
Abstract
Background
A major pathological hallmark of Alzheimer's disease (AD), the most common cause of dementia, is the accumulation of amyloid-β (Aβ) plaques. However, currently approved therapeutic agents fail to fundamentally halt disease progression and have limitations in terms of efficacy, sustainability, and safety. CD300c is an immunoregulatory molecule that modulates monocyte differentiation and activates macrophages. We recently developed a fully human anti-CD300c antibody, CB201, demonstrating its therapeutic potential in early and late 5xFAD mice.
Objective
This study investigated the efficacy of CB201 in late-stage AD using the APP/PS1 transgenic mouse model. We analyzed the effects of administering CB201 on changes in memory and cognitive function and Aβ and tau protein accumulation in the brain.
Methods
To assess long- and short-term memory improvements, behavioral tests were conducted using the Morris water maze and fear conditioning. Immunostaining was performed to quantify changes in Aβ and tau accumulation.
Results
CB201-treated late-stage AD mice demonstrated improved cognitive performance and memory, comparable to wild-type controls. Histopathological analysis further revealed that CB201 treatment reduced Aβ and tau accumulation.
Conclusions
CB201 exerts significant therapeutic effects on functional impairments and pathological alterations in late-stage AD. These results confirm CB201 as a potential immunotherapeutic for the treatment of AD.
Introduction
Approximately 90% of dementia cases are estimated to be attributable to Alzheimer's disease (AD), underscoring its dominant role among dementia subtypes. 1 AD is characterized primarily by two pathological features: amyloid-β (Aβ) plaque formation and the formation of neurofibrillary tangles due to tau protein aggregation following hyperphosphorylation. 2 These abnormal accumulations of Aβ and tau proteins lead to the development of fibrillar lesions, which are typically observed in AD pathology. 3 Five treatments are currently approved for AD, but their primary function is to manage symptoms rather than prevent disease progression. 4 Donanemab and lecanemab are both monoclonal antibodies that target Aβ, and both have been approved for the treatment of early symptomatic AD. 5 Donanemab binds to N-terminally truncated forms of Aβ and clears plaques through microglia-mediated phagocytosis. 6 Patients receiving donanemab have reported side effects including hospitalization for amyloid-related imaging abnormalities. 7 In contrast, lecanemab binds to protofibrillar Aβ and intervenes at early stages to prevent the development of amyloid plaques. 8 However, questions remain about the therapeutic effectiveness and possible adverse effects of this treatment.
The CD300 family comprises seven proteins, designated CD300a through CD300f, that are the products of genes localized to human chromosome 17.9,10 CD300c is expressed across a range of immune cells, including T cells; NK cells; neutrophils; antigen-presenting cells, such as B cells, dendritic cells, monocytes, and macrophages; and certain cancer cells.11,12 CD300c protein consists of 224 amino acids with a transmembrane domain, cytoplasmic domain, and an extracellular domain. CD300c interacts with DNAX-associated adaptor proteins (e.g., DAP12 and DAP10), as well as the Fc receptor gamma chain (FcRγ). 13 Functioning as a co-inhibitory receptor, CD300c, the sequence of which resembles B7 family proteins, attenuates T-cell activation and limits cytokine secretion. 11
Recent studies have demonstrated that treatment with an anti-CD300c antibody results in monocyte differentiation into M1 macrophages and activates the MAPK and NF-κB signaling pathways. 14 In our previous study, administration of anti-CD300c antibody in a mouse model of AD, 5xFAD mice, led to significant improvements in behavioral deficits; in the Y-maze test, a 44% increase in spontaneous alternation was observed, and in the Morris water maze (MWM), time spent at the platform site increased by 70%. 15
Based on these results, we used a synthetic human phage display library to develop a fully human monoclonal antibody, CB201 (CL7), which has cross-reactivity between mouse and human CD300c antigen. This antibody targets CD300c and was investigated as a therapeutic candidate in AD.
In previous studies, 5xFAD mice were utilized for the model's combination of mutations in presenilin-1 (PSEN1; M146L and L286V) and APP [Swedish (K670N/M671L), Florida (I716V), and London (V717I)], which together result in elevated levels of Aβ42. 14 In this model, the AD pathology progresses quickly, with intracellular Aβ42 accumulation initiating at 1.5 months and extensive amyloid plaque deposition observed by approximately 2 months. 16 In contrast, the present work employed the APP/PS1 mouse model, characterized by a comparatively slower progression of AD pathology. Human APP695 with the Swedish mutation (hAPP695swe) is over-expressed in these mice in the presence of the PSEN1 M146 V mutation (APPswe/PS1.M146V). Accumulation of Aβ in the brain begins at approximately 6 to 7 months of age and is followed by pronounced cognitive decline and memory impairments. 17 Although the 5xFAD model demonstrates rapid pathology and a robust phenotype, the APP/PS1 model's gradual pathological progression more closely reflects the course of AD in humans. 18
The present study employed the APP/PS1 mouse model, which is another representative model distinct from the 5xFAD model used in previous studies, to evaluate and confirm the therapeutic effects of CB201 in induced AD through behavioral tests assessing cognitive improvement, as well as analyses of Aβ and tau protein accumulation.
Methods
Animal experiments
Mice were housed in an individual ventilated cage system (manufactured by Allentown Inc.) in an animal facility registered with the Ministry of Food and Drug Safety (Registration No. 232). The housing conditions were a 12-h light/dark cycle at a relative humidity <60% and constant temperature of 22 ± 2°C.
To investigate the therapeutic effects of CB201, 8-month-old APPswe/PS1.M146V transgenic mice, which overexpress human APP695 with the Swedish mutation (hAPP695swe) and carry the presenilin-1 M146V mutation, 19 were used together with age-matched wild-type (WT) littermates (C57BL/6 background). All mice used were male. In this experiments, APPswe/PS1.M146V (n = 18) and WT littermates (n = 16) were intraperitoneally administered either PBS or CB201 at a dose of 10 mg/kg on days 0 and 3.
In the experiments, 8-month-old wild-type C57BL/6 mice (n = 16) and APP/PS1 mice (n = 18, all males) were intraperitoneally administered PBS or 10 mg/kg CB201 on days 0 and 3. Isotype controls were not used in this study since there was no effect on all of the biological activities in the previous several settings of the experiment under the conditions established in our laboratory. The Institutional Animal Care and Use Committee (IACUC) of Kyungpook National University approved all animal procedures (Approval No. KNU2023-0532).
Morris water maze
We used the MWM to assess spatial learning and memory. 20 CB201 (10 mg/kg) was administered twice at 8 months of age, and behavioral testing was conducted over 11 days, including a habituation phase (day 0), training phase (days 1–10), and a probe test (day 11).
The habituation phase started with the mice being habituated to handling (5 min) before being trained to swim and climb onto a platform in a triangular sectioned tank without visual cues. Each mouse performed 10 repetitions of swimming and platform climbing, with varied approach angles to promote adaptation.
Training trials were conducted daily in the same environment with consistent spatial cues placed on all four walls of the tank to support orientation learning. The platform was kept in the same location throughout the test period to enable spatial memory acquisition. Escape latency was automatically recorded for each trial as the time to reach the hidden platform (cut-off: 60 s).
The platform was removed on day 11 for the probe trial. Time spent in each quadrant, platform crossing frequency, path length, and swimming speed were recorded. All procedures were performed under randomized, open-blinded conditions.
Fear conditioning
Fear conditioning was conducted over three consecutive days to assess contextual and cued memory. During the training phase on day 1, mice were exposed to an auditory cue paired with a mild foot shock to induce associative learning. Mice were placed in a 310 × 220 mm chamber for 3 min. Two tone-shock pairings were delivered: a tone from 50 to 60 s followed by a 2 mA foot shock from 60 to 62 s, and a second tone from 110 to 120 s followed by a 2 mA shock from 120 to 122 s. Freezing behavior was automatically measured using TimeFX1 software.
On day 2, we tested contextual memory by returning the mice to the original chamber without any stimuli for 5 min. The mice were re-exposed to the same environment without any stimuli to evaluate memory retention related to the context.
On day 3, cued memory was assessed in a novel environment with a tone delivered from 60 to 120 s without a foot shock, and freezing was recorded for 3 min. The mice were re-exposed to the same environment without any stimuli to evaluate memory retention related to the tone.
Freezing percentage was used to quantify memory performance, and behavioral outcomes were evaluated based on direct quantitative and statistical comparisons between experimental groups rather than on a predefined percentage cutoff.
Immunohistochemistry
After the above tests, the mice were transcardially perfused with 4% paraformaldehyde following blood collection. The brains were then extracted and 30-μm-thick floating sections obtained using a vibratome.
To visualize dense-core amyloid plaques, thioflavin S (ThioS) staining was carried out. 16 Brain sections were transferred to a 12-well plate and incubated with 1 mL phosphate-buffered saline (PBS) at room temperature (RT) while shaking at 33 rpm for 5 min. We removed the PBS and incubated the sections for 10 min with 0.5% ThioS (prepared in ethanol). The sections were washed twice with 50% ethanol (RT, 33 rpm, 2 min) and once with PBS (RT, 33 rpm, 5 min). After the final wash, sections were mounted using DAPI-containing mounting medium.
AT8 staining was carried out for Aβ and phosphorylated tau, respectively.3,16 Sections were washed with PBS (RT, 33 rpm, 5 min) before permeabilizing for 1 h with 0.2% Triton X-100 in PBS (RT, 33 rpm). After permeabilization, blocking was performed for 1 h using blocking solution (1 mL, RT, 33 rpm). The sections were incubated with the following primary antibodies diluted 1:1 in a mixture of blocking buffer and 0.05% Triton X-100 in PBS (500 μL total volume per well) at 4°C overnight: AT8 (mouse anti-phospho-Tau, 1:100, MN1020, RRID:AB_223647, Thermo Fisher Scientific). The next morning, sections were washed with PBS plus Tween-20 (PBS-T; RT, 33 rpm, 3× 15 min each). Anti-mouse Alexa Fluor 488 diluted 1:1000 in blocking solution was applied as the secondary antibody for 2 h (RT, 33 rpm), followed by another three washes with PBS-T (RT, 33 rpm, 15 min each). The sections were then mounted using DAPI-containing mounting medium.
Three brain sections per animal were selected. For each section, six cortex regions and four hippocampus regions were imaged by confocal microscopy at 20× magnification. Quantification of the percentage area stained was performed using MetaMorph software.
Statistical analysis
In general, statistical methods were not used to recalculate or predetermine sample sizes. Variance was similar within comparable experimental groups. Individuals performing the experiments were blinded to the identity of experimental groups until the end of data collection and analysis for at least one of the independent experiments. Comparisons between two groups were performed with a two-tailed student's t test. In cases where more than two groups were compared to each other, one-way analysis of variance (ANOVA) was used, followed by Tukey's HSD test. Classical statistical analyses were performed using GraphPad Prism 8.0 software. p < 0.05 was considered as statistically significant.
Results
Enhanced cognitive performance following CB201 administration in AD
To evaluate spatial learning and memory, we carried out a 10-day MWM test, followed by a probe test on day 11. During the training period, the wild-type (WT)+PBS and WT + CB201 groups showed a progressive reduction in escape latency, reaching 16.2 s and 18.4 s, respectively, on day 10. However, the APP/PS1 + PBS group exhibited impaired learning, maintaining a high escape latency of 37.9 s on day 10.
APP/PS1 + CB201 mice had a reduced escape latency of 20.8 s on day 10. The area under the curve for latency revealed a significant 26.5% reduction compared to the APP/PS1 + PBS group according to one-way ANOVA with Tukey's post hoc test (p < 0.01; Figure 1B).
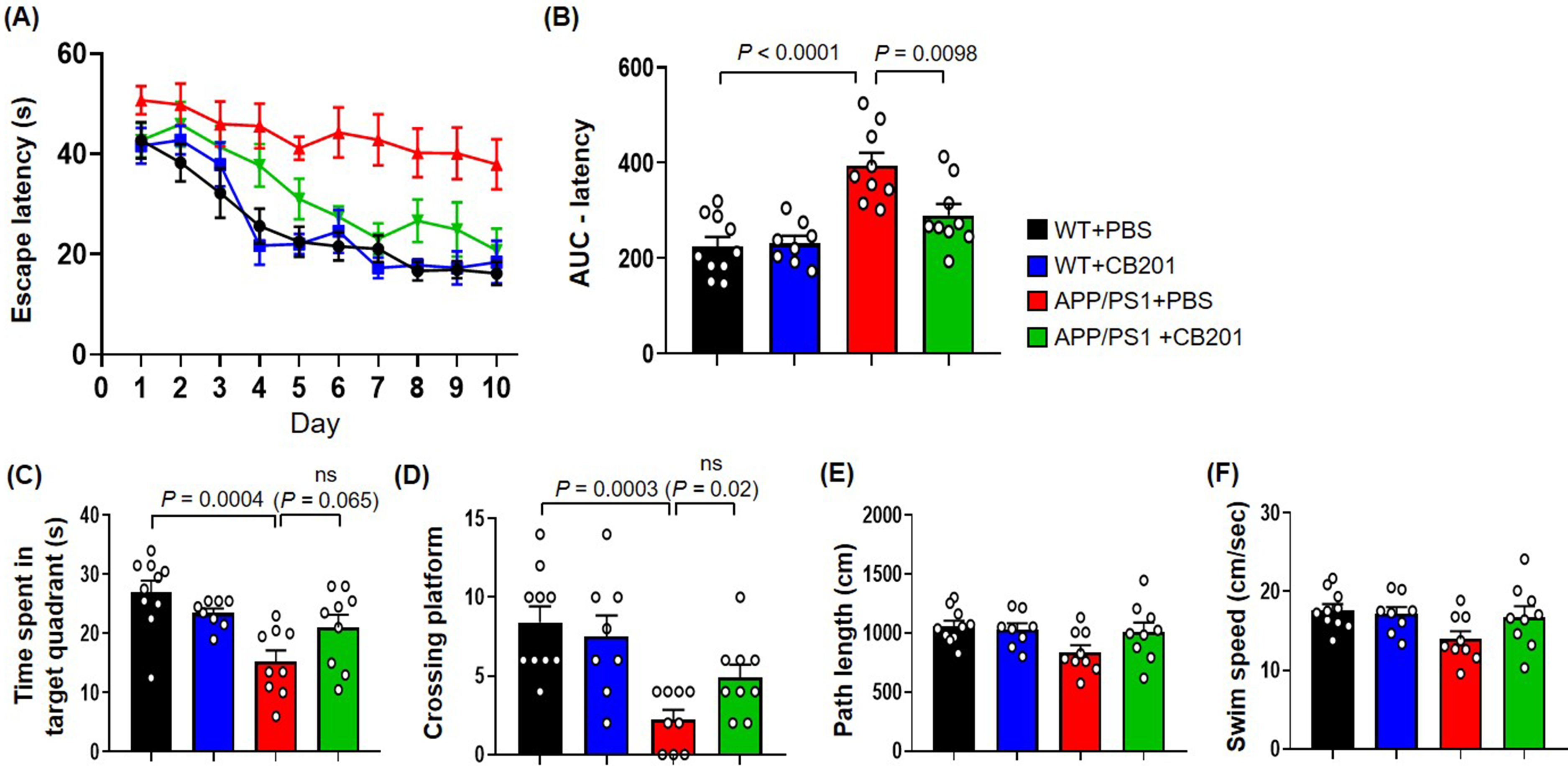
Effects of CB201 treatment on spatial learning and memory in APP/PS1 mice in the Morris water maze (MWM) test. (A) Escape latency in the MWM over 10 days of training. APP/PS1 mice treated with PBS (red) exhibited prolonged escape latency compared to other groups. WT + PBS (n = 10 mice), WT + CB201 (n = 8 mice), APP/PS1 + PBS (n = 9 mice), and APP/PS1 + CB201 (n = 9 mice). (B) Area under the curve (AUC) analysis of escape latency, showing significant improvement in the APP/PS1 + CB201 group (green) compared to the APP/PS1 + PBS group. WT + PBS (n = 10 mice), WT + CB201 (n = 8 mice), APP/PS1 + PBS (n = 9 mice), and APP/PS1 + CB201 (n = 9 mice). (C) Time spent in the target quadrant during the probe trial. WT + PBS (n = 10 mice), WT + CB201 (n = 8 mice), APP/PS1 + PBS (n = 9 mice), and APP/PS1 + CB201 (n = 9 mice). (D) Number of platform crossings. WT + PBS (n = 10 mice), WT + CB201 (n = 8 mice), APP/PS1 + PBS (n = 9 mice), and APP/PS1 + CB201 (n = 9 mice). (E–F) Total path length and swim speed were not significantly different among groups, suggesting no motor impairments. WT + PBS (n = 10 mice), WT + CB201 (n = 8 mice), APP/PS1 + PBS (n = 9 mice), and APP/PS1 + CB201 (n = 9 mice). One-way analysis of variance, Tukey's post hoc test. p < 0.05 was considered as statistically significant. ns = not significant. All data are presented as mean ± SEM (Color figure available online).
In the probe test, the APP/PS1 + PBS group crossed the previous platform location fewer times and spent less time in the target quadrant than the WT controls (Figure 1C, D). Although these changes were not significant, the APP/PS1 + CB201 group showed a trend toward improvement in both parameters, suggesting potential recovery of memory function. In addition, APP/PS1 mice treated with CB201 exhibited a tendency toward increased path length and swim speed compared to the PBS-treated APP/PS1 group, in which these parameters were reduced (Figure 1E, F).
These findings suggest that CB201 treatment may be associated with improvements in spatial learning and long-term memory, while also potentially influencing locomotor activity and anxiety-related behavior.
Recovery of impaired hippocampus-dependent memory with CB201 treatment
Fear conditioning was used to assess hippocampus-dependent memory function. Memory-related performance was evaluated by assessing freezing responses (Figure 2).
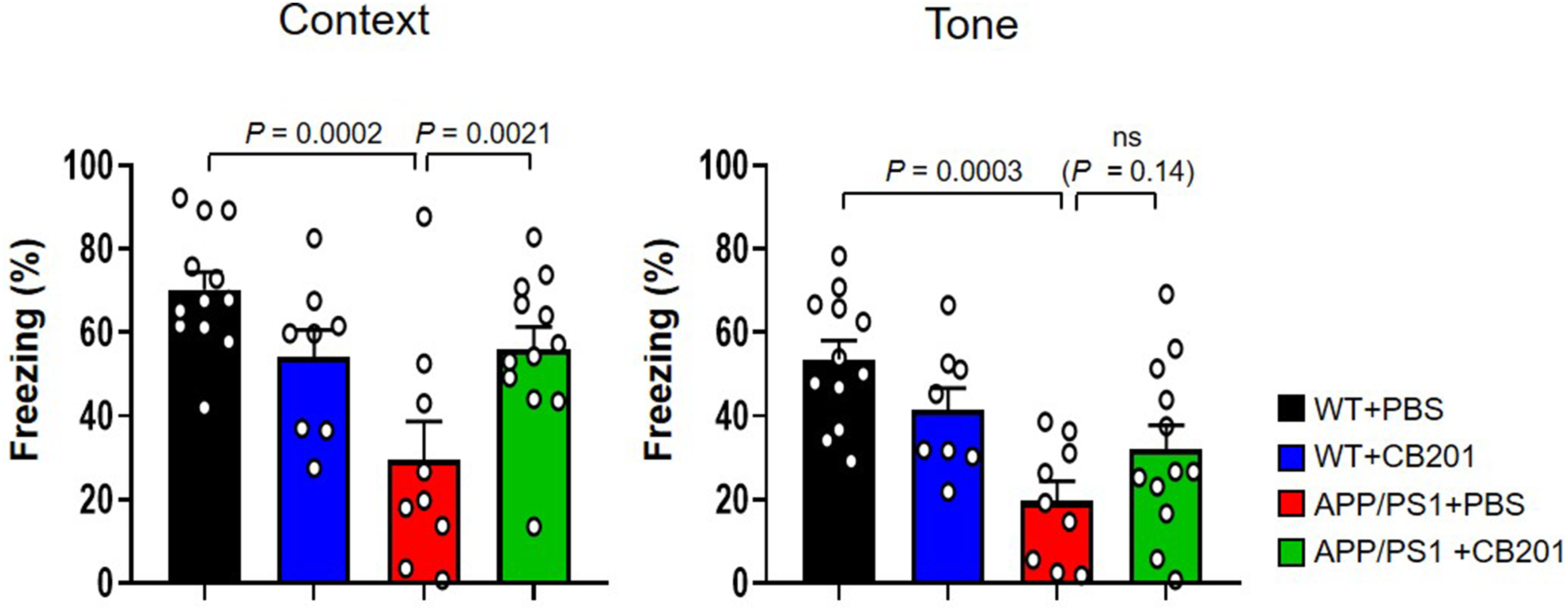
Effects of CB201 treatment on contextual memory in the fear conditioning test. (A) Quantification of freezing behavior (%) in the contextual recall phase (day 2). APP/PS1 mice treated with PBS showed significantly reduced freezing compared to WT + PBS controls, indicating impaired contextual memory. Treatment with CB201 significantly rescued this deficit in APP/PS1 mice. WT + PBS (n = 12 mice), WT + CB201 (n = 8 mice), APP/PS1 + PBS (n = 9 mice), APP/PS1 + CB201 (n = 12 mice). (B) Quantification of freezing behavior (%) in the tone-cued recall phase (day 3). APP/PS1 mice treated with PBS exhibited significantly reduced freezing compared to WT + PBS controls, indicating impaired contextual memory. Treatment with CB201 significantly rescued this deficit in APP/PS1 mice. WT + PBS (n = 12 mice), WT + CB201 (n = 8 mice), APP/PS1 + PBS (n = 9 mice), APP/PS1 + CB201 (n = 12 mice). One-way analysis of variance, Tukey's post hoc test. p < 0.05 was considered as statistically significant. ns = not significant. All data are presented as mean ± SEM.
In the contextual fear conditioning test, the WT + PBS group exhibited a high freezing response of approximately 70%, indicating intact memory function. Similarly, the WT + CB201 group had a comparable freezing response of approximately 60%, suggesting that CB201 did not impair memory function. In contrast, the APP/PS1 + PBS group demonstrated a markedly reduced freezing response of approximately 58% (p = 0.0002) compared to WT + PBS group, confirming the impairment of hippocampus-dependent memory typically observed in AD models.
Notably, the APP/PS1 + CB201 group exhibited significant improvement in memory performance, with an 89.8% increase in freezing behavior compared to the APP/PS1 + PBS group (p = 0.0021; Figure 2A).
In the tone-cued fear conditioning test, the WT-PBS group again exhibited a high freezing response of approximately 55%, indicating intact memory function. Similarly, the WT + CB201 group had a comparable freezing response of approximately 40%, suggesting that CB201 did not impair memory function. However, the APP/PS1 + PBS group demonstrated a markedly reduced freezing response of approximately 20% (p = 0.0003).
Although the improvement did not reach statistical significance, the APP/PS1 + CB201 group showed a 62.9% increase in freezing behavior compared to the APP/PS1 + PBS group, indicating a trend toward recovery of tone-associated memory (Figure 2B).
These results suggest that CB201 treatment was associated with improved hippocampus-dependent memory performance in APP/PS1 mice, supporting its potential relevance for cognitive enhancement in this model.
CB201 reduces Aβ and tau accumulation in the brain
To evaluate the changes in Aβ accumulation following CB201 administration in APP/PS1 mice, ThioS staining were performed. ThioS staining showed that APP/PS1 + CB201 mice had reduced Aβ accumulation across the entire brain compared to the PBS-treated group. Specifically, Aβ accumulation in the cortex and hippocampus was reduced by 53.0% and 44.8%, respectively according to a two-tailed student's t test (cortex, p = 0.026; Figure 3A).
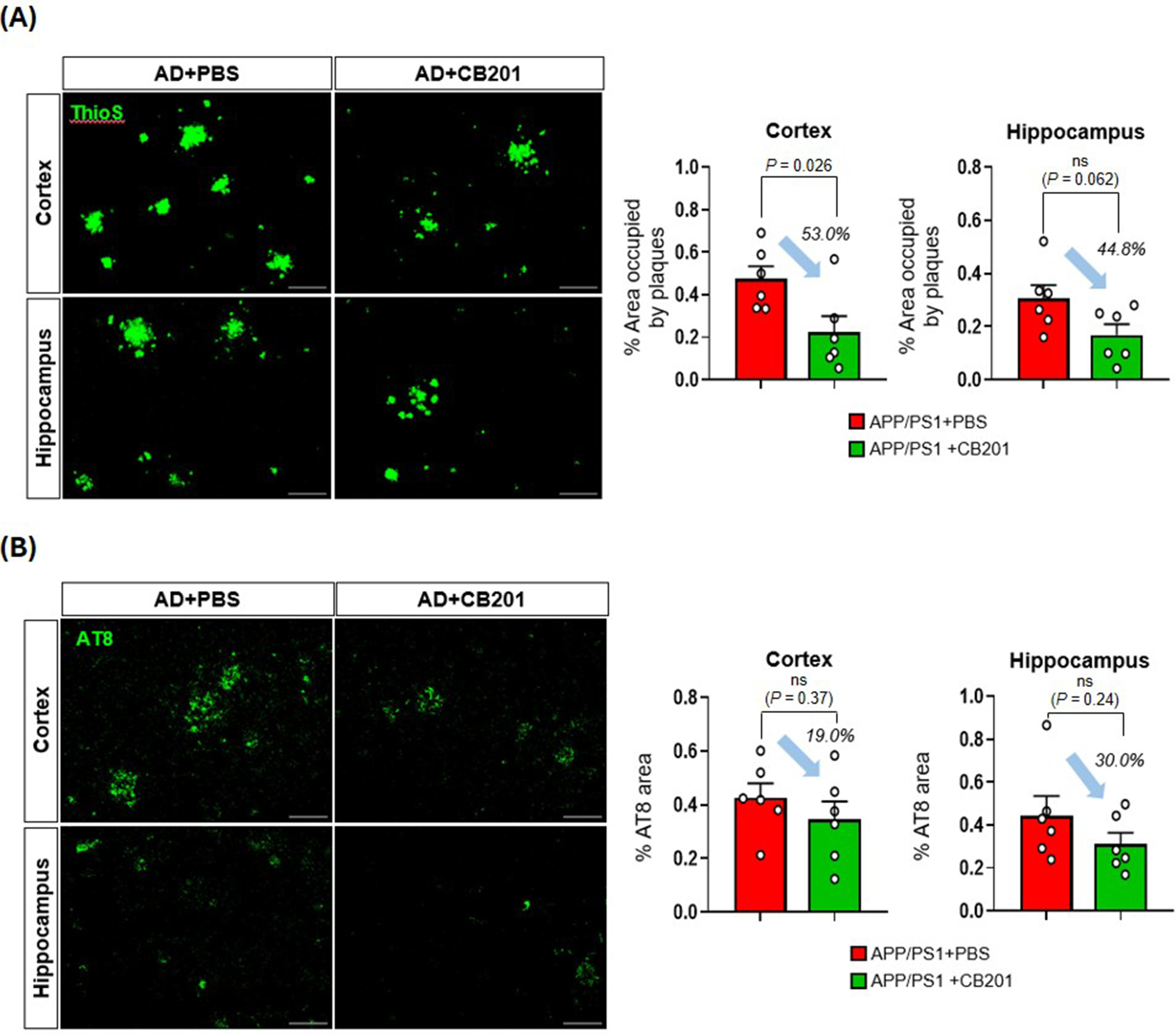
CB201 reduces Aβ and tau accumulation in APP/PS1 mice. (A) Quantification of Aβ accumulation in the cortex and hippocampus by ThioS immunostaining (n = 6 mice per group). Scale bar, 100 μm. (B) Quantification of tau accumulation in the cortex and hippocampus by AT8 immunostaining (n = 6 mice per group). Scale bar, 100 μm. Two-tailed student's t test. p < 0.05 was considered as statistically significant. ns = not significant. All data are presented as mean ± SEM.
We used AT8 staining to examine pathological phosphorylation of tau protein. Although the differences did not reach statistical significance, the CB201-treated group showed an approximately 20-30% reduction in phosphorylated tau protein compared to controls (PBS group; Figure 3B).
Overall, these results suggest that CB201 treatment was associated with reduced Aβ accumulation and a trend toward decreased tau burden in the brain tissues of AD model mice, consistent with a potential effect on pathological features associated with the disease in this experimental context.
Discussion
Here, we used the APP/PS1 transgenic mouse model to evaluate the therapeutic efficacy of CB201 in late-stage AD, focusing on changes in memory, cognitive function, and the accumulation of Aβ and tau proteins in the brain.16,18
This study demonstrated that CB201 treatment was associated with improved cognitive performance in a transgenic model of AD. In the MWM, APP/PS1 mice exhibited spatial learning and memory deficits, as indicated by a prolonged escape latency and spending less time in the target quadrant.17,20 However, CB201-treated mice showed improvements in performance, including a recovered learning curve and a reduction in the area under the curve for latency.15,21,22 Furthermore, the APP/PS1 + CB201 group exhibited increased swim speed and path length compared to untreated APP/PS1 mice, suggesting that CB201 may also contribute to changes in motor-related performance, anxiety-like behavior, or motivational state alongside cognitive enhancement.
A similar trend was observed in the fear conditioning test, which evaluates hippocampus-dependent memory. The APP/PS1 + PBS group showed a marked reduction in freezing behavior during contextual and tone-cued recall, reflecting significant memory impairment.16,21 In contrast, APP/PS1 + CB201 mice had a significant increase in freezing behavior, reaching levels comparable to those of WT controls. 13 Because fear conditioning relies less on locomotor performance, these results provide more direct support for an association between CB201 treatment and improved associative memory performance.
CB201 was also evaluated for its effects on Aβ accumulation and changes in tau phosphorylation. In APP/PS1 mice treated with CB201, ThioS staining revealed significantly reduced Aβ deposition in the cortex and hippocampus. These findings indicate an association between CB201 treatment and reduced fibrillar Aβ burden, which may reflect alterations in Aβ accumulation or clearance processes.8,23,24
AT8 staining, which detects pathological tau phosphorylation, demonstrated a tendency towards reduced tau burden in the CB201 group, indicating that CB201 could also modulate tau pathology and hinder its progression.3,25–27 Nevertheless, the APP/PS1 model used in this study does not develop robust tauopathy or neurofibrillary tangles, and thus the interpretation of tau-related findings is limited. In addition, although CB201 is a monoclonal antibody, an isotype-matched control antibody was not included in the experimental design. While prior studies conducted under similar conditions did not observe significant effects attributable to non-specific immunoglobulin administration, the absence of an isotype control in the current study represents a methodological limitation. Consequently, non-specific immunoglobulin-related effects on behavioral and histological outcomes cannot be fully excluded.
While this study provides robust evidence within male group to establish a foundational understanding of the therapeutic efficacy of CB201 in alleviating cognitive deficits and AD-related pathologies, further investigation involving both sexes will enhance the generalizability of these findings.
Accordingly, tau-related outcomes observed following CB201 treatment are regarded as secondary and exploratory rather than primary endpoints, and the primary conclusions of this study are based on behavioral outcomes and amyloid-related pathology. To address these limitations, further validation using models with more pronounced tau accumulation or human-derived samples will be required to clarify the potential effects of CB201 on tau pathology. Moreover, future studies incorporating appropriate isotype controls will be essential to further validate the specificity and translational relevance of CB201 and to more precisely define its mode of action in the context of AD pathology.
Monocyte-derived macrophages can penetrate the brain via the vasculature, phagocytose Aβ and tau, modulate microglial functions directly and indirectly, and restore inflammatory balance, making them key immune-based therapeutic targets in AD.28–31 In addition, activation of microglia can promote the clearance of Aβ and phosphorylated tau through phagocytosis and enhance neuroprotection via the release of anti-inflammatory cytokines.23,24,29,31 Therapeutic modulation toward a protective microglial phenotype may reduce neuroinflammation and mitigate disease progression.29,32–34 Thus, microglial activation may play an important role in restoring immune homeostasis and alleviating AD pathology.
Based on prior literature and the known immunological role of CD300c, these immune-related pathways may represent potential mechanisms through which CB201 exerts its effects; however, the present study did not directly assess microglial or macrophage activation, cytokine production, or immune signaling pathways. Accordingly, in this study we interpret the immunomodulatory implications of CB201 as preliminary and hypothesis-generating rather than as experimentally established mechanisms.
Conclusions
These findings collectively suggest that in the APP/PS1 model, CB201 may be associated with neuroprotective effects, potentially involving modulation of key pathophysiological processes implicated in AD, including synaptic plasticity, neuroinflammation, and neurodegeneration.1,4,15,27–29 More specifically, CB201 treatment was associated with improved cognitive performance and reduced amyloid pathology in this model, while the underlying cellular and molecular mechanisms remain to be elucidated. The consistent improvements observed in both spatial and associative memory paradigms support the potential relevance of CB201 for modulating cognitive function related to contextual and tone-cued memory in this experimental context.15,20–22 Moreover, the simultaneous reduction of Aβ and tau pathology positions CB201 as a potential therapeutic candidate within the context of late-stage pathology in the AD.3,8,25–27 Further investigation is warranted to elucidate the molecular and cellular interactions between the innate immune system and CB201, including detailed analyses of microglial and macrophage activation states, measurement of inflammatory cytokines, and functional assays assessing phagocytic activity and immune signaling pathways, to determine how CB201 may influence immune-related processes in the context of amyloid and tau clearance.
Limitations
Because the primary aim of the present study was to evaluate the therapeutic efficacy of the fully human anti-CD300c antibody CB201 in the APP/PS1 mouse model at the behavioral and pathological levels, several limitations should be acknowledged. First, although CB201 is a monoclonal antibody, an isotype-matched control antibody was not included in the experimental design. While prior studies conducted under similar conditions did not observe significant effects attributable to non-specific immunoglobulin administration, the absence of an isotype control in the current study represents a methodological limitation. Consequently, non-specific immunoglobulin-related effects on behavioral and histological outcomes cannot be fully excluded. Second, due to this limitation, the treatment-associated improvements observed in behavioral performance and pathological markers should be interpreted with caution. The findings demonstrate phenotypic changes associated with CB201 administration; however, they do not unequivocally establish antibody-specific or target-specific mechanisms. Accordingly, definitive conclusions regarding the specificity of CB201-mediated effects cannot be drawn from the present data alone. In addition, the study was conducted using a single transgenic mouse model (APP/PS1) and included only male animals, which may limit the generalizability of the findings across disease stages, genetic backgrounds, and sexes. Furthermore, the study focused on behavioral and pathological outcomes and did not include direct mechanistic analyses, such as immune cell profiling or signaling pathway assessment.
Future studies incorporating appropriate isotype controls, additional animal models, both sexes, and mechanistic investigations will be essential to further validate the specificity and translational relevance of CB201 and to more precisely define its mode of action in the context of AD pathology.
Footnotes
Acknowledgements
The authors have no acknowledgments to report.
Ethical considerations
The Institutional Animal Care and Use Committee (IACUC) of Kyungpook National University approved all animal procedures (Approval No. KNU2023-0532).
Consent to participate
Not applicable
Consent for publication
Not applicable
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Korea Drug Development Fund funded by the Ministry of Science and ICT; Ministry of Trade, Industry and Energy; and Ministry of Health and Welfare, Republic of Korea [grant number RS-2021-DD121403 (1711200672) to Jae-Won Jeon] and the Korea Dementia Research Center funded by the Ministry of Science and ICT; Ministry of Health and Welfare; and Korea Health Industry Development Institute, Republic of Korea [grant number RS-2023-KH-136029 to Jae-Won Jeon].
Declaration of conflicting interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Suin Lee, Sujin Park, Jeongeun Kim and Jae-Won Jeon were employed by the company CentricsBio, Inc. All authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Data availability statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.