
Abstract
Background
Immunological dysregulation is a hallmark of Alzheimer's disease (AD), a neurodegenerative disorder characterized by amyloid-β (Aβ) plaque accumulation and hyperphosphorylated Tau protein pathology. AD is associated with altered humoral immunity, which may play a role in its pathogenesis.
Objective
This study aimed to investigate alterations in plasma levels of B lymphocyte-associated cytokines and their clinical relevance in AD.
Methods
We performed quantitative detections of 13 cytokines associated with B lymphocytes (TNF-β, IL-13, IFN-γ, TNF-α, IL-2, BAFF, IL-6, CD40L, IL-10, IL-12p70, IL-4, IL-17A, and IL-7) in plasma and analyzed their associations with cognitive functions and biomarkers of AD.
Results
We found that plasma levels of CD40L, BAFF, TNF-β, IL-6 and IL-17A were increased in AD patients. However, the plasma IL-10 concentrations were decreased in Aβ-PET+ subjects. Plasma levels of CD40L, BAFF, TNF-β, IL-6, IL-17A were negatively associated with the plasma Aβ42/40 ratio. Plasma levels of IL-10 were negatively associated with pTau181. Plasma levels of BAFF were negatively associated with MMSE scores.
Conclusions
These findings demonstrate an altered B lymphocyte-associated cytokine secretion profile in AD patients, which correlates with the clinical severity and biomarkers of the disease.
Introduction
Alzheimer's disease (AD) is the most common form of dementia among the elderly population. 1 AD is manifested by progressive cognitive decline and causes a heavy burden to the society and families.2,3 Amyloid-β (Aβ) is the major pathological agent and therapeutic target in AD.4,5 Aβ accumulation in the brain triggers a downstream cascade, including tau hyperphosphorylation, neuroinflammation, and neuronal degeneration. 6
In recent years, Aβ-targeted immunotherapies have achieved a slowing of cognitive decline by substantially clearing Aβ plaques.7–10 However, strategies capable of halting or reversing disease progression are still lacking, underscoring the complexity of the AD pathogenesis. 11 This highlights the importance of further elucidating AD pathological mechanisms to identify potential therapeutic targets. Over the past decades, large-scale population-based genetic studies have identified multiple AD risk gene loci, with approximately half being associated with immune functions, 12 suggesting a strong connection between immune dysregulation and the development of AD.
B lymphocytes are critical effector cells in humoral immunity, and B cell aging represents a significant component of immunosenescence.13,14 The senescent B cell subpopulation is defined as age-associated B cells (ABCs), characterized by a restricted B cell receptor (BCR) repertoire, diminished antibody responses, impaired ability to combat infections or respond to vaccines, 15 increased production of pathological autoantibodies targeting self-antigens, and proinflammatory phenotypic transformation of B cells. 16 We have previously reported that the phenotype of B lymphocytes were altered in AD patients and associated with the clinical severity of the disease. 17
In this study, we investigated the association between B-cell-associated cytokines with cognitive functions and biomarkers of AD. The findings are valuable for understanding the pathogenesis of AD from an immune perspective.
Methods
Participants
Individuals reporting memory complaints were recruited from the Department of Neurology at Daping Hospital and screened for eligibility. A total of 44 AD patients and 42 non-AD individuals confirmed by amyloid-PET scanning were recruited from the Chongqing Aging and Dementia cohort, Daping Hospital. Subjects were excluded if they fulfilled any of the following conditions: (1) had severe cardiac, pulmonary, hepatic, or renal failure, or had any type of tumor; (2) had any type of autoimmune diseases that could have altered humoral immunity phenotypes; (3) had chronic infectious disease or acute infections during the past 4 weeks which may have altered inflammatory factors; (4) refused to participate. Written informed consent for participation and blood sampling was obtained from all participants and their legal representatives.
General cognitive function was initially assessed using the Mini-Mental Status Examination (MMSE). Participants with a MMSE score below 27 subsequently underwent a comprehensive neuropsychological evaluation, including the Clinical Dementia Rating (CDR), Montreal Cognitive Assessment (MoCA), and Activities of Daily Living (ADL) scales. The CDR scale categorized participants as cognitively normal (CDR=0), mild cognitive impairment (MCI; CDR=0.5), or dementia (CDR≥1). All participants underwent structural MRI, amyloid-PET imaging with the 18F-florbetapir tracer, and fluorodeoxyglucose (FDG)-PET to confirm the etiology of cognitive impairment and verify amyloid-negative status. According to the AMYPAD consortium recommendations, a centiloid value below 10 excludes the presence of Aβ pathology, whereas a value above 30 is indicative of pathological Aβ burden. Accordingly, a centiloid value of 30 or higher was defined as Aβ-positive. 18
All participants underwent APOE genotyping and a standardized panel of blood tests, including complete blood count, fasting glucose, lipid profile (cholesterol), hepatic and renal function markers, and levels of vitamin B12, folic acid, and thyroxine. A diagnosis of AD was established in accordance with the National Institute on Aging and Alzheimer's Association (NIA-AA) diagnostic criteria.19,20 The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Institutional Review Board of Daping Hospital (Ethical Approval Number: YYLS2023.147).
Detection of B cell-secreted cytokines
Approximately 10 mL of blood was collected from each participant into EDTA tubes. The samples were left to stand at room temperature for 30 min, then centrifuged at 2000 × g for 10 min to separate the plasma from the cellular components. The plasma samples were stored at −80°C for further use.
Plasma levels of 13 B lymphocyte-associated cytokines (TNF-β, IL-13, IFN-γ, TNF-α, IL-2, BAFF, IL-6, CD40L, IL-10, IL-12p70, IL-4, IL-17A, IL-7) were quantified using the RayPlex® Human B Cell Cytokine Array 1 (cat. no. FAH-BC-1-96, RayBiotech, USA) via bead-based flow cytometry. This panel was deliberately selected to comprehensively capture the functional diversity of B lymphocyte-mediated humoral immunity and its potential relevance to AD pathogenesis. The selection encompasses: (1) key regulators of B cell survival, differentiation, and activation, including BAFF (a critical survival and maturation factor) 21 and CD40L (which mediates T cell-dependent B cell activation and germinal center formation) 22 ; (2) signature cytokines defining distinct T-helper cell subsets that guide B cell responses—Th1-type (IFN-γ, IL-2, TNF-α, TNF-β, IL-12p70), Th2-type (IL-4, IL-13), and Th17-type (IL-17A, IL-6)—thereby capturing the major axes of T cell help that shape B cell effector functions 23 ; (3) the immunoregulatory cytokine IL-10, produced by regulatory B cells and other immune cells, which serves as a critical counterbalance to proinflammatory responses 24 ; and (4) the homeostatic cytokine IL-7, essential for B cell development and maintenance of peripheral B cell pools. 25 This comprehensive panel was therefore designed to provide a broad profile of the humoral immune environment, encompassing B cell-intrinsic regulatory molecules, T cell-derived helper signals, and effector cytokines that collectively shape B cell responses.
Plasma samples were diluted 1:1 with phosphate-buffered saline (PBS), achieving a final 4-fold dilution after mixing with microbeads. Lyophilized protein standards were reconstituted in Human Serum/Plasma Diluent and serially diluted (Std1–Std7), alongside negative controls. Following reagent preparation (vortexed bead cocktail, 1X Wash Buffer, and Streptavidin-PE working solution), 25 µL of bead suspension and 25 µL of prepared samples/standards were loaded into Filter 96-well plates. After overnight incubation at 4°C with shaking at 1000 rpm, unbound analytes were removed via two washes with 1X Wash Buffer. Biotinylated detection antibodies (1-h incubation) and Streptavidin-PE (30-min incubation) were sequentially applied for signal amplification. Beads were resuspended in 150 µL Wash Buffer and analyzed using a flow cytometer equipped with blue/green (PE: 488/578 nm) and red (APC: 640/660 nm) lasers. Filter plates and standardized protocols minimized inter- and intra-assay variability (CVs <10%), with standard curve linearity confirmed (R2 > 0.98). All samples were measured in a single batch in duplicate, and the average of the two replicates was used for statistical analyses.
Plasma biomarker detection
The detection of plasma biomarkers of AD was determined according to our previous protocol. 26 Plasma biomarkers, including pTau181 (cat. no. 81288, Lumipulse G, USA), Aβ40 (cat. no. 81298, Lumipulse G, USA), and Aβ42 (cat. no. 81301, Lumipulse G, USA), were quantified using the Fujirebio Lumipulse G1200 automated immunoassay system. The instrument was calibrated according to the manufacturer's standardized protocols, with operational parameters and assay conditions optimized to ensure analytical precision. Rigorous quality control procedures, including adherence to internal validation criteria and manufacturer-recommended standard operating procedures (SOPs), were implemented to verify result accuracy. Prior to analysis, samples were pre-incubated with target-specific monoclonal antibodies (AT270 for pTau181, HJ2 for Aβ40, and HJ7.4 for Aβ42, which are included in the plasma biomarker kit) to ensure selective binding. Quantitative measurements were performed via the integrated chemiluminescent enzyme immunoassay (CLEIA) technology of the Lumipulse platform. Biomarker concentrations were derived using the Lumipulse software package (version 2.5), with raw data normalized against validated reference ranges and standard curves generated from kit-provided calibrators.
Statistical analyses
Demographic and clinical characteristics of the participants were analyzed using descriptive statistics. Continuous variables were expressed as means ± standard deviations (SDs) or median (IQR), while categorical variables were reported as absolute frequencies and percentages. For group comparisons, independent samples t-tests were used to evaluate differences in B-cell related cytokine levels between Aβ-PET+ and Aβ-PET− participants. No formal normality test was performed. Given the sample size of over 40 per group, the central limit theorem ensures that the sampling distribution of the mean approximates normality, rendering the t-test robust to moderate violations of the normality assumption. 27 Visual inspection of histograms and boxplots confirmed that the data were approximately symmetric and free of extreme outliers. For correlation analyses, Spearman rank correlation coefficient was employed to assess the associations between B-cell associated cytokines and AD biomarkers as well as cognitive measures. This non-parametric method does not assume bivariate normality; therefore, an assessment of normality was not required for these analyses. No formal outlier removal or Winsorization was performed, and we explicitly note the criteria that would have been used if exclusion had been considered (e.g., values beyond ±3 standard deviations or outside 1.5×IQR). All statistical analyses were two-tailed, and significance was defined as a threshold of p < 0.05. GraphPad Prism 10 was used for performing statistical analyses and generating the figures.
Results
Sociodemographic characteristics of participants
This study included 44 Aβ-PET+ subjects and 42 Aβ-PET− subjects. Table 1 shows the sociocharacteristics of the participants. There were no significant differences in the mean age, mean educational attainment, sex distribution, proportion of APOE ε4 carriers, and ADL scores between the two groups. Aβ-PET+ subjects had lower MMSE, MoCA scores, and higher CDR scores than did Aβ-PET− subjects (Table 1).
Demographic characteristics of participants.
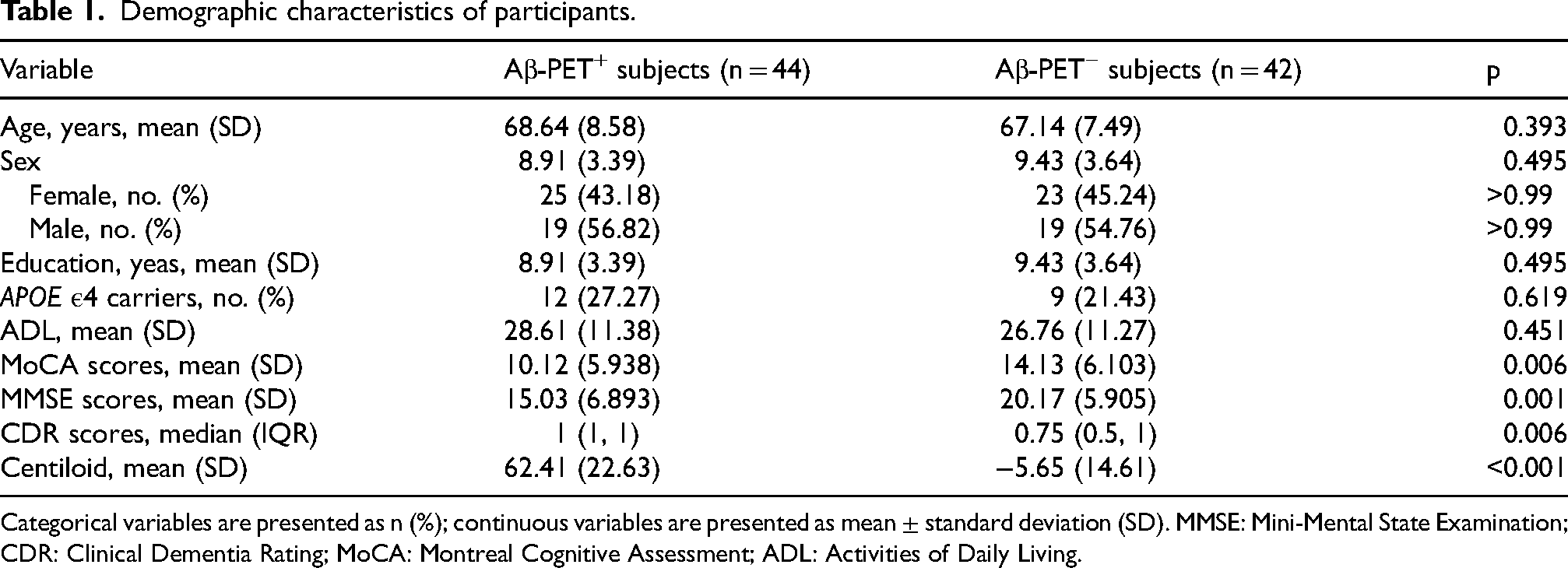
Categorical variables are presented as n (%); continuous variables are presented as mean ± standard deviation (SD). MMSE: Mini-Mental State Examination; CDR: Clinical Dementia Rating; MoCA: Montreal Cognitive Assessment; ADL: Activities of Daily Living.
Comparison of B lymphocyte-associated cytokines between Aβ-PET+ and Aβ-PET− subjects
We found that Aβ-PET+ subjects had significantly higher plasma levels of TNF-β, BAFF, IL-6, CD40L and IL-17A than Aβ-PET− subjects (Figure 1A-C, H, L). On the contrary, plasma levels of IL-10 were significantly lower in Aβ-PET+ subjects in comparison with Aβ-PET− subjects (Figure 1I). However, the plasma levels of IL-13,IFN-γ, TNF-α, IL-2, IL-12p70, IL-4 and IL-7 were comparable between Aβ-PET+ and Aβ-PET− subjects (Figure 1D-G, J, K, M). These findings indicate that Aβ-PET+ subjects may have an altered B lymphocytes-associated cytokine secretion profile.
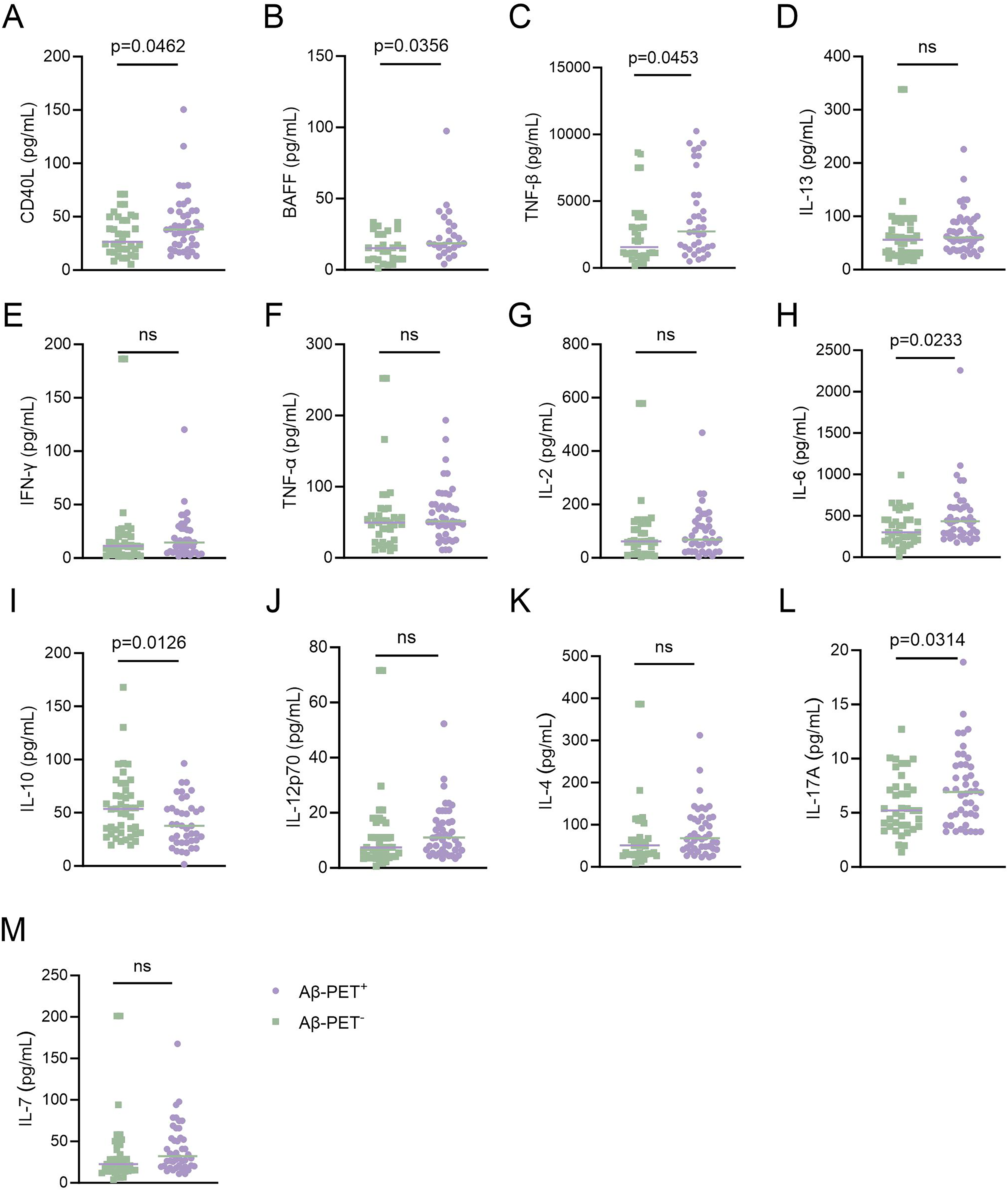
Comparison of plasma B lymphocyte-associated cytokines between Aβ-PET+ and Aβ-PET− subjects. Comparison of the plasma levels of (A) CD40L (t = 2.026, df = 77, p = 0.046) (B) BAFF (t = 2.161, df = 49, p = 0.0356) (C) TNF-β (t = 2.039, df = 69, p = 0.0453) (D) IL-13 (t = 0.2219, df = 82, p = 0.8250) (E) IFN-γ (t = 0.4046, df = 74, p = 0.6870) (F) TNF-α (t = 0.2786, df = 79, p = 0.7813) (G) IL-2 (t = 0.2096, df = 69, p = 0.8346) (H) IL-6 (t = 2.312, df = 81, p = 0.0233) (I) IL-10 (t = 2.552, df = 81, p = 0.0126) (J) IL-12p70 (t = 0.4310, df = 83, p = 0.6676) (K) IL-4 (t = 0.7181, df = 82, p = 0.4747) (L) IL-17A (t = 2.191, df = 80, p = 0.0314) (M) IL-7 (t = 0.8310, df = 83, p = 0.4083) between Aβ-PET+ and Aβ-PET− subjects. Independent t-test. The bar represents the median value.
Associations of B lymphocyte-associated cytokines with cognitive functions and biomarkers of AD
Next, we performed correlation analyses of plasma levels of B lymphocyte-associated cytokines with biomarkers of AD (Figure 2A-M and Figure 3A-M) and cognitive functions (Figure 4A-M). We found that plasma levels of CD40L, BAFF, TNF-β, IL-6, and IL-17A were negatively associated with Aβ42/40 (lower Aβ42/40 indicate higher amyloid burden in the brain) (Figure 2A-C, J, 1, K). However, no significant association was found between plasma levels of Aβ42/40 with IL-13, IFN-γ, TNF-α, IL-10, IL-12p70, IL-4, IL-7, or IL-2 (Figure 2D-F, H-J, L, M). In addition, plasma levels of pTau181 were not associated with CD40L, BAFF, IL-13, IFN-γ, TNF-α, IL-6, IL-12p70, IL-4, IL-7, or IL-2 (Figure 3A, B, D-G, 1, J, L, M). Plasma levels of TNF-β were positively associated with pTau181 (higher pTau181 indicates more severe neurodegeneration in the brain) (Figure 3C). Plasma levels of IL-10 were negatively associated with pTau181 (Figure 3H). Plasma levels of CD40L, TNF-β, IL-13, IFN-γ, TNF-α, IL-6, IL-10, IL-12p70, IL-4, IL-17A, IL-7, or IL-2 were not associated with MMSE scores (Figure 4A, C-M). Plasma levels of BAFF were negatively associated with MMSE scores (Figure 4B). These findings suggest that plasma levels of B lymphocyte-associated cytokines were associated with the severity of AD as reflected by cognitive functions and biomarkers.
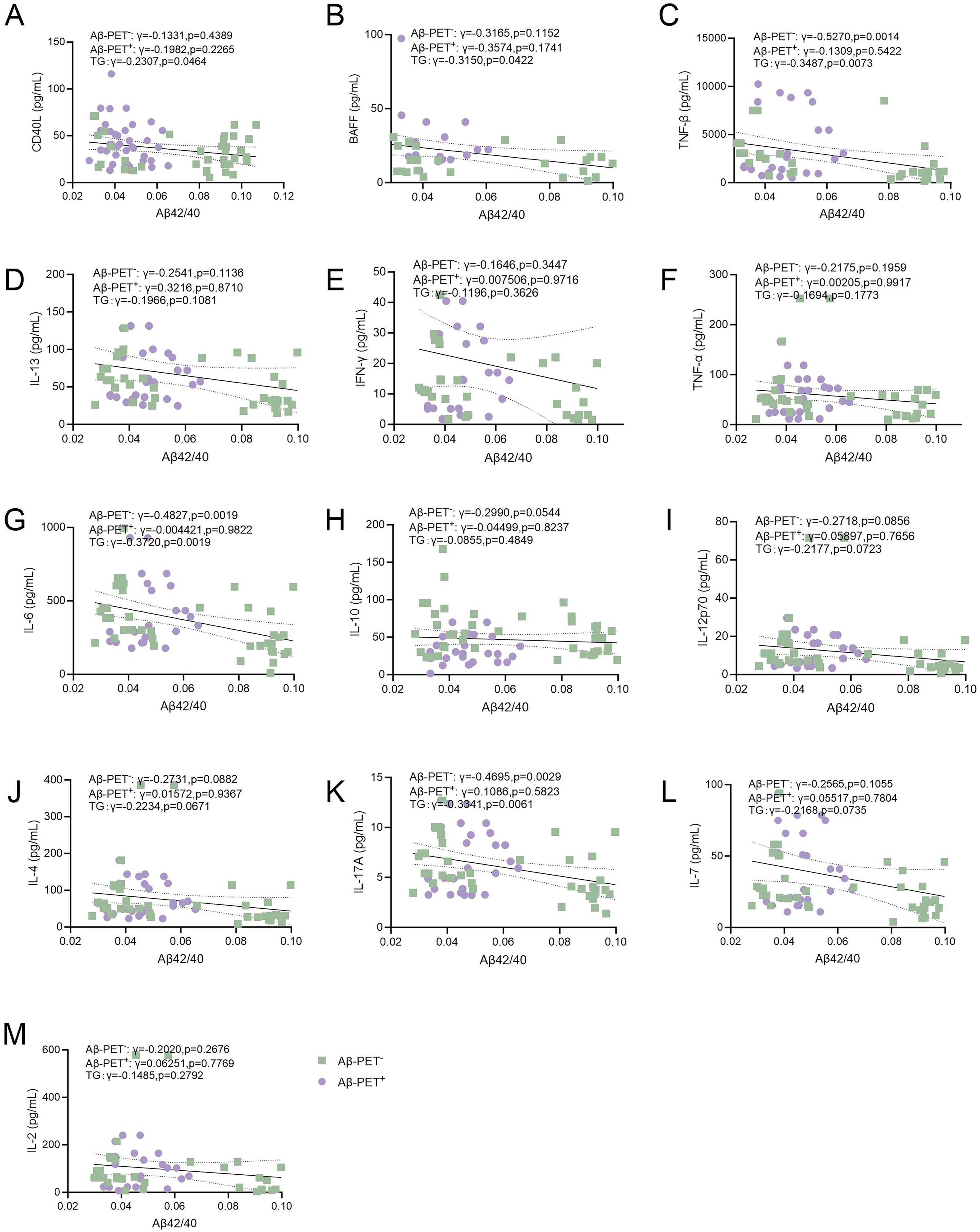
Association of B lymphocyte-associated cytokines with Aβ42/40. Correlations of plasma levels of (A) CD40L (B) BAFF (C) TNF-β (D) IL-13 (E) IFN-γ (F) TNF-α (G) IL-6 (H) IL-10 (I) IL-12p70 (J) IL-4 (K) IL-17A (L) IL-7 (M) IL-2 with Aβ42/40. Spearman correlation analysis. The solid line represents the fitted regression line, and the dashed lines denote the 95% confidence interval.
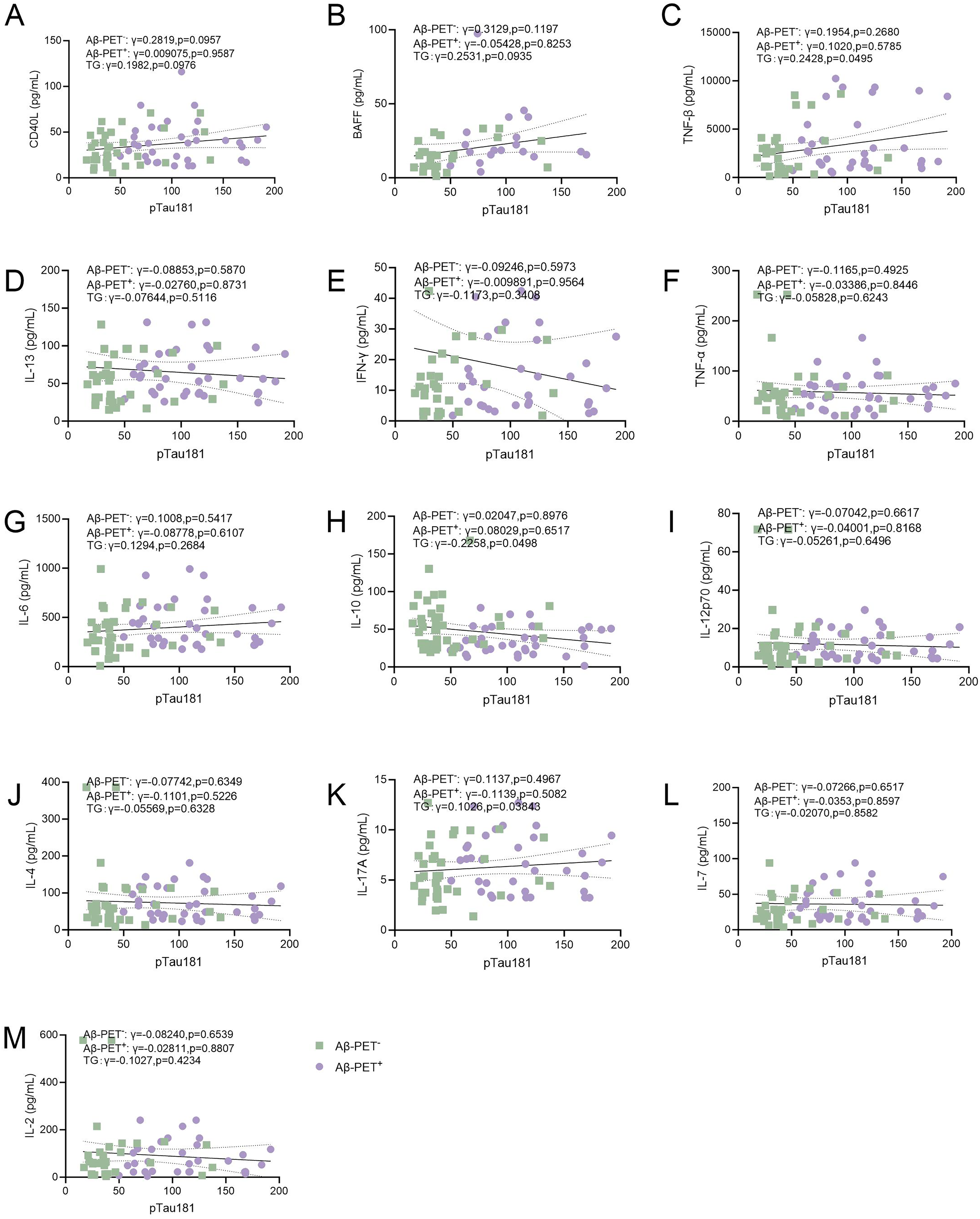
Association of B-lymphocyte secreted cytokines with pTau181. Correlations of plasma levels of (A) CD40L (B) BAFF (C) TNF-β (D) IL-13 (E) IFN-γ (F) TNF-α (G) IL-6 (H) IL-10 (I) IL-12p70 (J) IL-4 (K) IL-17A (L) IL-7 (M) IL-2 with pTau181. Spearman correlation analysis. The solid line represents the fitted regression line, and the dashed lines denote the 95% confidence interval.
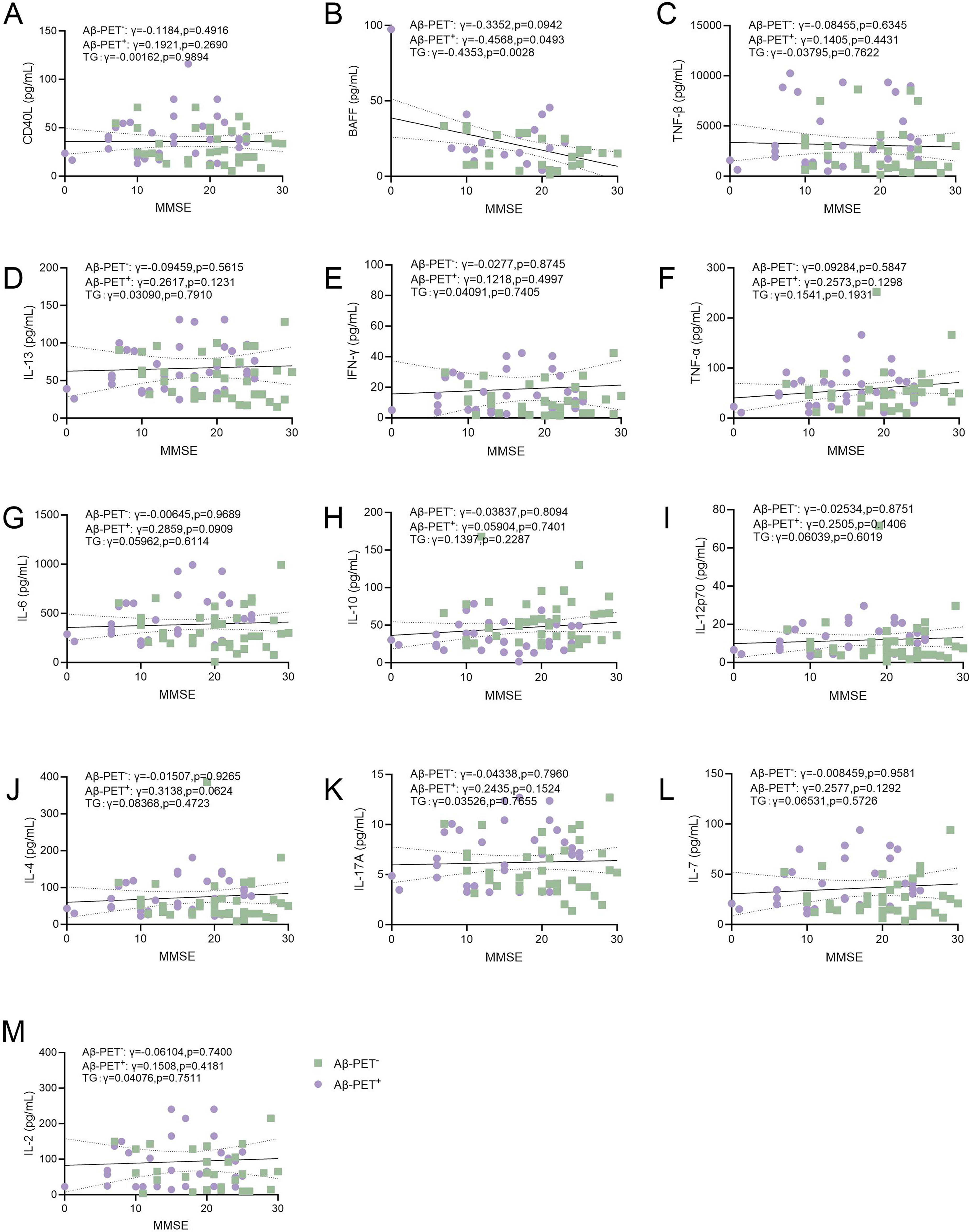
Association of B-lymphocyte secreted cytokines with cognitive functions. Correlations of plasma levels of (A) CD40L (B) BAFF (C) TNF-β (D) IL-13 (E) IFN-γ (F) TNF-α (G) IL-6(H) IL-10 (I) IL-12p70 (J) IL-4(K) IL-17A (L) IL-7 (M) IL-2 with MMSE scores. Spearman correlation analysis. The solid line represents the fitted regression line, and the dashed lines denote the 95% confidence interval.
Discussion
In this study, we found that plasma levels of TNF-β, BAFF, IL-6, CD40L, and IL-17A were increased but the levels of IL-10 were decreased in Aβ-PET+ subjects. Furthermore, plasma levels of these B lymphocyte-associated cytokines were associated with cognitive functions and biomarkers of AD.
The role of B lymphocytes in the pathogenesis of AD has garnered substantial support in recent years. 13 Previous studies have reported alterations in the gene expression profile of B lymphocytes during the disease progression. 16 In addition, recent findings indicate that depletion of B lymphocytes aids in alleviating cognitive impairment and AD-type pathologies, 28 suggesting a potential role of B cells in AD. To support this notion, we recently found that fingolimod, which is a sphingosine-1-phosphate (S1P) receptor modulator and inhibits the infiltration of lymphocytes into the brain, could effectively ameliorate multiple pathological features of AD. 29 These findings collectively suggest that B lymphocyte may have a pathogenic role in the development of AD.
B cells may promote the development of AD through several mechanisms. For example, they produce brain-reactive autoantibodies, which may penetrate the brain through the leakage of the blood-brain barrier. In the AD brain, increased autoantibodies were observed in association with Aβ accumulation. 30 Our group has identified multiple pathogenic autoantibodies in AD patients, which promoted the development of AD31–34. We previously reported that autoantibodies targeting BACE1, which is a major β-secretase for generating Aβ, can promote Aβ production via activating BACE1 during the development of AD. 35 Furthermore, we also identified dysregulated B-cell subset distributions and altered activation profiles in AD patients. Notably, AD patients demonstrated a reduced proportion of switched memory (SwM) B cells and double-negative (DN) B cells, alongside an elevated proportion of unswitched memory (USwM) B cells. Following ex vivo stimulation, B cells from AD patients preferentially adopted a proinflammatory phenotype, characterized by elevated frequencies of granulocyte-macrophage colony-stimulating factor (GM-CSF), IFN-γ, and TNF-α-producing subsets. These B-cell phenotypic alterations correlated significantly with cognitive decline and biomarkers of AD pathology. 26 As observed in the present study, the secretory phenotype of B cells in Aβ-PET+ subjects is also altered, being characterized by increased proinflammatory cytokines and decreased anti-inflammatory cytokines in circulation. The findings from this study, demonstrating that alterations in CD40L, BAFF, TNF-β, IL-6, IL-17A and IL-10 are associated with the clinical phenotype of AD, align with our earlier observations.
Emerging evidence highlights proinflammatory cytokines produced by B lymphocytes, such as TNF-β, IL-6, and IL-17A, key contributors to AD pathogenesis. 36 These molecules are consistently observed to be elevated in AD models and patient samples, with studies linking their overexpression to neuroinflammatory cascades. 37 CD40, an immunoregulatory molecule ubiquitously expressed on immune cells, plays a critical role in maintaining BBB integrity through its interaction with CD40 ligand CD40L on B lymphocytes. 38 Dysregulation of this signaling pathway may compromise immune system homeostasis. Emerging evidence indicates the CD40/CD40L dyad contributes to AD pathogenesis by aberrant driving microglial activation, a process that promotes excessive production of proinflammatory cytokines in the brain.39,40 BAFF and BAFF receptors signaling pathway controls the fate of B lymphocytes 41 and is suggested to be associated with neuronal survival, 42 but its role in AD has not been reported yet. Our finding of elevated plasma BAFF in Aβ-PET+ subjects aligns with the observed humoral immune dysregulation in AD. Notably, BAFF levels correlated positively with brain amyloid burden and negatively with cognitive scores (lower Aβ42/40 ratio indicates higher cerebral amyloid burden), suggesting a potential link between BAFF-driven B cell activity and disease progression. In addition, BAFF inhibitors (e.g., belimumab) are approved for systemic autoimmune diseases. 43 Given the shared features of immune dysregulation,35,44,45 investigating whether BAFF pathway modulation could temper the potentially pathogenic B cell response in AD represents a novel therapeutic avenue. Future studies should determine if BAFF elevation is a compensatory response or a primary driver of neuroinflammation in AD. In contrast to the elevated proinflammatory cytokines, we observed a significant decrease in plasma IL-10 levels in Aβ-PET+ subjects. IL-10 is a potent anti-inflammatory cytokine produced by regulatory B cells (Bregs) and other immune cells, known to mitigate neuroinflammation and promote neuronal survival. 46 Reduced IL-10 levels align with our previous findings where IL-10 producing cells were decreased in AD. 17 This finding is further corroborated by two recent systematic reviews demonstrating a shift toward pro-inflammatory cytokines (e.g., increased IL-1β, IL-6, and TNF-α) alongside decreased anti-inflammatory cytokines (such as IL-10) in AD.47,48 This process possibly involves impaired Breg function, thus failing to restrain the proinflammatory environment in AD. This decrease in IL-10, coupled with elevated proinflammatory cytokines like IL-6 and IL-17A, points to a profound imbalance between pro- and anti-inflammatory humoral signals, which may collectively exacerbate neuroinflammation and pathology.
In conclusion, this study provides clinical evidence that the humoral immune phenotype differs between Aβ-positive AD patients and Aβ-negative cognitive impairment patients. First, its cross-sectional design establishes correlative associations but cannot determine causality or temporal dynamics between cytokine changes and AD progression. Second, the modest sample size, while sufficient to detect the reported significant differences, necessitates validation in larger, independent cohorts to confirm the robustness and generalizability of our findings. Third, a major limitation of this study is the lack of cellular specificity. Although the cytokines measured are functionally linked to B cell activity and were assessed using a B-cell-focused array, they are not exclusively produced by B lymphocytes—other immune cell types may also contribute to their expression. As plasma levels represent the net output from multiple cellular sources, the observed cytokine alterations may reflect the combined contribution of various immune cells rather than changes in B cells alone. Therefore, our findings do not directly establish B cells as the definitive source of these cytokine variations. Whether these changes originate from B cells, involve other immune populations, or reflect broader immune dysregulation cannot be resolved from the current data. However, together with our previous study showing aberrant B-cell subclass composition, 17 these results further implicate dysregulation of humoral immunity in AD. A critical future direction is to validate these findings in a large, multi-center cohort and to establish longitudinal studies to track cytokine changes over time in relation to cognitive decline and biomarker progression. Furthermore, in vitro and in vivo functional studies are needed to mechanistically dissect how specific B cell-derived cytokines influence AD-related pathophysiology.
Footnotes
Acknowledgements
We thank all participants and their families for their contribution to this study.
Ethical considerations
This work is approved by the Institutional Review Board of Daping Hospital, Third Military Medical University.
Consent to participate
Written informed consent for participation and blood sampling was obtained from all participants and their legal representatives.
Consent for publication
Not applicable
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the National Natural Science Foundation of China (82222023 and 82471461 to Y.H.L.), the National-Level Reserve Talent Program of the Chongqing Health Commission (HBRC202402 to Y.H.L.), the Natural Science Foundation of Chongqing (No. CSTB2022NSCQ-LZX0042 to LYH), and the Program for Innovation Capacity Enhancement of Third Military Medical University (No. 2022XRC04 to LYH).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The data generated in this study are available from the corresponding author upon reasonable request.