
Abstract
Background
Alzheimer's disease (AD) is a neurodegenerative disorder and the most common cause of dementia. The integrated stress response (ISR) contributes to impaired synaptic plasticity, neuronal dysfunction, and cognitive deficits in AD. However, research targeting the ISR as a therapeutic strategy for AD remains limited due to insufficient mechanistic insight.
Objective
This study aimed to evaluate the effects of 2BAct, an ISR inhibitor, on behavioral symptoms, amyloid-β (Aβ) and tau accumulation, and neuroinflammation in 5xFAD mice.
Methods
Ten-month-old 5xFAD mice received daily intraperitoneal (IP) injections of either 2BAct (10 mg/kg/day), donepezil (2 mg/kg/day; positive control), or vehicle for 23 consecutive days. Anxiety-like behavior and cognitive function were assessed using the open field test (OFT), novel object recognition test (NORT), and Morris water maze (MWM). Amyloid-β (Aβ), tau, and neuroinflammation markers were analyzed by immunofluorescence staining. ISR inhibition was evaluated by examining the phosphorylation level of eukaryotic initiation factor 2 alpha (eIF2α) using immunofluorescence staining and by analyzing ISR-related markers via RNA sequencing.
Results
2BAct treatment significantly improved object recognition performance and attenuated microglial activation and tau accumulation, without reducing Aβ burden. Reduced levels of phosphorylated eIF2α were also confirmed by immunofluorescence staining.
Conclusions
These findings suggest that 2BAct treatment improves cognitive performance and mitigates neuroinflammation while reducing tau accumulation. Although the therapeutic effects are limited, targeting the ISR with inhibitors such as 2BAct represents a potential therapeutic approach for AD. Further studies are required to elucidate the underlying molecular mechanisms and to address the limitations of ISR-based interventions.
Keywords
Introduction
Alzheimer's disease (AD) is a debilitating neurodegenerative disorder and the most common cause of dementia, currently affecting over 50 million people worldwide, with this number projected to triple by 2050 due to population aging. 1 Clinically, AD manifests as progressive memory impairment, cognitive decline, and behavioral changes, severely compromising the quality of life for both patients and caregivers.2,3 Pathologically, AD is characterized by the accumulation of extracellular amyloid-β (Aβ) plaques, intracellular neurofibrillary tangles composed of hyperphosphorylated tau protein, synaptic loss, neuronal degeneration, and pronounced neuroinflammation.4–6 Despite extensive research, effective therapeutic strategies targeting the underlying pathology remain limited, and current medications primarily provide symptomatic relief without altering disease progression. 4
Emerging evidence has highlighted the integrated stress response (ISR) as a critical pathway involved in neurodegeneration7–16 and cognitive impairment associated with AD.7,8,10,17–20 The ISR is a conserved cellular mechanism activated by various stressors, including oxidative stress, protein misfolding, and inflammation.21–23 Activation of the ISR leads to phosphorylation of eukaryotic initiation factor 2 alpha (eIF2α) by four kinases (PERK, GCN2, HRI, and PKR). This process reduces global protein synthesis while allowing selective translation of stress-response proteins. While this response is initially protective, sustained and aberrant ISR activation—particularly elevated phosphorylation of eIF2α—has been consistently associated with impaired synaptic plasticity, neuronal dysfunction, and cognitive deficits in AD12,19,20,24 and is correlated with disease severity and progression in postmortem AD brain tissues.
Supporting this notion, knockdown of PERK, a kinase responsible for eIF2α phosphorylation, alleviates synaptic plasticity impairments and spatial memory deficits in AD mouse models. 19 Similarly, pharmacological modulation of the ISR with the ISR inhibitor ISRIB reduces levels of ATF4, a key downstream mediator of eIF2α signaling, thereby restoring impaired synaptic plasticity and memory in AD models. 20 Collectively, these findings suggest that excessive ISR activation contributes to neuronal dysfunction and provide a rationale for targeting this pathway in AD.
A central regulator of ISR signaling is eukaryotic translation initiation factor 2B (eIF2B), a guanine nucleotide exchange factor (GEF) that controls the activity of eIF2 during the initiation of mRNA translation. 25 By promoting the exchange of GDP for GTP on eIF2, eIF2B enables the formation of the translation initiation complex required for protein synthesis.25–28 Under stress conditions, phosphorylation of eIF2α inhibits eIF2B function, leading to suppression of global protein synthesis while selectively promoting translation of stress-related transcripts.29–31 Because chronic ISR activation and sustained translational repression have been implicated in neurodegenerative diseases, modulation of eIF2B activity has emerged as a promising strategy to restore translational homeostasis and neuronal resilience during cellular stress.
Consistent with the therapeutic promise of targeting eIF2B, recent small-molecule activators such as ISRIB and 2BAct have demonstrated robust efficacy in restoring impaired translational control by suppressing ISR-mediated inhibition of eIF2B.11,32,33 ISRIB, a well-characterized eIF2B activator, has shown potent effects in restoring protein synthesis rates, alleviating cognitive impairments, and ameliorating synaptic dysfunction across multiple animal models of neurodegenerative diseases, including AD. Similarly, 2BAct, a structurally distinct eIF2B activator featuring a bicyclo[1.1.1]pentyl core structure, is notable for being the first orally bioavailable compound in its class, possessing favorable pharmacokinetic properties and proven in vivo efficacy. In a preclinical model of vanishing white matter (VWM) disease, 2BAct effectively prevented disease progression, suppressed pathological ISR signaling, and normalized transcriptomic and proteomic alterations. 32 Moreover, ISRIB restored eIF2B activity and rescued pathological phenotypes in mouse models harboring mutations such as R195H in the eIF2Bε subunit. 33 Despite the accumulating evidence highlighting the therapeutic potential of these eIF2B-targeting compounds, the efficacy of 2BAct in the context of Alzheimer's disease remains insufficiently explored, warranting comprehensive preclinical evaluation.
In the present study, we investigated whether 2BAct could ameliorate cognitive deficits and pathological features in the widely used 5xFAD transgenic mouse model of AD. This mouse model exhibits aggressive amyloid pathology, tau hyperphosphorylation, neuroinflammation, and cognitive impairments that recapitulate key features of human AD. Our findings reveal that chronic administration of 2BAct significantly improves recognition memory deficits while also modulating ISR signaling, tau pathology, and microglial activation. This study provides important insights into the therapeutic potential of ISR modulation and highlights eIF2B activators such as 2BAct as promising candidates for further preclinical investigation toward disease-modifying strategies in AD.
Methods
Animals
5xFAD transgenic mice were from the Jackson Laboratory (RRID:MMRRC_034848-JAX) and were continuously inbred to maintain a stable line until the required number of animals was reached for the experiments. Male heterozygous 5xFAD transgenic mice and wild-type (WT, C57BL/6J) littermate controls aged 7 months were used for all behavioral and histological experiments. Behavioral experiments were performed with 8 animals per group, whereas histological analyses were conducted using 4–5 animals per group. In total, 32 mice were used for behavioral experiments and 16–20 mice were used for histological analyses. Animals were randomly assigned to experimental groups prior to drug administration and behavioral testing. 5xFAD transgenic mice overexpress three mutations, Swedish (K670N, M671L), Florida (I716 V), and London (V717I), in the human amyloid beta precursor protein (APP) and two mutations, M146L and L286 V, in human PS1. Homozygous 5xFAD mice exhibited significant weight loss and abnormal behaviors, such as paw clasping, by 5 months of age compared to WT and heterozygous mice. Due to the severe pathology and reduced survival rates observed in 5xFAD homozygous mice, heterozygous 5xFAD mice were utilized for experiments to ensure adequate viability and experimental consistency. 34
All animals were housed in the Yeungnam University Experimental Animal Center under controlled conditions (12-h light/dark cycle, lights on from 08:00 to 20:00; humidity: 50 ± 5%; temperature: 22 ± 1 °C). Food and water were provided ad libitum, except during behavioral testing. All experimental procedures involving animals were conducted in accordance with the guidelines of Yeungnam University Institutional Animal Care and Use Committee (IACUC) and were approved under protocol number 2023-021.
Mouse genotyping
We used three sets of primers for polymerase chain reaction (PCR): Common Forward (5'-ACCCCCATGTCAGAGTTCCT-3’), WT Reverse (5'-TATACAACCTTGGGGGATGG-3’), and Mutant Reverse (5'-CGCGCCTCTTCGCTATTAC-3’). Standard PCR was performed using AccuPower Taq PCR PreMix (Bioneer, K-2602) and DNA extracted from tail snips. The PCR products were loaded onto a 1% agarose gel and analyzed by electrophoresis. A single band at 216 bp indicated a WT genotype, while two bands at 129 bp and 216 bp indicated a heterozygous 5xFAD genotype.
Drug preparation
The vehicle solution was prepared using 5% DMSO (Sigma, D8418) and 95% corn oil (Sigma, C8267). Donepezil and 2BAct were first dissolved in DMSO to prepare stock solutions and subsequently diluted with corn oil to achieve final concentrations of 0.2 mg/ml and 1 mg/ml, respectively. For vehicle controls, 5% DMSO was mixed with 95% corn oil to ensure consistency with the experimental conditions. DMSO stock solutions for all drugs were used within one week of preparation, and injection solutions were freshly prepared daily prior to use. Donepezil and 2BAct were generously provided by the ALBORADA Drug Discovery Institute.
Injection procedure
All drugs and chemicals were administered intraperitoneally (IP) using 1 mL Kovax syringes (Koreavaccine, A047242). Injections were performed using a sterile needle inserted at a slight angle into the central lower abdomen, just below the umbilical region, to minimize the risk of puncturing internal organs. The injections were administered daily at the same time, with doses adjusted to 2 mg/kg for Donepezil and 10 mg/kg for 2BAct according to the individual body weight of each mouse. The injections commenced two weeks prior to the start of the behavioral tests.
Pharmacokinetic analysis and extrapolations
To support pharmacokinetic characterization, 2BAct was administered intraperitoneally (10 mg/kg) to male C57BL/6 mice (8–10 weeks old). Blood samples were collected at 0.25, 0.5, 1, 2, 4, 8, 12, and 24 h post-dose (n = 3 mice per time point, sparse sampling) via retro-orbital sinus puncture into heparinized tubes. Samples were gently mixed and kept on ice prior to centrifugation. Plasma was separated within 30 min by centrifugation at 4000 rpm for 10 min at 4°C and stored at −70 ± 10°C until analysis.
At designated time points, mice were euthanized immediately following blood collection by cervical dislocation, and brain tissues were rapidly extracted within 2–3 min post-mortem to minimize degradation. Brains were carefully removed, briefly rinsed in ice-cold saline to remove residual blood, blotted on filter paper, weighed, and snap-frozen prior to storage at −70 ± 10°C. For analysis, brain tissues were homogenized in phosphate-buffered saline (PBS) at a ratio of 1:3 (w/v) under cold conditions using a mechanical homogenizer to ensure consistent tissue disruption.
Concentrations of 2BAct in plasma and brain homogenates were quantified using a fit-for-purpose LC-MS/MS method with a calibration range of 10–10,000 ng/mL. Pharmacokinetic parameters were calculated using non-compartmental analysis (Phoenix WinNonlin, version 8.3, Certara). All sample collection, processing, and storage procedures were performed in accordance with standardized protocols described in the pharmacokinetic study report to minimize variability across biological matrices.
Pharmacokinetic extrapolations were guided by prior data from Wong et al. (2019), who reported that 2BAct achieves effective brain exposure with an unbound brain-to-plasma ratio of ∼0.3 and unbound brain levels ∼15-fold above the in vitro EC50 at a 30 mg/kg oral dose. Given the higher systemic exposure associated with intraperitoneal administration, our 10 mg/kg IP regimen was expected to yield similar central concentrations (∼0.8–1 µM). As shown in Supplemental Figure 1, brain levels reached 3650 ± 1300 ng/g at 0.5 h (∼8.3 µM, based on molecular weight 440.8 g/mol) 35 and remained above the EC50 for several hours, supporting target engagement.
Open field test
The open field test (OFT) is widely used due to its quick and relatively simple procedure, providing a range of behavioral data from general locomotor activity to emotional aspects such as anxiety in experimental animals. 36 The experiment was conducted in a square acrylic arena with white walls and floor (40 × 40 cm), and a ceiling-mounted camera connected to a computer was used for recording and analysis of the trials. Each mouse was placed at the edge of the arena and allowed to freely explore the area for 10 min. Before each trial, the arena was sanitized with alcohol and allowed to dry completely for at least 3 min before proceeding with the experiment. The entire movement of each mouse was recorded, and parameters such as total distance moved and duration in the center (20 × 20 cm) were measured and analyzed using EthoVision XT 11.5.
Novel object recognition test
The novel object recognition test (NORT) is one of the most commonly used behavioral tests in rodents, providing an index of recognition memory. 37 The experiment was conducted in a square acrylic arena (white walls and floor, 30 × 30 cm), and a ceiling-mounted camera connected to a computer was used for recording and analysis of the trials. On the first day (Habituation), which coincided with the OFT, the mice were placed in an empty open field and allowed to freely explore. On the second day (Familiarization session), two identical objects (50 mL conical tubes filled with white-painted water) were fixed in diagonally opposite corners, 10 cm away from the walls. Each mouse was placed at the edge of the arena, away from the objects, and allowed to freely explore the arena for 10 min. On the third day (Test session), one of the objects was replaced with a novel object (a ceramic piece of similar size). Again, each mouse was placed at the edge of the arena, away from the objects, and allowed to freely explore the arena for 10 min. Before each trial, the arena and objects were sanitized with alcohol and allowed to dry completely for at least 3 min before proceeding with the experiment. The entire movement of the mice was recorded using EthoVision XT 11.5. The time spent interacting with each object during each trial was scored by an experimenter using recorded video. The experimenter was blinded to the group identity of all mice during the analysis.
Morris water maze
This experiment is one of the behavioral tests designed by Morris and is widely used to study spatial learning and memory in rodents. 38 The experiment was conducted in a circular swimming pool (100 cm in diameter, 40 cm in depth), and a ceiling-mounted camera connected to a computer was used for recording and analysis of the trials. The pool was filled with water of 22–25°C, which was mixed with non-toxic white paint. On the first trial of the initial day, the platform was located 1 cm above the water, and mice were located in a pool to find the visible platform above the water surface for 60 s. After finding the platform or being guided to it, the mice were placed on the platform for 15 s before being transferred to a warmed cage. For all subsequent trials, the platform was hidden 1 cm below the surface. The mice were trained to freely swim for 60 s to find the hidden platform. If a mouse failed to find the platform within the allotted time, it was guided to the platform and allowed to remain there for 15 s before being transferred to a warmed cage. Four trials per day were performed for five consecutive days, with the starting points randomized for each trial. The probe test was conducted on the sixth day. Mice were released from the start position farthest from the previous platform location, with the platform removed, and were allowed to swim freely for 60 s. The entire movement of the mice was recorded, and the time spent in the target quadrant as well as the total distance to the platform were measured using EthoVision XT 11.5.
Perfusion and tissue preparation
Before perfusion, inhalation anesthesia was performed by placing cotton soaked with 2 mL of Isoflurane (Hana Pharm. Co., 657801261) in a sealed container. The mouse was placed inside, and after 45 s, the anesthetized mouse was removed and secured. Perfusion was performed sequentially using 1× PBS to remove blood, immediately followed by 4% paraformaldehyde to achieve in situ fixation. The extracted brain was fixed overnight in 4% paraformaldehyde (pH 7.4, Sigma, 158127). After overnight fixation, the brain was transferred to 30% sucrose (Sigma, S0389 in 1X PBS) for 2 days for dehydration. The dehydrated brain tissue was preserved in optimal cutting temperature compound (O.C.T. Compound, Sakura Finetek, 4583) and stored at −80°C until further use. For immunofluorescence studies, 50 µm-thick brain slices were sectioned using a cryostat Microtome (Kalstein, YR426-1). Sectioned brain slices were stored in 1X PBS at 4°C until use. If storage exceeded one week, the slices were kept in 1X PBS containing 0.05% sodium azide.
Immunohistochemical analysis
Immunofluorescence studies were performed on 50 µm-thick slices of perfused mouse brains. PBS-T solution was prepared by adding 0.3% Triton X-100 (Sigma, T8787) to 1X PBS, and the blocking solution consisted of PBS-T with 4% normal donkey serum (Sigma, S30-M). After cryosectioning, the frozen brain slices were rehydrated by placing them in 1X PBS in a 12-well plate for at least 30 min prior to the experiment. Brain slices were then washed four times for 5 min each with 1X PBS on a shaker. Blocking was performed with a blocking solution at 4°C for 1 h. The slices were then incubated overnight at 4°C with primary antibodies diluted in the blocking solution. The following day, the slices were washed again four times for 5 min each with 1X PBS and incubated with secondary antibodies diluted in the blocking solution at room temperature for 2 h. After incubation, the slices were washed four times for 5 min each with 1X PBS, followed by DAPI staining (1:1000 in 1X PBS) at room temperature for 10 min. Subsequently, the slices were washed four additional times with 1X PBS. The stained slices were mounted onto glass slides using Mounting Medium (Vector Laboratories, Inc., H-5700) and covered with 24 × 40 mm coverslips (Marienfeld, 0101192). Each glass slides contained brain slices from one mouse, with approximately 6–12 slices per mouse. Detailed information about the primary and secondary antibodies used is provided in Supplemental Table 1. The slices were visualized and imaged using an inverted fluorescence microscope (Leica DMi8). FITC and RHOD_LP filters were used to detect fluorescence signals corresponding to green and red markers, respectively. The signal intensity of each staining was measured using Leica Application Suite X software (LAS X, the software associated with Leica microscopes). Signal area, puncta count, and average puncta size were analyzed using ImageJ software (further details available at https://fiji.sc).
RNA sequencing
Hippocampal tissues were collected from 5xFAD and wild-type mice following PBS perfusion, homogenized in TRIzol, and stored at −80 °C. Total RNA was extracted and assessed for integrity (RIN ≥ 8.0). mRNA libraries were prepared using the TruSeq Stranded mRNA Kit and sequenced on an Illumina NovaSeq 6000 platform (paired-end 151 bp). Data processing was performed by Macrogen Inc. (Seoul, South Korea) using FastQC, Trimmomatic, HISAT2, and StringTie with the mm10 (GRCm39) reference genome. Gene expression was quantified in TPM for downstream analysis.
Statistical analysis
All statistical analyses were performed using GraphPad PRISM V6 software. Data were expressed as mean ± S.E.M. Statistical significance was set at p < 0.05. Significance levels were indicated as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. For experiments involving four experimental groups (WT, 5xFAD vehicle, 5xFAD + donepezil, and 5xFAD + 2BAct), statistical comparisons were performed using ordinary one-way ANOVA followed by Dunnett's multiple-comparisons test with the vehicle-treated 5xFAD group as the reference condition. For analyses involving two independent factors, ordinary two-way ANOVA was applied. In the novel object recognition test (NORT), comparisons between object types within groups were analyzed using two-way ANOVA followed by Sidak's multiple-comparisons test. For other two-way ANOVA analyses, Dunnett's multiple-comparisons test was applied where appropriate.
Results
2BAct does not affect locomotor activity and anxiety-related behavior in 5xFAD mice
Ten-month-old WT and 5xFAD mice received daily intraperitoneal injections of either 2BAct (10 mg/kg/day), donepezil (2 mg/kg/day; positive control), or vehicle for 23 consecutive days. Behavioral experiments began with low-stress protocols, starting with the OFT for habituation in preparation for the NORT (Figure 1A). Donepezil, an acetylcholinesterase inhibitor and a clinically approved and widely used medication for AD,3,6,39 served as a positive control. Donepezil has been shown to alleviate learning and memory deficits as well as inflammation in AD patients.3,40,41
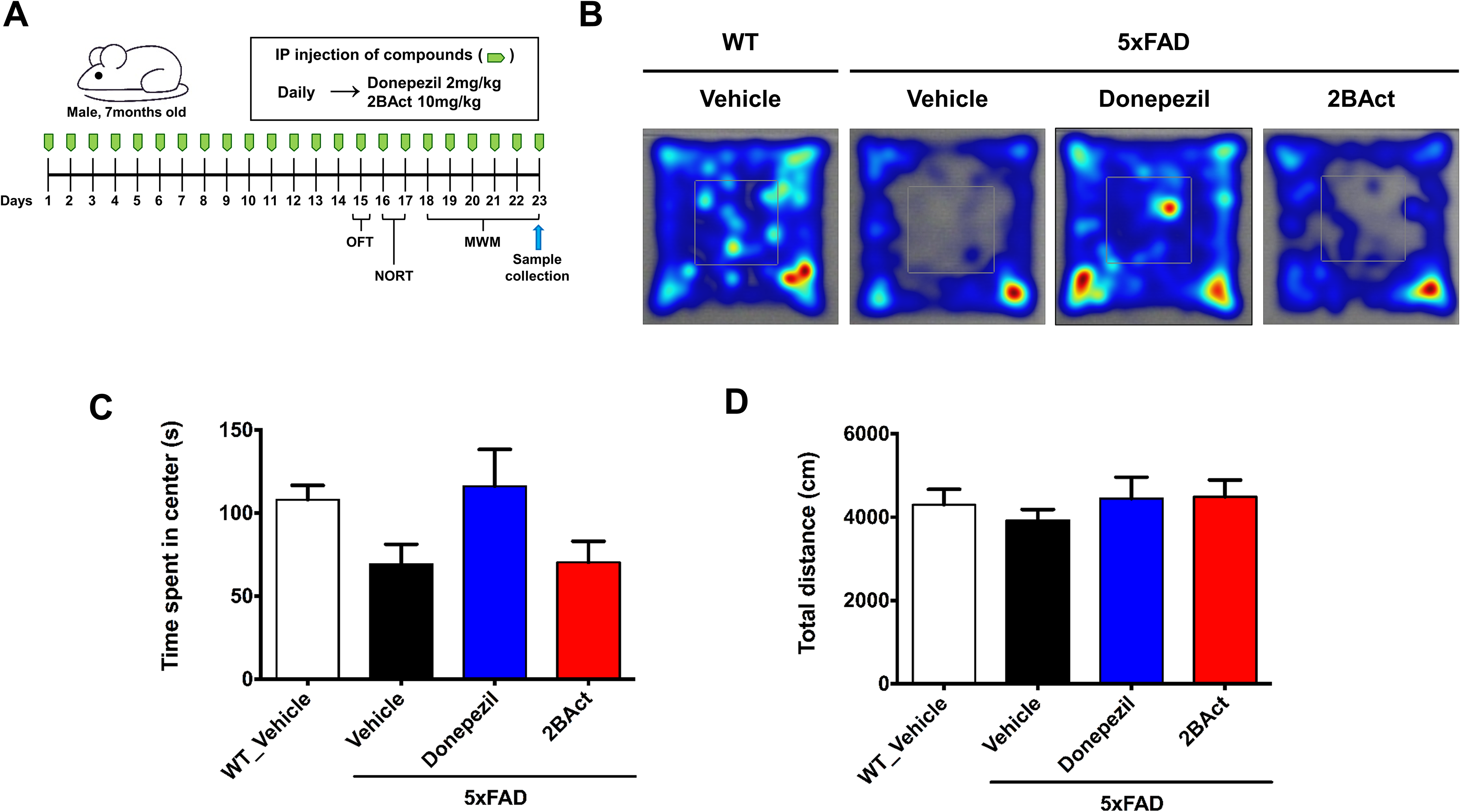
2BAct does not affect anxiety-related behavior in 5xFAD mice. (A) Schematic representation of treatment and behavioral experiments. Mice were administered vehicle, donepezil (2 mg/kg), or 2BAct (10 mg/kg) daily. After completion of behavioral tests, brains were collected following perfusion for tissue staining. (B) Heat map illustrating the 10-min exploration patterns of mice in the OFT. (C) Time spent in the center zone of the field (analyzed using ordinary one-way ANOVA followed by Dunnett's post hoc test; F (3, 28) = 2.802, p = 0.0582). (D) Total distance traveled within the field during the entire duration of the test (analyzed using ordinary one-way ANOVA followed by Dunnett's post hoc test; F (3, 28) = 0.4184, p = 0.7412). Comparisons were made vs vehicle-treated 5xFAD mice. All results are presented as mean ± SEM, n = 8 mice per group.
We first evaluated whether 2BAct impacts general locomotion and anxiety-like behaviors using the OFT. Analysis of exploratory behavior showed that both WT and Donepezil-treated mice spent more time in the center zone compared to vehicle-treated 5xFAD mice; however, these differences did not reach statistical significance (Figure 1B, C). In contrast, 2BAct treatment did not significantly change the time spent in the center zone compared to vehicle-treated 5xFAD mice. These results indicate no statistically significant effects of the treatments on anxiety-like behavior under the conditions tested. Additionally, total locomotor activity, measured by the total distance traveled, revealed no significant differences among WT, vehicle-treated 5xFAD, 2BAct-treated 5xFAD, and donepezil-treated groups, confirming that neither 2BAct nor donepezil altered overall motor function or activity levels (Figure 1D). Throughout the experimental period, body weight changes among groups were also comparable (Supplemental Figure 2).
2BAct rescues object recognition memory deficits in 5xFAD mice
Recognition memory deficits, characteristic of AD, were assessed using the NORT. During the familiarization phase, no significant differences in exploration time for identical objects were observed across all groups, confirming similar initial exploratory behaviors (Supplemental Figure 3). After 24 h, during the test phase in which one familiar object was replaced by a novel one, all groups of mice showed a preference for the novel object (Figure 2A). At this time point, vehicle-treated 5xFAD mice exhibited significantly reduced recognition of the novel object, reflecting impaired memory function. Notably, treatment with 2BAct significantly restored novel object preference in 5xFAD mice to WT levels, mirroring the effects of donepezil. This improvement was reflected in the discrimination index (DI), which was significantly increased in 2BAct- and donepezil-treated mice compared to vehicle-treated 5xFAD controls (Figure 2B). These findings suggest that 2BAct improves performance in the novel object recognition test, indicating a task-specific enhancement of recognition memory in 5xFAD mice.
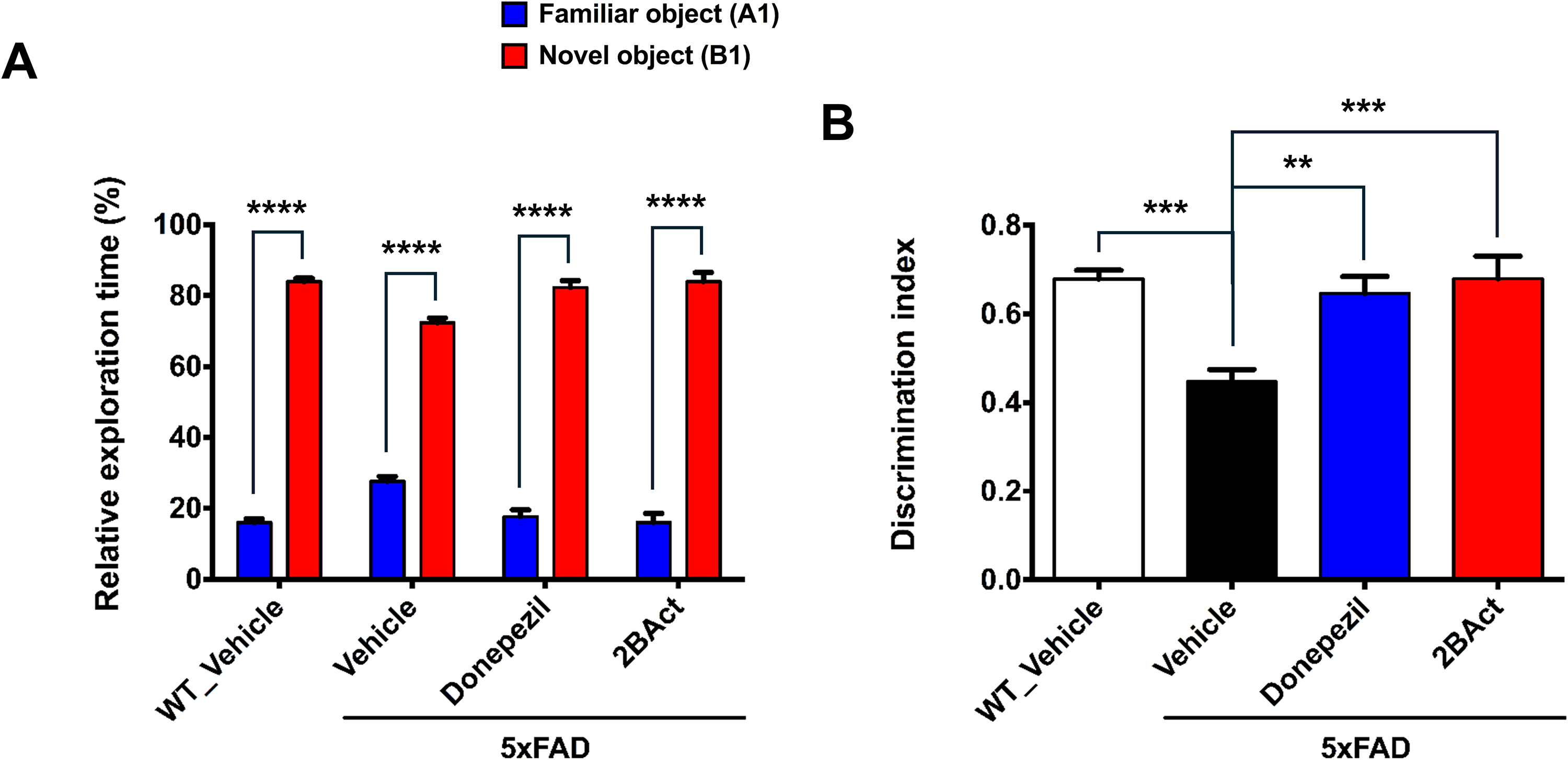
2BAct restores object recognition memory deficits in 5xFAD mice. The mice were exposed to two identical objects, and 24 h later, one object was replaced with a novel object. (A) Relative preference ratio for the familiar and novel objects after 24 h (analyzed using ordinary two-way ANOVA followed by Sidak's post hoc tests; interaction: F (3, 56) = 19.32, p < 0.0001; group factor: F (3, 56) = 1.15 × 10−12, p > 0.9999; object factor: F (1, 56) = 2329, p < 0.0001). ****p < 0.0001 for familiar vs novel object within each group. (B) The discrimination index (DI) for the NORT is shown (analyzed using ordinary one-way ANOVA followed by Dunnett's post hoc test; F (3, 28) = 9.658, p = 0.0002). DI = (novel − familiar) / (novel + familiar). **p < 0.01, ***p < 0.001 versus vehicle-treated 5xFAD mice. All results are presented as mean ± SEM, n = 8 mice per group.
2BAct does not significantly improve spatial memory deficits in 5xFAD mice
Spatial learning and memory, evaluated via the Morris water maze (MWM), are commonly compromised in AD. During the training phase, all groups demonstrated progressively reduced latency times to locate the hidden platform, indicating similar learning capacities (Figure 3A). However, in the 60-s probe trial assessing memory retention, vehicle-treated 5xFAD mice spent less time in the target quadrant compared to WT mice, although this difference did not reach statistical significance. Treatment with donepezil significantly improved spatial memory, whereas 2BAct did not significantly affect performance in the probe trial (Figure 3B, C). Importantly, no differences in swimming distance were observed among groups during the probe trial, indicating comparable swimming performance (Figure 3D).
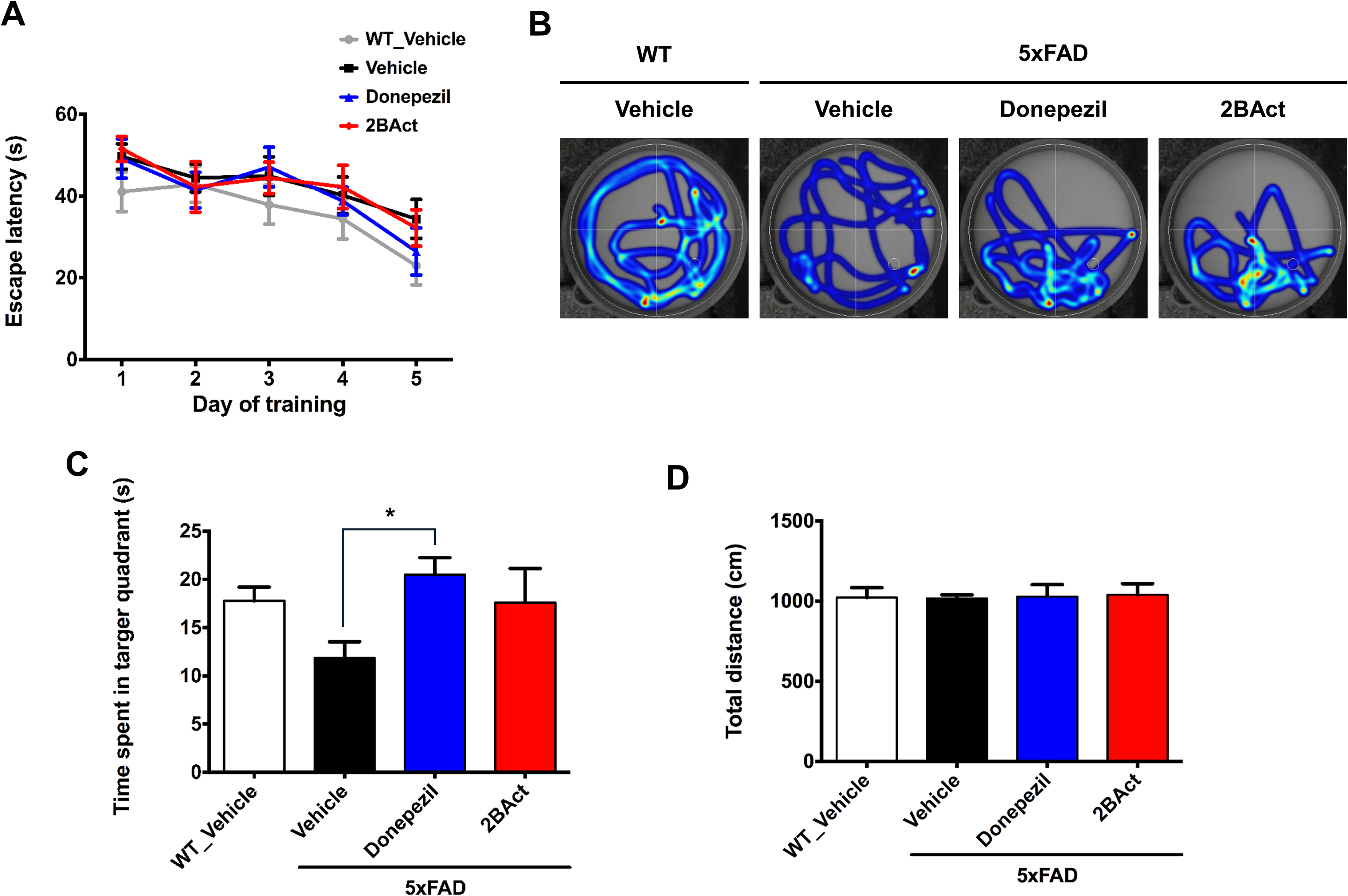
2BAct does not affect spatial learning and memory deficits in 5xFAD mice. The training trials were conducted four times daily for five consecutive days. A probe test was performed 24 h after the final training session, with the platform removed. (A) Time taken to reach the hidden platform during the training trials (analyzed using ordinary two-way ANOVA followed by Dunnett's post hoc tests; interaction: F (12, 140) = 0.3101, p = 0.9867; group factor: F (4, 140) = 9.710, p < 0.0001; time factor: F (3, 140) = 2.443, p = 0.0667). Comparisons were made vs vehicle-treated 5xFAD mice at each training day. (B) Tracking lines of mice during the probe test on the sixth day. (C) Time spent in the target quadrant during the 60-s probe trial (analyzed using ordinary one-way ANOVA followed by Dunnett's post hoc test; F (3, 28) = 2.572, p = 0.0741). *p < 0.05 versus vehicle-treated 5xFAD mice. (D) Total distance traveled within the water maze during the 60-s probe trial (analyzed using ordinary one-way ANOVA followed by Dunnett's post hoc test; F (3, 28) = 0.02626, p = 0.9941). Comparisons were made versus vehicle-treated 5xFAD mice. All results are presented as mean ± SEM, n = 8 mice per group.
2BAct attenuates microglial activation and tau pathology but does not affect astrocytic activation or Aβ pathology in 5xFAD mice
We further evaluated pathological and inflammatory responses via immunofluorescence staining for astrocytes (GFAP), microglia (Iba1), Aβ deposition (6E10), and tau pathology (Tau-5). Consistent with AD pathology, vehicle-treated 5xFAD mice exhibited increased hippocampal GFAP immunoreactivity compared to WT mice; however, neither 2BAct nor donepezil significantly altered GFAP levels compared to vehicle-treated 5xFAD mice (Figure 4A, B). Likewise, Aβ deposition assessed by 6E10 immunostaining remained unchanged after both treatments (Figure 4A, C). In contrast, 2BAct treatment significantly reduced microglial activation, as evidenced by decreased Iba1 immunoreactivity in the hippocampus compared to vehicle-treated 5xFAD mice (Figure 5A, B). Tau pathology was also significantly reduced following 2BAct treatment, as shown by Tau-5 staining (Figure 5A, C). These results indicate that 2BAct reduces microglial activation and tau pathology, without affecting astrocytic activation or amyloid deposition.
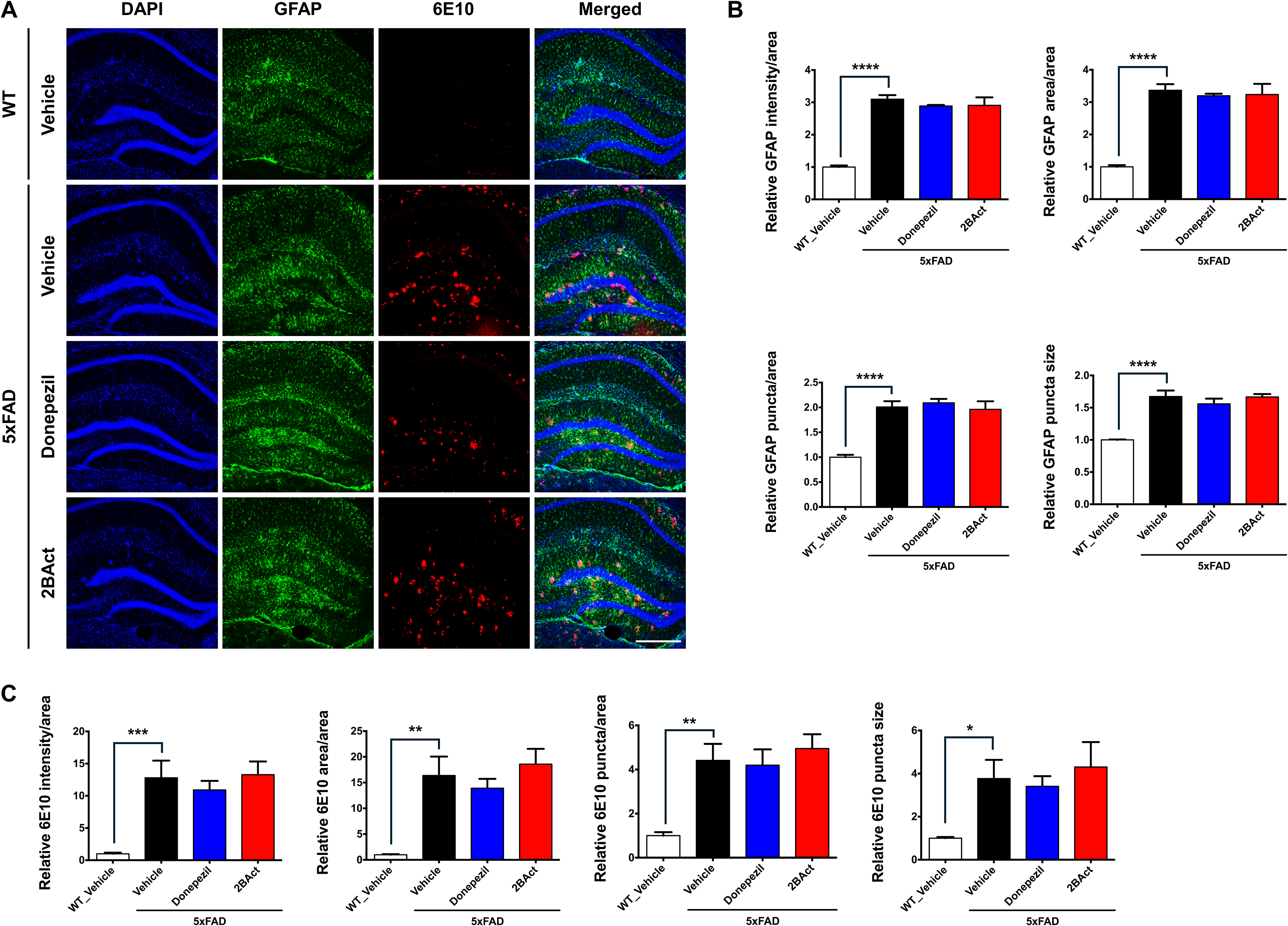
2BAct does not affect astrocytic activation or Aβ deposition in 5xFAD mice. (A) Immunohistochemical results showing that 2BAct does not alter GFAP (green) and 6E10 (red) immunoreactivity in the hippocampus of 5xFAD mice. Nuclei were counterstained with DAPI (blue). GFAP and 6E10 immunoreactivity were significantly elevated in 5xFAD mice compared to WT mice. Scale bar: 200 µm. (B) Quantification of GFAP immunoreactivity, including signal intensity per total area, signal area per total area, puncta count per total area, and average puncta size (all analyzed using ordinary one-way ANOVA followed by Dunnett's post hoc tests; intensity: F (3, 15) = 64.54, p < 0.0001; area: F (3, 15) = 44.74, p < 0.0001; puncta: F (3, 15) = 26.23, p < 0.0001; puncta size: F (3, 15) = 22.76, p < 0.0001). ****p < 0.0001 versus vehicle-treated 5xFAD mice. (C) Quantification of 6E10 immunoreactivity, including signal intensity per total area, signal area per total area, puncta count per total area, and average puncta size (all analyzed using ordinary one-way ANOVA followed by Dunnett's post hoc tests; intensity: F (3, 15) = 10.39, p = 0.0006; area: F (3, 15) = 10.06, p = 0.0007; puncta: F (3, 15) = 8.499, p = 0.0015; puncta size: F (3, 15) = 4.291, p = 0.0225). *p < 0.05, **p < 0.01, ***p < 0.001 versus vehicle-treated 5xFAD mice. All results are presented as mean ± SEM, n = 4–5 mice per group.
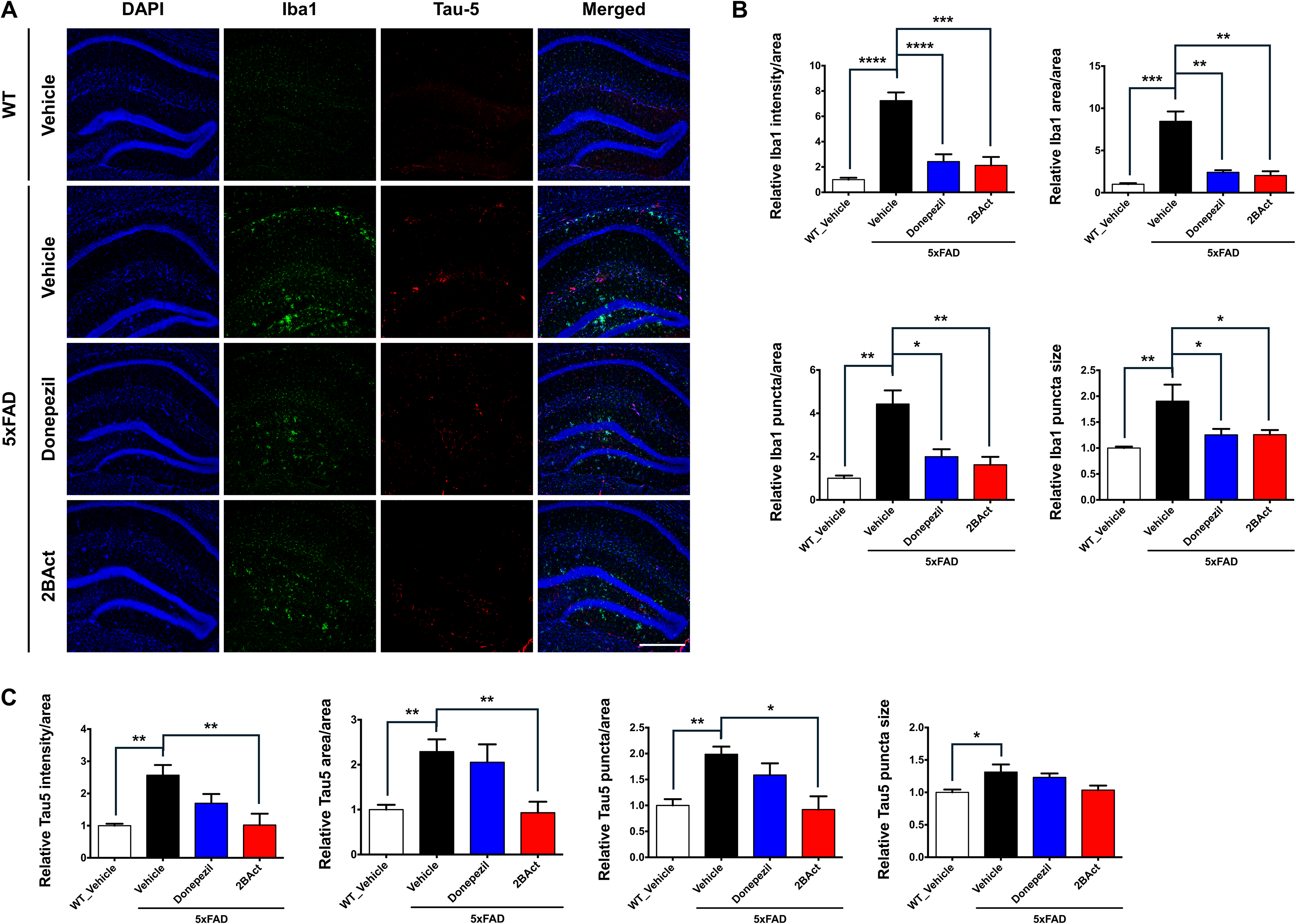
2BAct ameliorates microglial activation and Tau accumulation in 5xFAD mice. (A) Immunohistochemical results showing that 2BAct reduces Iba1 (green) and Tau-5 (red) immunoreactivity in the hippocampus of 5xFAD mice. Nuclei were counterstained with DAPI (blue). Iba1 and Tau-5 immunoreactivity were significantly elevated in 5xFAD mice compared to WT mice. Scale bar: 200 µm. (B) Quantification of Iba1 immunoreactivity, including signal intensity per total area, signal area per total area, puncta count per total area, and average puncta size (all analyzed using ordinary one-way ANOVA followed by Dunnett's post hoc tests; intensity: F (3, 13) = 16.86, p < 0.0001; area: F (3, 13) = 12.49, p = 0.0004; puncta: F (3, 13) = 7.681, p = 0.0033; puncta size: F (3, 13) = 5.547, p = 0.0113). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs vehicle-treated 5xFAD mice. (C) Quantification of Tau-5 immunoreactivity, including signal intensity per total area, signal area per total area, puncta count per total area, and average puncta size (all analyzed using ordinary one-way ANOVA followed by Dunnett's post hoc tests; intensity: F (3, 13) = 7.911, p = 0.0030; area: F (3, 13) = 7.186, p = 0.0043; puncta: F (3, 13) = 7.118, p = 0.0045; puncta size: F (3, 13) = 4.205, p = 0.0277). *p < 0.05, **p < 0.01 versus vehicle-treated 5xFAD mice. All results are presented as mean ± SEM, n = 4–5 mice per group.
2BAct attenuates hippocampal ISR activation at the phospho-eIF2α level in 5xFAD mice
Phosphorylation of eIF2α, a critical ISR marker associated with impaired synaptic function and cognitive decline,20,22 was significantly elevated in vehicle-treated 5xFAD mice compared to WT controls. Contrary to our initial hypothesis, both donepezil and 2BAct significantly reduced hippocampal p-eIF2α immunoreactivity (Figure 6A, B). Given that neither drug is known to directly target the phosphorylation of eIF2α, the observed reduction likely results indirectly from alleviation of pathological states in AD. It is plausible that improvements in inflammatory signaling, tau pathology, or other downstream pathways linked to ISR activation contribute to this indirect modulation of eIF2α phosphorylation. Overall, these findings are consistent with indirect attenuation of eIF2α phosphorylation accompanying the amelioration of AD-related pathology by 2BAct, rather than demonstrating a direct causal effect.
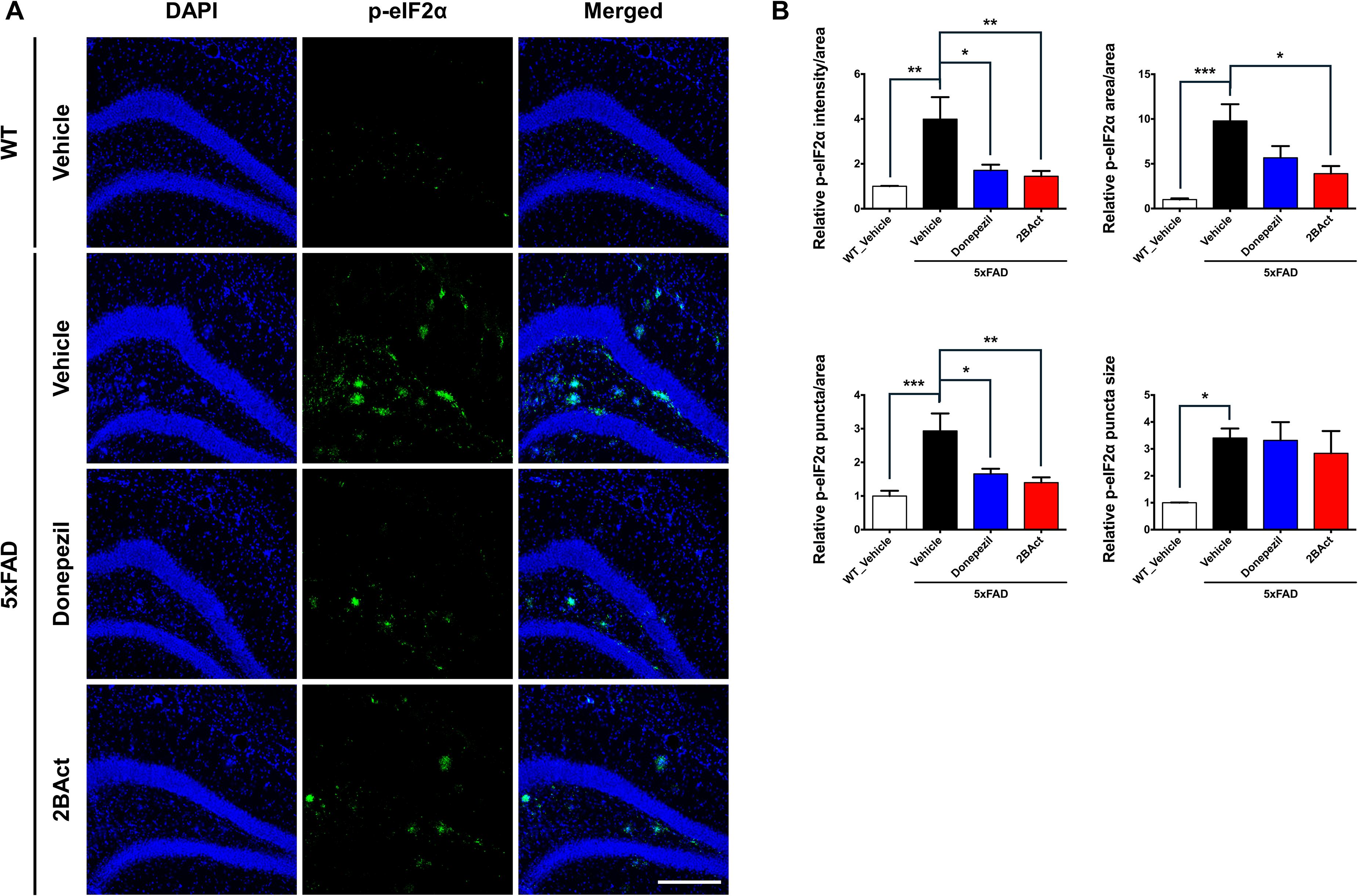
2BAct decreases phosphorylation of eIF2α in 5xFAD mice. (A) Immunohistochemical results showing that 2BAct reduces p-eIF2α (green) immunoreactivity in the hippocampus of 5xFAD mice. Nuclei were counterstained with DAPI (blue). p-eIF2α immunoreactivity was significantly elevated in 5xFAD mice compared to WT mice. Scale bar: 100 µm. (B) Quantification of p-eIF2α immunoreactivity, including signal intensity per total area, signal area per total area, and puncta count per total area, and average puncta size (all analyzed using ordinary one-way ANOVA followed by Dunnett's post hoc tests; intensity: F (3, 13) = 6.923, p = 0.0050; area: F (3, 13) = 8.333, p = 0.0024; puncta: F (3, 13) = 8.535, p = 0.0022; puncta size: F (3, 13) = 3.555, p = 0.0447). *p < 0.05, **p < 0.01, ***p < 0.001 versus vehicle-treated 5xFAD mice. All results are presented as mean ± SEM, n = 4–5 mice per group.
To further examine ISR-related changes at the transcriptional level, we analyzed hippocampal expression of ISR-related genes using RNA sequencing. Notably, ATF4, a key transcriptional effector downstream of eIF2α phosphorylation and a robust molecular indicator of ISR activation,20,22 showed no statistically significant difference between groups. Similarly, expression levels of additional ISR-related genes, including DDIT3, PPP1R15A, and ATF5, were not significantly altered by 2BAct or donepezil treatment (Supplemental Figure 4). Because the dataset consisted of normalized TPM values from a small cohort, formal FDR-based differential expression testing was not applicable. Therefore, these RNA-seq data are presented as descriptive observations. Under these conditions, no statistically significant transcriptional changes in ISR-related genes were detected. These findings indicate that no robust transcriptional alterations were detected under the current experimental conditions and should be interpreted in the context of the protein-level changes observed.
Discussion
The present study provides experimental evidence supporting the therapeutic potential of the eIF2B activator 2BAct in improving recognition memory performance in the 5xFAD mouse model of Alzheimer's disease. Our findings suggest that 2BAct significantly improves object recognition memory, while effectively reducing hippocampal eIF2α phosphorylation, a core marker of ISR activation. Although the drug did not alter Aβ plaque burden or astrocytic activation, it showed significant reductions in microglial activation and tau accumulation, suggesting a selective modulation of specific pathological mechanisms underlying AD.
An important distinction between 2BAct and donepezil observed in the present study lies in their qualitative profiles rather than the magnitude of behavioral efficacy. While donepezil exerted robust effects on cognitive performance, consistent with its established role as a symptomatic treatment, 2BAct produced limited behavioral improvements. Notably, 2BAct significantly reduced tau accumulation, whereas both 2BAct and donepezil comparably attenuated microglial activation, indicating partially overlapping but mechanistically distinct pathological effects. In the OFT, 2BAct did not alter locomotor activity or significantly affect anxiety-like behavior, indicating that the observed behavioral effects are not attributable to changes in general locomotor activity or sedation. Together, these findings indicate that 2BAct is associated with changes in ISR signaling and tau pathology, rather than primarily enhancing neurotransmission as observed with donepezil.
Previous studies have highlighted the role of chronic ISR activation in exacerbating cognitive dysfunction and neurodegeneration in AD.8,10,20,24,42 In particular, Lourenço et al. 43 demonstrated that Aβ oligomers trigger a TNF-α–PKR–eIF2α signaling cascade, leading to translational repression, synaptic dysfunction, and memory impairment in both mouse and primate models of AD. This work provides mechanistic evidence that inflammatory signaling can drive ISR-dependent cognitive decline. Our observation that 2BAct reduces p-eIF2α levels is consistent with attenuation of maladaptive ISR signaling, which may in turn help restore protein synthesis necessary for synaptic function. However, this relationship remains correlative, and further studies are required to directly establish eIF2B activation as the mechanistic driver. The improvements in tau pathology and microglial activation further support the role of ISR in modulating neuroinflammatory responses and tau dynamics, possibly through mechanisms involving microglial signaling pathways, although this requires further molecular validation.
Recent findings by Flury et al. 44 further support this interpretation, demonstrating that chronic ISR activation in microglia induces a neurotoxic “dark microglia” phenotype via aberrant lipid metabolism and secretion, leading to synaptic loss and cognitive deficits in AD. In our study, 2BAct reduced p-eIF2α levels and significantly ameliorated microglial activation and tau accumulation. These findings are consistent with the possibility that pharmacological activation of eIF2B attenuates maladaptive microglial phenotypes, potentially through indirect suppression of ISR signaling. Therefore, ISR inhibition could exert therapeutic effects not only through restoring neuronal homeostasis but also by modulating microglial-driven neuroinflammation.
One notable finding is the lack of impact of 2BAct on amyloid deposition and astrocytic activation. This suggests that targeting ISR may be insufficient as a monotherapy to resolve all facets of AD pathology. Given that AD pathology is multifaceted, combining ISR-targeting drugs like 2BAct with agents addressing amyloid or tau aggregation might yield enhanced therapeutic efficacy. Indeed, combined drug approaches have demonstrated superior outcomes in various neurodegenerative models.45–47
Importantly, the interpretation of our findings should consider the limitations of transgenic models such as 5xFAD, which do not fully capture the complexity of sporadic AD. Although the ISR is generally protective by reducing protein synthesis under stress, chronic and maladaptive ISR activation has been implicated in synaptic dysfunction and cognitive decline.12,19,20–24
In this context, modulation of the ISR by 2BAct should not be viewed as a simple suppression of a protective pathway, but rather as a strategy to restore translational homeostasis under chronic stress conditions. This perspective provides a conceptual counterbalance to approaches that aim to reduce protein stress by suppressing protein synthesis,48,49 suggesting that restoring appropriate levels of protein translation may represent a complementary therapeutic strategy in AD. Nevertheless, further studies in diverse models will be required to validate these findings.
Our study has several limitations. While behavioral outcomes strongly indicate cognitive improvement, further molecular characterization of ISR downstream targets and synaptic proteins would help clarify the mechanisms underlying these effects. We did not directly measure eIF2B activity or expression, which precludes a definitive demonstration of target engagement. The relatively short treatment duration and modest sample size (n = 8 per group, within the typical range for AD behavioral studies)20,40 also warrant cautious interpretation.
Although the behavioral improvement was most evident in the novel object recognition test, no statistically significant effects were observed across multiple behavioral paradigms, including the MWM. Further studies are required to clarify the effects of 2BAct on cognitive performance. Replication of these findings in independent cohorts and through additional behavioral paradigms will be essential to confirm this interpretation.
Behavioral and molecular analyses were conducted in the same cohort, which may influence stress-related pathways. However, testing was performed in a low-stress order (OFT → NORT → MWM) with recovery intervals, and all groups underwent identical handling to minimize procedural bias. Moreover, our immunohistochemical analysis did not identify the specific cell types exhibiting changes in p-eIF2α. Future studies combining phospho-ISR markers with neuronal and glial markers (NeuN, Iba1, GFAP) will be required to delineate cell-type-specific ISR modulation.
Finally, although 2BAct has shown a favorable safety profile in rodent studies, previous high-dose canine data reported cardiac abnormalities. 16 These effects were species-specific and occurred at exposures far exceeding those used here, but they underscore the need for comprehensive long-term toxicological evaluation, particularly regarding cardiovascular safety.
While our results demonstrate correlations between ISR attenuation and behavioral improvements, we acknowledge that direct mechanistic evidence linking eIF2B activation to cognitive recovery remains to be established. Future studies should aim to elucidate the precise molecular mechanisms of 2BAct's action, including assessing the downstream ISR mediators such as ATF4 and related signaling pathways. Moreover, longer-term studies and combination therapies should be explored to comprehensively evaluate the disease-modifying potential of ISR inhibition in AD. Collectively, our findings highlight eIF2B activation as a promising therapeutic avenue for AD, providing preclinical evidence to support the further development and evaluation of ISR modulators.
Supplemental Material
sj-docx-1-alz-10.1177_13872877261450638 - Supplemental material for Pharmacological targeting of the integrated stress response by 2BAct improves object recognition memory and reduces neuroinflammation in the 5xFAD model of Alzheimer's disease
Supplemental material, sj-docx-1-alz-10.1177_13872877261450638 for Pharmacological targeting of the integrated stress response by 2BAct improves object recognition memory and reduces neuroinflammation in the 5xFAD model of Alzheimer's disease by Daegeon Kim, Hojung Jeong, Hye Kyung Lim, Soo Min Ahn and Minseok Song in Journal of Alzheimer's Disease
Footnotes
Acknowledgements
We gratefully acknowledge the Alborada Drug Discovery Institute for the synthesis of 2BAct.
Ethical considerations
All experimental procedures involving animals were conducted in accordance with the guidelines of Yeungnam University Institutional Animal Care and Use Committee (IACUC) and were approved under protocol number 2023-021.
Consent to participate
Not applicable
Consent for publication
Not applicable.
Author contribution(s)
Funding
This research was supported by the 2025 RISE Program at Yeungnam University.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The data supporting the findings of this study will be made available by the corresponding author upon reasonable request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.