
Abstract
We describe a model-based strategy for defining an optimal lighting schedule for strengthening the circadian rhythms of patients in intensive care units as a means of reducing delirium rates in hospitals. The effect of different lighting schedules on the amplitude of the circadian rhythms was assessed via computer simulations using a model of the circadian system. The simulation results show a large boost of the amplitude of circadian rhythms when the photoperiod is within the 6 h–16 h duration range at relatively modest light levels. It is concluded that a fast recovery of endogenous circadian rhythm can be supported by daily light pulses of about 6 h without necessarily causing visual discomfort to subjects.
1. Introduction
In recent years, weak circadian rhythms of melatonin have been reported to be associated with increased delirium rates in intensive care units (ICUs). Given that delirious patients tend to stay hospitalised for longer periods of time and have a worse prognosis, reducing the odds of delirium is a matter of great importance for public health. A plausible strategy for reducing delirium rates is the strengthening of circadian rhythms by creating appropriate lighting conditions in patient rooms.
Although the definition of lighting conditions for achieving fast re-establishment of strong circadian rhythms in critical care patients is best pursued experimentally, the large solution space of this problem makes this approach impractical without some form of a guiding strategy. Mathematical models of the circadian system can be instrumental in defining a potential strategy for accomplishing this goal.
2. Objective
The primary aim of this paper is to describe a model-based strategy for defining an optimal lighting schedule of strengthening the circadian rhythms of patients in intensive care units as a means of reducing delirium rates in hospitals. The lighting schedule resulting from the study is recommended for future research addressing the role of light and circadian rhythms in creating healing environments in hospitals.
3. Background and significance
As a way of conveying the basis and relevance of reducing delirium with lighting, the following paragraphs introduce basic concepts and present a review of the scientific literature describing the putative effects of lighting on the odds of ICU delirium.
3.1 Delirium
Delirium is an etiologically nonspecific organic cerebral syndrome characterised by concurrent disturbances of consciousness and attention, perception, thinking, memory, psychomotor behaviour, emotion, and the sleep-wake schedule. 1 It is a state of confusion whose intensity may rapidly fluctuate over time.
The exact cause of delirium is currently unknown and it is likely to be multiple. It is a quite prevalent condition in ICUs with rates normally reported to be two digits high in different studies. Ely et al. 2 estimated that 60–80% of the patients in an ICU develop delirium while Maldonado 3 reported an 18% incidence in an acute ICU (medical and surgical patients combined).
Whatever the cause of delirium, it represents a burden for patients who report terrifying hallucinations and for the hospital staff who have to cope with uncooperative and irrational behaviour. Furthermore, patients who develop delirium are more likely to stay longer in the hospital and die earlier in comparison to patients who do not develop the condition. 4 Surviving patients who experienced delirium are also at a greater risk of developing long-term cognitive impairments. 5 All in all, delirium is a serious problem that deserves attention due to its high prevalence and negative associations.
3.2 Lighting effects
Despite its prevalence and potential negative consequences there is currently no specific treatment for delirium, and interventions are addressed mainly to control the symptoms once the condition is manifested. In this context, prevention can be regarded as the most important strategy for addressing ICU delirium.
Effective prevention requires the identification of causes or at least precipitating factors of delirium. Among these factors there is a body of evidence suggesting that lighting conditions affect the odds of delirium in the ICU population.
Wilson, in 1972, reported a difference in delirium rates between hospitals patients in rooms with and without windows, i.e. subject to distinct illumination conditions. 6 This trend was also observed and reported by Keep et al. in a similar study. 7 Furthermore, delirium rates have been observed to display seasonality with higher rates in the winter months, when the photoperiod is shorter, and lower rates during the summer, when daylight is plentiful. 8
While these studies were observational, Taguchi et al. conducted an interventional experiment with a control group to assess the impact of light on delirium rates in which the morning bright light of 5000 lux was applied to postoperative ICU patients. 9 The results show a trend for reducing delirium in the bright light group in comparison to the control group. Finally, a recent prospective cohort study conducted by Rompaey et al. 10 concluded that patients in rooms with visible daylight have lower chances of developing delirium than patients deprived of it (odds ratio 2.39; confidence interval 1.28–4.45).
3.3 Potential mediation of the circadian system
The apparent link between lighting and delirium in the ICU suggests a potential causal relationship which could be leveraged to reduce delirium rates in acute care settings. Assuming this causality holds, knowledge of the mechanism of action is instrumental in designing an optimal lighting system aimed at preventing delirium. A possible mechanism exists in the way lighting influences the amplitude of circadian rhythms, such as plasma levels of melatonin secreted by the pineal gland. 11
Plasma melatonin levels follow a circadian rhythm in which very low levels are observed during the day and high levels at night. The difference between the maximum and minimum levels defines the amplitude of the melatonin circadian rhythm. In post-operative ICU patients, such rhythm tends to be abnormal in the first day after admission but typically normalises by the end of the second day. 12 – 14 Nonetheless, ICU patients with low melatonin amplitude lasting for three or more days have been reported to be at greater risk of developing delirium. 15 The correlation between the blunted circadian rhythm of melatonin and delirium rates provides a hint that melatonin itself or some of its regulatory agents, e.g. lighting, could be precipitating factors of delirium.
Lighting is a primary regulator of plasma melatonin levels throughout the day. This influence is mediated by a cluster of neurons in the brain referred to as the suprachiasmatic nucleus (SCN). 16 The neurons in the SCN fire at varying frequencies, keeping an approximate 24 h cycle which is coupled to the light–dark cycle via lighting information from the retina. The dynamics of lighting has the effect of promoting a strong alignment of the firing rhythms of the SCN cells, causing periods of fast firing to happen in close synchrony. When the SCN cells are in close synchrony, the circadian rhythm of melatonin is well defined and displays a large amplitude.17,18 Consequently, variations in the ambient lighting conditions can indirectly impact the amplitude of the circadian rhythm of melatonin. The effect of light on the amplitude of melatonin has in fact been observed in several experiments: light during the day is able to increase melatonin amplitude at night 19 ; and a lower amplitude of the melatonin circadian rhythm can be observed in subjects exposed to days with dimmer or shorter photoperiods.19,20 Besides delayed effects, light induces a strong and immediate suppression of melatonin in humans.21,22
The circadian system comprises a multiplicity of circadian oscillators and pacemakers of which the SCN is responsible for overall synchronisation via electrical or humoral (e.g. melatonin) pathways. 23 This broad spectrum of action suggests that the impact of lighting on delirium, if any, could be mediated by changes in the amplitude of circadian rhythms in the body.
Our working hypothesis on delirium can thus be summarised as follows: Lighting affects delirium rates and circadian rhythms as observed in plasma melatonin levels; a low amplitude in the circadian rhythm of melatonin is associated with higher rates of delirium in intensive care units; hence delirium can be prevented by using lighting to strengthen circadian rhythms in the body, of which melatonin is a reliable marker. Note that this hypothesis does not necessary imply that melatonin is in the causal pathway of delirium. It does, however, imply that lighting conditions suitable for strengthening the circadian rhythm of melatonin are also suited to have a positive impact on delirium rates, no matter what underlying process is in place.
A plausible strategy for reducing delirium rates is thus the strengthening of circadian rhythms by creating appropriate lighting conditions in patient rooms. The use of mathematical models to create suitable lighting strategies in ICUs is the main object of this document.
4. Document organisation
The rest of this paper is organised as follows: Section 5 describes a mathematical model of the circadian system that accepts lighting of time-varying intensities as input and outputs levels of plasma melatonin concentration over time. Section 6 presents and discusses simulation results using the aforementioned model to address the problem of finding a lighting schedule that maximises the amplitude of melatonin circadian rhythm of subjects with blunted rhythms. Section 7 further evaluates three candidate schedules for ICU lighting. Section 8 presents conclusions of the study.
5. Circadian system model
The literature of circadian physiology is rich in conceptual and quantitative models of the circadian system aimed at describing biological rhythms. 24 – 28 Conceptually, at the core of all these models lies a pacemaker, which is influenced by the environment via receptors and influences physiological processes via effectors.
In this section we describe a mathematical model of the circadian system for exploring the impact of different lighting schedules on resynchronising circadian rhythms. Although the model integrates concepts from different studies, it differs from previous models by its focus on the lighting control of melatonin levels as required by the objective of our study. The existing model to date with closest focus was proposed by St Hilaire et al. 29 to simulate the effects of ocular light and the circadian system on plasma melatonin. Although the model describes the impact of lighting on the circadian dynamics of melatonin, it is not explicit in how these dynamics are endogenously generated by the circadian pacemaker. The value of our proposed model is in bridging this gap.
5.1 Model description
The circadian system model accepts lighting at time-varying intensities as input and outputs levels of plasma melatonin concentration over time. The model comprises three components as depicted in Figure 1. The photic receptors translate the amount of lighting reaching the retina into an excitatory stimulus to the circadian system. The master pacemaker generates oscillations whose amplitude and phase are regulated by the stimuli received from the photic receptors. Such oscillations then drive the production of melatonin by the pineal gland. The remainder of this section describes in general terms these three model components. A detailed presentation of the model can be found in Appendix A1.
Schematic view of the circadian system model. The model, comprising three components, accepts as input lighting (L) at time-varying intensities and outputs levels of plasma melatonin concentration (C) over time. Photic receptors define the number (Agate) of gate cells which are active (i.e. firing) at any point in time; the percentage of firing neurons (Pout) at a certain instant defines the concentration of plasma melatonin in the body (C)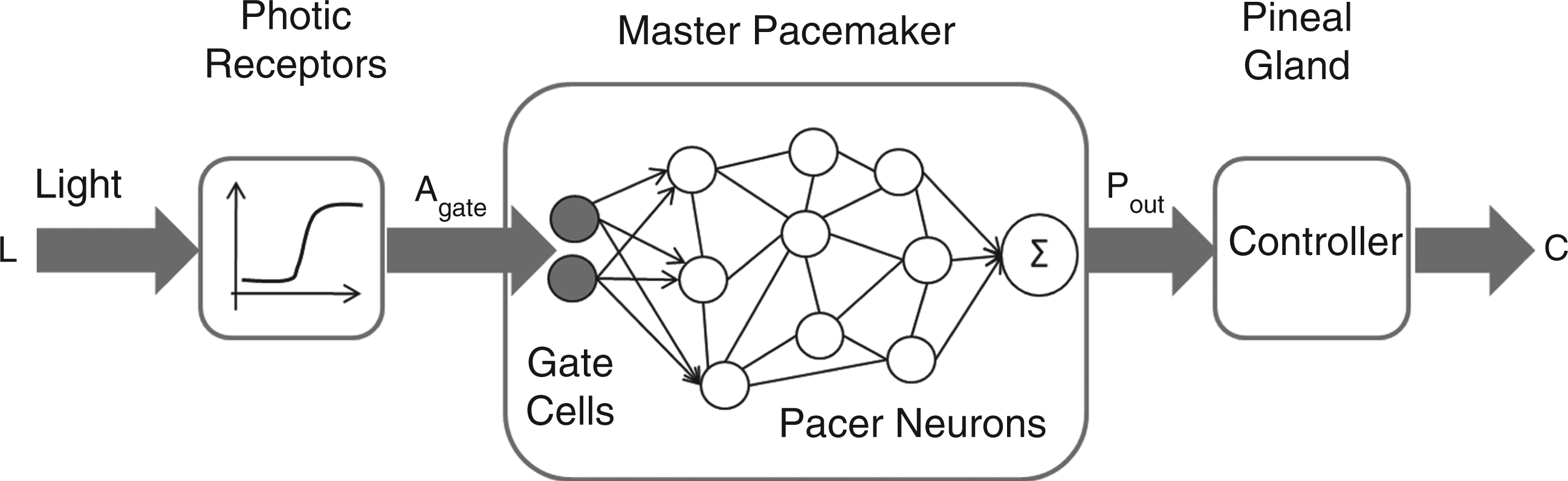
5.1.1 Photic receptors
The impact of light intensity on the pacemaker is modelled as a function that translates light intensity in lux reaching the eye to a number of gate neurons excited in the SCN. A logistic function is used to describe the excitatory phenomena.
A logistic-like behaviour is naturally expected in describing the activation of gate cells in the SCN since a minimum amount of light is needed to excite these cells. Furthermore, there must be a point beyond which most or all gate cells are already excited and more light cannot provide large changes.
The model assumes the light intensity level L as coming from a uniform cool-white light source of about 4100 K as a proxy for light exposure. This assumption reflects the type of light source used in previous experiments whose data were used for model calibration (cf. Section 5.2). The circadian effects of lighting are likely to vary with its spectrum30,31 or spatial distribution. 32 The contribution of these factors was not counted in the model because the quantification of their impact is still in an early stage of development. In particular, interactions between different spectral components affect circadian response in ways that just recently have been appreciated,33,34 and consequently it is currently premature to adopt models of such effects. 35
5.1.2 Master pacemaker
The master pacemaker model comprises two groups of neuronal cells 36 : Gates and oscillators. Gate neurons are excited in the presence of light stimulus mediated by the photic reception function. In the absence of light, most of these cells do not fire. Conversely, oscillator cells display autonomic oscillation and fire with a circadian period of approximately 24 h. Each oscillator cell fires for a number of hours each day and remains quiescent for the rest of the time.
Coupling between the SCN cells contributes to the generation of synchronised firing patterns by continuously adjusting the drift between pacers with different frequencies. The output of the master pacemaker is a signal Pout that represents an adjusted percentage of firing neurons at a certain instant in time. Thus, if the oscillator cells are tightly synchronised, Pout will be close to 1 during parts of the day and close to 0 in other parts. In other words, the master pacemaker will display a high amplitude (peak value). Conversely, a pacemaker with desynchronised pacers will display low amplitude.
Although a very weak or lacking circadian rhythm happens when the SCN individual neurons fire out of phase, this phenomenon can only be temporary: pacer neurons are coupled and will eventually resynchronise by themselves, even in the absence of light. The strength of this coupling is given by parameter K1, described in Appendix A1. Light is thus not generating the circadian rhythm in the model, only modulating the process: the activation of gate neurons by light influences the speed with which the oscillator cells align their firing, the strength of alignment (via parameter K2, see Appendix A1), as well as the final alignment phase.
The model encompasses photoperiodism by making the duration of the firing pattern of an oscillator vary with the amount of light exposure. Thus during winter, when days are short, the firing duration of pacers will tend to be larger than during summer.
5.1.3 Pineal gland
The third component of the circadian system model is the pineal gland which controls the release of melatonin in the body. It is assumed that melatonin is metabolised at a fast enough rate for its plasma levels to depend primarily on the production by the pineal gland. This assumption is corroborated by evidence that humans given melatonin orally display a biphasic elimination pattern with half-lives of about only 3 and 45 min, respectively. 37
In the model, it is the firing of SCN neurons that suppresses melatonin. The more the firing of neurons are in phase alignment, the longer is the period in which little or no firing occurs and melatonin production is high. As the firing alignment is primarily determined by lighting stimuli, this explains why appropriate lighting during the day can increase melatonin production during the night. 19
5.2 Model calibration and validation
The circadian system model described in Section 5.1 contains a set of parameters whose values need to be specified in a manner that deems it capable of representing real world phenomena. A calibration process was undertaken to choose adequate values by varying these parameters until the model was able to closely reproduce data from real-world experiments involving humans.
Parameter values were randomly selected from a space of possible candidates and used in the simulation of experiments assessing the phase response curve of melatonin rhythm to one pulse of light,
38
to intermittent light
39
and to varying illuminance levels.
17
Experiments assessing melatonin suppression at varying illuminance levels
17
and how changes in daylight affect the duration of nocturnal melatonin secretion
20
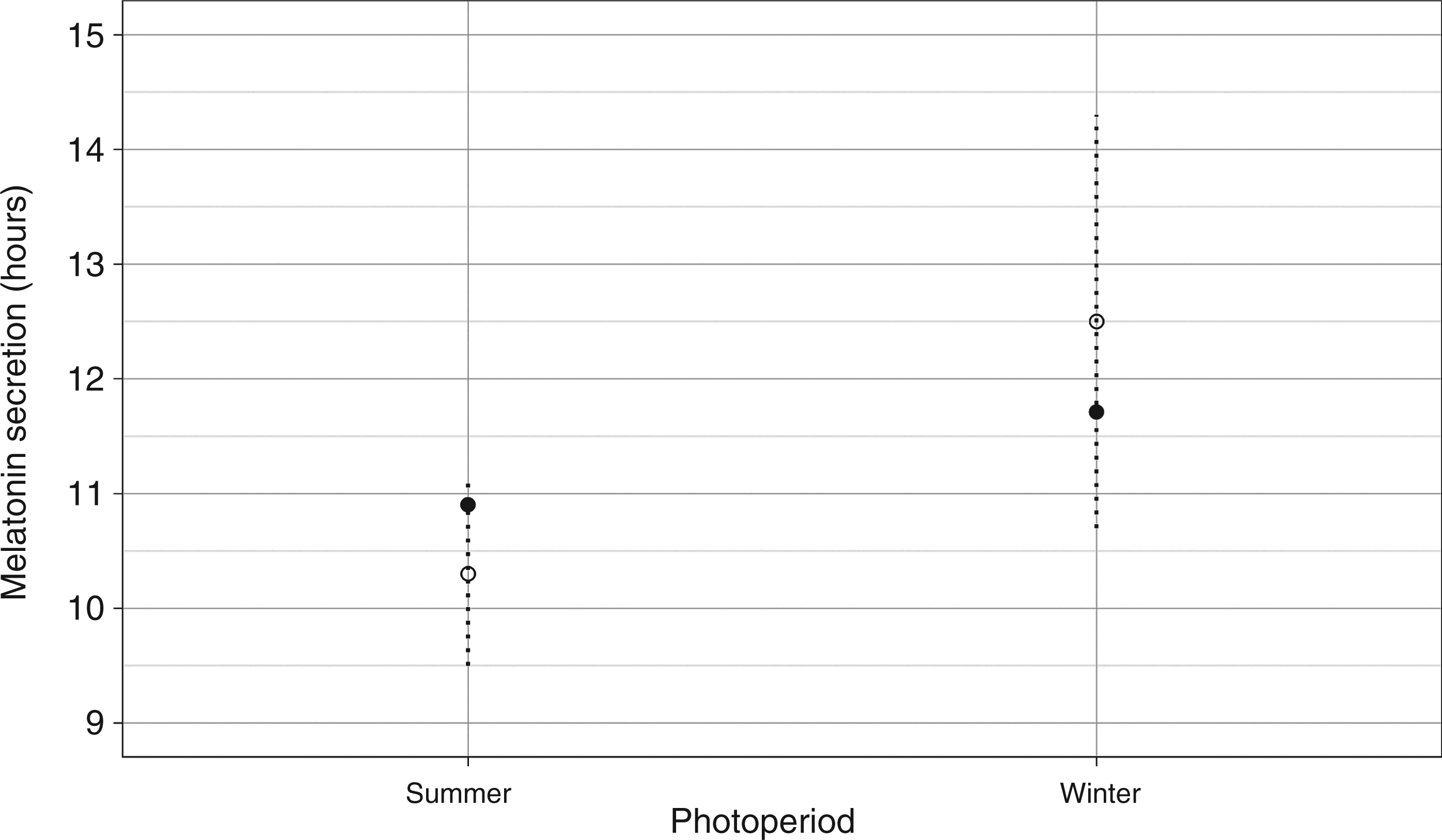
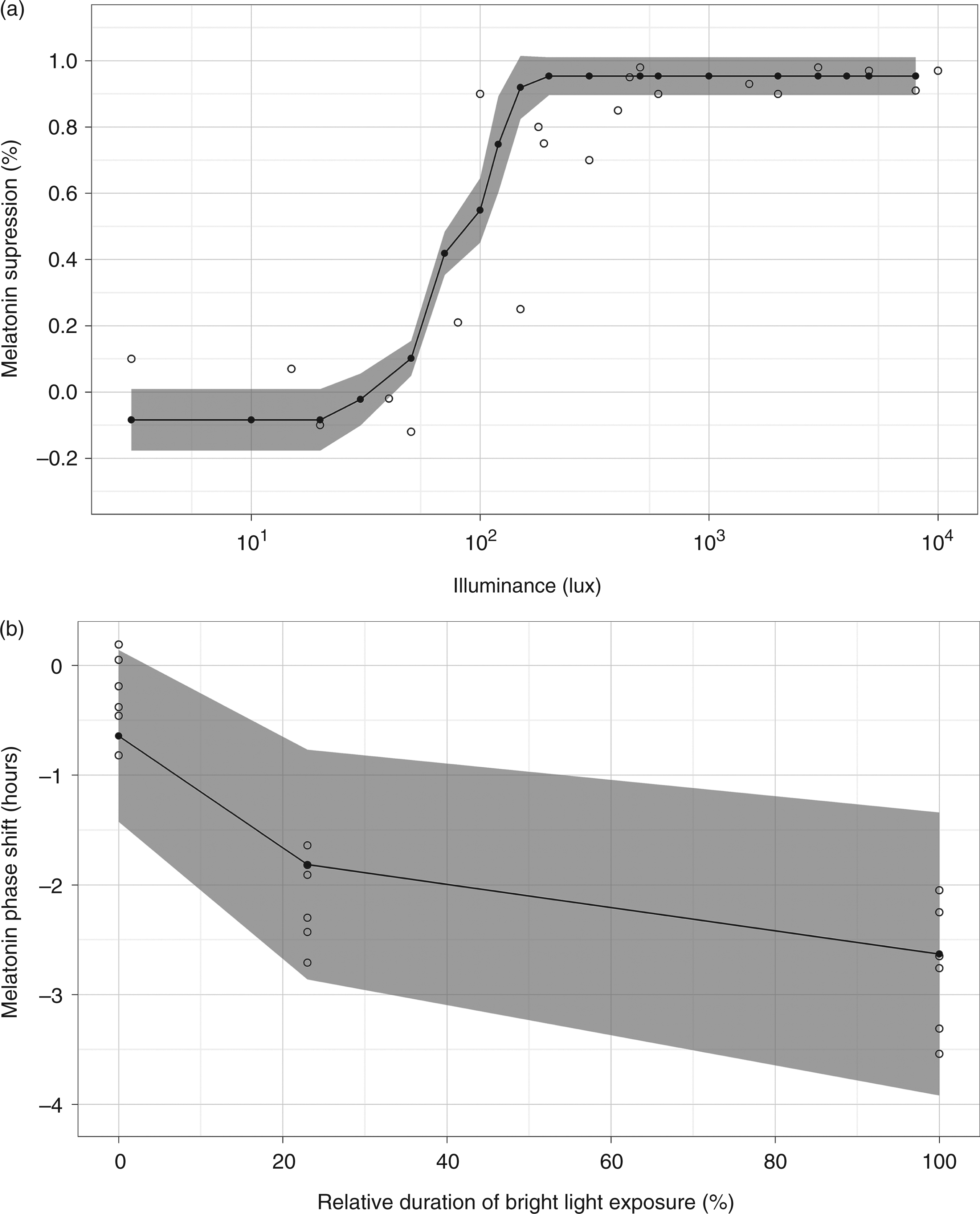
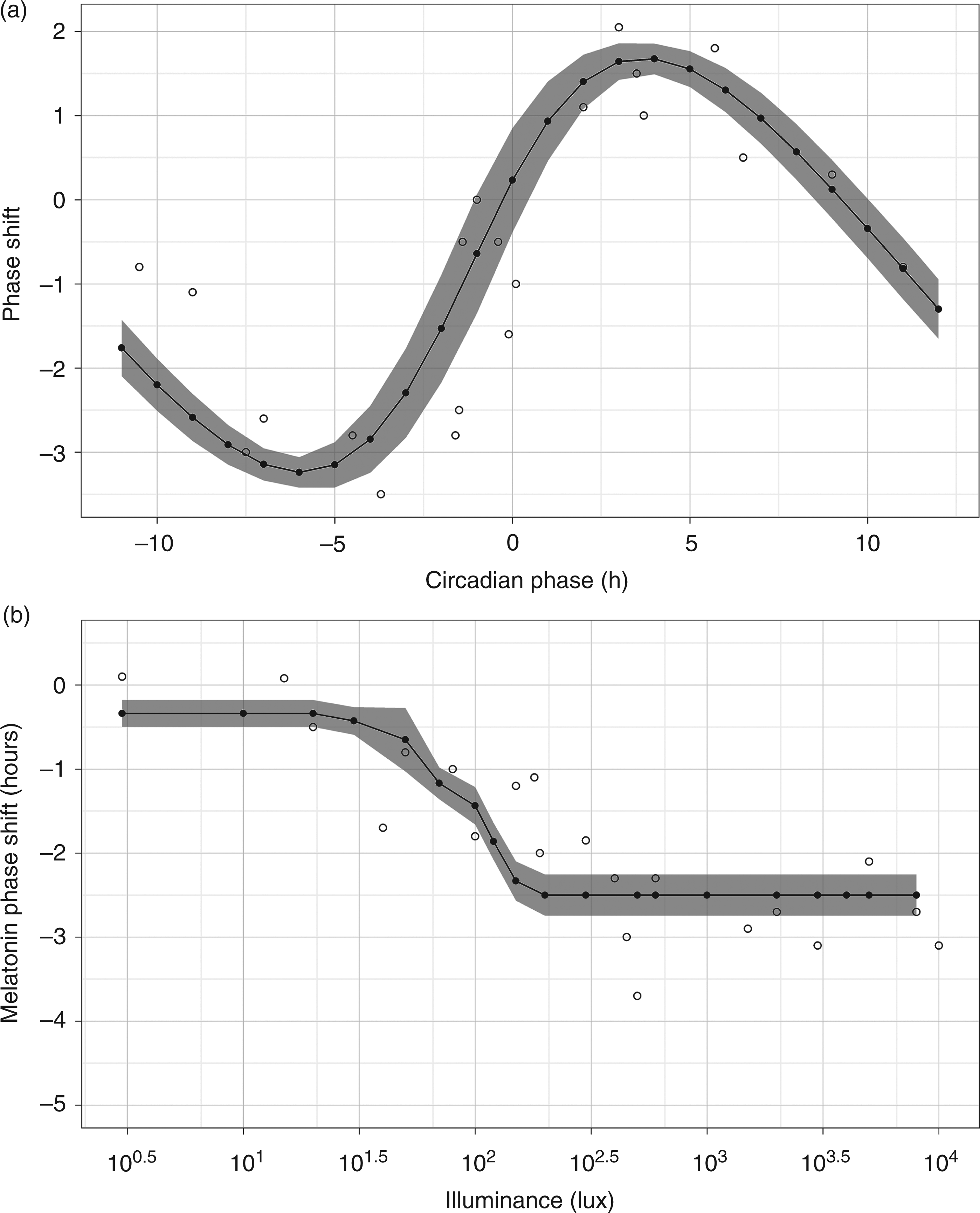
were also simulated. The mean squared error of the curves generated in each simulation was used to compare among candidate parameter sets. The set of parameters with smallest errors was used as the calibrated values of the validated model. A list of model parameters and calibrated values can be found in Appendix A2. Figures 2, 3 and 4 display comparisons of simulated and experimental results.
Comparison of simulated and experimental results. Nocturnal melatonin secretion duration as a function of a long (summer) and short (winter) photoperiod. Experimental data from Wehr.
20
The open (○) circle represents experimental data and the filled (•) circle represents simulated data. A simulated data point is the average of 50 simulations. The dotted line in the graph represents the 99% confidence interval of the experimental data Comparison of simulated and experimental results. The open (○) circle represents experimental data and the filled (•) circle represents simulated data. A simulated data point is the average of 50 simulations. The shaded areas cover two standard deviations of simulated data. (a) Suppression response curve to one light pulse using changes in the area under the curve of melatonin as a suppression marker. Experimental data from Zeitzer et al.
22
(b) Phase responses to very dim light (0%), intermittent bright light (23%) and bright light (100%). Experimental data from Gronfier et al.
39
Comparison of simulated and experimental results. The open (○) circle represents experimental data and the filled (•) circle represents simulated data. A simulated data point is the average of 50 simulations. The shaded areas cover two standard deviations of simulated data. (a) Phase response curve to one light pulse stimulus using melatonin midpoints as the circadian phase marker. Experimental data from Khalsa et al.
38
(b) Phase response curve to one light pulse at different illuminance levels using melatonin midpoints as the circadian phase marker. Experimental data from Zeitzer et al.
22
6. Boosting the amplitude of blunted circadian rhythms
Given a subject with complete absence of circadian rhythms, our aim is to define a lighting schedule that maximises the amplitude of melatonin circadian rhythm by the end of the third day (Figure 5). In this section we use the circadian system model outlined in Section 5.1 to find a solution to this problem and discuss potential adaptations to the schedule when applying it to a hospital healing environment.
Boosting circadian rhythm amplitude. The top graph shows the output Pout of the circadian pacemaker in terms of percentage of active neurons at a given point in time. The bottom graph represents the corresponding melatonin concentration over time. The amplitude of melatonin in the third day is marked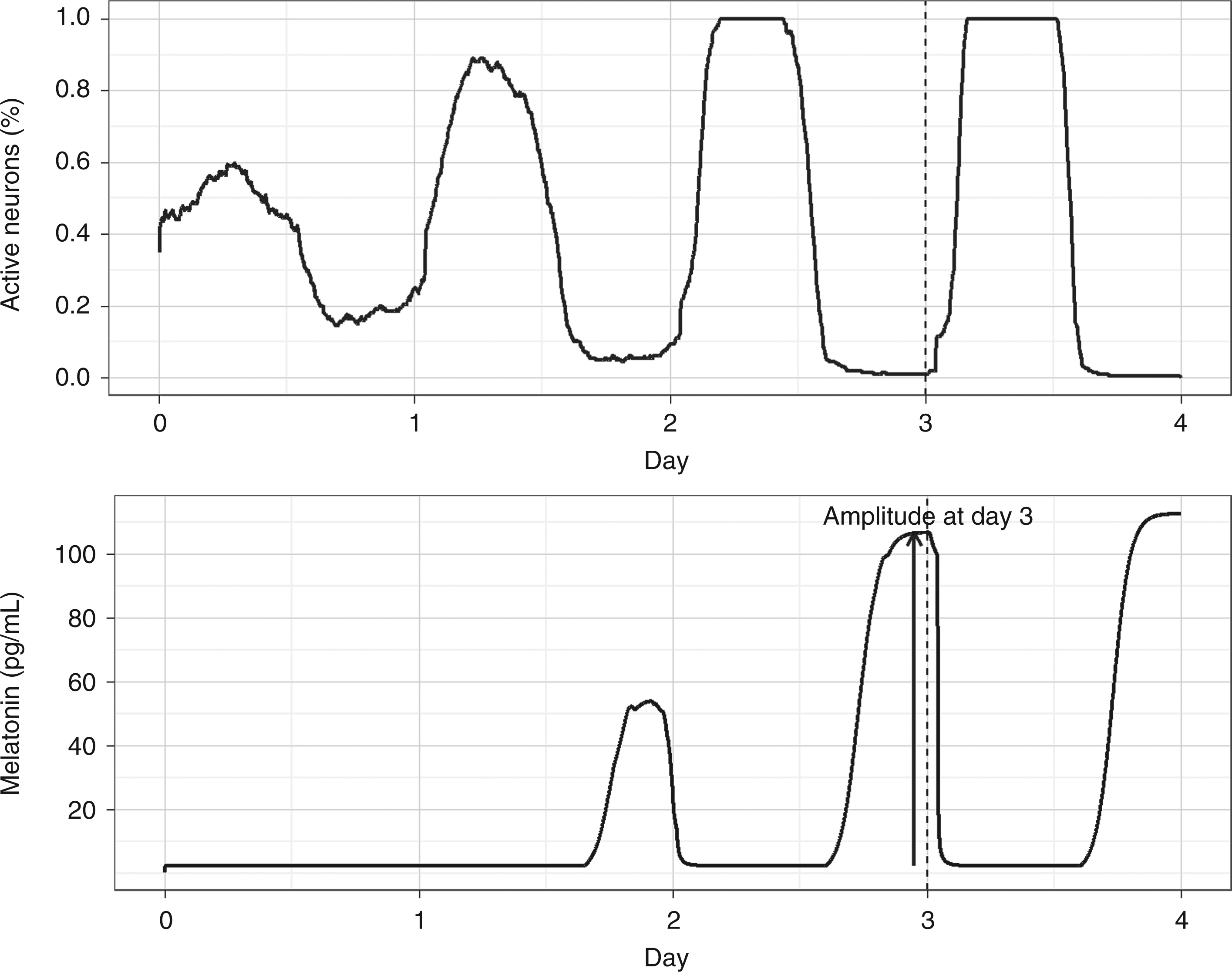
6.1 Methods
The effects of different lighting schedules on the amplitude of circadian rhythms were assessed via computer simulations. At the start of the simulation, the pacemaker of each simulated subject was fully desynchronised, meaning that the firing phase of the oscillator neurons in the pacemaker is uniformly distributed, and consequently no circadian rhythm is apparent. Melatonin levels were initially basal. The amplitude of the circadian rhythm of melatonin was defined in terms of its 24 h period peak value. Further information on the values of parameters used in the simulations can be found in Appendix A2.
6.1.1 Lighting schedules
Each simulated day was divided into 24 one-hour time intervals and within each hour the light could be on or off. In order to maintain the solution space of the problem small, lighting schedules were constrained in terms of light levels and time distribution. In general, a single schedule displayed only two light levels at the subject's eyes (e.g. 0/100 lux; 0/500 lux; 0/1,000 lux, etc.) and each simulated day repeated exactly the same schedule. Figure 6 shows the lighting schedule pattern used in the study.
Lighting strategy patterns. Schedules are divided into one hour slots. Unshaded blocks represent light pulses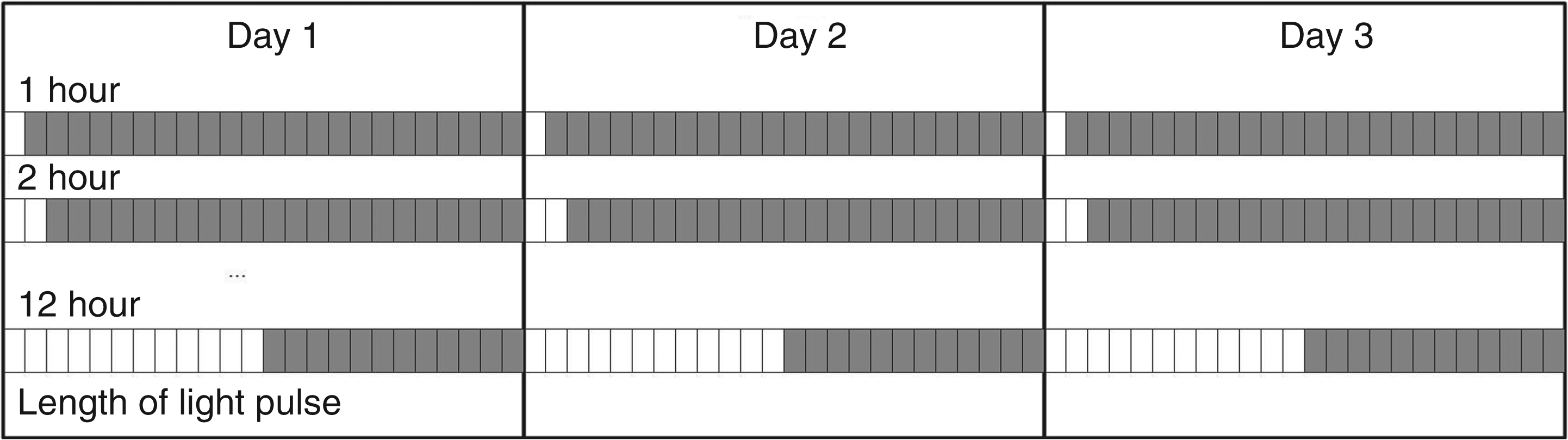
Deviations of the bi-level lighting schedule patterns were allowed to take into consideration the effects of light sources that may not be controllable in real-world scenarios. In the forthcoming discussion we thus refer to two types of lighting sources: therapy lighting, which we can in principle adjust at will; and ambient lighting, which represents ‘noise' in the lighting schedule.
A second type of noise, besides ambient lighting, was represented by the time a subject remained with eyes closed (e.g. napping) during the therapy photoperiod. This noise was considered by introducing random periods of darkness in the lighting schedule to emulate the effects of closed eyes. A patient confined to a bed for several days is likely to close his or her eyes more often than a healthy subject under normal routine. The random periods were used as a first approximation to investigate the impact of napping behaviour on the results of the lighting intervention.
6.2 Results
6.2.1 Effect of light pulse duration and intensity
Applying a light schedule of the form represented in Figure 6 with different pulse durations and intensities yields the graph shown in Figure 7. Irrespective of the light intensity it can be seen from the graph that the highest final amplitudes of melatonin are reached for pulses of about 12 h duration, which more or less equally divide the day into a light and a darkness period.
Effect of pulse duration and intensity on final melatonin amplitude. Melatonin amplitude after three days as a function of the duration of the photoperiod at different light intensities. Each day the start of light pulses coincided with time zero. Bars indicate standard deviations. The largest amplitudes resulted from photoperiods in the duration range from 6 to 16 hours. Pulses with light intensity beyond 200 lux provided only marginal increases in the final melatonin amplitude. A simulated data point is the average of 50 simulated subjects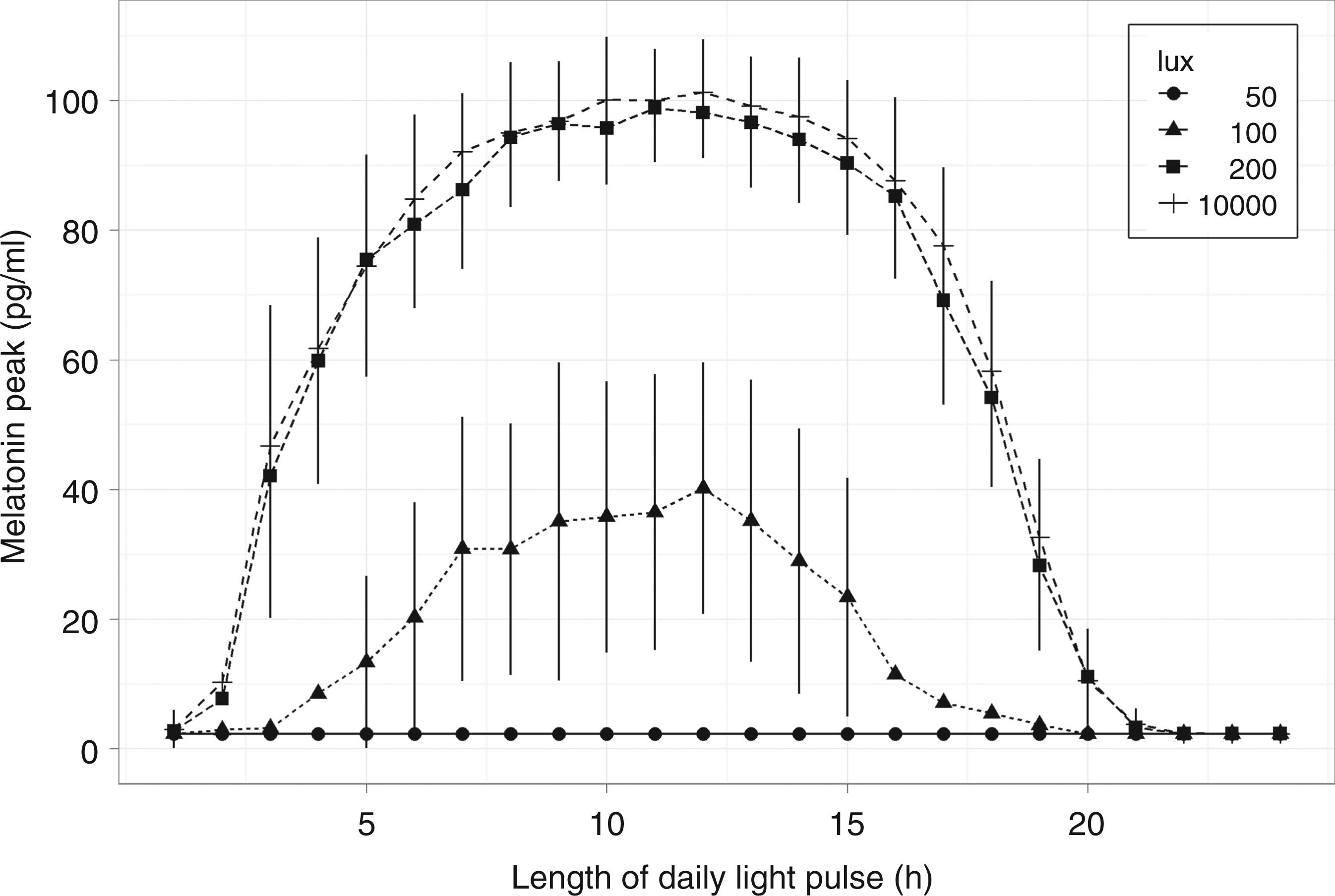
The graph also clearly indicates that light pulses above 200 lux do not significantly increase the final melatonin amplitude. Considering these effects together, the results suggest that 6–16 h pulses of at least 200 lux are able to produce near maximum amplitude boost of blunted circadian rhythms.
6.2.2 Effect of closed eyes and ambient lighting
The graph in Figure 8 shows the effect of a subject having eyes closed on the final melatonin amplitude for two distinct lighting schedules involving a 12 h continuous pulse of 200 lux therapy lighting. The closed-eye probability in a given hour slot is assumed independent from that in all other intervals. When eyes are closed, the circadian system is assumed to be free of photic stimulation. Intuitively in this situation, the curves in the graph must present a negative slope, reflecting the fact that amplitude boost is weaker when light stimuli becomes scarce. The graph also indicates a difference in the boost achieved by the two lighting schedules which differ only by the presence of ambient lighting. Having an additional source of light superimposed on the therapy light but lasting longer (18 h), even at a relatively low level (100 lux), may induce significant reductions in the boosting effect.
Effect of closed eyes and ambient lighting. Melatonin amplitude after three days as a function of closed eye probability in any given hour. Therapy lighting intervention consists of a continuous pulse of 12 hours duration at 200 lux delivered at the beginning of each day for three days. The more a subject remains with eyes closed during therapy the lower the final melatonin peak. The graph also depicts the effect of having an additional source of light superimposed on the therapy light but lasting longer (18 h). Even at low levels, such additional light induces significant reductions in the final melatonin amplitude. A simulated data point is the average of 50 simulated subjects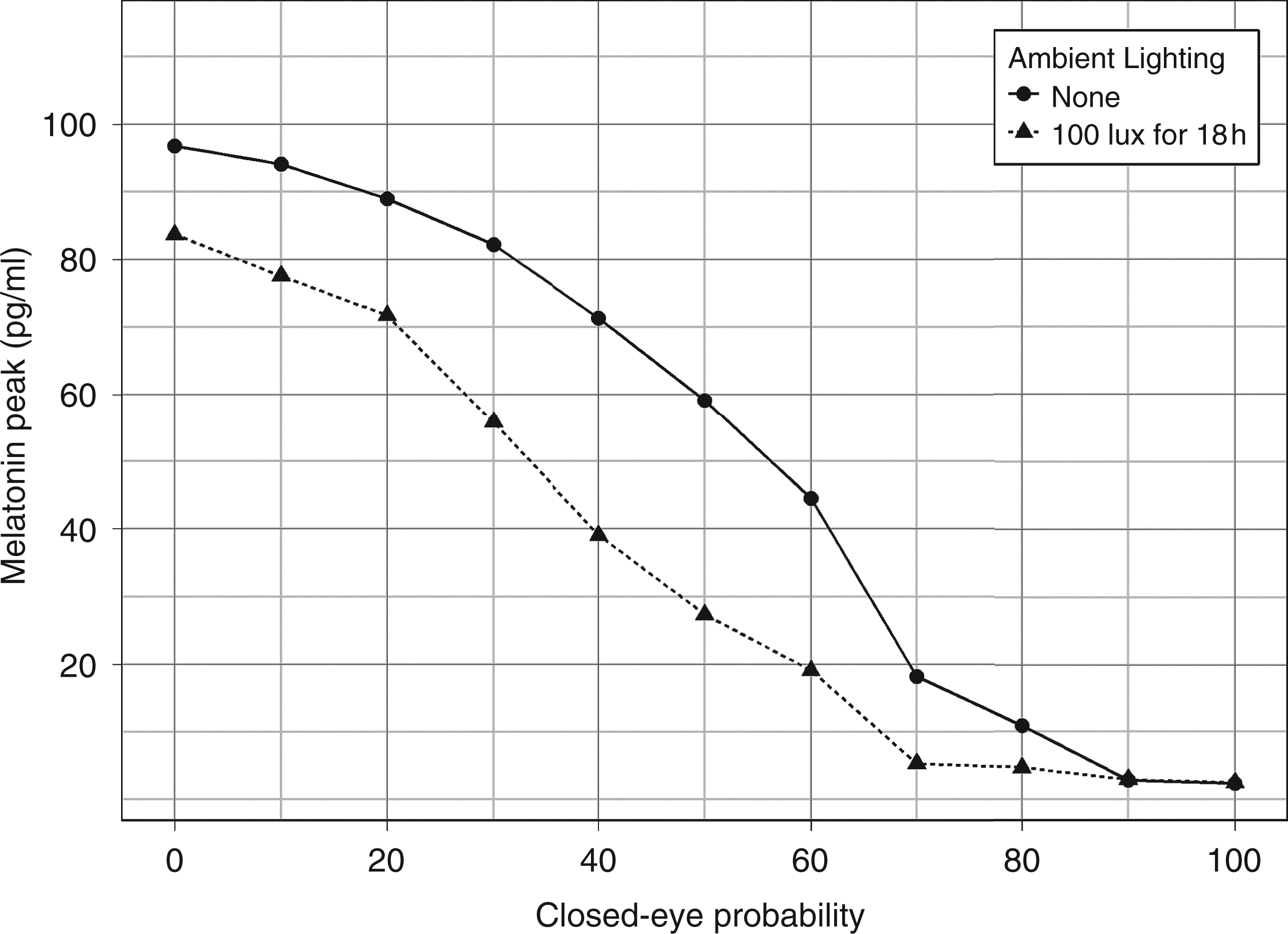
6.2.3 Effect of light compensation
A compensation light therapy strategy extends the photoperiod of light therapy exposure beyond its initial value to compensate for time intervals in which the subject maintained his or her eyes closed. During the extended light session, subjects must remain with their eyes open. For example, if light therapy is set to 6 h and during this period the patient remains for 2 h with his or her eyes closed, compensation requires the light therapy to be prolonged by an extra 2 h. Consequently, the patient will be exposed to 8 h of bright light instead of the original planned 6 h.
The three curves in Figure 9 were produced with 16 h of ambient lighting of 100 lux superimposed on therapy lighting of 12 h without compensation, 6 h without compensation and 6 h with compensation. A 6 h therapy lighting with compensation induces high levels of final melatonin amplitude independent of closed-eye probability during initial light exposure.
Effect of light compensation. Melatonin amplitude after three days as a function of closed eye probability in any given hour. A compensation light therapy strategy extends the photoperiod beyond its initial value to ensure the subject has his or her eyes open for the total duration intended. During the extended light session subjects remain with eyes open. The three curves were produced with the therapy light is superimposed on 16 h of ambient lighting of 100 lux. A simulated data point is the average of 50 simulated subjects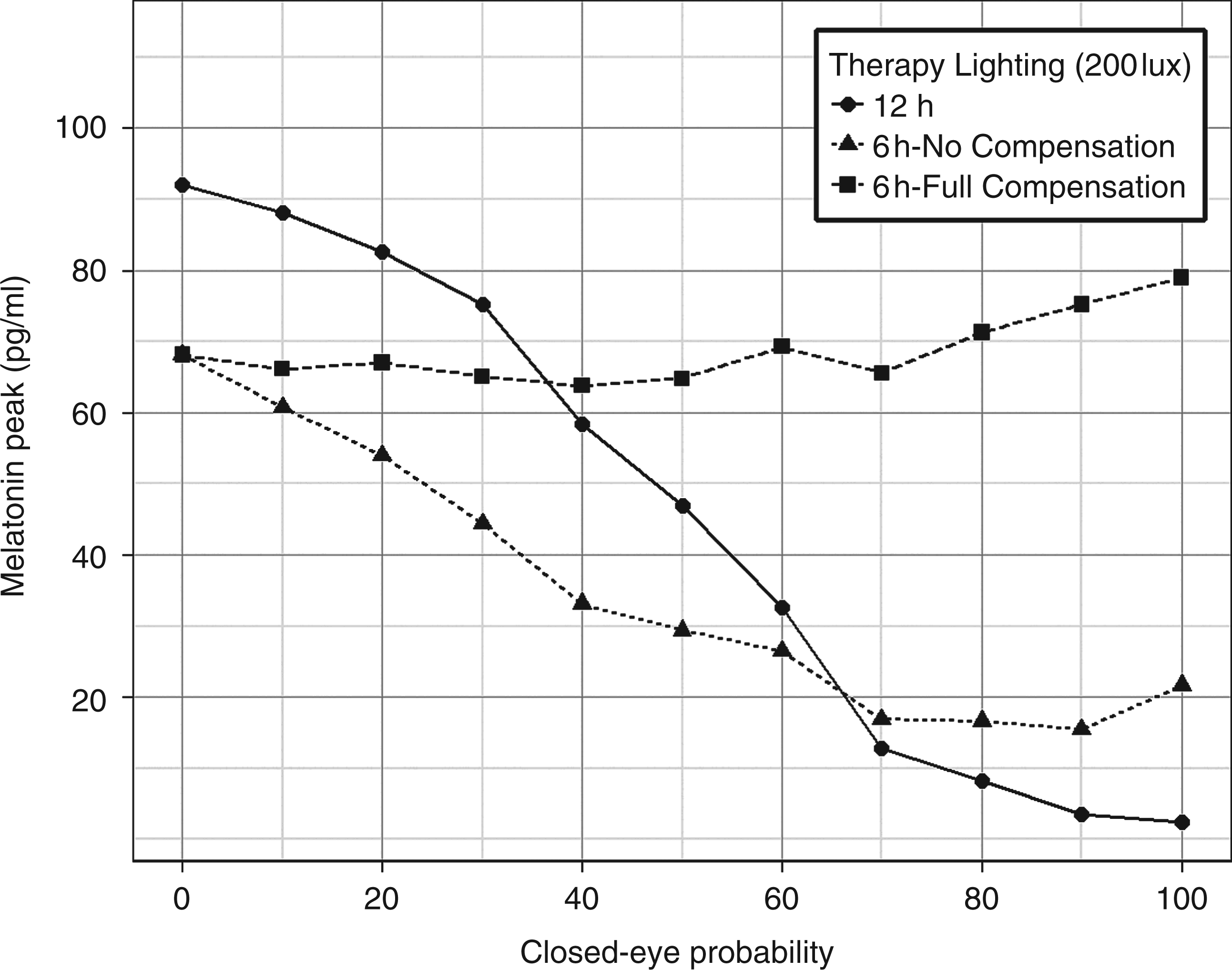
6.3 Discussion
The simulation results show a large boost of the amplitude of circadian rhythms when the photoperiod is within the 6–16 h range. Differences in the final amplitude for values within this range are relatively narrow (less than 20%) indicating that longer than 6 h exposure to daily light does not necessarily provide significant benefits (Figure 7). Interestingly, the lower cutoff value of such a photoperiod range coincides with the minimum amount of daylight available throughout the year for regions of the Earth between parallels 60 degrees north and 60 degrees south where almost the entire world population resides. This close match could be a result of optimal evolutionary adaptation. 40
Regarding the intensity of the lighting stimulus, relatively modest light levels are able to produce rhythms with amplitudes close to those generated with much brighter levels. Therefore, a fast recovery of endogenous circadian rhythm can be supported by daily light pulses of about 6 h without necessarily causing visual discomfort to subjects.
These results were, however, rather sensitive to two factors: light noise during periods of darkness and closed eyes during the photoperiod. When darkness periods were disturbed by the presence of light, even at low levels, the final circadian amplitude was greatly reduced. This observation reinforces the notion that darkness is as important as light in the regulation of the circadian system. As for the second factor, frail subjects in hospitals cannot be expected to remain awake or with eyes opened for many consecutive hours. In the simple model assigning independent equal probability of having a subject with eyes closed during each one-hour interval, the final circadian amplitude for photoperiods of 6 h was shown to be negatively impacted. A potential solution to balance the requirements of the subject’s comfort and maximum impact on circadian rhythm amplitude is to use variable photoperiods that are extended based on the amount of time subjects remain with their eyes closed.
Although we assumed for the sake of simplicity that eyelids are able to completely block the light, in reality a certain amount is always transmitted to the retina. The transmittance of light through human eyelids has been observed to be up to 15% at 700 nm and 3% below 600 nm in measurements conducted in vivo.41,42 However, the effective impact of this residual light on the circadian system is likely to be very low, if any at all.43,44
7. Simulation of candidate lighting schedules
We used the lessons of the simulation study to generate and further evaluate three candidate schedules for ICU lighting. The schedule in Figure 10(a) exhibits a bright light pulse of approximately 6 h duration starting in the morning. The schedule in Figure 10(b) is a slight variation of the previous schedule with a dip in the light intensity level around noon. Finally, Figure 10(c) presents a schedule that provides only about 3 h of bright light during the morning and is used for comparison purposes. All schedules provide 8 h of darkness and at least 300 lux throughout the day.
Lighting Schedules for the ICU. (a) 6 hour continuous bright light; (b) 6 hour bright light with noon dip. (c) 3 h continuous bright light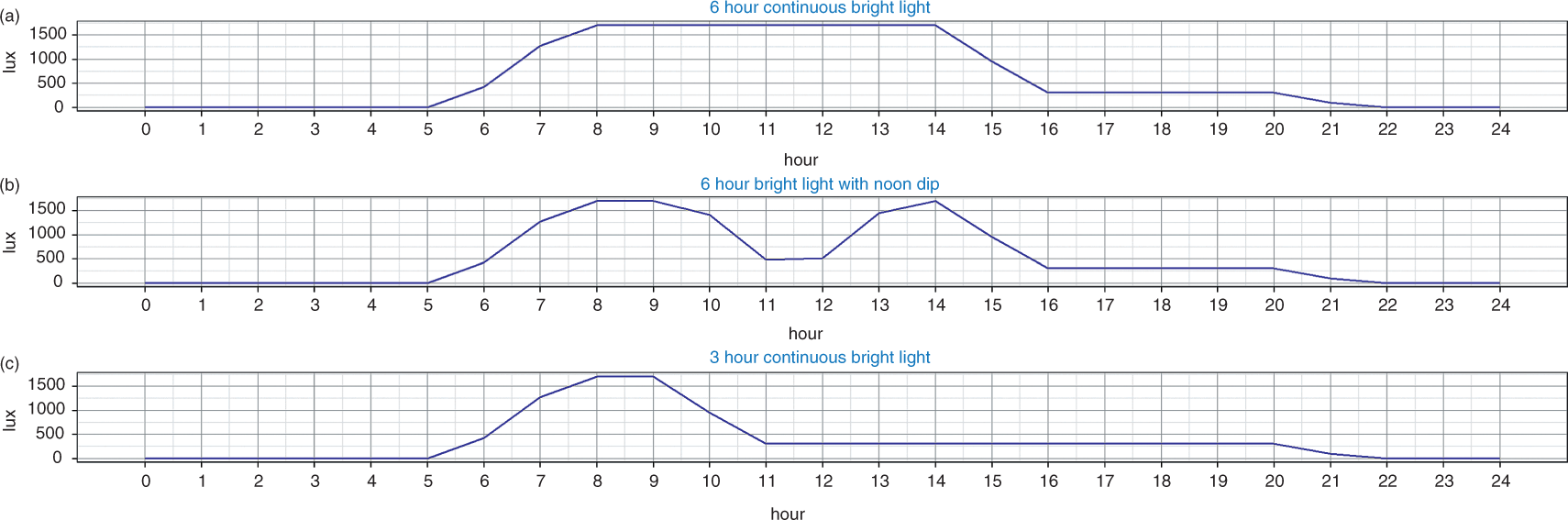
The effectiveness of these schedules in restarting the circadian rhythm of melatonin was evaluated through simulations using the model described in Section 5. Lighting levels in these schedules are assumed to be measured close to the patient head in the horizontal plane. It is further assumed that the light level reaching the patient’s retina is only 20% of the level indicated in the schedule due to shadowing of eyebrows, etc.
The results are depicted in Figure 11 as a function of the closed-eye probability. Using a value of 50 pg/mL or about 50% of the maximum amplitude as the minimum acceptable level after three days of lighting therapy, it can be seen that both schedules with 6 h bright light are effective in restarting the circadian rhythm even when patients remain with eyes closed about one-third of the day. On the other hand, the 3 h bright light schedule requires patients to maintain open eyes during almost all day in order to reach the 50 pg/mL threshold.
Melatonin amplitude after three days as a function of closed eye probability in any given hour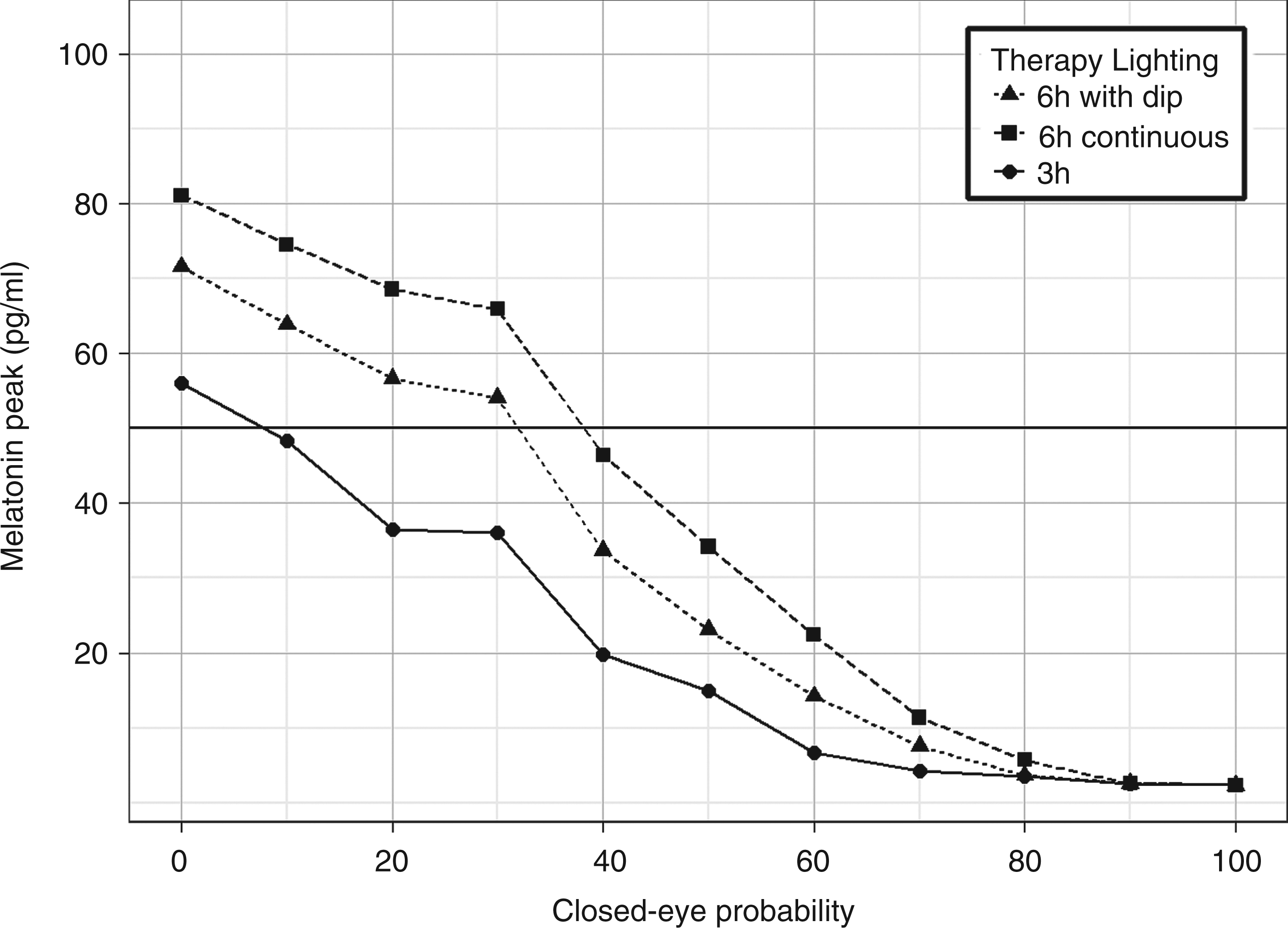
8. Conclusions
In the final analysis, a practical setting for lighting conditions to boost the amplitude of circadian rhythms of subjects would consist of ensuring 6 h of bright lighting from the early morning, followed by normal lighting conditions (e.g. room artificial lighting or sunlight if present) for no more than 10 h, and complemented by a period of 8 h of darkness. This setting should be able to balance patient comfort and sleep with the needs of hospital staff. It should also be able to provide a large amplitude boost for subjects closing their eyes for up to one-third of the time during the bright light exposure, even without resorting to a compensation strategy. Schedules compliant with this strategy are recommended for future research addressing the role of light and circadian rhythms in enabling healing environments in hospitals.
As a final note, we would like to reflect on the limitations of this study which may affect the applicability of the results in ICU environments and beyond. The circadian system model proposed is a simplification of the complex architecture of the circadian time keeping in humans, which involves a network of peripheral oscillators in the body. 45 Critically ill patients may develop systemic asynchronous rhythms which even a strong SCN drive would have trouble normalising. Medications and environmental noise light intensity may fragment sleep making it harder for the resynchronisation process to succeed. Furthermore, the proposed model was calibrated with data collected from healthy individuals and it is not clear how well these represent the circadian dynamics of ill patients. In view of these limitations, it is important to stress that the results of this study should be seen as hypotheses to be validated in clinical experiments.
Footnotes
Acknowledgments
The authors would like to thank Thomas Falck, Els Møst, Verus Pronk and anonymous reviewers for their valuable suggestions to improve this manuscript.
Funding
This research was funded by Royal Philips Electronics and received no additional specific grant from any funding agency in the public, commercial, or not-for-profit sectors.