
Abstract
Significance:
Hydrogen sulfide (H2S), a vital gasotransmitter in liver physiology and pathology, is produced by cystathionine γ-lyase (CSE), cystathionine β-synthase (CBS), and 3-mercaptopyruvate sulfurtransferase (MPST). Although CSE and CBS have been extensively studied in liver diseases, the role of MPST has received less attention, despite its significant and complex regulatory functions in hepatic biology and metabolism.
Recent Advances:
The involvement of MPST in antioxidant processes, mitochondrial function, and protein persulfidation highlights its multifaceted role in various liver diseases, where it can act either as a protective factor or as a contributor to disease progression, primarily through H2S metabolism. Notably, MPST also participates in crosstalk between various organs, including adipose–liver, gut–liver, skeletal muscle–liver, and pancreas–liver interactions. We also summarize current genetic models and pharmacological modulators targeting MPST. This review highlights the importance of advancing our understanding of the MPST/H2S pathway to develop promising therapeutic strategies for liver diseases.
Critical Issues:
Current research focuses mainly on changes in the expression of MPST in liver diseases, but its underlying mechanisms require further elucidation. The available genetic and pharmacological tools are still limited, underscoring the need for specific modulators and tissue- or cell-specific conditional MPST knockout or knock-in models to advance future investigations.
Future Directions:
Future research should focus on delineating stage-specific and cell type-dependent mechanisms of MPST in liver diseases, identifying upstream regulators, and elucidating detailed downstream molecular pathways mediating the effects of MPST. Ultimately, these efforts should aim to establish the clinical significance of MPST as a translational target. Antioxid. Redox Signal. 00, 000–000.
Introduction
Hydrogen sulfide (H2S), a gaseous transmitter, plays important roles in modulating physiological and pathological processes within mammalian tissues. It is produced by enzymes such as cystathionine γ-lyase (CSE), cystathionine β-synthase (CBS), and 3-mercaptopyruvate sulfurtransferase (MPST) (Wang, 2012). H2S exerts its biological functions through diverse interactions with proteins, DNA, and reactive species, including reactive oxygen species (ROS) and reactive nitrogen species (Wang, 2012). The liver has a remarkable capacity for H2S metabolism (Norris et al., 2011), which influences glucose homeostasis (Zhang et al., 2013), insulin sensitivity (Mani et al., 2013; Yusuf et al., 2005), lipoprotein synthesis (Hwang et al., 2013), and mitochondrial bioenergetics and biogenesis (Lagoutte et al., 2010; Módis et al., 2013). Accumulating evidence has linked H2S dysregulation to various liver diseases (Mani et al., 2014; Nguyen et al., 2024).
With respect to H2S-generating enzymes, previous research has focused primarily on the roles of CBS and CSE in liver diseases. CBS and CSE provide the liver with a high trans-sulfuration capacity (Sarna et al., 2015) and are essential for maintaining hepatic redox homeostasis, mitigating oxidative stress, and regulating lipid metabolism (Mani et al., 2014). The loss of CBS induces severe hyperhomocysteinemia, which leads to liver injury (Robert et al., 2005). Furthermore, CBS and CSE have been identified as potential therapeutic targets for conditions such as ischemia/reperfusion-induced liver injury, metabolic dysfunction-associated steatotic liver disease (MASLD), liver fibrosis, and liver cancer (Ascenção and Szabo, 2022; Sarna et al., 2015; Sun et al., 2021). However, the role of MPST in liver diseases is frequently overlooked.
MPST has unique biological characteristics that distinguish it from CBS and CSE. While CBS and CSE are predominantly cytosolic enzymes (Fiorucci et al., 2006), MPST is localized in both the cytosol and mitochondria (Kuo et al., 1983), enabling it to perform compartment-specific functions, such as transfer tRNA (tRNA) thiolation (Noma et al., 2009) and the maintenance of mitochondrial electron flow (Módis et al., 2013). Additionally, CBS and CSE are pyridoxal-5′-phosphate-dependent enzymes that use cysteine and/or homocysteine as substrates to generate H2S (Cirino et al., 2023). In contrast, MPST catalyzes H2S production with 3-mercaptopyruvate (3-MP) as its substrate (Shibuya et al., 2009). MPST also participates in cyanide detoxification (Williams et al., 2003), further highlighting its functional divergence from CBS and CSE. Emerging evidence highlights the significant regulatory roles of MPST in liver pathophysiology. This review emphasizes the contributions of MPST across various hepatic disorders.
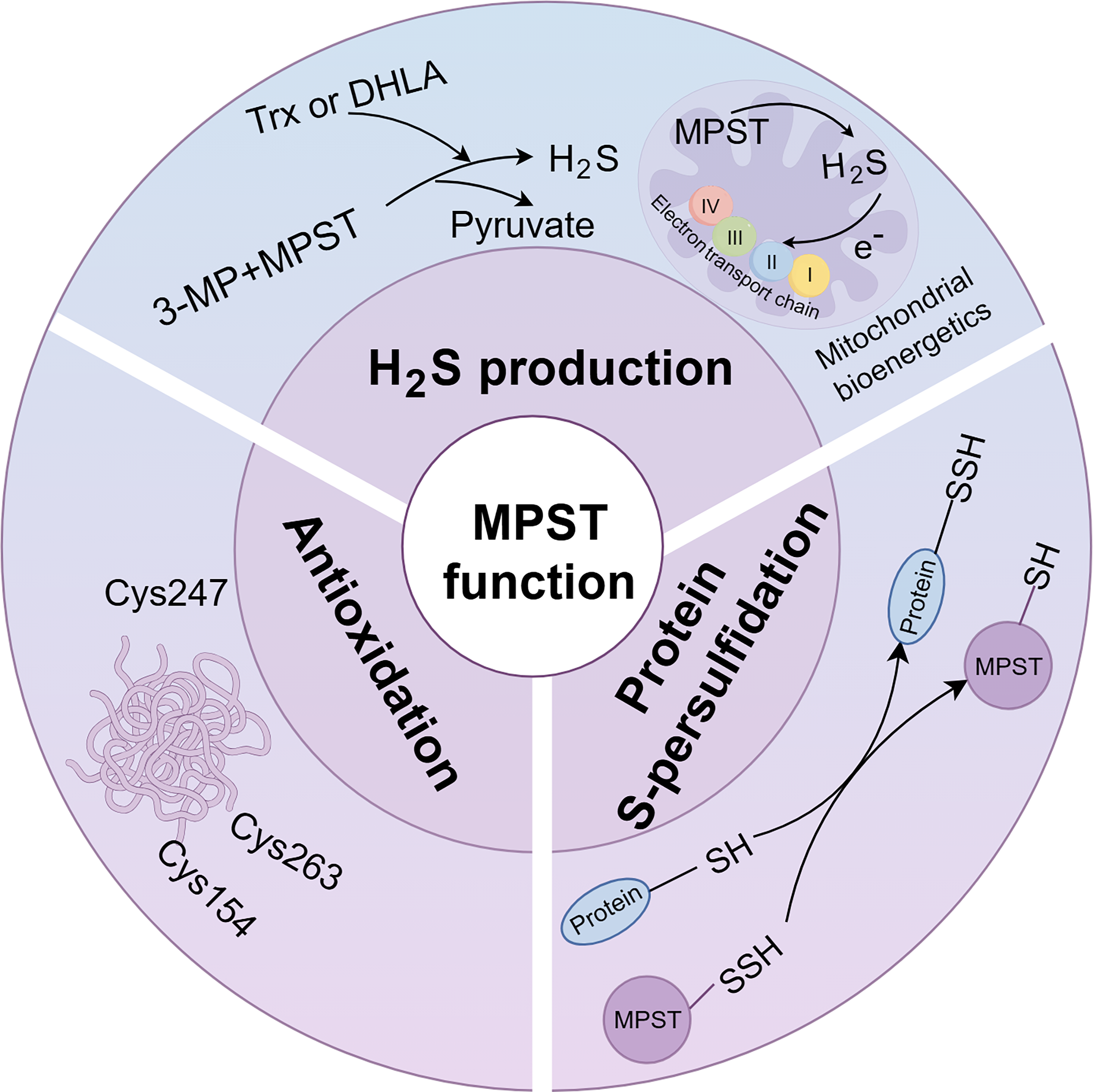
MPST, a Multifunctional Enzyme Responsible for H2S Generation, Protein Persulfidation, and Antioxidation
MPST is widely distributed, although its specific localization varies across organs (Tomita et al., 2016). It is predominantly found in the proximal tubular epithelium of the kidney, pericentral hepatocytes of the liver, cardiac cells of the heart, and neuroglial cells of the brain (Nagahara et al., 1998; Tomita et al., 2016). With respect to its subcellular distribution, MPST is localized to both the cytoplasm and the mitochondria (Kuo et al., 1983). MPST catalyzes H2S production with 3-MP as the substrate. Cells generate 3-MP primarily through two pathways (Fig. 1): the metabolism of
Role of MPST in Liver Detoxification
The liver plays a pivotal role in detoxification processes. Exposure to heavy metals such as cadmium (Cd) significantly disrupts the activity of detoxification enzymes. In adult frogs, Cd exposure reduces the enzyme activities of MPST and CSE while increasing glutathione (GSH) levels and rhodanese activity (Sura et al., 2006). Similarly, Kaczor-Kamińska and colleagues reported decreased hepatic MPST activity in frogs after 10 days of Cd exposure, despite unchanged MPST messenger RNA (mRNA) levels (Kaczor-Kamińska et al., 2020). These findings suggest post-translational regulation of MPST; however, the precise mechanism by which Cd suppresses MPST activity without affecting transcription remains to be elucidated.
Acrylonitrile (AN) and cyanide are common cytotoxic agents found in both natural and manufactured sources. AN undergoes two metabolic pathways: enzymatic conversion to cyanide or conjugation to GSH and protein sulfhydryl groups (Kopecký et al., 1980). Notably, acute AN exposure reduced hepatic H2S levels in a dose-dependent manner in rats, which was accompanied by reduced CBS, CSE, and MPST protein expression in a rat liver cell line (Yang et al., 2021). In contrast, subchronic AN exposure upregulated CBS and CSE protein expression while downregulating MPST expression in the liver; however, levels of hepatic H2S remained unchanged (Yang et al., 2022a).
Similarly, oral administration of potassium cyanide dose-dependently suppressed hepatic MPST activity in mice. Significant reductions were observed at doses of 2.0 and 4.0 times the median lethal dose (LD50), whereas no changes occurred at 0.5 and 1.0 LD50 (Singh et al., 2013). These findings were corroborated by Wróbel et al. (2004), who administered an intraperitoneal injection of a sublethal dose of cyanide (4 mg/kg) to mice. Collectively, these studies demonstrate that cytotoxins differentially regulate hepatic MPST expression and activity, with exposure dose and duration critically influencing outcomes. Nevertheless, the precise molecular mechanisms governing MPST modulation during detoxification remain incompletely understood.
MPST in Liver Injury
Liver injury can result from various endogenous and exogenous factors, including chemical substances, toxic metabolites (e.g., lipopolysaccharide [LPS]), and drugs (Mao et al., 2024). A previous study demonstrated that LPS stimulation upregulated MPST expression in the liver (Ahmad et al., 2016). MPST-mutant mice exhibited significantly reduced levels of cytokines, including tumor necrosis factor α, interleukin-10 (IL-10), and IL-12, while the plasma levels of IL-5 and granulocyte–macrophage colony-stimulating factor were greater in these mice than in wild-type (WT) control mice. However, their survival rates did not differ significantly following LPS challenge (Ahmad et al., 2016). Furthermore, the functional effect of MPST deficiency on the liver during endotoxemia remains unclear and requires confirmation in liver-specific MPST knockout mice.
The therapeutic effects of nonsteroidal anti-inflammatory drugs (NSAIDs) are well established; however, prolonged use or overdose can induce hepatic toxicity (Panchal and Sabina, 2023). Aspirin, a common NSAID, has been shown to increase the hepatic activities of CSE and MPST. This elevation increases the GSH to GSSG ratio, thereby increasing H2S production and providing protection against oxidative stress (Wróbel et al., 2017).
MPST in MASLD
MASLD, previously known as nonalcoholic fatty liver disease (NAFLD), is diagnosed on the basis of evidence of hepatic steatosis plus one of the following three criteria: overweight/obesity, the presence of type 2 diabetes mellitus, or evidence of metabolic dysregulation (Eslam et al., 2020). Subsequent analyses have shown minimal differences between MASLD and NAFLD, as 99% of NAFLD patients meet MASLD criteria (Hagström et al., 2024; Song et al., 2024). Accordingly, this review uses the term MASLD when referring to previous studies. MASLD is currently the most common chronic liver disease, affecting more than 30% of the global adult population (Miao et al., 2024). Excessive intake of methionine, an essential amino acid, is positively correlated with steatosis, inflammation, and fibrosis in MASLD (Navik et al., 2021). Yang et al. (2022b) reported that mice fed a high-methionine diet (HMD; 2.58% methionine) for 11 weeks exhibited decreased hepatic H2S levels. This decrease was accompanied by reduced mRNA expression and activity of CSE and MPST, along with a declining trend in CBS activity. Methionine restriction has been shown to reduce intrahepatic lipid content in a randomized controlled trial (Plaisance et al., 2011). Interestingly, a methionine-restricted diet (MRD; 0.17% methionine) reversed HMD-induced liver steatosis, injury, and fat accumulation (Yang et al., 2022b). Furthermore, MRD-fed mice exhibited reduced hepatic CBS activity but increased CSE and MPST activities, suggesting compensatory mechanisms for H2S production when CBS function is impaired (Tamanna et al., 2018). The role of MPST activity in methionine restriction treatment requires further research to be clearly defined.
Notably, the role of MPST in MASLD development appears to vary depending on the context (Fig. 2). Recently, Katsouda et al. (2025) reported that the double ablation of CSE and MPST induced early-onset liver steatosis in lean mice fed a normal diet, a process mechanistically linked to cholesterol metabolism. Our previous study revealed that free fatty acid-induced upregulation of MPST expression contributes to MASLD pathogenesis by regulating H2S in a CSE-dependent manner and modulating the sterol regulatory element binding transcription factor 1 (SREBP-1) pathway. Partial knockdown of MPST via adenovirus-mediated delivery of Mpst short hairpin RNA or heterozygous deletion of the Mpst gene significantly ameliorated hepatic steatosis in mice fed a high-fat diet (HFD) for 8 weeks (Li et al., 2018). Furthermore, partial knockdown of MPST reduces oxidative stress and inflammation by inhibiting c-Jun N-terminal kinase (JNK) phosphorylation. Additionally, given that Mpst is partially regulated by nuclear factor κB (NF-κB), the ability of astaxanthin to inhibit NF-κB makes it a promising therapeutic approach against MASLD (Davinelli et al., 2022; Li et al., 2018). Notably, short-term HFD consumption (4–8 weeks) typically induces hepatic steatosis that does not progress to severe steatohepatitis and fibrosis (Gallage et al., 2022). However, contrasting findings have been reported with longer periods of HFD feeding. For example, Peh et al. reported decreased CSE and MPST expression but increased CBS expression in mice fed a HFD for 12 or 16 weeks. Plasma H2S levels in these mice showed no significant changes compared with those of the controls (Peh et al., 2014). Furthermore, Katsouda et al. reported increased liver steatosis in Mpst−/− mice fed a HFD for 16 weeks (Katsouda et al., 2022). The biphasic expression pattern of MPST observed in HFD-fed mice over varying durations has potential clinical implications for MASLD stratification, which will be important for the development of stage-specific therapeutic strategies. However, several critical knowledge gaps remain to be addressed. Specifically, future studies must identify the precise point at which MPST decreases and simultaneously characterize corresponding clinical parameters, such as MASLD activity scores, serological biomarkers, radiological findings, and metabolic risk factors. Establishing these correlations is essential for integrating the MPST/H2S pathway into precision treatment strategies.
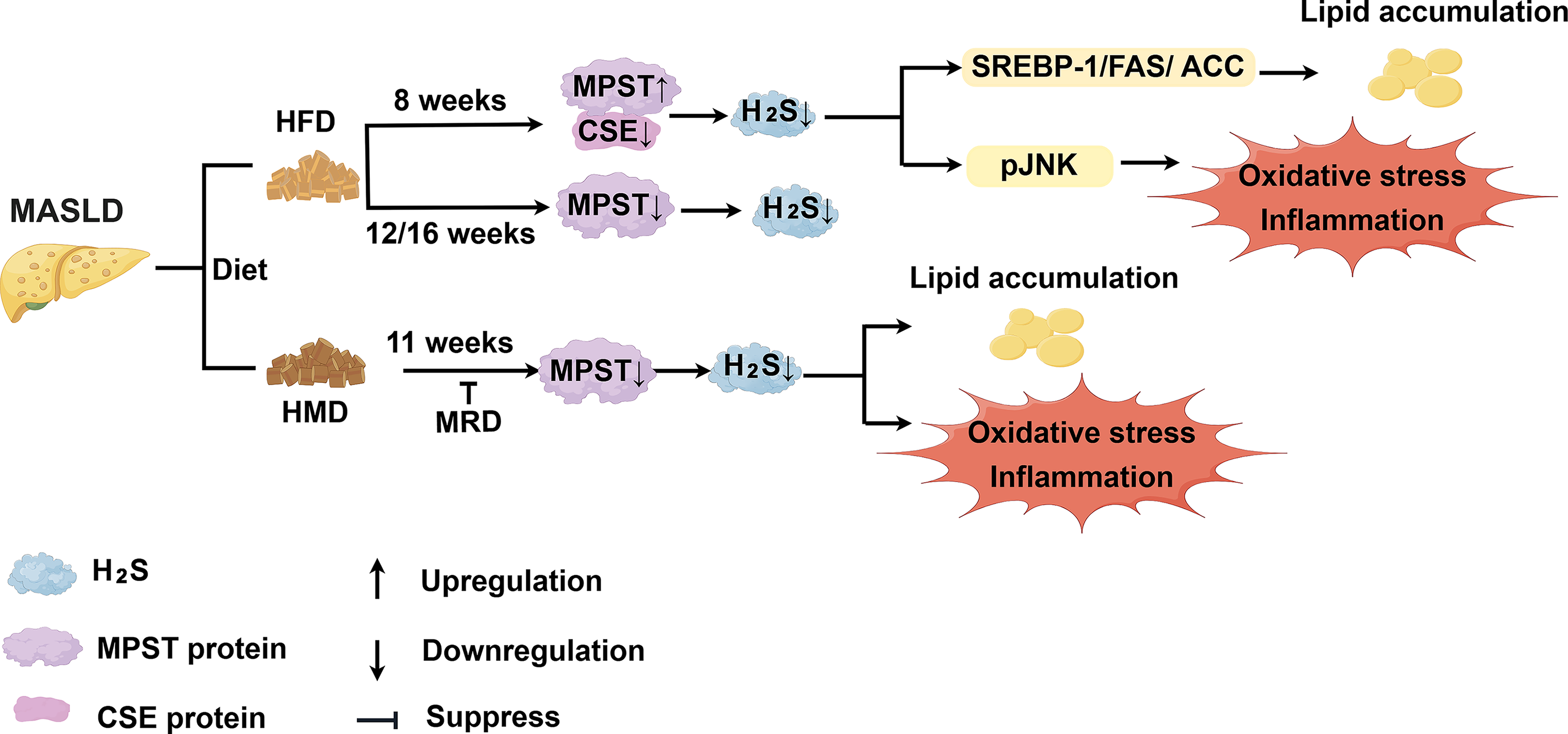
Therapeutic interventions also modulate MPST pathways. Physical exercise increases hepatic H2S levels and upregulates the mRNA expression of CBS, CSE, and MPST (Wang et al., 2017), although the underlying mechanisms require further investigation. Because MPST expression appears to be downregulated following long-term HFD-induced MASLD, exercise may modulate hepatic MPST levels and activity, thereby preventing disease progression in the advanced stages of MASLD. Additionally, vitamin D exerts anti-inflammatory effects in MASLD mice (Roth et al., 2012) and suppresses profibrotic signaling in hepatic stellate cells (HSCs; Beilfuss et al., 2015). Clinically, vitamin D supplementation for 4 months led to a reduction in serum high-sensitivity C-reactive protein and malondialdehyde levels in patients with MASLD (Sharifi et al., 2014). Previous studies have indicated that the active form of vitamin D, 1,25-dihydroxycholecalciferol, exerts anti-inflammatory and antiapoptotic effects, which may be mediated through the downregulation of MPST expression (Ibrahim et al., 2023; Liu et al., 2020). However, the specific mechanism by which 1,25-dihydroxycholecalciferol regulates MPST expression in MASLD remains unknown. Because Mpst ablation exacerbates liver steatosis in mice fed a HFD for 16 weeks (Katsouda et al., 2022), whether vitamin D supplementation exerts the same protective effects in patients with MASLD who are already in an MPST-deficient stage remains unclear. Further studies are needed to determine whether MPST status influences the response to vitamin D therapy in patients with MASLD.
MPST in Alcoholic Liver Disease
Diallyl trisulfide (DATS), an organosulfur compound derived from garlic, attenuates acute ethanol-induced liver injury and mitochondrial dysfunction (Zeng et al., 2008). In alcohol-induced fatty liver and ethanol-exposed human hepatocytes, DATS restores the ethanol-induced suppression of CSE and CBS expression while inhibiting hepatocyte apoptosis through an H2S-mediated mechanism (Chen et al., 2016). Additionally, DATS alleviates alcohol-induced hepatocellular damage by suppressing oxidative stress-mediated pyroptosis by increasing H2S levels (Zhu et al., 2022). Collectively, these findings indicate a protective role for H2S and its generating enzymes in alcoholic liver disease (ALD); however, the involvement of MPST in the pathogenesis of ALD and its regulatory mechanisms remains unexplored. The first step is to determine the expression and activity levels of MPST in ALD.
MPST in Liver Fibrosis and Cirrhosis
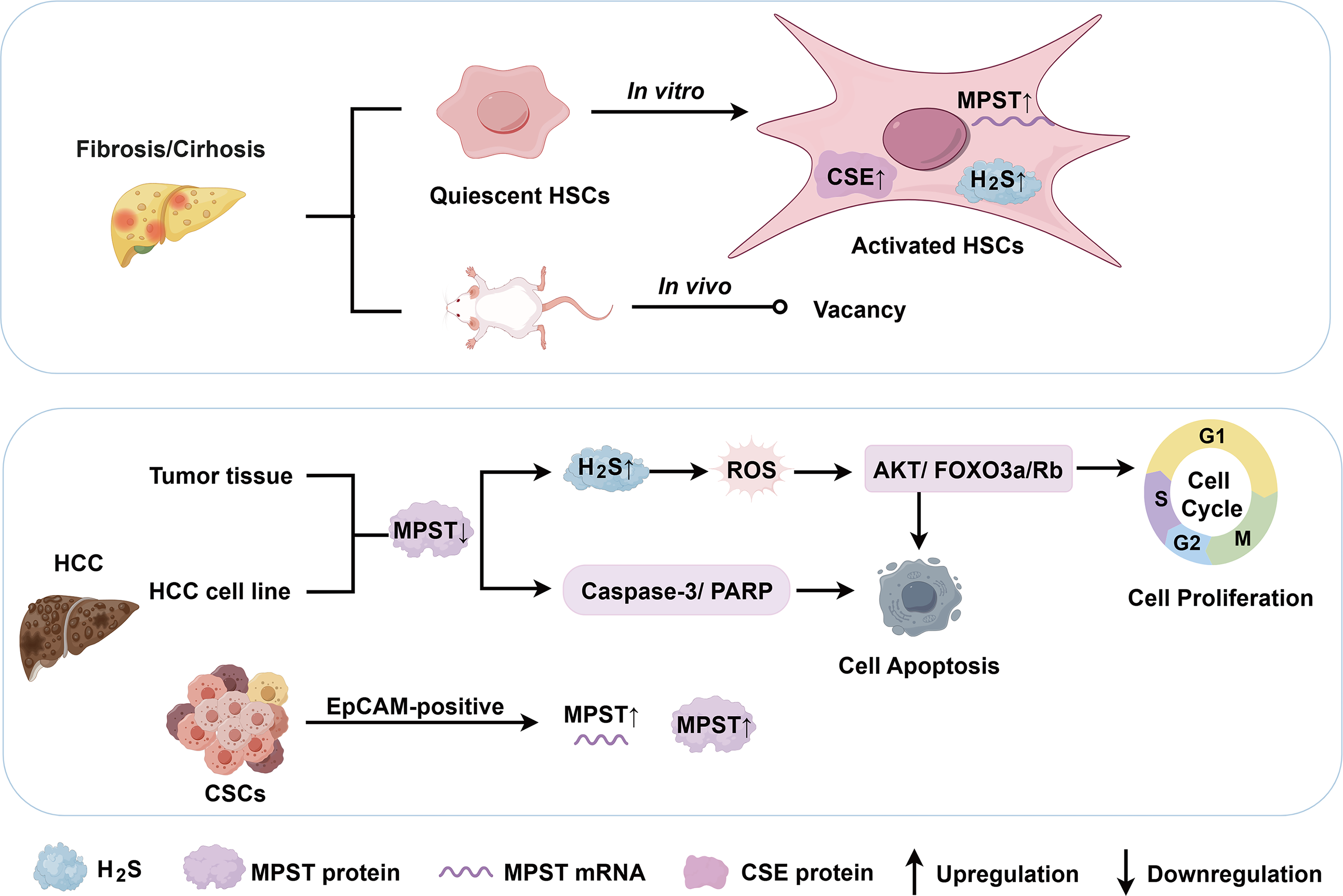
Hepatic fibrosis is a dynamic process characterized by the accumulation of extracellular matrix following chronic liver injury. Common etiologies include chronic viral hepatitis, ALD, and metabolic dysfunction-associated steatohepatitis (MASH; Kisseleva and Brenner, 2021). HSCs play a central role in fibrogenesis (Fig. 3). Damba and colleagues reported lower expression levels of CSE, CBS, and MPST in primary quiescent HSCs from rats than in hepatocytes and Kupffer cells. After 7 days of in vitro activation, HSCs exhibited increased CSE expression and slightly upregulated MPST expression, accompanied by increased H2S levels (Damba et al., 2019). H2S production by HSCs may represent a potential target for antifibrotic intervention. However, in vitro models have limitations for investigating the functions of H2S-generating enzymes, necessitating validation using in vivo fibrosis models.
Cirrhosis results from persistent liver inflammation, leading to diffuse hepatic fibrosis that disrupts the liver vasculature. This structural distortion increases resistance to portal blood flow, which is the primary cause of portal hypertension (Ginès et al., 2021). Notably, reduced expression of the CBS and CSE enzymes and impaired H2S biosynthesis may contribute to endothelial dysfunction and subsequent portal hypertension (Fiorucci and Distrutti, 2016). While these pathways highlight the therapeutic potential of H2S signaling, the specific role of MPST in regulating hepatic endothelial function and circulatory homeostasis remains unexplored. Clarifying endothelial MPST variations and its subsequent contribution to portal hypertension is essential for a comprehensive understanding of H2S-mediated mechanisms in portal hypertension.
MPST in Hepatocellular Carcinoma
Hepatocellular carcinoma (HCC) is a malignant primary liver cancer and is the leading cause of cancer-related mortality worldwide. HCC typically develops in the context of chronic liver diseases of various etiologies, including MASLD, ALD, and chronic hepatitis B or C infections (Hwang et al., 2025). HCC originates from hepatic cancer stem cells (CSCs), which exhibit self-renewal, differentiation, tumorigenicity, and chemoresistance (Zhao et al., 2024). Notably, increased MPST mRNA and protein levels were observed in epithelial cell adhesion molecule (EpCAM)-positive CSCs compared with both EpCAM-negative HCC cells and EpCAM-positive cells from control liver tissues (Khosla et al., 2019). In contrast, our previous study revealed significantly decreased MPST mRNA and protein expression in HCC tissue samples (Fig. 3). Clinically, low MPST expression was shown to correlate with increased tumor size and poor overall survival. MPST overexpression inhibited HCC cell proliferation, induced apoptosis, and suppressed tumor xenograft growth in nude mice. Mechanistically, MPST suppressed the cell cycle via H2S signaling and inhibited the protein kinase B/forkhead box O3a/retinoblastoma protein (AKT/FOXO3a/Rb) pathway during HCC development. In addition, considering that H2S is involved in regulating mitochondrial function and cellular bioenergetics, including the generation of ROS, we found that the overexpression of MPST significantly reduced ROS levels in both in vitro and in vivo HCC models (Li et al., 2022). Another study revealed that genetic silencing of MPST in HepG2 cells exacerbated multiple parameters associated with endoplasmic reticulum (ER) stress and its downstream responses, indicating a role for MPST in the adaptability of HCC cells to ER stress (Panagaki et al., 2020). The MPST–H2S system has demonstrated angiogenic properties and bioenergetic effects in other malignancies (Augsburger and Szabo, 2020), and increasing evidence suggests that H2S plays a biphasic role in HCC pathogenesis (Zhao et al., 2023). Nevertheless, the specific contributions of MPST to HCC development require more in-depth mechanistic investigation. Collectively, MPST is a potential predictor of clinical outcomes in patients with HCC and may serve as both a prognostic indicator and a therapeutic target. MPST-low tumors may constitute a patient subgroup that might benefit from inhibitors targeting the AKT/FOXO3a/Rb axis or ER stress. Given the broad physiological roles of the MPST/H2S system across various organs and systems, off-target effects may result in adverse cardiovascular, neurological, or metabolic consequences (Rao et al., 2023). Considering the biphasic role of H2S in HCC, its effects likely depend on factors such as concentration, duration of exposure, disease context, and complexity of the regulatory mechanisms involved. Historically, the direct therapeutic use of exogenous H2S donors, such as sulfide salts (NaHS and Na2S), has been significantly limited by their uncontrolled release profiles and lack of tissue specificity (Wallace and Wang, 2015). The pharmacology of H2S donors is often complicated by the formation of unpredictable reactive by-products. However, the development of H2S prodrugs has advanced markedly over the past decade. Precision delivery has also been partially achieved through organelle-targeted donors, most notably mitochondrion-targeted AP-39 (Gerő et al., 2016). Recent innovations include slow-release hydrolysis-triggered agents (e.g., ATB-346) and thiol-triggered donors (e.g., arylthioamides; Powell et al., 2018). Additionally, the field has seen the emergence of dual carbonyl sulfide/H2S donors, such as N-thiocarboxyanhydrides (Levinn et al., 2020). Researchers have also developed microenvironment-responsive nanocarriers (e.g., ROS-triggered polymeric micelles; van der Vlies et al., 2023) and therapeutic hybrids that conjugate H2S-releasing moieties to active pharmacophores to achieve synergistic biological effects (Bege et al., 2025). More high-quality in vivo studies are needed to accurately define the therapeutic window of H2S for specific targets and to identify the exact active species responsible for its therapeutic effects. It is promising that new long-acting H2S-releasing agents, which have fewer side effects, could be precisely applied in the clinical treatment of selected HCC patient subtypes in the future. Nevertheless, this represents a significant long-term challenge and a key focus for future research.
Potential Role of MPST in Interorgan Crosstalk
Adipose tissue–liver crosstalk
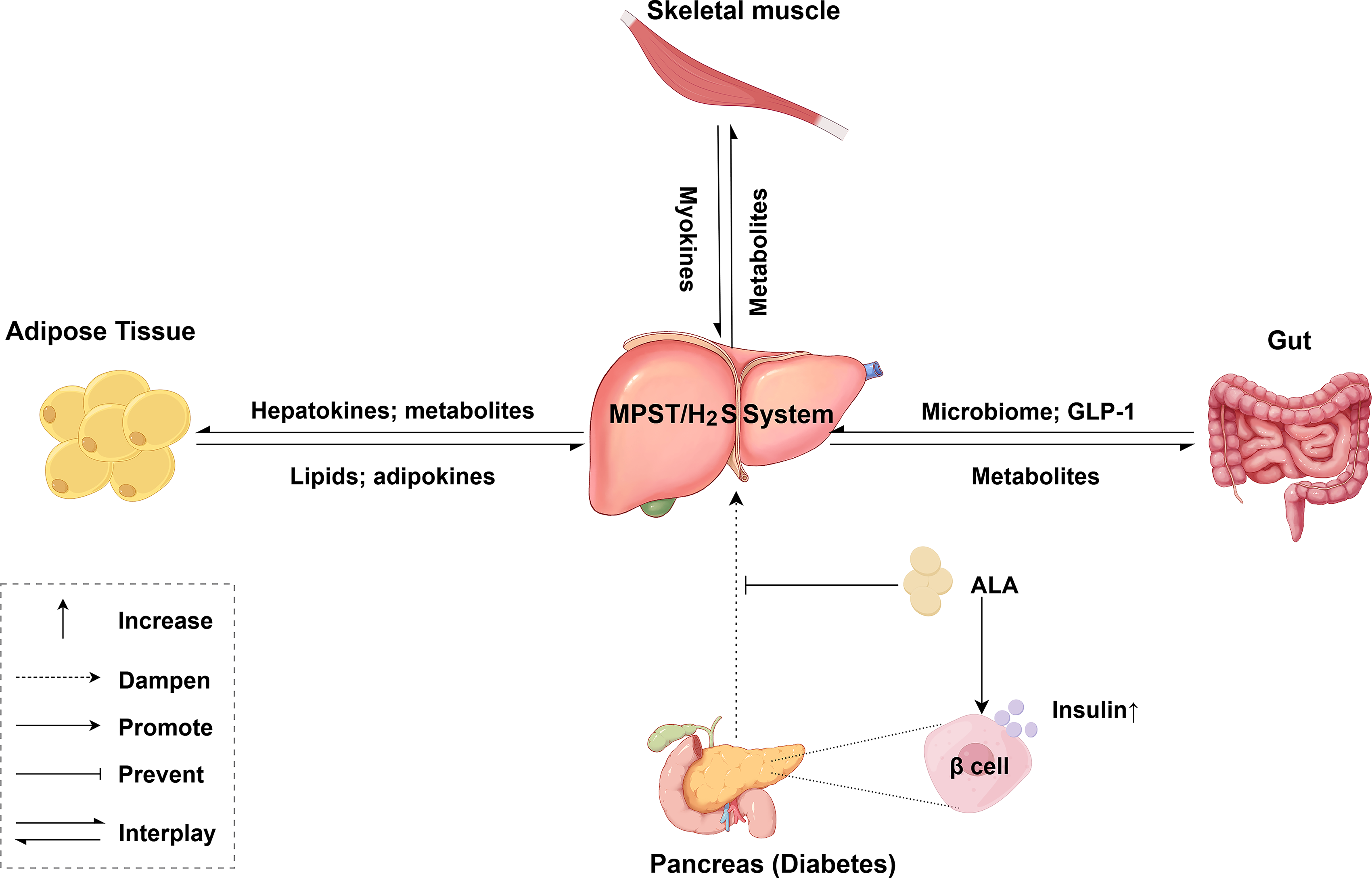
The metabolic interplay between adipose tissue and the liver in obesity is complex and multifaceted. There is increasing evidence that adipose tissue dysfunction contributes to lipid accumulation in the liver through pathological alterations in the secretome, including changes in the levels of adipokines and lipids (Fig. 4; Qureshi and Abrams, 2007). Conversely, hepatic metabolites and hepatokines also modulate adipose tissue physiology. The G protein-coupled bile acid receptor 1 (also known as TGR5) axis induces beige remodeling of white adipose tissue following cold exposure or HFD feeding (Velazquez-Villegas et al., 2018). In addition, energy stress-induced dysregulation of hepatokines impairs adipose tissue metabolism and promotes inflammation, cell proliferation, fibrosis, and other related processes (Zhang et al., 2023). Katsouda and colleagues reported that global MPST deficiency increased white adipose tissue mass and exacerbated hepatic steatosis in HFD-fed mice (Katsouda et al., 2022). To precisely define the metabolic effects of MPST on adipose tissue–liver communication, future studies utilizing adipose- or liver-specific MPST knockout or knock-in models are essential.
Gut–liver axis
The gut microbiome and metabolites are altered in MASLD and MASH (Fig. 4; Li et al., 2003; Loomba et al., 2017). Qi and colleagues demonstrated that H2S produced by Desulfovibrio, a bacterium abundant in HFD-fed mice, suppresses glucagon-like peptide-1 (GLP-1) production, thereby impairing GLP-1-mediated metabolic regulation (Qi et al., 2024). Although hepatocytes lack GLP-1 receptors, GLP-1 receptor agonists effectively attenuate hepatic steatosis, inflammation, and fibrosis in MASH (Yabut and Drucker, 2023). Whether enzyme-mediated H2S production (e.g., via CBS, CSE, or MPST) and/or protein persulfidation participate in the mechanisms by which gut-derived peptides modulate the progression of chronic liver diseases remains unknown. This hypothetical axis requires further investigation.
Muscle–liver crosstalk
Skeletal muscle is a secretory organ that releases hundreds of myokines through paracrine and autocrine signaling (Pedersen and Febbraio, 2012; Fig. 4). Sprenger and colleagues demonstrated that exercise training induces the accumulation of ergothioneine within skeletal muscle mitochondria. In this context, ergothioneine directly targets and activates MPST, enhancing mitochondrial function and exercise performance (Sprenger et al., 2025). Adverse muscle composition, which is associated with poor muscle health, is prevalent among patients with MASLD. These patients exhibit impaired muscle function and a high incidence of metabolic comorbidities (Linge et al., 2021). Moreover, sarcopenia, defined as the progressive and generalized loss of skeletal muscle mass and strength, is observed in 40%–60% of patients with end-stage liver disease (Bhanji et al., 2017). Although the therapeutic benefits of exercise for MASLD and obesity are well established, whether the ergothioneine–MPST axis mediates skeletal muscle–liver crosstalk to regulate hepatic metabolic homeostasis remains unclear.
Pancreatic–liver crosstalk
Glucagon and insulin, which are secreted by pancreatic islets, play critical roles in regulating hepatic glucose and lipid metabolism. Conversely, liver-derived hormones, cytokines, and microRNAs (miRNAs) can directly modulate pancreatic islet function or indirectly influence it by altering metabolite levels (Hædersdal et al., 2023; Xourafa et al., 2024). MASLD and type 2 diabetes frequently coexist because of shared pathogenic features such as insulin resistance (Smith and Adams, 2011; Younossi et al., 2024). Although type 1 diabetes is characterized primarily by insulin deficiency, MASLD remains relatively common among patients with type 1 diabetes (Souza et al., 2025). Alpha-lipoic acid (ALA) prevents liver injury by upregulating hepatic CSE and MPST protein expression (Fig. 4). It also restores pancreatic β-cell function and insulin secretion (Dugbartey et al., 2022). However, further investigations are needed to elucidate whether the precise mechanism of ALA-mediated protection is through direct hepatic effects of MPST/CSE upregulation or via the modulation of pancreas–liver crosstalk mediators. The reduced form of ALA (DHLA) serves as a critical sulfur acceptor during H2S production mediated by MPST (Mikami et al., 2011). ALA alleviates certain symptoms associated with diabetes-related neuropathy, obesity, and schizophrenia because of its antioxidant properties (Salehi et al., 2019). ALA supplementation may represent an effective therapeutic strategy for liver diseases involving MPST deficiency; however, this requires further controlled and robust clinical trials. An overview of the complex interorgan crosstalk is shown in Figure 4.
MPST Involvement in Liver Disorders Associated with Systemic Diseases
Aging
MPST expression exhibits stage-specific and genotype-specific changes during aging. TejJ89 mice subjected to 40% dietary restriction presented increased hepatic MPST and H2S levels and experienced an extended lifespan (Wilkie et al., 2020). In contrast, TejJ48 mice did not exhibit lifespan extension under the same dietary restriction, and their H2S levels decreased. Additionally, TejJ114 mice had a shortened lifespan without changes in hepatic H2S production during dietary restriction. These findings indicate genotypic specificity in the MPST/H2S system, particularly in its response to aging modulation through dietary restriction (Wilkie et al., 2020; Fig. 5). In support of age-related changes, He and colleagues reported increased hepatic MPST expression in 12-month-old mice (He et al., 2024). Regardless of genetic factors, MPST expression appears to undergo stage-specific fluctuations during aging. Szlęzak and colleagues reported decreased hepatic MPST mRNA levels and activity in 60-week-old spontaneously hypertensive rats (SHRs) compared with those of 16-week-old SHRs. Sulfane sulfur levels significantly increased, potentially because of the increased activity of the CSE enzyme. Compared with age-matched Wistar Kyoto controls, the 60-week-old SHRs presented significantly reduced MPST activity despite unchanged gene expression, suggesting hepatic MPST involvement in hypertension development (Szlęzak et al., 2021).
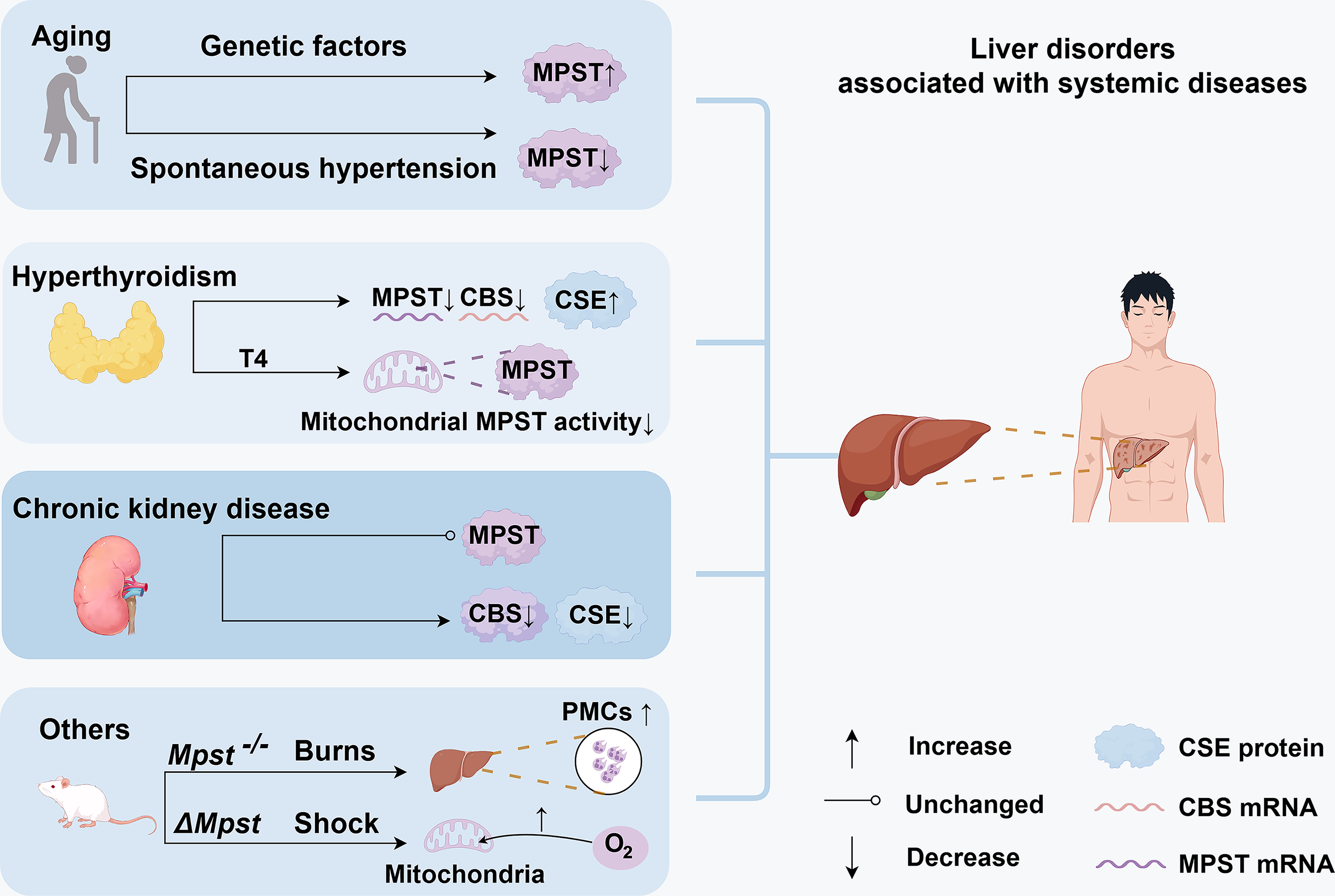
Hyperthyroidism
The liver regulates thyroid hormone activation, deactivation, transport, and metabolism, whereas thyroid hormones modulate hepatocyte activity and metabolic processes (Piantanida et al., 2020). Jeddi and colleagues reported decreased hepatic MPST and CBS mRNA levels and increased H2S levels in hyperthyroid rats, which was partially attributable to increased hepatic CSE expression (Jeddi et al., 2019). Exogenous thyroxine (T4) administration decreased mitochondrial MPST activity without affecting cytosolic MPST activity (Wróbel et al., 2000; Fig. 5). These results suggest that elevated thyroid hormone levels regulate hepatic MPST expression and activity, although the exact mechanisms remain unknown.
Chronic kidney disease
Patients with chronic kidney disease exhibit oxidative stress and significantly reduced plasma H2S concentrations, which correspond to diminished renal expression of CBS, CSE, and MPST. Although hepatic CBS and CSE expression are similarly decreased, hepatic MPST expression remains unchanged (Aminzadeh and Vaziri, 2012). Patients with chronic renal failure have elevated plasma urea levels, which can serve as a source of cyanate. Experimental cyanate treatment in rats reduced hepatic MPST activity, which was consistent with decreased levels of sulfane sulfur and nonprotein sulfhydryl groups (Sokolowska et al., 2011).
Other systemic conditions: Burns and shock
Severe burn injuries cause multiorgan failure and systemic inflammation. WT and Mpst knockout (Mpst−/−) mice exhibited comparable elevations in plasma markers of hepatic injury following burn injury. Plasma H2S levels increased significantly in WT mice in response to burn but remained unchanged in Mpst−/− mice. Mpst−/− mice also exhibited increased accumulation of polymorphonuclear cells in the liver 7 days postburn (Fig. 5). There were no differences between the Mpst−/− and WT groups in inflammatory cytokine levels or burn wound healing. The absence of the H2S-producing enzyme MPST thus slightly exacerbates the development of multiorgan dysfunction (Ahmad et al., 2019). In a murine model of hemorrhagic shock combined with blunt chest trauma, ΔMpst mice exhibited increased mitochondrial oxygen consumption in the kidney and liver (Fig. 5). Despite this, the hemodynamics, metabolism, and mortality did not differ from those of WT mice (Gröger et al., 2019). Whether organ–organ interactions contribute to these processes requires further investigation (Fig. 4).
Promising Models for Studying the Role of MPST in Physiology and Pathology
Genetic mouse model
Nagahara and colleagues established an Mpst-knockout (Mpst-KO) mouse model using the Cre/loxP system, in which Cre was expressed throughout all tissues, and the mice exhibited anxiety-like behaviors without abnormal morphological changes in the brain. Additionally, the male heterozygous KO mice were frequently infertile. Defects in MPST function, such as antioxidant insufficiency and deficiencies in H2S and/or SOx, may underlie these behavioral abnormalities (Nagahara et al., 2013). The authors further reported increased urinary 3-mercaptolactate excretion and enhanced passive systemic anaphylactic responses in Mpst-KO mice generated by CRISPR/Cas9 technology; however, these mice did not exhibit any behavioral disorders (Akahoshi et al., 2020). Our laboratory developed Mpst genetic KO mice on a C57BL/6 background using a frameshift mutation generated by the transcription activator-like effector nuclease system (Li et al., 2018; Wefers et al., 2013). Given the widespread tissue distribution of MPST, its dysfunction may significantly disrupt systemic physiology. To elucidate the liver-specific role of MPST, future studies can use Cre/loxP systems to develop hepatic MPST repression and overexpression models.
MPST activators/agonists
MPST can be activated within the reducing system that includes glutaredoxin, GSH reductase, GSH, and nicotinamide adenine dinucleotide phosphate (Nagahara, 2020). Diallyl disulfide (DADS), a compound derived from garlic, increased hepatic sulfane sulfur levels and CSE and MPST activities in healthy mice without altering hepatic GSH levels. In Ehrlich ascites tumor-bearing mice, DADS restored GSH and sulfane sulfur concentrations while rescuing suppressed CSE activity. Notably, DADS did not affect these parameters in tumor cells, indicating liver-specific efficacy (Iciek et al., 2007). Thus, these findings may offer a promising avenue for the development of MPST activators.
Inhibitors of MPST
To date, potent and selective pharmacological inhibitors of MPST remain limited (Table 1), which has somewhat hindered progress in MPST-related research. α-Keto acids, such as pyruvate, inhibit MPST activity in a concentration-dependent manner (Porter and Baskin, 1996), as does the 3-mercaptopyruvate analog 3-chloropyruvate (Nagahara et al., 2004). However, neither α-keto acids nor 3-chloropyruvate exhibits high selectivity. Additionally, 2-mercaptopropionic acid and 3-mercaptopropionic acid have been identified as noncompetitive inhibitors of MPST with respect to 3-MP (Porter and Baskin, 1995). Incubating U373 cells with 20 μM menadione for 1 h resulted in a 20% decrease in MPST activity, along with ∼23% and 12% reductions in sulfane sulfur and GSH levels, respectively (Wróbel and Jurkowska, 2007). A novel selective MPST inhibitor, HMPSNE (2-[(4-hydroxy-6-methylpyrimidin-2-yl)sulfanyl]-1-(naphthalen-1-yl)ethan-1-one), was used to inhibit MPST with 80%–90% efficacy and had IC50 values in the micromolar range (Hanaoka et al., 2017). HMPSNE significantly promoted lipid accumulation in adipocytes, accompanied by increased intracellular H2S levels, potentially through the upregulation of CBS and CSE expression (Casili et al., 2022). However, its pharmacological profile and applications in liver diseases require further validation.
Characterization of 3-Mercaptopyruvate Sulfurtransferase Inhibitors
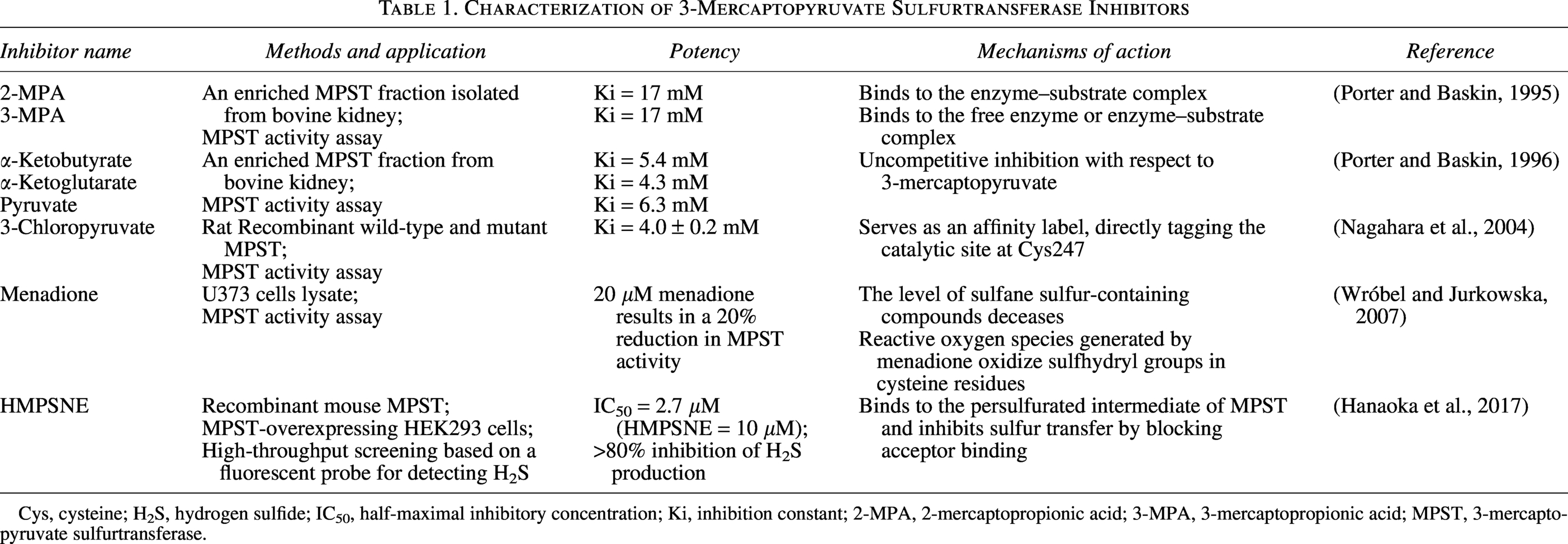
Cys, cysteine; H2S, hydrogen sulfide; IC50, half-maximal inhibitory concentration; Ki, inhibition constant; 2-MPA, 2-mercaptopropionic acid; 3-MPA, 3-mercaptopropionic acid; MPST, 3-mercaptopyruvate sulfurtransferase.
Unanswered Questions and Future Directions
Divergent alterations of MPST in heterogeneous liver pathologies
In this review, we provide an overview of current studies on MPST in the context of liver diseases, including MASLD, ALD, liver fibrosis and cirrhosis, HCC, liver injury, and liver disorders associated with systemic diseases. MPST expression varies across different contexts, showing either upregulation or downregulation (Table 2). This raises the following question: Is MPST a friend or a foe? Resolving this issue requires the consideration of three criteria, namely, context dependency, disease stage, and cell type-specific selectivity.
Alterations in Hepatic Hydrogen Sulfide, Hydrogen Sulfide-Producing Enzymes, and Related Molecules in Liver Disorders
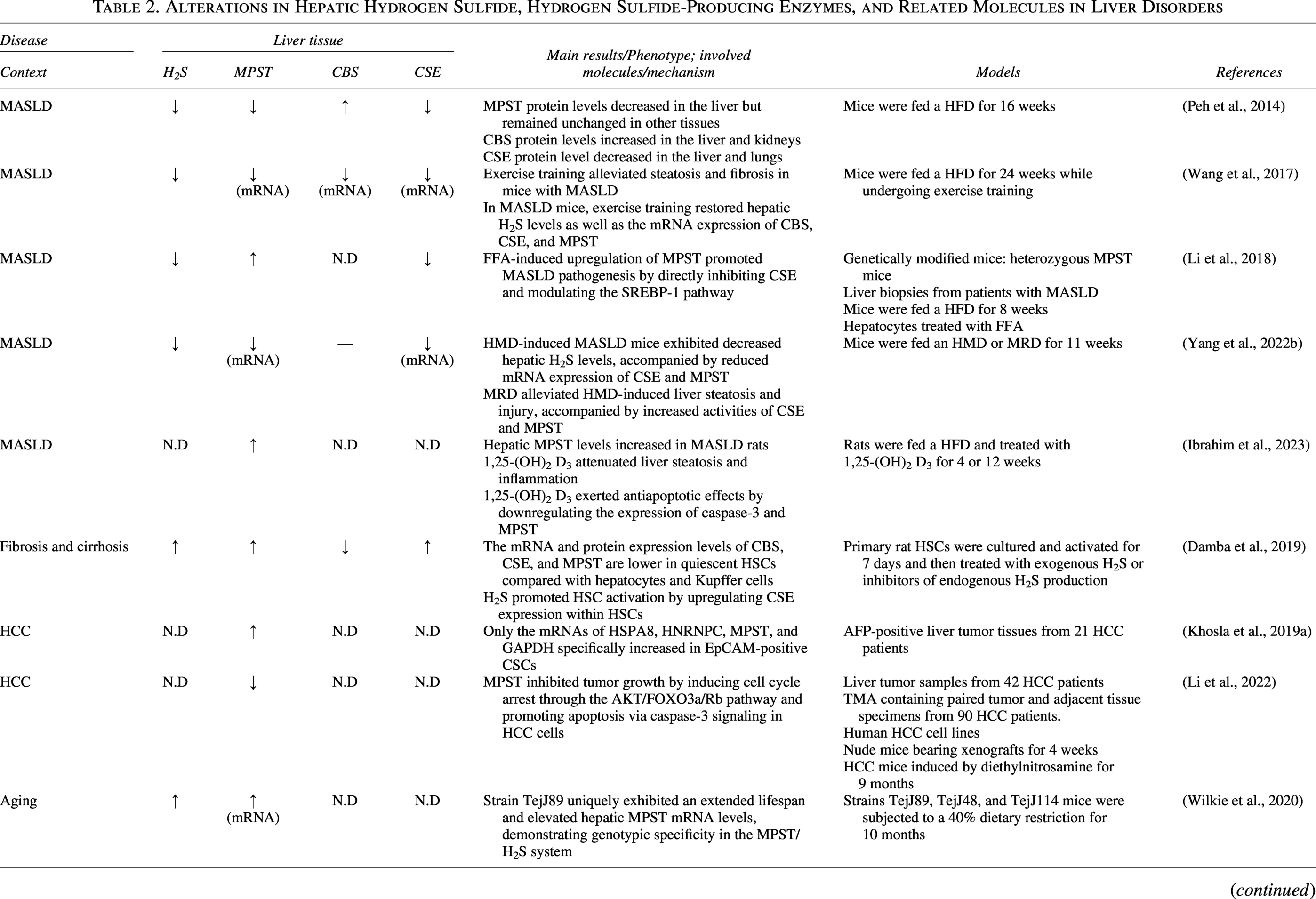
↑, increase; ↓, decrease; -, no significant change; N.D, not detected. Unless otherwise specified, the above symbols refer to changes in protein expression levels.
AN, acrylonitrile; CBS, cystathionine β-synthase; CSE, cystathionine γ-lyase; EpCAM, epithelial cell adhesion molecule; FFA, free fatty acid; HFD, high-fat diet; HMD, high methionine diet; HCC, hepatocellular carcinoma; HSC, hepatic stellate cell; LPS, lipopolysaccharide; MASLD, metabolic dysfunction-associated steatotic liver disease; MPST, 3-mercaptopyruvate sulfurtransferase; MRD, methionine-restricted diet; SHR, spontaneously hypertensive rat; TMA, tissue microarray.
The regulatory function of MPST is context dependent and is partly influenced by the underlying etiologies. Hepatic MPST expression decreases in HMD-induced MASLD but, conversely, increases in HFD-induced MASLD (Li et al., 2018; Yang et al., 2022b). Interestingly, MRD reverses MPST suppression in HMD-induced MASLD mice (Tamanna et al., 2018), highlighting the metabolic link between MPST and methionine availability. Furthermore, the role of MPST in HCC warrants etiology-specific investigation to gain a more in-depth mechanistic understanding. Our previous work reported a general downregulation of hepatic MPST in patients with HCC with various etiologies (Li et al., 2022). Given the increasing prevalence of MASLD/MASH-driven HCC (Huang et al., 2021), future research is needed to characterize and clarify the role of MPST in the progression of MASLD/MASH-related HCC, as well as HCC induced by other etiologies, to advance precision medicine. Moreover, research on the potential role of MPST in ALD, viral hepatitis, and autoimmune liver diseases remains limited. Impaired H2S biosynthesis promotes spontaneous autoimmune disorders and exacerbates immune-mediated diseases (Dilek et al., 2020). Mpst-KO mice exhibit enhanced passive systemic anaphylactic responses, indicating a role for MPST in immune regulation (Akahoshi et al., 2020). Future studies are essential to elucidate whether and how MPST regulates autoimmune liver diseases. Additionally, exploring the MPST/H2S axis in the context of chronic liver disease complications, such as hepatogenic diabetes and hepatorenal syndrome, represents an important research direction, given its regulatory role in glucose and lipid metabolism and its wide tissue distribution (Clementi et al., 2025; García-Compean et al., 2009). The regulatory mechanisms of MPST also show context dependence. Changes in MPST mRNA levels result from transcriptional modulation in response to toxin exposure, metabolic remodeling, inflammation, and aging. In other contexts, MPST may undergo post-translational modifications (PTMs) in response to excessive T4 stimulation.
The role of MPST is critically dependent on disease severity, particularly during the dynamic progression from MASLD to MASH, fibrosis, cirrhosis, and ultimately HCC. In MASLD, discrepancies in MPST expression may result from variations in dietary intervention durations. MPST deletion ameliorated liver steatosis in mice fed a HFD for 8 weeks (Li et al., 2018) but exacerbated steatosis after 16 weeks of HFD consumption (Katsouda et al., 2022; Li et al., 2018). MPST expression may be upregulated during the early onset of MASLD but become downregulated at later stages. Additionally, fibrosis severity is the critical indicator of mortality in patients with MASH, with a higher risk of liver-related complications and death, especially in patients with stage F3 and F4 fibrosis. Given this clinical significance, elucidating the stage-dependent role of MPST in MASH and MASH-associated liver fibrosis requires further study.
The effect of MPST is also influenced by cell-type specificity. Complex crosstalk among multiple cell types occurs during fibrosis and HCC development. MPST expression is increased in steatotic hepatocytes (Li et al., 2018), activated cultured HSCs (Damba et al., 2019), and EpCAM-positive CSCs (Khosla et al., 2019) but decreased in cancer cells during advanced HCC progression (Li et al., 2022), which demonstrates divergent expression patterns. Therefore, cell specificity should be considered when the role of MPST is analyzed.
Mechanistic insights into the role of MPST
Targeting MPST is a promising strategy for managing multiple hepatic disorders. Although most studies have reported alterations in MPST expression or activity, the regulatory mechanisms governing MPST are not yet fully understood. On the basis of previous studies and the biological characteristics of MPST, the underlying mechanisms involve both transcriptional and post-translational control (Pope and Medzhitov, 2018).
We demonstrated that MPST could be partially regulated by NF-κB in FFA-induced steatotic hepatocytes (Li et al., 2018). Other putative transcription factor-binding sites were identified in the promoter region of Mpst, suggesting that multiple potential mechanisms are involved in the transcriptional regulation of Mpst (Fig. 6). Epigenetic mechanisms, including DNA methylation, histone modification, chromatin remodeling, and noncoding RNA modifications (Toh et al., 2017), may also be involved. For example, the expression of the epigenetic reader bromodomain containing 9 (BRD9), which regulates chromatin structure and gene expression by modulating remodeling complexes and selectively recognizing acetyl-lysine markers to recruit transcriptional regulators (Ali et al., 2023), is upregulated in HCC (Cleary et al., 2013). Mutated BRD9 (c.183G>C) increases MPST expression by promoting nuclear factor erythroid 2-related factor 2 binding to the MPST promoter in the presence of cysteine, whereas WT BRD9 inhibits MPST expression in A375 melanoma cells (Hipólito et al., 2024). These findings suggest an epigenetic pathway of MPST regulation in cancer cells (Fig. 6). Typically, miRNAs bind to the 3′-untranslated region of target mRNAs to inhibit their function through translational repression or degradation, but they can also bind to coding sequences to suppress transcription or reduce mRNA stability (Pu et al., 2019). In EpCAM-positive CSCs, increased MPST mRNA and protein levels were associated with decreased expression of miRNAs, including miR-29b-3p and let-7c-5p (Fig. 6); however, the underlying mechanisms remain unclear (Khosla et al., 2019). Although relatively rare, alternative splicing could be another possible post-transcriptional mechanism that may modulate MPST. This process allows a single gene to produce multiple mRNA variants, resulting in different protein isoforms that may have diverse biological functions (Black, 2003). The liver demonstrates complex RNA processing and stress-responsive alternative splicing of numerous metabolic enzymes (Jobbins et al., 2023). Because human MPST has been shown to exist in two isoforms with distinct subcellular localizations (Fräsdorf et al., 2014), it is plausible that MPST is regulated by alternative splicing under specific conditions. However, direct evidence of stress-induced alternative splicing of MPST in the liver is currently lacking, representing a promising avenue for future research.
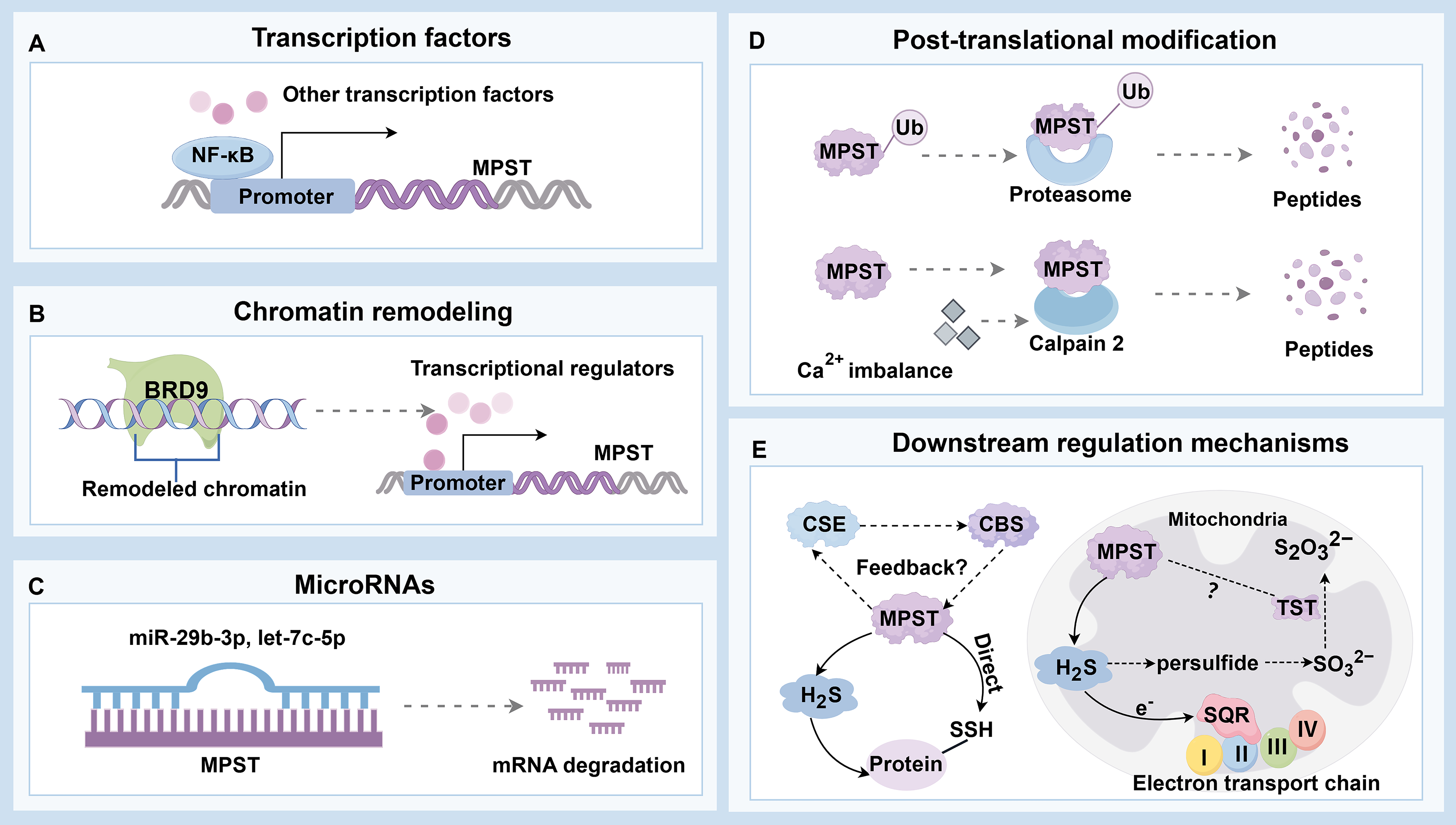
MPST activity or protein levels can change without corresponding transcriptional alterations in some cases, indicating the involvement of PTMs. Owing to computational predictions and advanced detection technologies, more than 400 types of PTMs have been identified (Ramazi and Zahiri, 2021). The cysteine residue, which forms the catalytic site of MPST (Nagahara, 2018), can theoretically undergo at least 10 types of PTMs, such as palmitoylation, ubiquitination, and methylation (Ramazi and Zahiri, 2021). However, the functional impact of these modifications on MPST activity remains unknown. PTMs also critically regulate proteolytic stability, with ubiquitination playing a key role in mediating protein degradation through both the ubiquitin–proteasome system and the autophagy–lysosome pathway (Dikic, 2017; Lee et al., 2023). Ubiquitination can target 20 different amino acids but occurs predominantly on lysine residues (Wu et al., 2023). MPST is ubiquitously expressed across multiple organs and diverse cell types (Tomita et al., 2016). In certain liver disorders, MPST protein levels decrease without corresponding changes in mRNA expression, suggesting that ubiquitination may regulate MPST degradation; this finding warrants further investigation. In addition, cysteine proteases have emerged as potential modulators of MPST expression (Fig. 6). Calpain 2, a calcium-activated cytoplasmic cysteine protease, mediates MPST protein degradation in cerebrovascular damage (Zhu et al., 2025). It is also overexpressed in HCC, promoting tumor proliferation and invasion (Ma et al., 2022). Calcium signaling contributes to multiple pathophysiological processes in the liver, including bile secretion, metabolism, cell proliferation, apoptosis, ER stress, and lipid accumulation (Ali and Petrovsky, 2019; Amaya and Nathanson, 2013). Therefore, calcium-dependent activation of calpain 2 is a potential contributor to MPST degradation in various liver diseases. Collectively, the current evidence indicates that MPST can be regulated at both the transcriptional and post-translational levels. However, the precise upstream regulatory mechanisms of MPST in liver pathologies require further study, especially considering its close association with cysteine metabolism.
With respect to downstream signaling (Fig. 6), MPST expression frequently covaries with changes in CBS and CSE expression in various liver pathologies (Table 2). We demonstrated that hepatic MPST directly interacts with and inhibits CSE expression in MASLD mice (Li et al., 2018). This coordinated modulation suggests potential crosstalk among these three H2S-producing enzymes, although the precise regulatory networks and molecular mechanisms have only been partially elucidated. Additionally, MPST participates in metabolic regulation through the SREBP-1 pathway and contributes to oxidative stress and inflammation via JNK phosphorylation (Li et al., 2018). In the context of HCC, MPST suppresses cell proliferation and promotes apoptosis by inhibiting the AKT/FOXO3a/Rb pathway, effects associated with H2S (Li et al., 2022). Despite these important findings, the detailed molecular mechanisms by which H2S, regulated by MPST, influences these specific cell signaling pathways remain poorly understood. The roles of MPST in other liver diseases beyond MASLD and HCC are also largely unknown. Notably, MPST is involved in direct protein-to-protein transpersulfidation reactions (Pedre et al., 2023), suggesting potential downstream functional mechanisms. Protein persulfidation is involved in the biosynthesis of lipoic acid, thiamine, biotin, iron–sulfur clusters, sulfur-rich bases, and sulfur transfer during RNA synthesis (Mueller, 2006). In adipocytes, proteomic data revealed that the combined partial knockdown of MPST and CBS reduced the persulfidation of proteins involved in adipocyte adipogenesis and function, including the Ras-related protein Rab-4A, isocitrate dehydrogenase 2, aconitase 1, low-density lipoprotein receptor-related protein 1, and galectin 1. Additionally, this knockdown increased persulfidation of the proinflammatory transcription factors, including transcription factor p65, signal transducer and activator of transcription 1, and signal transducer and activator of transcription 3 (Latorre et al., 2022). Although the role of MPST in mediating protein persulfidation in various liver diseases remains unclear, similar modifications may occur in the liver as they do in other organs (Fig. 6). Interestingly, in HEK293 cells, proteins such as α-hemoglobin, triosephosphate isomerase, and heterogeneous nuclear ribonucleoprotein Q have been identified as primary targets of MPST-mediated persulfidation (Pedre et al., 2023). The specific cysteine residues that are modified in these proteins remain to be determined. Mapping the persulfidome in hepatic tissue and at the single-cell level using selective probes such as DCP-Bio1 coupled with liquid chromatography–tandem mass spectrometry (Zivanovic et al., 2019) holds significant promise. This approach might enable the identification of downstream persulfidated targets whose functional integrity is compromised in liver diseases and the discovery of secretory persulfides that may facilitate organ or cell communication. Dynamic alterations in hepatic persulfides can then serve as clinical predictors reflecting disease progression or therapeutic response. In summary, we propose that a vertical MPST signaling pathway is involved in distinct liver pathologies: Upstream transcriptional or post-translational regulation affects the MPST/H2S system, leading to reshaping of the persulfidome and modulating protein function and subsequently contributing to the progression of liver diseases.
Given the distinct mitochondrial localization of MPST and its resulting compartment-specific functions, these factors should be considered. Within mitochondria, MPST-derived H2S maintains bioenergetics by utilizing sulfide quinone oxidoreductase (SQR; an enzyme linked to complex II) to transfer electrons into the mitochondrial electron transport chain (Módis et al., 2013). The loss of MPST triggers a compensatory response in the mitochondrial sulfide oxidation pathway (Nagahara et al., 2019). Thiosulfate sulfurtransferase (TST; also known as rhodanese), which is evolutionarily related to MPST and is specifically localized in mitochondria (Nagahara et al., 1995), was found at increased levels in the hepatic mitochondrial fractions of Mpst-KO mice (Nagahara et al., 2019). TST participates in H2S metabolism within mitochondria, where H2S is initially oxidized to persulfides by SQR. These persulfides are then converted into sulfite (SO32−) by persulfide dioxygenase. Finally, TST further oxidizes the resulting compounds. Sulfite (SO32−) is subsequently converted into thiosulfate (S2O32−) (Kabil and Banerjee, 2014). In addition, both MPST and TST may serve as antioxidant proteins (Nagahara, 2008). To date, evidence demonstrating MPST-regulated liver diseases involving TST processing is limited. It is possible that TST participates in the compensatory response within the mitochondrial sulfide oxidation pathway or oxidative stress when MPST activity changes (Fig. 6). The regulation of MPST may be compartment dependent, as an earlier study revealed that exogenous T4 administration decreases mitochondrial MPST activity without affecting cytosolic MPST activity (Wróbel et al., 2000). Taken together, these findings suggest that the specific localization of MPST in the mitochondria plays a unique role in both energy regulation and sulfur metabolism. Investigating whether and how cytosolic and mitochondrial MPST perform distinct functions in liver disease is a promising direction for advancing MPST-targeted research.
Investigating the hepatic functions of MPST requires careful consideration of interorgan communication and intercellular signaling pathways. Alterations in systemic physiology may affect the hepatic MPST/H2S system, which, in turn, can reciprocally influence other organs (Fig. 4).
Experimental tools for dissecting the functional landscape of MPST in liver diseases: The need for tissue-/cell-specific conditional MPST knockout models
Emerging evidence reveals a paradoxical expression pattern of MPST in HCC: Although MPST expression is markedly decreased in tumor tissues compared with that in adjacent nontumor regions, it is unexpectedly increased in EpCAM+ CSCs relative to that in both EpCAM− HCC cells and EpCAM+ cells from control liver tissues (Khosla et al., 2019; Li et al., 2022). This cell type-specific duality highlights the context-dependent functionality of MPST in hepatic pathologies. Current studies predominantly utilize global Mpst knockout murine models, which inadequately distinguish tissue- or cell-specific effects. Furthermore, global knockout models can introduce systemic confounding factors, such as anxiety-like behaviors and passive systemic anaphylactic responses. To overcome this limitation, it is essential to generate conditional Mpst knockout or knock-in models, such as hepatocyte-, HSC-, or immune cell-specific manipulation models, to elucidate their distinct roles in hepatic pathophysiology. Moreover, developing and validating specific MPST inhibitors or activators remains challenging but holds promise for advancing both biological understanding and therapeutic innovation in the management of hepatic disorders.
Conclusion
This review highlights the significant regulatory functions of MPST in various liver diseases that extend beyond its fundamental roles in H2S generation and liver detoxification. MPST influences lipid metabolism in MASLD and affects the activation of HSCs, possibly through H2S signaling. It also plays a dual role in HCC, depending on the overall tumor tissue context or specific cell types. Furthermore, this review addresses the involvement of MPST in liver disorders associated with systemic diseases and its potential role in interorgan communication, particularly in gut–liver, adipose–liver, muscle–liver, and pancreas–liver crosstalk. As discussed in the text, the role of MPST in a broad spectrum of liver diseases, such as ALD, viral hepatitis, and autoimmune liver diseases, remains unexplored (Akahoshi et al., 2020; Nagahara et al., 2013), underscoring the need for more comprehensive and interdisciplinary studies to elucidate the underlying mechanisms. Specifically, the upstream regulatory mechanisms of MPST expression and activity are largely unknown, and the detailed downstream molecular pathways that mediate its effects on various liver diseases require further investigation. The role of MPST is critically dependent on the disease context and severity, particularly during the dynamic progression from MASLD to MASH, fibrosis, cirrhosis, and HCC. Given the significant prognostic value of fibrosis severity in MASH, elucidating the precise stage-dependent role of MPST in MASH-induced liver fibrosis and HCC is essential. Furthermore, studies should aim to elucidate the cell type-specific functions and regulatory mechanisms of MPST within the complex liver microenvironment. Particular focus should be placed on better understanding protein persulfidation and the interplay among the three enzymes responsible for H2S biosynthesis. Continuous basic research would benefit from more specific modulators and advanced experimental approaches, such as tissue- or cell-specific conditional MPST knockout or knock-in models. A deeper understanding of the biological mechanisms described in this review will eventually lead to more clinical trials with the goal of unlocking the therapeutic potential of the MPST/H2S axis in liver diseases.
Authors’ Contributions
Y.X.: Writing—original draft and methodology. M.L.: Conceptualization, supervision, writing—review, editing, and validation. Both authors approved the final version of the article before submission.
Footnotes
Acknowledgments
Ethical Considerations
This article does not contain any studies with human or animal participants.
Author Disclosure Statement
The authors declare that there are no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding Information
This work was supported by the Natural Science Foundation of Zhejiang Province, China (no. LY22H030010).