
Abstract
Background
Triple-negative breast cancer (TNBC) remains clinically challenging due to its aggressive nature, high recurrence rate, and lack of targeted therapies. To address these limitations, combination chemotherapies that synergistically enhance antitumor efficacy while permitting dose reduction and supporting dose-sparing strategies are urgently needed. Cisplatin has a broad spectrum of anticancer activity. However, its resistance and dose-limiting toxicities constrain clinical benefit. Digitoxin is a cardiac glycoside that has anticancer activity.
Objective
We aimed to assess the nature of interaction between digitoxin and cisplatin in TNBC cells, define the most synergistic area (MSA), and estimate dose-sparing potential.
Methods
We profiled digitoxin-cisplatin effects in MDA-MB-231 TNBC cell line by checkerboard and fixed-ratio designs, quantifying interaction with Chou-Talalay combination index/dose-reduction index by CompuSyn, and multiple matrix models by SynergyFinder.
Results
The CI analysis demonstrated synergy across all tested ratios, with the strongest synergistic interaction at a digitoxin/cisplatin ratio of 1:200 (CI = 0.30 at ED97). Dose-reduction analysis showed up to 27-fold cisplatin sparing (ratio 1:25, ED97). Matrix-based models confirmed synergy (Bliss 6.9, Loewe 18.6, ZIP 7.1, and HSA 14), converging on MSA at 31.25 nM digitoxin and 6.25 µM cisplatin.
Conclusion
Our results highlight a robust synergistic interaction between digitoxin and cisplatin in MDA-MB-231 cells with a dosing window that maximizes cytotoxic effect while potentially reducing cisplatin exposure.
Introduction
Breast cancer remains one of the most prevalent malignancies, 1 and triple-negative breast cancer (TNBC) is an aggressive subtype lacking the expression of estrogen receptors, progesterone receptors, and HER2. It comprises 15-20% of all breast cancers, and is associated with poor prognosis. 2 The absence of druggable molecular targets makes combination chemotherapy a mainstay for TNBC, used to enhance efficacy, delay resistance, and mitigate monotherapy toxicity. 3 Rational drug combinations, however, require quantitative frameworks to distinguish true synergy from simple additivity and to identify dose windows with genuine therapeutic benefit. Prior work from our group demonstrated that natural-product chemosensitizers, such as cucurbitacin B in cisplatin-resistant ovarian cancer 4 and gingerols/cucurbitacins in resistant breast and ovarian models, 5 can restore platinum sensitivity, motivating the search for additional synergistic partners.
Cisplatin is one of the earliest platinum-based drugs to enter clinical use (1970s) and remains a backbone therapy across multiple tumor types. 6 Cisplatin and other platinum-based compounds exhibit a broad spectrum of antitumor activity and are used in the treatment of various cancers, including testicular, lung, ovarian, bladder, cervical, head and neck, and gastric cancers.7,8 Mechanistically, cisplatin interferes with DNA synthesis by forming intra- and inter-strand crosslinks between purine bases, disrupting DNA replication and leading to cell cycle arrest and apoptosis.9,10 However, cisplatin is not highly selective toward cancer cells, and the development of dose-limiting toxicities and acquired resistance remain the principal challenges that constrain its clinical benefit.11–14
Digitoxin is a cardiac glycoside and Na+/K+-ATPase inhibitor that has been explored for anticancer repurposing.15–17 At nanomolar concentrations, digitoxin activates the Na+/K+-ATPase signalosome and suppresses AP-1 and NF-κB-driven transcription of survival and cell-cycle genes,17,18 rapidly extinguishes NFAT-driven c-MYC expression, 19 and depends on Src-EGFR-ERK1/2 effectors coupled to an interferon-response program for its cytotoxicity. 20 Cardiac glycosides trigger a robust DNA damage response characterized by single- and double-strand breaks, ROS-mediated oxidative lesions, and the suppression of homologous recombination and nucleotide excision repair, providing a direct mechanistic basis for sensitization to DNA-crosslinking agents such as cisplatin.21,22 Consistent with this rationale, cardiac-glycoside pretreatment more than doubles cisplatin’s antitumor effect in a syngeneic breast cancer model 23 and produces Src-dependent synergistic cytotoxicity with cisplatin in cervical carcinoma cells. 24 Together, these observations provide a coherent rationale for testing digitoxin as a chemosensitizing partner for cisplatin in TNBC.
To assess the nature of the interaction between digitoxin and cisplatin, we adopted two complementary quantification approaches: fixed-ratio analysis using the Chou-Talalay method (CompuSyn) to obtain combination indices (CI) and dose-reduction indices (DRI) across effect levels, 25 and full-matrix analysis using four reference models in SynergyFinder to calculate synergy scores and map the most synergistic area (MSA).26,27 We profiled digitoxin-cisplatin combinations in MDA-MB-231 cells using a checkerboard design and summarized interaction patterns at both the fixed-ratio and matrix levels to delineate dose windows of potential therapeutic synergy.
Materials and methods
Cell lines and culture
The TNBC cell line MDA-MB-231 was obtained from Nawah Scientific (Cairo, Egypt). Cells were maintained in DMEM high glucose media (Cegrogen Biotech, Germany) supplemented with 1% antibiotic (penicillin (100 U/ml) and streptomycin (1000 µg/ml)) and 10% fetal bovine serum (Biowest, USA) in humidified, 5% CO2 atmosphere at 37°C. Mycoplasma PCR was routinely performed every 2 weeks.28,29 Digitoxin was purchased from Universal Fine Chemicals, Cas No. 71-63-6.
Cell viability assay
MDA-MB-231 cells were seeded at 5000 cells/well in 96-well plates and allowed to adhere overnight at 37°C and 5% CO2. On the next day, digitoxin and cisplatin were added to the cell plate using a checkerboard design with twofold serial dilutions for each agent (cisplatin 1.56-50 µM, digitoxin 7.8-250 nM). After 48 h of drug exposure, 10 µl of MTT solution (5 mg/ml in 1X PBS) was introduced to each well and the plates were incubated for 4 h. To solubilize the resulting formazan crystals, 100 µl of 10% SDS in 0.01 N HCl was added to each well. The plates were then incubated for further 14 h to ensure complete solubilization. Absorbance readings were taken using microplate reader (Azure Biosystems, USA) at wavelength of 570 nm.
The viability experiment was conducted in three independent biological replicates (n = 3). For downstream interaction quantification, the mean normalized response across the replicates was used to construct the response matrix submitted to CompuSyn and SynergyFinder.
Synergy analysis
Drug-interaction quantification was performed under two complementary analytical frameworks to ensure that any observed synergy is robust and not an artefact of a single null-interaction assumption. First, a fixed dose-ratio analysis was conducted using the Chou-Talalay method (CompuSyn), which provides clinically interpretable CI and DRI. Second, the full dose-response matrix was analyzed using four reference models in SynergyFinder, each resting on a different null-interaction assumption. Because the reference surfaces generated by these models are non-equivalent, their numerical synergy scores are not directly interchangeable in absolute magnitude, and the position of their respective MSA can shift between models. Reporting all four scores, together with a conservative consensus, provides cross-validation: an interaction supported simultaneously by frameworks with non-overlapping null assumptions is unlikely to be an artefact of any single reference surface.
Fixed-ratio analysis: Combination index and dose-reduction index
The CI determines the type of drug interaction where CI < 1 indicates synergy, CI = 1 indicates additivity, CI > 1 indicates antagonism.
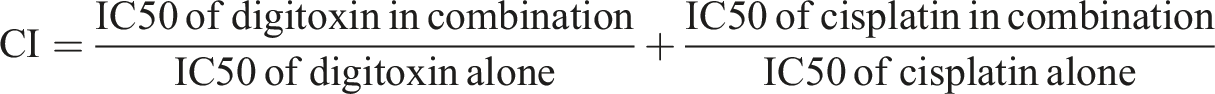
The DRI measures the degree of dose reduction for a given drug in a combination to reach the same effect as the drug alone, where DRI >1 indicates that less of the drug is needed in the combination than when used alone, DRI = 1 indicates no dose reduction, and DRI <1 indicates antagonism or higher dose of the drug is needed in combination.

The CI and DRI were computed by Chou-Talalay equation 25 utilizing CompuSyn software version 1.0.
Matrix-based analysis by SynergyFinder
The full dose-response matrix was analyzed using SynergyFinder under four reference models. 26 For all models, synergy scores are reported in δ-units: positive values indicate synergy, values near zero indicate non-interaction, and negative values indicate antagonism.
Bliss independence model assumes that the two agents act through statistically independent mechanisms, and the expected combined effect is calculated from the product of the single-agent fractional responses. 30 Bliss synergy scores were computed at each dose pair and summarized across the matrix to identify regions of synergy.
Loewe additivity model assumes that the two agents act like dilutions of the same agent and defines non-interaction as dose additivity along isoeffect contours.
The ZIP model builds a no-interaction reference surface by overlaying the single-agent dose-response curves and then computes a synergy score at each dose pair as the deviation of the observed combination effect from that reference. 27
The HSA model evaluates whether the effect of a drug combination is better than the maximum effect achieved by either single drug at the same doses. A combination is called synergistic when the combined effect exceeds the highest single-agent effect. 27
In addition, SynergyFinder provides a conservative Bliss-Loewe consensus synergy score. At each dose pair the consensus expectation is taken as the maximum expected effect across the Bliss, Loewe, and HSA surfaces; the consensus synergy is the deviation of the observed response from this expectation. ZIP is not included in the consensus because it shares the multiplicative survival principle with Bliss and may bias the consensus interpretation.
Results
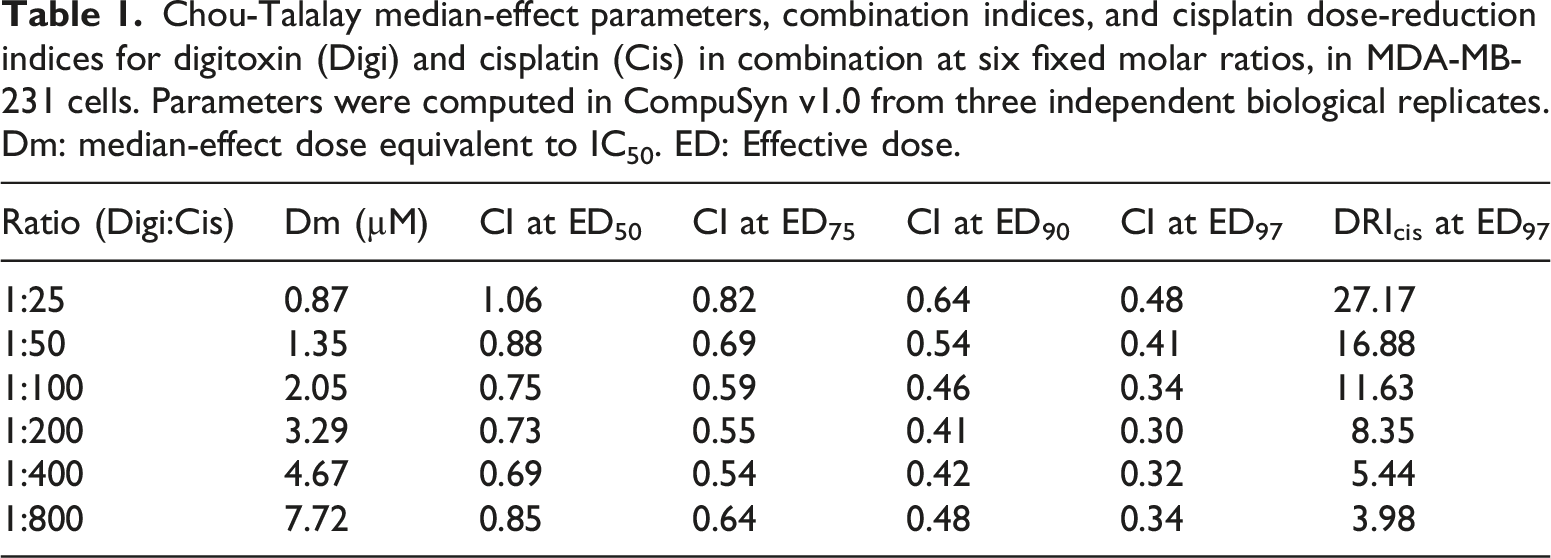
Chou-Talalay median-effect parameters, combination indices, and cisplatin dose-reduction indices for digitoxin (Digi) and cisplatin (Cis) in combination at six fixed molar ratios, in MDA-MB-231 cells. Parameters were computed in CompuSyn v1.0 from three independent biological replicates. Dm: median-effect dose equivalent to IC50. ED: Effective dose.
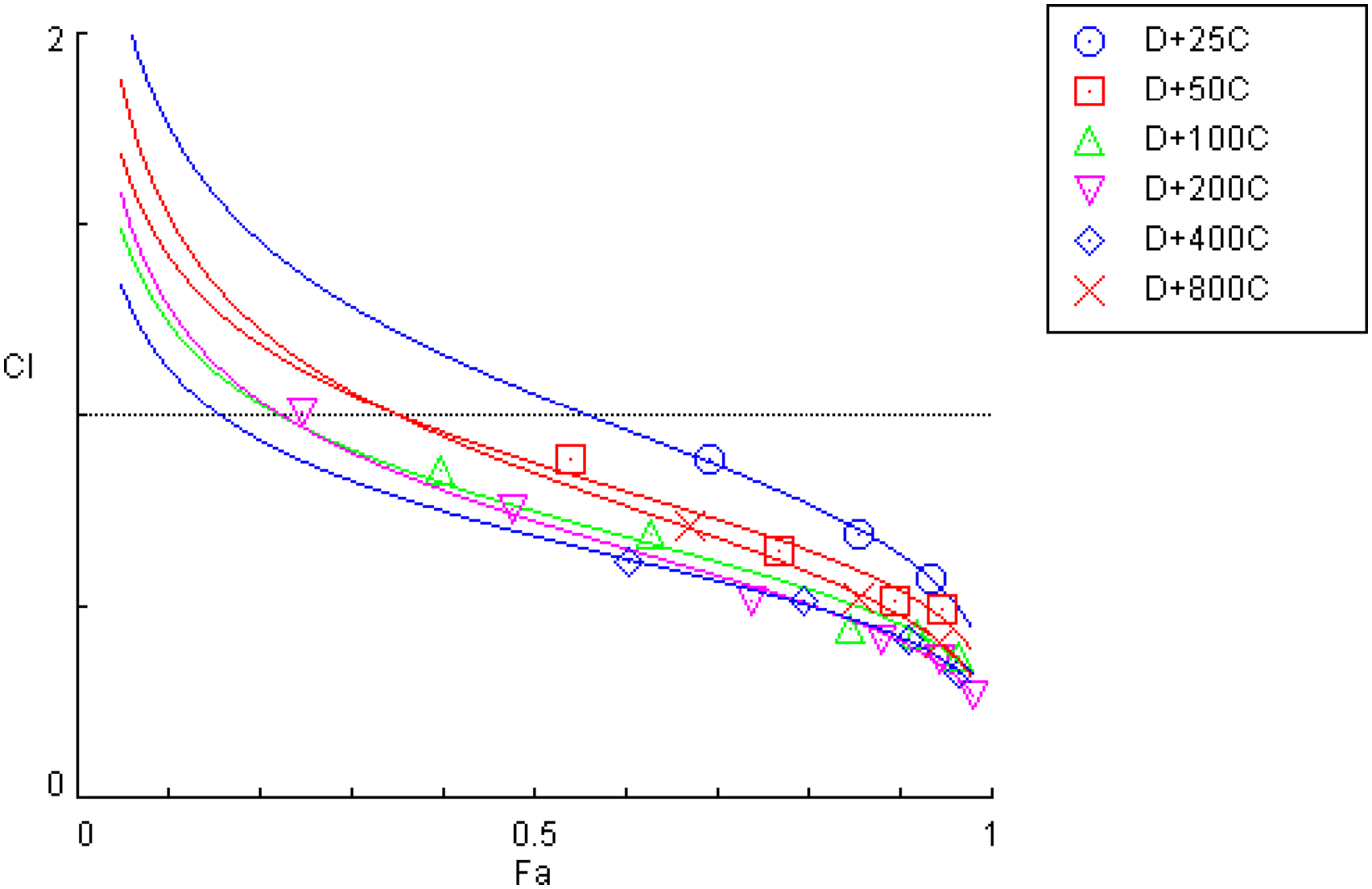
Combination index (CI) versus fraction affected (Fa) plot for the digitoxin-cisplatin combination (D + C) at fixed ratios (D:C = 1:25, 1:50, 1:100, 1:200, 1:400, and 1:800) generated using CompuSyn software. The horizontal reference line indicates CI = 1 (additivity), CI < 1 indicates synergy and CI > 1 indicates antagonism. Numerical CI values at ED50, ED75, ED90, and ED97 for all six ratios are tabulated in Table 1.
DRI further supports these findings and suggests that digitoxin enhances cisplatin efficacy. The strongest cisplatin sparing occurred at ratio 1:25 where DRIcis at ED97 was 27.17 indicating 27-fold dosage reduction compared to monotherapy (Table 1). At higher ratios of cisplatin, the sparing persisted but was smaller; for example, at 1:50 the DRIcis was 16.88 and at 1:100 the DRIcis was 11.63.
After quantifying the interaction between the two drugs at fixed ratios by CompuSyn, we analyzed the full dose-response matrix across a wide range of concentrations, with cisplatin ranging from 1.56 to 50 µM and digitoxin from 7.8 to 250 nM, in SynergyFinder using Bliss, Loewe, ZIP, and HSA models to calculate synergy scores, map the MSA, and cross-validate the results. As shown in the inhibition matrix (Figure 2), both drugs exhibit concentration-dependent cytotoxicity, with increasing levels of cellular inhibition observed as either drug concentration increases. Notably, the combination treatment leads to significantly higher inhibition compared to single-agent treatments at equivalent doses, suggesting a cooperative effect. At the highest tested concentrations, near-complete cell death (98%) was achieved, indicating potent combined activity. Dose-response inhibition matrix for digitoxin-cisplatin combination in MDA-MB-231 cells generated using SynergyFinder. The matrix shows % inhibition of cell viability across a range of drug concentrations (digitoxin 0-250 nM and cisplatin 0-50,000 nM). The color scale indicates increasing inhibition (white indicates ∼0% inhibition; darker red indicates progressively greater % inhibition).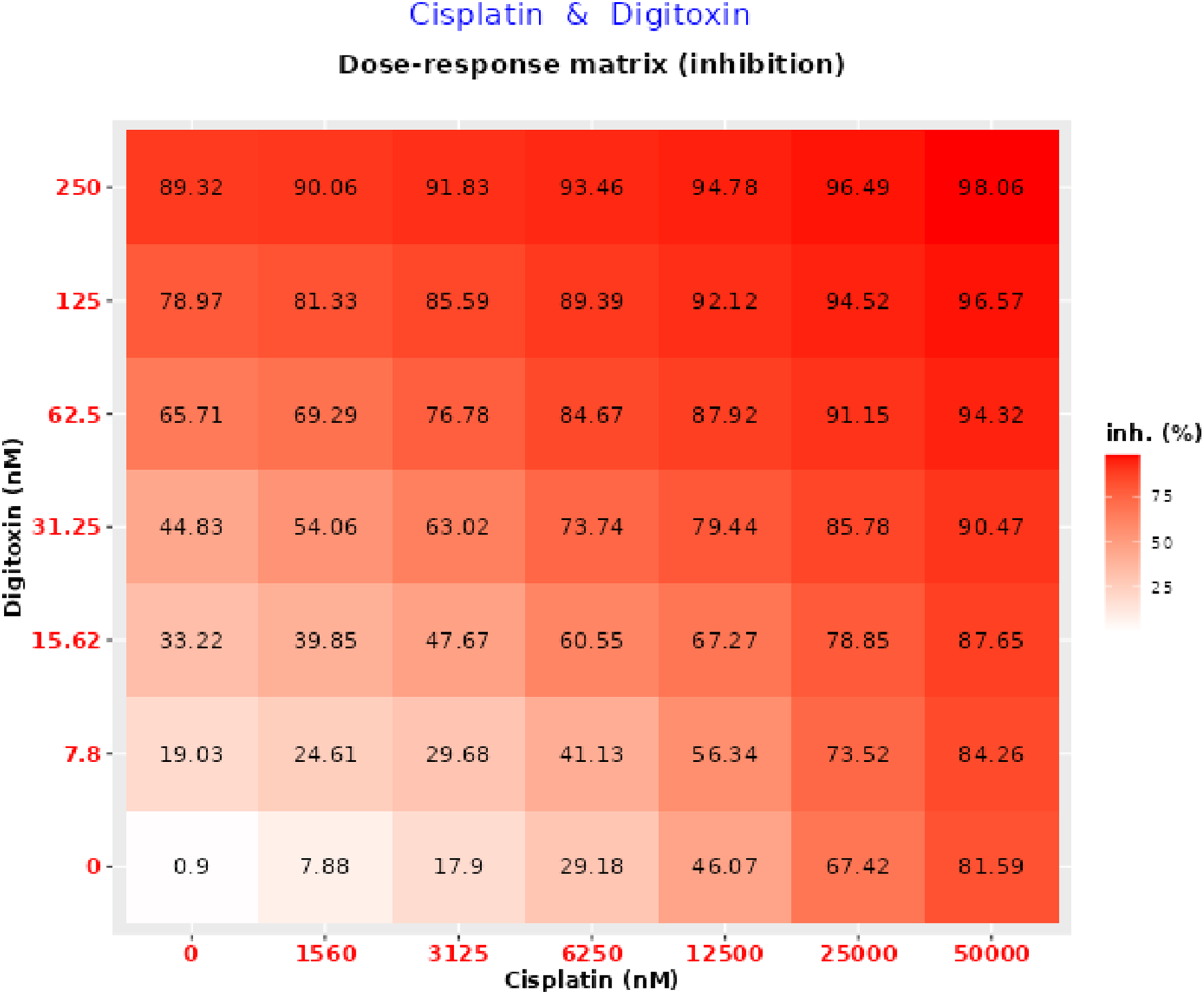
To quantify the nature and magnitude of this interaction, synergy analysis was performed using multiple reference models. The Bliss synergy map (Figure 3) reveals a broad region of strong positive synergy (deep red area), with an overall Bliss synergy score of 6.94 and a hotspot centered at 6.25 µM cisplatin and 31.25 nM digitoxin. The spatial distribution of red contours suggests a reproducible synergistic interaction across multiple concentration combinations. Bliss synergy contour map for digitoxin-cisplatin combination in MDA-MB-231 cells generated using SynergyFinder. The heatmap shows the Bliss synergy score (δ-units) at the indicated dose pair. The color scale represents the magnitude and direction of interaction (white corresponds to Bliss scores ≈0 (Bliss independence); red indicates positive Bliss scores (synergy); green indicates negative Bliss scores (antagonism)). The annotated hotspot indicates the MSA.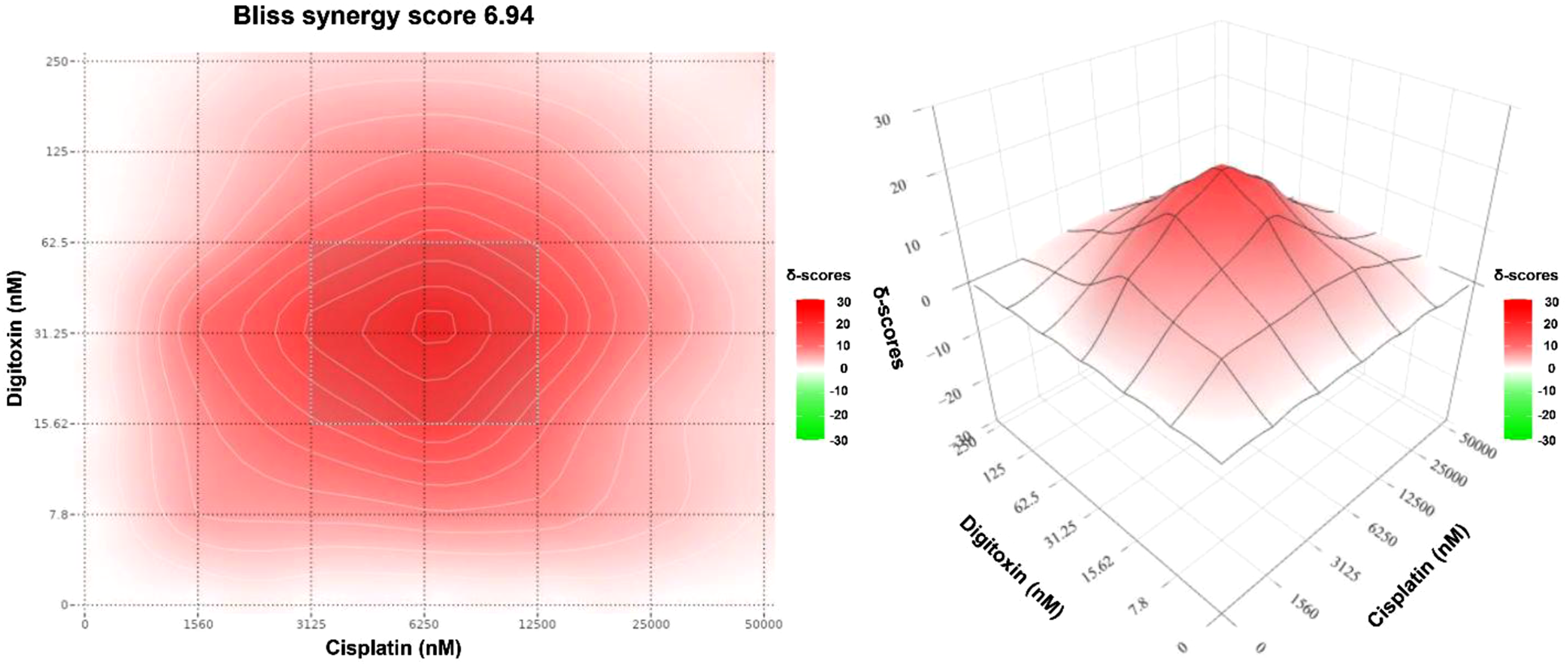
In contrast, the Loewe synergy map (Figure 4) demonstrates an even more pronounced synergistic profile, with an overall Loewe synergy score of 18.66 and a hotspot centered at 6.25 µM cisplatin and 125 nM digitoxin. However, a distinct green region at low doses of both drugs reflects antagonism, suggesting that at suboptimal doses the drugs may not cooperate efficiently and may engage compensatory survival responses. Loewe synergy contour map for digitoxin-cisplatin combination in MDA-MB-231 cells generated using SynergyFinder. The heatmap shows the Loewe synergy score (δ-units) at the indicated dose pair. The color scale represents the magnitude and direction of interaction (white corresponds to scores ≈0 (Loewe additivity); red indicates positive scores (synergy); green indicates negative scores (antagonism)). The annotated hotspot indicates the MSA.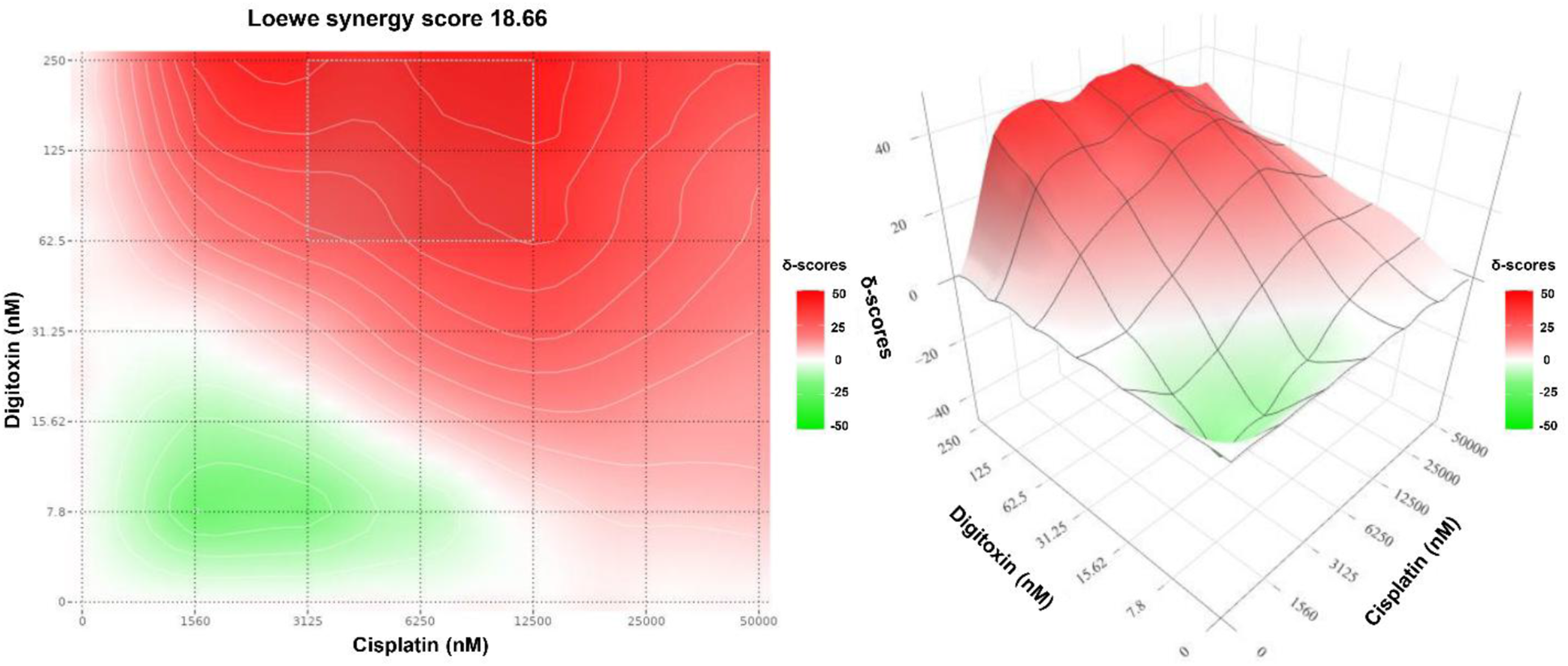
Complementing the Bliss and Loewe results, the ZIP model also indicated synergy (Figure 5). The ZIP surface showed predominantly positive synergy across the interior of the matrix, yielding a ZIP synergy score of 7.1, consistent with moderate synergy. The most positive region was centered near 6.25 µM cisplatin and 31.25 nM digitoxin, matching the hotspots identified by Bliss and Loewe. ZIP synergy contour map for digitoxin-cisplatin combination in MDA-MB-231 cells generated using SynergyFinder. The heatmap shows the ZIP synergy score (δ-units) at the indicated dose pair. The color scale represents the magnitude and direction of interaction (white corresponds to scores ≈0 (no change in potency); red indicates positive scores (synergy); green indicates negative scores (antagonism)). The annotated hotspot indicates the MSA.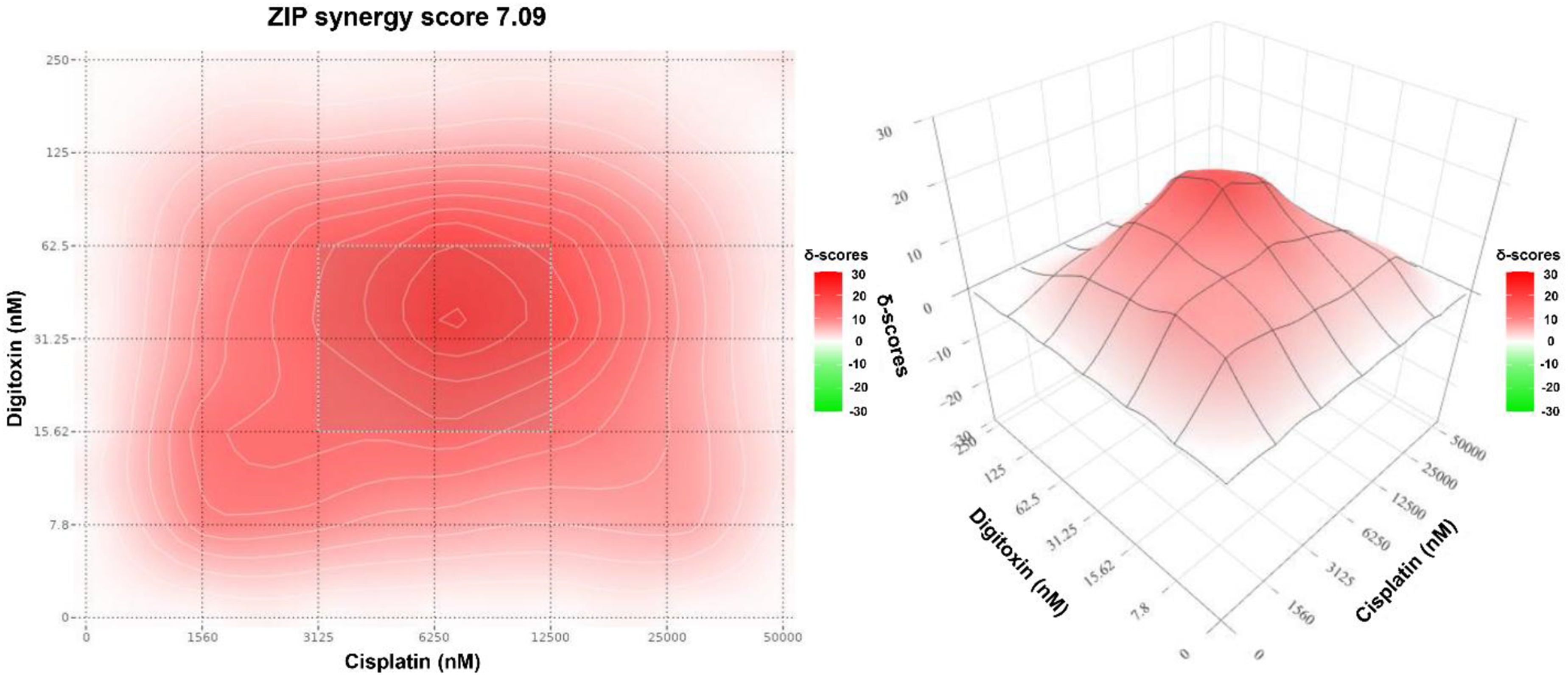
The Bliss-Loewe consensus score was evaluated (Figure 6), which provides a more conservative cross-assumption summary of interaction behavior across the dose matrix. The overall consensus synergy score was 3.05, indicating modest net synergy when averaged across the full matrix. Importantly, the consensus map still localizes the MSA to the same intermediate-dose band highlighted by Bliss, Loewe, and ZIP, supporting the robustness of the synergy hotspot across distinct models. Bliss-Loewe consensus synergy contour map for digitoxin-cisplatin combination in MDA-MB-231 cells generated using SynergyFinder. The heatmap shows the Bliss-Loewe consensus synergy score (δ-units) at the indicated dose pair. The color scale represents the magnitude and direction of interaction (white corresponds to scores ≈0 (consensus expectation met); red indicates positive scores (synergy); green indicates negative scores (antagonism)). The annotated hotspot indicates the MSA.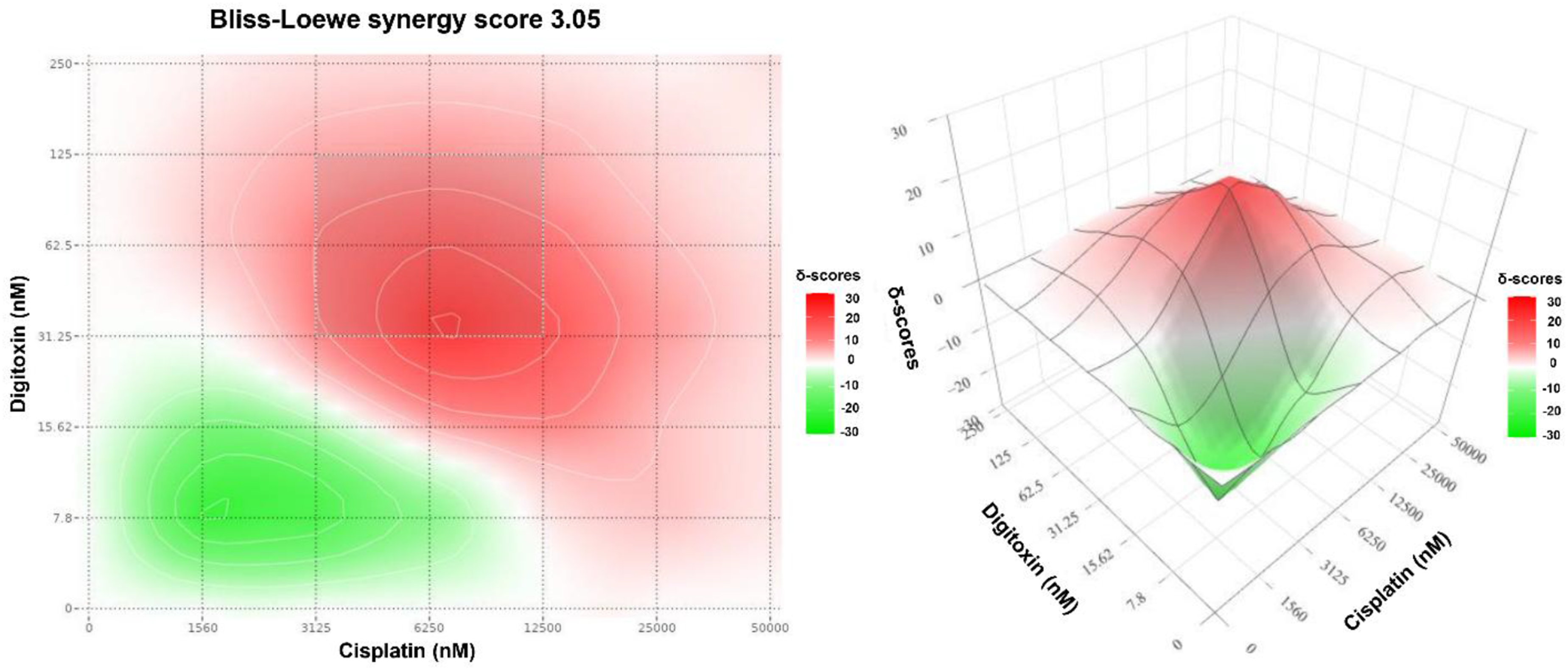
The HSA map showed moderate synergy for digitoxin-cisplatin with HSA score of 14. In the heatmap (Figure 7), the deepest red island indicates the MSA at 12.5 µM cisplatin and 31.25 nM digitoxin, where the combination outperforms the best monotherapy by 40 δ-units. Notably, the HSA model localized its MSA at 12.5 µM cisplatin, whereas Bliss, Loewe, and ZIP models identified a common hotspot at 6.25 µM cisplatin. This difference is expected because HSA measures improvement over the highest single-agent effect at matched doses, while the other models quantify deviations from model-specific additivity reference surfaces. HSA synergy contour map for digitoxin-cisplatin combination in MDA-MB-231 cells generated using SynergyFinder. The heatmap shows the HSA synergy score (δ-units) at the indicated dose pair. The color scale represents the magnitude and direction of interaction (white corresponds to scores ≈0 (combination effect equals the best single agent); red indicates positive scores (the combination outperforms the best monotherapy); green indicates negative scores (the combination performs worse than the best single agent)). The annotated hotspot indicates the MSA.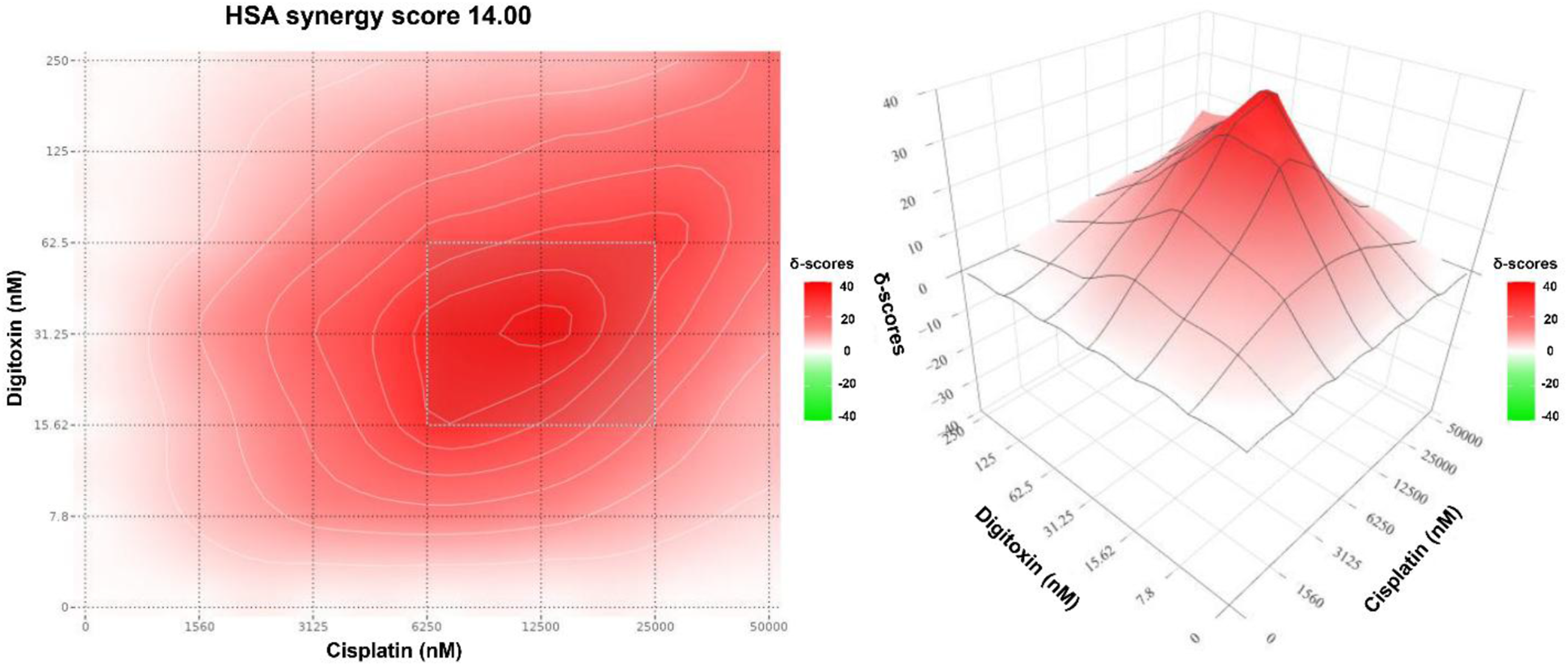
Cross-comparison of the four reference models therefore yields convergent and complementary information. Bliss, Loewe, ZIP and the Bliss-Loewe consensus all returned positive global scores and localized the strongest synergistic hotspot to the same intermediate-dose band (6.25 µM cisplatin and 31.25 nM digitoxin), despite their distinct null assumptions. The relative magnitudes of the scores reflect the underlying mathematics rather than any biological discrepancy: Loewe yields the largest values because dose-additivity is a stricter null than independence, while ZIP and Bliss return moderate scores reflective of effect level rather than dose-level deviations; the consensus is, by construction, the most conservative. The HSA map also confirmed synergy but shifted its peak to 12.5 µM cisplatin and 31.25 nM digitoxin, an expected consequence of HSA benchmarking the combination against the best single agent at matched doses rather than against a model-defined additivity surface. Given our dose-sparing aim, we prioritize the 6.25 µM cisplatin band, the consensus hotspot of the model-based methods, and the band that aligns with the strongest fixed-ratio synergy (1:200) and largest cisplatin DRI from the CI/DRI analysis, as the lead candidate dose-optimization window.
Discussion
The present work integrates fixed-ratio CI/DRI calculations with checkerboard-matrix analysis across four reference models to provide a multi-dimensional view of the digitoxin-cisplatin interaction in MDA-MB-231 cells. Our results showed that both frameworks converge on a lead candidate synergy window centered near 6.25 µM cisplatin and 31.25 nM digitoxin, with robust cisplatin dose-sparing.
Mechanistically, the synergy we observed integrates the rationale outlined in the Introduction with the topology of our dose matrix. Cisplatin’s therapeutic ceiling in MDA-MB-231 cells is set by efficient adduct repair, NF-κB–driven survival signaling, and dose-limiting toxicity31,32; precisely the nodes that digitoxin perturbs via Na+/K+-ATPase signalosome inhibition of NF-κB/AP-1 and NFAT-c-MYC axes,17–19 and through cardiac glycoside-class effects on DNA strand-break burden and repair efficiency. 21 The intermediate-dose hotspot we mapped (6.25 µM cisplatin and 31.25 nM digitoxin) is consistent with such a chemosensitization scenario, in which sub-cytotoxic digitoxin reshapes the survival and repair program of cells already burdened by platinum adducts, lowering the cisplatin dose required for a given lethal effect. Our DRI-based estimate of up to 27-fold cisplatin sparing extends, in a quantitative dose-matrix framework and in a TNBC context, prior cardiac glycoside-platinum observations that have been limited to digoxin and to non-TNBC models.23,24 The direction and magnitude of this interaction can nevertheless be context-dependent across cell line, schedule, and exposure window, 33 reinforcing the need for orthogonal validation in additional TNBC models stratified by BRCA1/2 and HRD status.
The deliberate use of four matrix-based reference models alongside the Chou-Talalay fixed-ratio framework also addresses a methodological concern that is increasingly emphasized in the synergy-quantification literature: any single model can produce false-positive or false-negative synergy calls depending on the geometry of the single-agent dose-response curves, on potency mismatch between partners, and on the effect level at which the score is integrated. The frameworks we deployed are not redundant; they probe different aspects of interaction. Across these conceptually independent methods, the results were concordant: all four reference models returned positive global scores, Bliss, Loewe, ZIP, and the consensus localized the MSA to the same intermediate-dose band (6.25 µM cisplatin and 31.25 nM digitoxin), and the matrix-derived hotspot aligned with the strongest fixed-ratio synergy (1:200) and the largest cisplatin DRI from the CI/DRI analysis. This convergence across frameworks with non-overlapping null assumptions constitutes stronger evidence of genuine pharmacological synergy than any single metric in isolation. The expected divergence of HSA’s peak toward 12.5 µM cisplatin is not a contradiction but a direct consequence of its single-agent benchmark and, in fact, reinforces the interpretation: at higher cisplatin exposure the combination still outperforms either monotherapy, even though the dose-additivity/independence-based criteria identify a more conservative low-dose synergy window better suited to dose-sparing. This methodological triangulation is also the reason we report the DRI in addition to CI and matrix scores: together they specify not only whether synergy exists but where in the dose space it is most useful for cisplatin dose-sparing.
From a translational perspective, three features of our results merit attention. First, the digitoxin concentration at the center of the synergy hotspot (31.25 nM) falls within the therapeutic plasma range routinely maintained in cardiac patients receiving digitoxin for heart failure (10-40 nM),34,35 indicating that clinically achievable digitoxin exposure is sufficient to potentiate cisplatin. Unlike many repurposed candidates that require supratherapeutic dosing, digitoxin’s anticancer-relevant concentrations overlap with its established cardiotonic window, substantially de-risking early clinical translation. Second, cisplatin’s principal dose-limiting toxicities are cumulative and dose-dependent,31,32 making any strategy that preserves efficacy at lower cisplatin doses directly relevant to patient safety. Our DRI-based estimate of up to 27-fold cisplatin sparing, if recapitulated in vivo, could substantially widen the therapeutic index by reducing cumulative platinum exposure while maintaining tumor control. Third, digitoxin is eliminated hepatically rather than renally, a pharmacokinetic property that is complementary to cisplatin-induced nephrotoxicity; co-administration would not be expected to compound renal injury or alter cisplatin clearance. 34 Taken together, these pharmacological features position the digitoxin-cisplatin combination as a rational candidate for dose-optimization studies in TNBC, particularly in patients for whom cisplatin toxicity constrains treatment duration.
Future work should extend these findings in three complementary directions. First, orthogonal functional validation should confirm that the intermediate-dose synergy window observed in short-term viability mapping translates to durable loss of reproductive capacity and defined cell-death modalities. Second, mechanistic profiling is needed to connect digitoxin-driven Na+/K+-ATPase signaling perturbations to downstream DNA-damage and replication-stress responses, particularly in the context of cisplatin-induced DNA crosslinks. Third, to strengthen clinical relevance in TNBC, subsequent studies should evaluate additional TNBC subtypes and incorporate combination benchmarks against contemporary standards of care.
This study quantified digitoxin-cisplatin interactions only in the MDA-MB-231 TNBC model. Given the molecular heterogeneity of TNBC and the enrichment of platinum sensitivity in BRCA-associated and broader homologous recombination-deficient (HRD) contexts, the magnitude and position of the synergy window may differ by BRCA1/2 and HRD status. Accordingly, an important next step is to profile the same digitoxin-cisplatin dose matrix across a TNBC panel stratified by BRCA/HRD to determine whether synergy is amplified in repair-deficient settings and to refine the dose window for translational relevance.
Finally, selectivity toward non-malignant cells was not assessed and should be addressed in follow-up work to better define the therapeutic window.
Conclusion
Digitoxin and cisplatin exhibit a potent synergistic interaction in MDA-MB-231 cells. Fixed-ratio CI/DRI analyses identified the strongest synergy at digitoxin:cisplatin ratio of 1:200 with substantial cisplatin dose-sparing, while checkerboard modeling localized a consensus intermediate-dose hotspot around 6.25 µM cisplatin and 31.25 nM digitoxin, where Bliss, Loewe, and ZIP were concordantly positive. HSA also supported benefit but shifted its maximal region toward 12.5 µM cisplatin consistent with its single-agent benchmark definition. These results provide a quantitative rationale to advance this combination into orthogonal functional validation, mechanistic studies linking Na+/K+-ATPase modulation to DNA-damage responses, and in vivo evaluation to define therapeutic index and safety.
Supplemental material
Supplemental material - Digitoxin as a novel synergistic partner of cisplatin in triple-negative breast cancer (TNBC) cells
Supplemental material for Digitoxin as a novel synergistic partner of cisplatin in triple-negative breast cancer (TNBC) cells by Sara A. Al-Shun, Magdy M. Youssef, and Farid A. Badria in Tumor Biology
Supplemental material
Supplemental material - Digitoxin as a novel synergistic partner of cisplatin in triple-negative breast cancer (TNBC) cells
Supplemental material for Digitoxin as a novel synergistic partner of cisplatin in triple-negative breast cancer (TNBC) cells by Sara A. Al-Shun, Magdy M. Youssef, and Farid A. Badria in Tumor Biology
Supplemental material
Supplemental material - Digitoxin as a novel synergistic partner of cisplatin in triple-negative breast cancer (TNBC) cells
Supplemental material for Digitoxin as a novel synergistic partner of cisplatin in triple-negative breast cancer (TNBC) cells by Sara A. Al-Shun, Magdy M. Youssef, and Farid A. Badria in Tumor Biology
Footnotes
Ethical considerations
Not applicable. This study did not involve human participants or experimental animals. MDA-MB-231 cells were obtained from Nawah Scientific (Cairo, Egypt) and confirmed mycoplasma-free prior to experiments.
Author contributions
All authors contributed to study conception and design. SAA performed all experiments, data analysis and interpretation, prepared all figures, and drafted the manuscript. MMY and FAB supervised the work, critically revised the scientific content, and contributed to manuscript writing. All authors read and approved the final manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Science, Technology and Innovation Funding Authority (STDF) within the German Egyptian Research Fund (GERF), Project ID 33601. The grant funded research activities only; article processing charges (APCs) are not an eligible cost and no funds are available for publication fees. No additional institutional open-access support is available.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data generated or analyzed during this study are included in this article. Additional materials are available from the corresponding authors upon reasonable request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.