moreover, we noted the heterogeneity in previous results and the limited body of research around our specific questions, making it challenging to compare our findings. Finally, the cross-sectional design limited our ability for causal inference and directionality in the relationships between napping, genetic risk and cognition. Despite these limitations, our study provides a foundation for further studies into the interplay of genetic risk, daytime napping and cognition.
Conclusion
Despite significant advances in our understanding of AD and its pharmacological treatments, effective prevention continues to be elusive. This, combined with a rising popularity in genetic testing 36 highlights the need for a better understanding of modifiable factors that could play a key role in personalized interventions, particularly in at-risk populations. Our findings suggest that the relationship between napping and memory may vary as a function of genetic risk for AD. Although more work is needed to elucidate the mechanisms underlying these relationships, these findings could inform personalized preventative treatment for AD based on genetic risk profiles. This is particularly relevant given the increasing access to genetic testing, especially APOE genotyping services. Our findings do not support a one-size fits all sleep advice that suggests that short naps may be beneficial for health while long naps may be detrimental, including for cognitive abilities.26,57 Within APOE ε4 carriers, there was a memory advantage of long naps compared to short naps. Similarly, at the lower end of the PRS spectrum of AD risk, we found that no naps were associated with better memory outcomes than short naps. This highlights the importance of factoring genetic risk for AD when providing health advice on napping.
Supplemental Material
sj-docx-1-alz-10.1177_13872877261427707 - Supplemental material for The relationship between napping and memory varies as a function of genetic risk for Alzheimer's disease
Supplemental material, sj-docx-1-alz-10.1177_13872877261427707 for The relationship between napping and memory varies as a function of genetic risk for Alzheimer's disease by Caroline Faucher, Renate Thienel, Anna Behler, Lina Gomez, Jurgen Fripp, Michael Breakspear and Michelle K. Lupton in Journal of Alzheimer's Disease
Footnotes
Acknowledgements
The Prospective Imaging Study of Aging: Genes, Brain and Behaviour (PISA) was funded by a National Health and Medical Research Council (NHMRC) Boosting Dementia Research Initiative—Team Grant [APP1095227]. We would like to acknowledge and thank all the participants who took part in the study, and the PISA team members working on recruitment and data collection. We also acknowledge the contribution of Professor Nick Martin at QIMR Berghofer, who has led genetic epidemiology studies over the past 40 years, from which the PISA population-based sample was recruited.
Ethical considerations
Ethics approval was obtained from the Human Research Ethics committees of QIMR Berghofer (P2210).
Consent to participate
Participants provided written informed consent prior to participating in the Prospective Imaging Study of Aging: Genes, Brain and Behaviour (PISA) and consented for anonymized information to be published in articles. Participation was voluntary.
Consent for publication
Not applicable
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the The Australian Government, Commonwealth Scientific and Industrial Research Organisation, (Research Training Program (RTP) Scholarship, postgraduate studentship program).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
Following completion of each wave (baseline, follow-up) and appropriate quality control, de-identified PISA data will be made available to other research groups upon request. Due to privacy, confidentiality and constraints imposed by the local Human Research Ethics Committee, a “Data Sharing Agreement” will be required before data will be released. Due to ethics constraints, data will be shared on a project-specific basis. Depending on the nature of the data requested, evidence of local ethics approval may be required.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.