
Abstract
Purpose:
Circular RNA (circRNA) has been proved to play a vital role in atherosclerosis (AS) progression, and vascular smooth muscle cells (VSMCs) are involved in the progression of AS. However, the in-depth mechanism by which circRNA regulates VSMC proliferation and migration remains to be elusive.
Methods:
We used tumor necrosis factor-alpha (TNF-α) to treat VSMCs to establish a cell model of AS. We used Cell Counting Kit-8, terminal-deoxynucleoitidyl transferase–mediated nick end labeling, and transwell assays to assess the proliferation, apoptosis, and migration in TNF-α-induced VSMCs. Moreover, the interaction between molecules was measured by RNA-binding protein immunoprecipitation, RNA pull-down, and luciferase reporter assays.
Results:
Our study found that a novel circRNA hsa_circ_0030042, which is derived from its host gene forkhead box O1 (FOXO1), was upregulated in TNF-α-induced VSMCs. Silencing of hsa_circ_0030042 inhibited proliferation and migration while promoting apoptosis in TNF-α-induced VSMCs. Mechanically, hsa_circ_0030042 positively regulated FOXO1 expression via sponging miR-514a-3p.
Conclusions:
Our study stated the vital role of the hsa_circ_0030042/miR-514a-3p/FOXO1 axis and provides a profound understanding about the circRNA in AS.
Introduction
Atherosclerosis (AS), as one of the main factors of coronary heart disease, featured lipid metabolic disorders and derivation from the intima of the vessel wall. 1 The proliferation of vascular smooth muscle cells (VSMCs) participates in the pathogenesis of AS, accompanied by the formation of collagen fibers and the accumulation of lipids. 2 As a consequence, a better understanding of the molecular mechanisms via which VSMCs participate in AS may contribute to finding effective treatments for this disease. In addition, tumor necrosis factor-alpha (TNF-α) has been demonstrated to promote the development of AS. 3 TNF-α-induced VSMCs have been widely used in the study of inflammatory effect of RNAs, such as NOD2 4 and Tanshinone A. 5 As atherosclerosis is a chronic inflammatory disease, 6 we adopted TNF-α-induced VSMCs as the simulation environment of AS.
Circular RNAs (CircRNAs) are a type of non-coding RNAs (ncRNAs) with a covalently closed continuous loop that are formed by back-splicing. 7 Mechanically, circRNAs exert different biological functions in various diseases via serving as competing endogenous RNAs (ceRNAs) to sequester microRNAs (miRNAs) and thereby modulate the levels of miRNA downstream target genes. 8 Recently, many circRNAs have been documented to play important roles in AS development. For example, circRNA‑0044073 is highly expressed in AS and promotes cell proliferation and invasion by interacting with miR-107. 9 CircRNA-PTPRA accelerates AS progression through sponging miR-636 to upregulate SP1. 10 CircRSF1 protects against oxidized low-density lipoprotein (ox-LDL)-induced endothelial cell injury in AS through the miR-758/CCND2 axis. 11 However, more underlying circRNAs which take part in AS pathogenesis need to be identified to improve therapeutic effectiveness.
MicroRNAs are short conserved ncRNAs with about 22 nucleotides in length and regulate gene expression by interacting with the 3′-untranslated region (3′ UTR) of target mRNAs at the level of post-transcription. 12 Accumulating evidence has unveiled that miRNAs are closely implicated in AS progression. MiR-126-5p increases endothelial proliferation and weakens AS via suppressing Dlk1. 13 MiR-652-3p repression elevates endothelial repair and inhibits AS via upregulating Cyclin D2. 14 MiR-let-7g lessens AS via targeting LOX-1. 15
In this study, we focused on exploring the expression of hsa_circ_0030042, derived from its host gene forkhead box O1 (FOXO1) in AS, which is 1 of the 4 mammalian FoxO isoforms of forkhead transcription factors (FoxO1, FoxO3, FoxO4 and FoxO6) and ubiquitously expressed.16,17 We used TNF-α to treat VSMCs to mimic a cell model of AS and found that hsa_circ_0030042 was significantly upregulated in TNF-α-induced VSMCs. Moreover, the functions and mechanisms of hsa_circ_0030042 in TNF-α-induced VSMCs were explored in depth.
Material and Methods
Cell Culture and Treatment
Human VSMCs and HEK293T cells were commercially acquired from ATCC (Manassas, VA, USA). Cells were routinely grown in Dulbecco’s Modified Eagle Medium with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin (Beyotime, Beijing, China) at 37°C in a humidified incubator with 5% CO2. Before stimulation with TNF-α, VSMCs were incubated in serum-free medium for 24 hours. Next, VSMCs were treated with different doses of TNF-α (0, 10, 20, 30, 40, 50 ng/mL) for 24 hours. It is found that cell viability and hsa_circ_0030042 expression were strongly enhanced after the treatment with 50 ng/mL of TNF-α. Therefore, VSMCs were treated with 50 ng/mL of TNF-α for subsequent experiments. Similarly, VSMCs were incubated with different doses of interleukin (IL)-6 (0, 10, 20, 30, 40, 50 ng/mL). It was found that treatment with 30 ng/mL of IL-6 significantly enhanced the cell viability and hsa_circ_0030042 expression, whereas treatment with 40 and 50 ng/mL failed to further improve them. Hence, VSMCs were treated with 30 ng/mL of IL-6 for subsequent experiments.
Vector Construction and Cell Transfection
The short hairpin RNAs (shRNAs) directed against hsa_circ_0030042 (ie, sh-hsa_circ_0030042#1/#2) and FOXO1 (ie, sh-FOXO1#1/#2) were chemically synthesized and procured from GenePharma (Shanghai, China). The overexpression vector pcDNA3.1-FOXO1 was constructed via inserting the sequence of FOXO1 into pcDNA3.1. In addition, miR-514a-3p mimics/inhibitor and the corresponding controls NC mimics/inhibitor were purchased from GenePharma. Lipofectamine 3000 (L3000075; Invitrogen, Carlsbad, CA, USA) was adopted for transfection as per the manufacturer’s protocols.
Cell Counting Kit-8
For detecting cell viability, a Cell Counting Kit-8 (CCK-8) assay kit (Dojindo, Japan) was used. Cells were plated in 96-well plates after transfection. Subsequent to the addition of 10 μL CCK8 solution in each well, cells were subjected to incubation at 37°C for 2 hours. Afterward, the optical density (OD) value at 450 nm of each group was measured with a microplate reader. The OD value at this time represents OD value at 0 hours in figures. Subsequently, the OD value was detected at different time points (24, 48, 72 hours). Assays were performed thrice.
5-Ethynyl-2′-Deoxyuridine
A 5-ethynyl-2′-deoxyuridine (EdU) kit (Ribobio, Guangzhou, China) was used to examine cell proliferation following the manufacturer’s suggestions. Cells were planted into 96-well plates after transfection. Then, 50 μmol/L of EdU solution was added to stain proliferative cells. Next, cells were incubated with anti-EdU working solution after treating with 0.5% Triton X-100 and 4% paraformaldehyde. The nucleus was stained by DAPI, and images were acquired via a fluorescent microscopy. Assays were performed thrice.
Terminal-Deoxynucleoitidyl Transferase–Mediated Nick End Labeling
After transfection cells were washed and fixed, and then permeabilized. Terminal-deoxynucleoitidyl transferase–mediated nick end labeling (TUNEL) assays were conducted following manufacturer’s instructions (Roche). Subsequently, cells were subjected to incubation with terminal dexynucleotidyl transferase (TdT) reaction cocktail, followed by treatment with Click-iT reaction cocktail. The nucleus was stained with DAPI. Images were captured via a fluorescence microscopy. Assays were performed thrice.
Flow Cytometry
Following the manufacturer’s requirements, the Annexin V-FITC/PI apoptosis detection kit (KeyGen, Nanjing, China) was applied for the detection of the apoptosis rate. FACSCanto II flow cytometer (BD Biosciences) was used for measurement. Assays were performed thrice.
Wound Healing
Wound healing assay assessed cell migration activity. After transfection cells were grown in 6-well plates. A scratch was made with a sterile 200 μL plastic pipette tip followed by phosphate-buffered saline (PBS) washing. Three to five randomly selected fields were photographed and recorded as the width of wound at the time point of 0 hours (control group). Cells were then incubated in the medium containing 2% FBS at 37°C with 5% CO2. After 24 hours, a microscope was used for image acquisition. Three to five randomly selected fields were photographed and recorded as the width of wound at the time point of 24 hours (experimental group). The relative distance of wound healing was the rate of migration distance, defined as follows: (the wound width of control group − the wound width of experimental one)/the wound width of control group. Assays were performed thrice.
Transwell
Transwell chambers detected cell migration. Transfected cells were placed in the upper chamber in serum-free medium, and medium containing 10% FBS was added to the lower chamber. After 24 hours, the cells on the upper surface of the insert were slightly abraded by a cotton swab. The cells passing through the lower surface of the insert were migrated cells, which were subsequently stained by crystal violet and then observed under an inverted light microscope. Three to five randomly selected fields were photographed. The results were displayed as the average number of migrating cells per field. Assays were performed thrice. The data were expressed as the average number of the results of 3 independent assays.
Quantitative Real-Time Polymerase Chain Reaction
Total RNA was isolated with the help of RNA iso Plus (Takara, Japan). Reverse transcription was performed using ReveAid First Strand cDNA synthesis kit (Thermo Fisher, USA) to obtain complementary DNA (cDNA). Quantitative polymerase chain reaction (qPCR) was performed using SYBR Green PCR Master Mix (Applied Biosystems, USA). Relative expression of hsa_circ_0030042 or FOXO1was normalized to GAPDH (a housekeeping reference gene for circRNA or mRNA), whereas that of miR-185-5p, miR-654-3p, miR-875-5p, miR-9-5p, miR-532-3p, miR-345-5p, miR-514a-3p, miR-224-5p, miR-5195-3p, miR-27b-3p, miR-145-5p, miR-552-3p, or miR-27a-3p was normalized against U6 (a housekeeping reference gene for miRNAs) and calculated using 2−ΔΔCt method. The PCR conditions were as follows: 95°C for 10 minutes, 95°C for 30 seconds, and 60°C for 30 seconds. Assays were performed thrice. Data were displayed as mean ± standard deviation (SD). The primers used for quantitative real-time polymerase chain reaction (RT-qPCR) are reported in Supplementary Table 1.
RNase R and Actinomycin D Treatment
RNase R (40 U) and actinomycin D (4 µg/mL) and were obtained from Sigma. The expression levels of hsa_circ_0030042 and linear-FOXO1 were assessed by RT-qPCR. Assays were performed thrice.
Western Blot
RIPA lysis buffer (Beyotime Biotechnology, China) was adopted for the extraction of proteins. The protein samples were subjected to 10% SDS-PAGE gel for separation and shifted onto polyvinylidene difluoride membranes (Millipore, Billerica, MA, USA). After blocking, the membranes were subjected to incubation with primary antibodies containing anti-FOXO1 (Abcam, 1/1000), anti-P65 (Abcam, 1/100), anti-IκB-α (Cell Signaling Technology, 1/1000), and anti-GAPDH (Abcam, 1/1000) overnight at 4°C, followed by incubation with secondary antibodies at room temperature for 1 hour. The immunoblots were measured using enhanced chemiluminescence (ECL) detection system (Thermo Scientific). Assays were performed thrice.
Subcellular Fractionation
To separate cytoplasmic and nuclear RNA, a PARIS kit (Life Technologies) was used following the supplier’s protocols. The collected cells were cultured and then placed on ice followed by resuspension with cell fractionation buffer. Afterward, cells were centrifuged to separate cytoplasmic fractions and nuclear pellets. Later, RT-qPCR assessed the levels of extracted RNAs. Assays were performed thrice.
Fluorescent In Situ Hybridization and Immunofluorescence Assay
Cellular co-localization of hsa_circ_0030042 and FOXO1 was determined by fluorescent in situ hybridization (FISH) and immunofluorescence (IF) assays. The FISH assay was performed using hsa_circ_0030042-specific probe. Cells were incubated with hsa_circ_0030042 probe after fixation, followed by incubating with FOXO1 antibody and P65 antibody (Abcam, 1/250) after washing and sealing. Then cells were incubated with secondary antibodies conjugated with Alexa Fluor 488. DAPI was applied for the counterstaining of nuclei. Images were acquired with a confocal microscope. Assays were performed thrice.
RNA-Binding Protein Immunoprecipitation
RNA-binding protein immunoprecipitation (RIP) experiments were conducted with the help of EZ-Magna RIP Kit (Millipore) based on the manufacturer’s instructions. The transfected cells were washed using ice-cold PBS and subjected to the mixture with RIP lysis buffer. Subsequently, the lysis products were incubated with Protein A/G Agarose magnetic beads conjugated with anti-Ago2 antibody or control anti-IgG antibody overnight at 4°C. The immunoprecipitated RNAs were subjected to RNA isolation and detected by RT-qPCR. Assays were performed thrice.
RNA Pull Down Assay
The transfected cells were washed with ice-cold PBS and then lysed in lysis buffer. Cell lysates were incubated with biotin (Bio)-labeled probes specifically against hsa_circ_0030042 (RiboBioa) at room temperature. Streptavidin-coupled Dynabeads (Invitrogen) was used for capturing the hsa_circ_0030042-bound miRNA complexes. The RT-qPCR was conducted to analyze the extracted miRNAs from the pull-down complexes after the brief wash of the beads with lysis buffer. Assays were performed thrice.
Luciferase Reporter Assay
The wild-type (WT) and mutant (MUT) sequences of hsa_circ_0030042 or FOXO1 3′ untranslated region (UTR) were sub-cloned into psiCHECK2 (Fisher Scientific) to generate corresponding reporter vectors, including psiCHECK-hsa_circ_0030042-WT, psiCHECK-hsa_circ_0030042-MUT, psiCHECK-FOXO1 3′ UTR-WT, or psiCHECK-FOXO1 3′ UTR-MUT. The TNF-α-induced VSMCs/HEK293T cells were maintained in 24-well plates overnight and transfected with luciferase reporter vector together with miR-514a-3p mimics or NC mimics. After 48 hours, the Firefly and Renilla luciferase activities were measured using dual-luciferase reporter assay system (Promega). The ratio of luminescence of firefly luciferase activity is normalized to the internal reference Renilla luciferase. Assays were performed thrice.
Statistical Analysis
Data were presented as mean ± SD from 3 experiments. Statistical significance was assessed using the Student t test or analysis of variance (ANOVA) followed by post hoc tests (Dunnett or Tukey). GraphPad Prism 7.0 was adopted to analyze statistics. Differences were considered to be statistically significant when p value was less than 0.05.
Results
The Characterization of hsa_circ_0030042 in TNF-α-Induced VSMCs
The VSMCs were treated with TNF-α for the follow-up experiments. And proinflammatory cytokine IL-6 was also applied to treat VSMCs to prove these data are generally applicable. CCK-8 and EdU assay kits were used to measure the effect of TNF-α and IL-6 on cell proliferation. As indicated in Figure S1A and S1B, TNF-α or IL-6 treatment evidently promoted VSMC proliferation. On the contrary, the apoptosis of VSMCs was declined after the treatment of TNF-α or IL-6 (Figure S1C and S1D). Moreover, through wound healing and transwell assays, we confirmed that TNF-α or IL-6 induced the migration in VSMCs (Figure S1E and S1F). It has been reported that FOXO1 plays an important role in AS progression. 18 To investigate AS-related circRNAs, we detected the expression of circRNAs derived from FOXO1 and found that hsa_circ_0030042 was significantly upregulated in VSMCs treated with TNF-α or IL-6 (Figure S1G). Thus, we selected hsa_circ_0030042 for follow-up experiments. We found that hsa_circ_0030042 was back-spliced of FOXO1 (Figure 1A). Besides, we designed divergent primers to characterize hsa_circ_0030042. The results suggested that hsa_circ_0030042 could only be amplified from cDNA by divergent primers, indicating the circular structure of hsa_circ_0030042 (Figure 1B). To further confirm the circular structure of hsa_circ_0030042, we used RNase R to treat both VSMCs and TNF-α-induced VSMCs and found that the linear transcripts of FOXO1 were degraded after RNase R treatment, while such treatment failed to degrade hsa_circ_0030042 (Figure 1C). At the same time, upon the treatment of actinomycin D, the half-life of linear form of FOXO1 was decreased, whereas that of hsa_circ_0030042 remained unchanged (Figure 1D). All these results demonstrated that hsa_circ_0030042 was highly expressed in TNF-α-induced VSMCs.
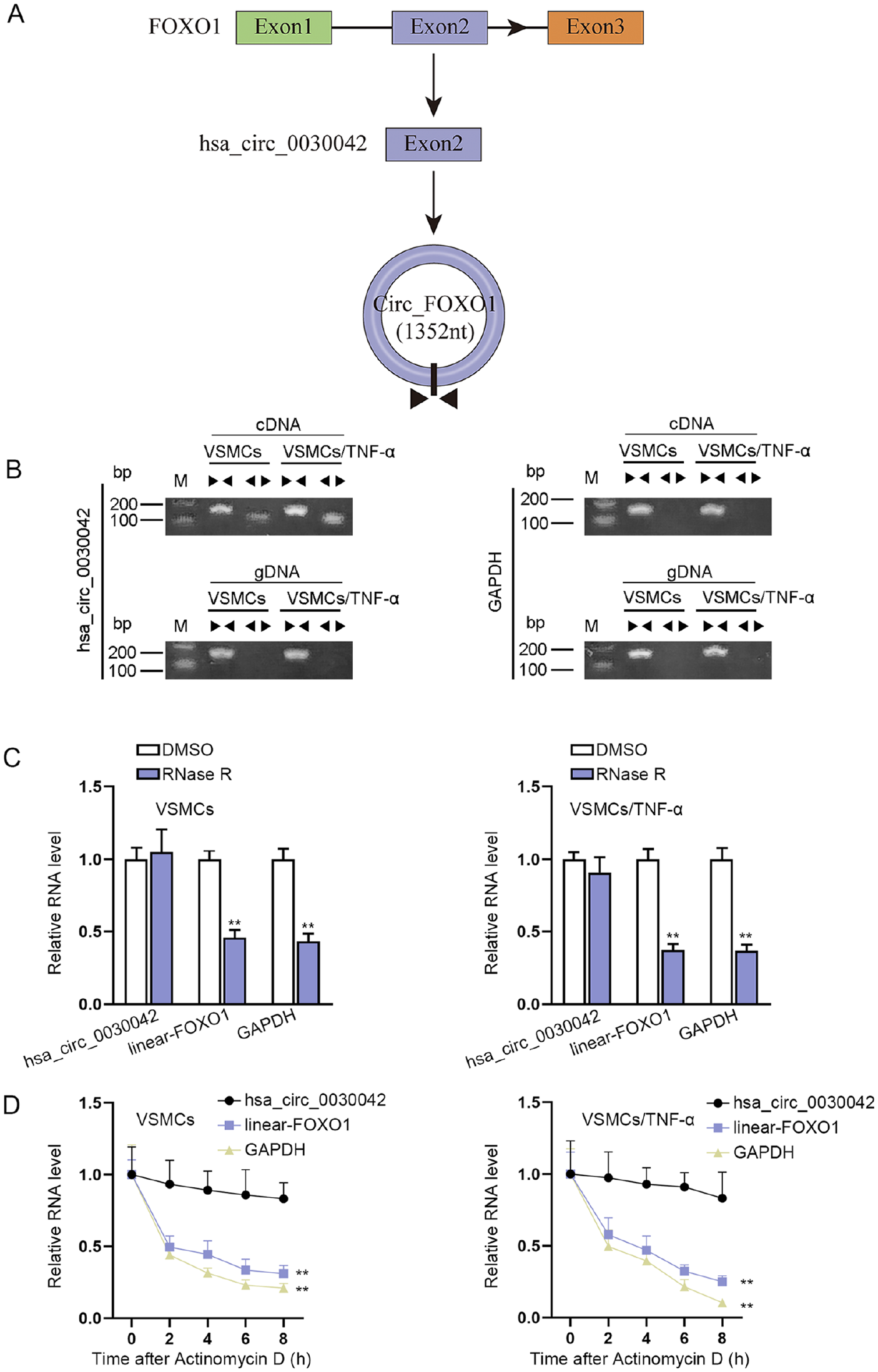
The characterization of hsa_circ_0030042 in TNF-α-induced VSMCs: (A) the “head-to-tail” splicing of hsa_circ_0030042, and (B) the existence of hsa_circ_0030042 was validated by RT-PCR. Divergent primers amplified hsa_circ_0030042 in cDNA but not in gDNA. GAPDH served as a negative control. (C) Hsa_circ_0030042 and linear transcripts of FOXO1 from cells were treated with or without RNase R for RT-qPCR, and (D) RNA stability of hsa_circ_0030042 and linear transcripts of FOXO1 was assessed after actinomycin D treatment. Statistical significance was determined using the Student t test in C; 1-way ANOVA in D. TNF-α, tumor necrosis factor-alpha; VSMCs, vascular smooth muscle cells; RT-PCR, real-time polymerase chain reaction; cDNA, complementary DNA; gDNA, genomic DNA; FOXO1, forkhead box O1; RT-qPCR, quantitative real-time polymerase chain reaction; ANOVA, analysis of variance.
Hsa_circ_0030042 Silence Inhibits TNF-α-Induced VSMC Proliferation and Migration While Elevating Apoptosis
Loss-of-function experiments conducted using shRNAs specifically targeting hsa_circ_0030042 were transfected into TNF-α-induced VSMCs (Figure 2A). CCK-8 and EdU assays depicted that hsa_circ_0030042 silence repressed the proliferative ability of TNF-α-induced VSMCs compared with the control group (Figure 2B and 2C). Besides, the apoptosis rate of VSMCs was elevated when hsa_circ_0030042 was knocked down after TNF-α treatment (Figure 2D and 2E). Moreover, hsa_circ_0030042 depletion definitely reduced the capacity of migration in VSMCs treated with TNF-α (Figure 2F and 2G). Overall, hsa_circ_0030042 silence inhibits TNF-α-induced VSMC proliferation and migration while elevating apoptosis.
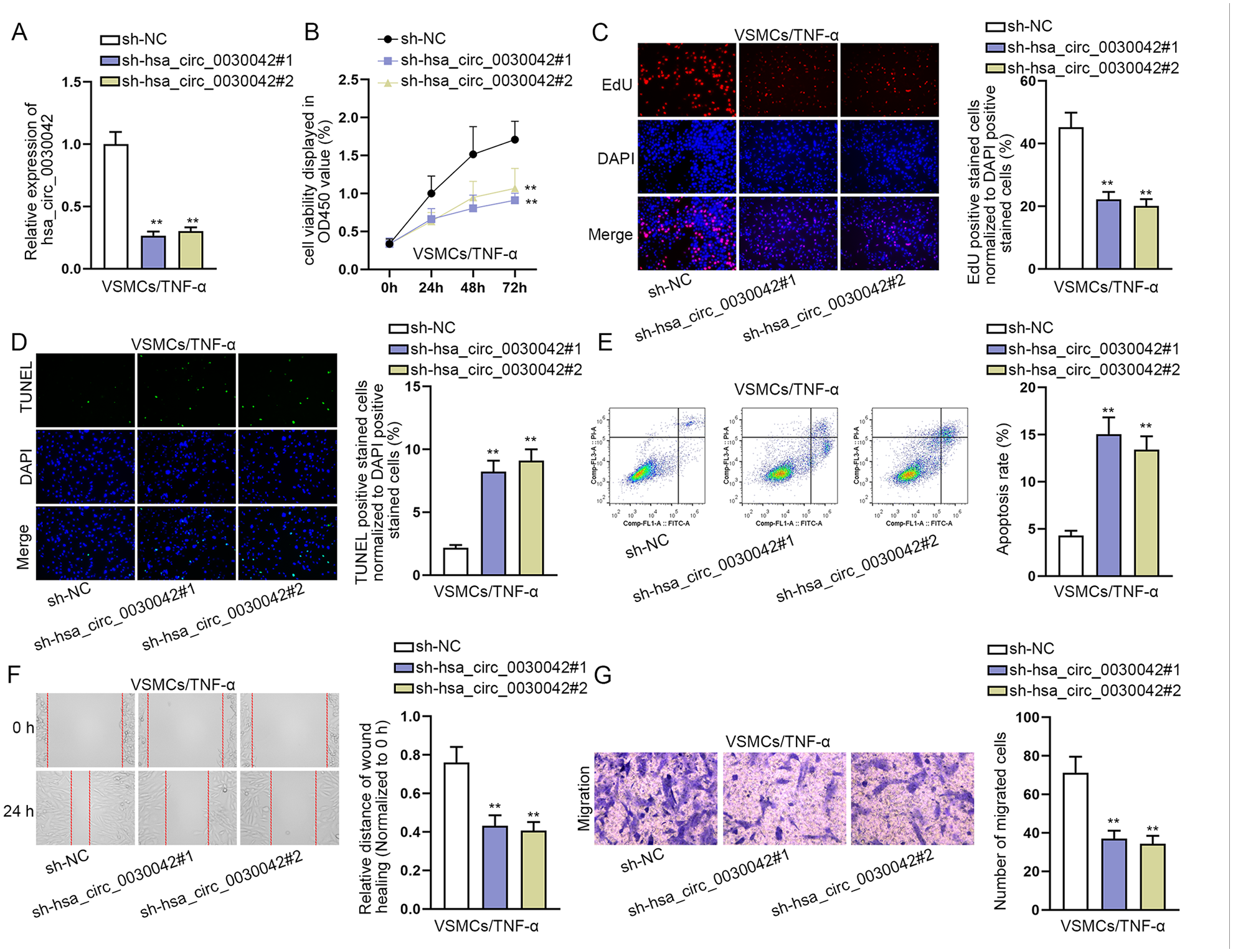
Hsa_circ_0030042 silence inhibits TNF-α-induced VSMC proliferation and migration while it elevates apoptosis: (A) silencing efficiency of sh-hsa_circ_0030042#1/2 was assessed by RT-qPCR, (B and C) impacts of hsa_circ_0030042 silence on TNF-α-induced VSMC proliferation by CCK-8 and EdU (the fluorescent color of proliferating cells was red and that of nuclei of cells was blue) assays, (D and E) impacts of hsa_circ_0030042 silence on TNF-α-induced VSMC apoptosis by TUNEL (the fluorescent color of cells undergoing apoptosis was green and that of nuclei of cells was blue) assay and flow cytometry analysis, and (F and G) impacts of hsa_circ_0030042 silence on TNF-α-induced VSMC migration by wound healing and transwell (the fluorescent color was purple) assays. Statistical significance was determined using 1-way ANOVA in A, B, C, D, E, F, and G. TNF-α, tumor necrosis factor-alpha; VSMCs, vascular smooth muscle cells; RT-qPCR, quantitative real-time polymerase chain reaction; CCK-8, cell counting kit-8; TUNEL, terminal-deoxynucleoitidyl Transferase Mediated Nick End labeling; ANOVA, analysis of variance.
Hsa_circ_0030042 Positively Regulates FOXO1 Expression
CircRNAs can exert their functions by cooperating with their host genes. To test whether hsa_circ_0030042 exerts biological functions in TNF-α-induced VSMCs via interacting with FOXO1, we first measured FOXO1 expression in VSMCs treated with or without TNF-α using RT-qPCR. As shown in Figure 3A, FOXO1 expression was elevated in VSMCs treated with TNF-α. We then detected FOXO1 expression in TNF-α-induced VSMCs after hsa_circ_0030042 silence and found that hsa_circ_0030042 downregulation declined FOXO1 expression (Figure 3B). Moreover, the results from subcellular fractionation assays illuminated that hsa_circ_0030042 was mainly expressed in the cytoplasm (Figure 3C). Meanwhile, we validated the co-localization of hsa_circ_0030042 and FOXO1 in the cytoplasm (Figure 3D). Existing evidence has suggested that cytoplasmic ceRNAs can regulate mRNA expression via sponging miRNAs. Thus, we performed RIP assays using Ago2 antibody, because Ago2 is a key component of the RNA-induced silencing complex (RISC) that binds miRNA complexes to target mRNAs. 19 We found that hsa_circ_0030042 and FOXO1 were preferentially enriched in the immunoprecipitates pulled down by the Ago2 antibody rather than IgG (Figure 3E), implying that hsa_circ_0030042 might positively regulate FOXO1 expression via sponging miRNAs. After the efficiency of sh-FOXO1#1/2 was assessed (Figure S2A), we performed functional experiments and found that FOXO1 silence significantly repressed VSMC proliferation and migration and promoted apoptosis after TNF-α treatment (Figure S2B–S2G). All these results indicated that hsa_circ_0030042 positively regulated FOXO1 expression.
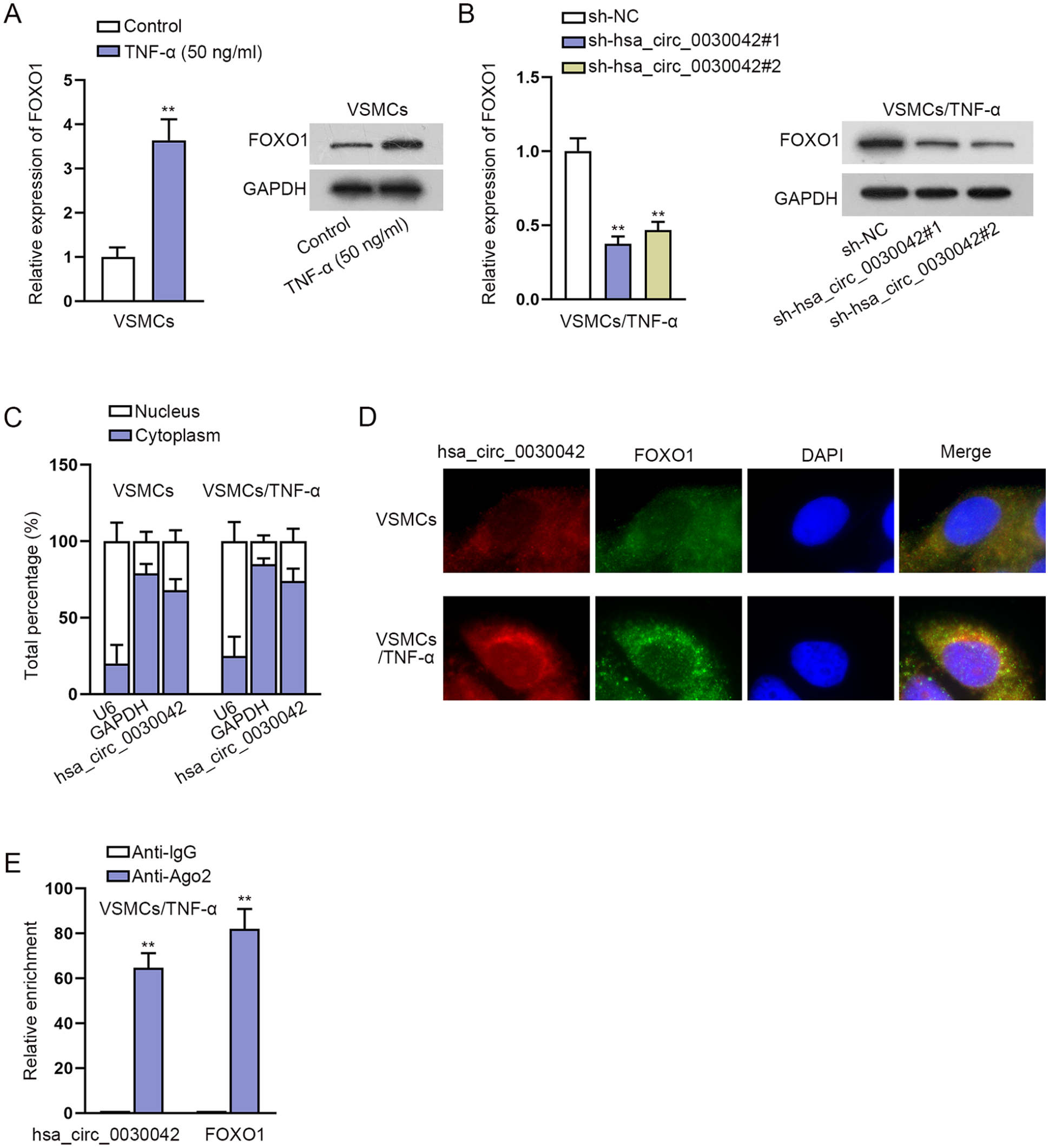
Hsa_circ_0030042 positively regulates FOXO1 expression: (A) FOXO1 expression in VSMCs treated with or without TNF-α using RT-qPCR and Western blot, (B) FOXO1 expression in TNF-α-induced VSMCs after hsa_circ_0030042 silence was measured by RT-qPCR and Western blot, (C) subcellular fractionation assays detected the cellular location of hsa_circ_0030042 in VSMCs and TNF-α-induced VSMCs, (D) FISH (the fluorescent color of hsa_circ_0030042 group was red, that of FOXO1 group was green, and that of nuclei was blue) assays detected the co-location of hsa_circ_0030042 and FOXO1 in VSMCs and TNF-α-induced VSMCs, and (E) RIP assays detected the enrichment of hsa_circ_0030042 and FOXO1 in Ago2 group. Statistical significance was determined using the Student t test in A, C and E; 1-way ANOVA in B for RT-qPCR. FOXO1, forkhead box O1; VSMCs, vascular smooth muscle cells; TNF-α, tumor necrosis factor-alpha; RT-qPCR, quantitative real-time polymerase chain reaction; FISH, fluorescent in situ hybridization; RIP, RNA-binding protein immunoprecipitation; ANOVA, analysis of variance.
Hsa_circ_0030042 Regulates FOXO1 Expression via Sponging miR-514a-3p and Modulates NF-κB Pathway Through P65 Translocation
To explore the miRNAs of hsa_circ_0030042 and FOXO1, starBase (http://starbase.sysu.edu.cn) database was used to predict which miRNAs interact with hsa_circ_0030042 or FOXO1. Consequently, 13 miRNAs were identified for hsa_circ_0030042 and FOXO1 in common (Figure 4A). To screen which miRNA truly interacts with hsa_circ_0030042 in TNF-α-induced VSMCs, RNA pull-down assays were carried out. The results showed that biotinylated probes against hsa_circ_0030042 could only pull down endogenous miR-514a-3p (Figure 4B). The RIP assays further confirmed that hsa_circ_0030042, miR-514a-3p, and FOXO1 coexisted in RISC (Figure 4C). To determine the function of miR-514a-3p in AS, we constructed miR-514a-3p overexpression plasmids. The results revealed that miR-514a-3p mimics significantly upregulated miR-514a-3p expression (Figure S3A). The proliferation ability of VSMCs transfected with miR-514a-3p mimics was decreased after the treatment of TNF-α (Figure S3B and S3C). Meanwhile, the apoptosis of TNF-α-induced VSMCs was increased after miR-514a-3p overexpression (Figure S3D and S3E). Besides, miR-514a-3p elevation inhibited VSMC migration under TNF-α treatment (Figure S3F and S3G). Together, miR-514a-3p could inhibit TNF-α-induced VSMC proliferation and migration while promoting cell apoptosis. Moreover, the binding sites of hsa_circ_0030042 and miR-514a-3p along with the mutant sites are displayed in Figure 4D, and luciferase reporter assays further proved that enforced miR-514a-3p expression significantly reduced the luciferase activity of hsa_circ_0030042 WT, but barely affected that of hsa_circ_0030042 mutant sites (Figure 4E). Furthermore, we performed RT-qPCR to detect the expression of hsa_circ_0030042 after the enhancement of miR-514a-3p. The results showed that miR-514a-3p overexpression could not influence the expression of hsa_circ_0030042, proving that miR-514a-3p was sponged by hsa_circ_0030042 (Figure 4F). Similarly, the complementary sequences of FOXO1 3′ UTR and miR-514a-3p as well as the mutant sequences were predicted, and miR-514a-3p elevation led to a decrease in the activity of FOXO1 3′ UTR-WT but not FOXO1 3′ UTR-MUT (Figure 4G and 4H). In addition, we determined whether hsa_circ_0030042 regulates FOXO1 expression via targeting miR-514a-3p. After detecting the efficiency of miR-514a-3p inhibitor (Figure 4I), we inhibited miR-514a-3p expression and found that miR-514a-3p suppression reversed the reduced FOXO1 expression caused by hsa_circ_0030042 silence in VSMCs after TNF-α treatment (Figure 4J). To expand the findings beyond the FOXO1 axis, we also explored whether the nuclear factor (NF)-kB pathway is also affected by hsa_circ_0030042. We performed IF and Western blot analyses to assess the effect of hsa_circ_0030042 on P65 nuclear translocation, finding that hsa_circ_0030042 inhibition translocated P65 to cytoplasm (Figure S4A and S4B). Taken together, hsa_circ_0030042 regulates FOXO1 expression via sponging miR-514a-3p and modulates NF-kB pathway through P65 translocation.
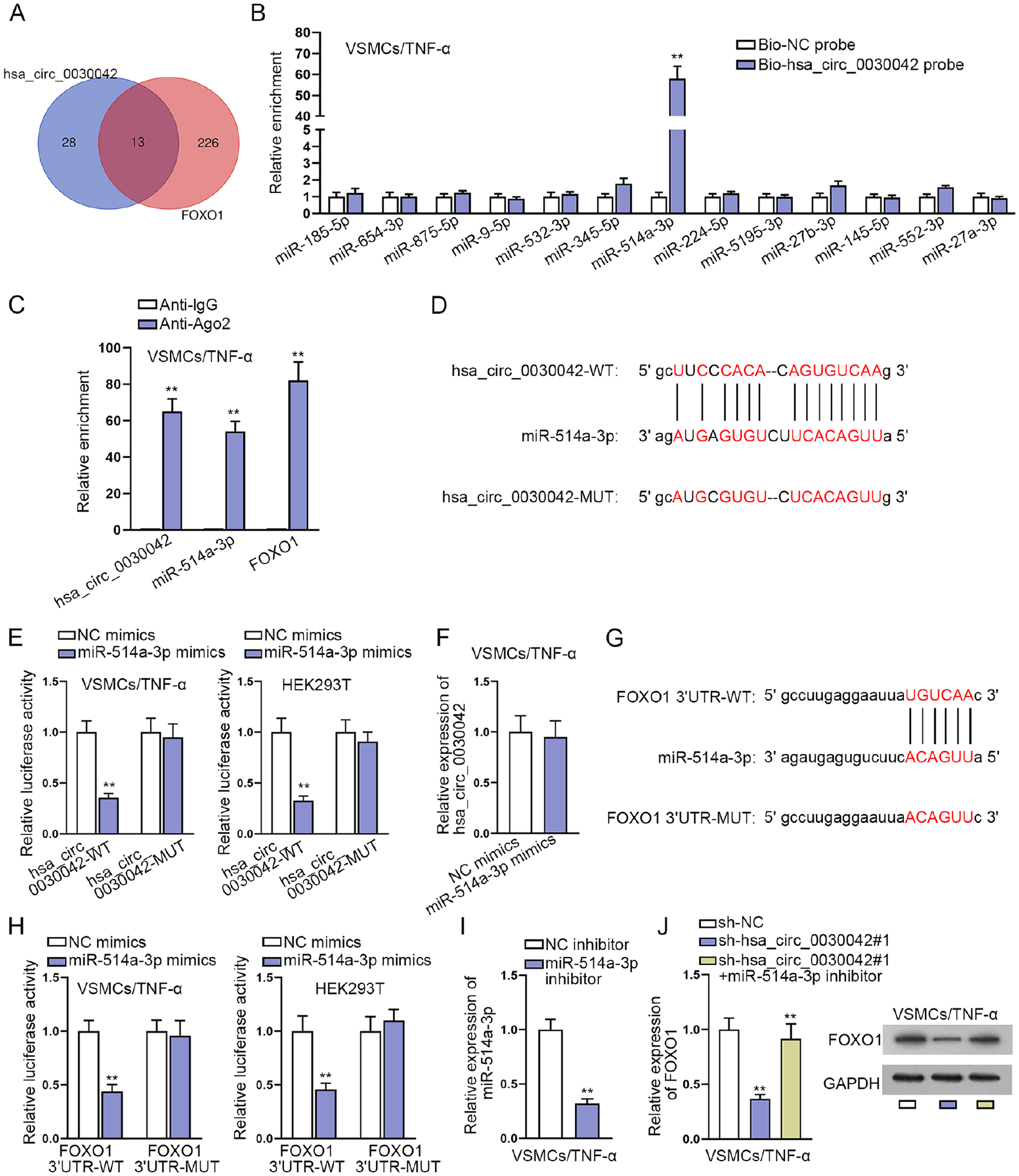
Hsa_circ_0030042 regulates FOXO1 expression via sponging miR-514a-3p: (A) Venn diagram showed the starBase prediction of the mutual target miRNAs of hsa_circ_0030042 and FOXO1, (B) RNA pull-down assay detected the combination of miRNAs and hsa_circ_0030042, (C) RIP assays detected the enrichment of hsa_circ_0030042, miR-514a-3p, and FOXO1 in Ago2, (D) binding sites between hsa_circ_0030042 and miR-514a-3p, (E) luciferase activities of hsa_circ_0030042-WT and hsa_circ_0030042-MUT were examined in TNF-α-induced VSMCs or HEK293T cells after miR-514a-3p overexpression, (F) RT-qPCR was performed to detect the expression of hsa_circ_0030042 after the overexpression of miR-514a-3p, (G) binding sites between FOXO1 3′ UTR and miR-514a-3p, (H) luciferase activities of FOXO1 3′ UTR-WT and FOXO1 3′ UTR-MUT were examined in TNF-α-induced VSMCs or HEK293T cells after miR-514a-3p overexpression, (I) silencing efficiency of miR-514a-3p inhibitor was assessed by RT-qPCR, and (J) the mRNA and protein levels of FOXO1 were measured in TNF-α-induced VSMCs transfected with sh-NC, sh-hsa_circ_0030042#1 and sh-hsa_circ_0030042#1+miR-514a-3p inhibitor using RT-qPCR and Western blot. Statistical significance was determined using the Student t test in B, C, E, F, H and I; 1-way ANOVA in J for RT-qPCR. miRNAs, microRNAs; FOXO1, forkhead box O1; RIP, RNA-binding protein immunoprecipitation; TNF-α, tumor necrosis factor-alpha; VSMCs, vascular smooth muscle cells; RT-qPCR, quantitative real-time polymerase chain reaction; 3′ UTR, 3′ untranslated region; ANOVA, analysis of variance.
Hsa_circ_0030042 Promotes TNF-α-Induced VSMC Proliferation, Migration and Inhibits Apoptosis via Targeting the miR-514a-3p/FOXO1 Axis
To further validate whether hsa_circ_0030042 promotes TNF-α-induced VSMC proliferation and migration and inhibits apoptosis via targeting the miR-514a-3p/FOXO1 axis, we upregulated FOXO1 expression (Figure 5A) and then severally transfected sh-NC, sh-hsa_circ_0030042#1, sh-hsa_circ_0030042#1+pcDNA3.1-FOXO1, and sh-hsa_circ_0030042#1+miR-514a-3p inhibitor into TNF-α-induced VSMCs for rescue assays. CCK-8 and EdU assays indicated that overexpression of FOXO1 or silence of miR-514a-3p offset the reduced proliferation caused by hsa_circ_0030042 silence in VSMCs after TNF-α treatment (Figure 5B and 5C). Moreover, the enhanced apoptosis in hsa_circ_0030042-depleted VSMCs was abolished after co-transfection of pcDNA3.1-FOXO1 or miR-514a-3p inhibitor upon TNF-α treatment (Figure 5D and 5E). In addition, when FOXO1 was upregulated or miR-514a-3p was downregulated, the lessened migration in TNF-α-induced VSMCs caused by hsa_circ_0030042 knockdown was restored (Figure 5F and 5G). Taken together, hsa_circ_0030042 promotes TNF-α-induced VSMC proliferation and migration and inhibits apoptosis via targeting the miR-514a-3p/FOXO1 axis.
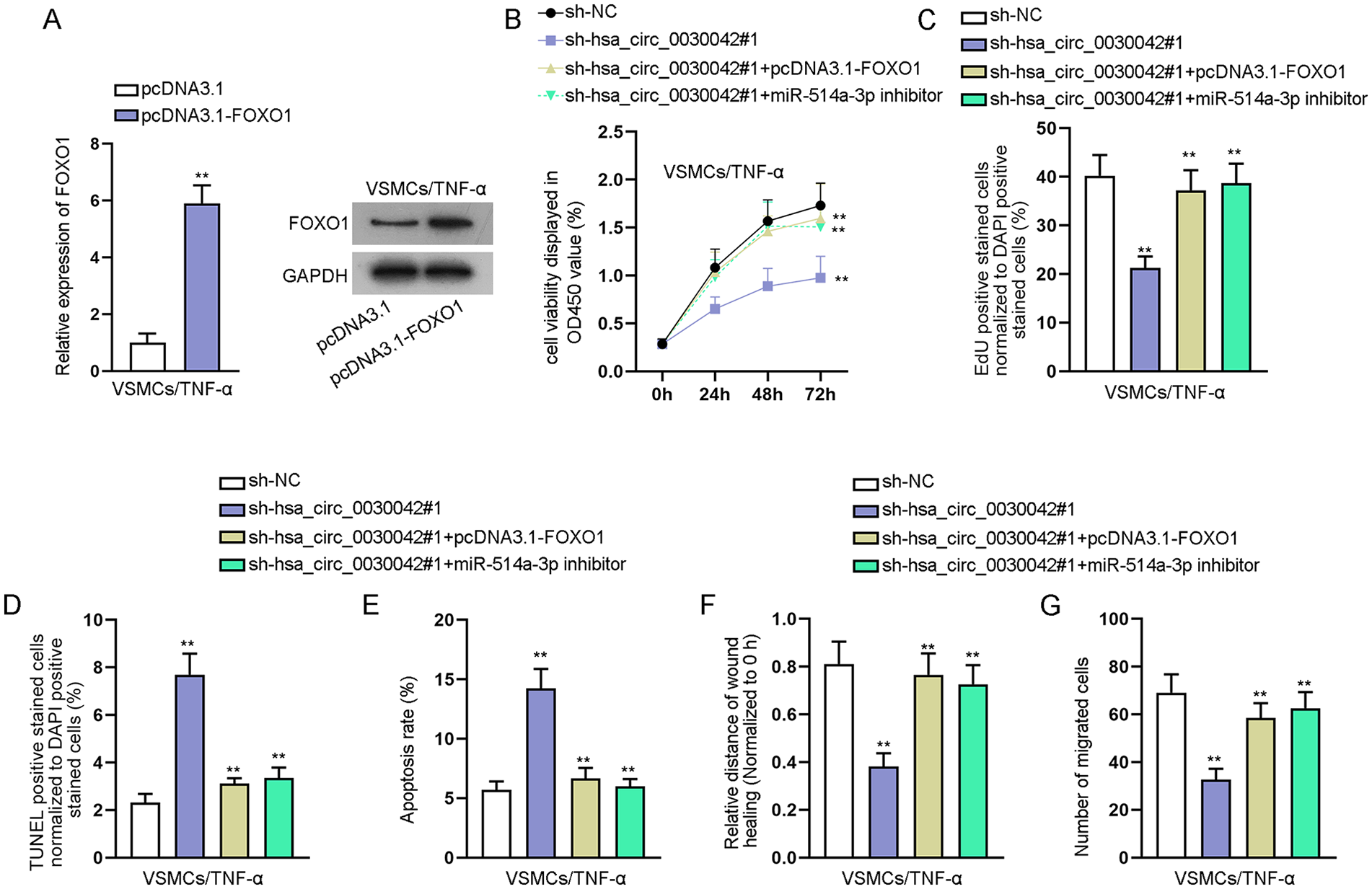
Hsa_circ_0030042 promotes TNF-α-induced VSMC proliferation and migration and inhibits apoptosis via targeting the miR-514a-3p/FOXO1 axis: (A) upregulation efficiency of pcDNA3.1-FOXO1 was detected in TNF-α-induced VSMCs using RT-qPCR and Western blot. Rescue assays were performed in TNF-α-induced VSMCs transfected with sh-NC, sh-hsa_circ_0030042#1, sh-hsa_circ_0030042#1+pcDNA3.1-FOXO1, and sh-hsa_circ_0030042#1+miR-514a-3p inhibitor, respectively, (B and C) CCK-8 and EdU assays detected proliferation, (D and E) apoptosis was examined by TUNEL assay and flow cytometry analysis, and (F and) migration was assessed by wound healing and transwell assays. Statistical significance was determined using the Student t test in A for RT-qPCR; 1-way ANOVA in B, C, D, E, F, and G. TNF-α, tumor necrosis factor-alpha; VSMCs, vascular smooth muscle cells; FOXO1, forkhead box O1; RT-qPCR, quantitative real-time polymerase chain reaction; CCK-8, cell counting kit-8; ANOVA, analysis of variance; TUNEL, terminal-deoxynucleoitidyl transferase–mediated nick end labeling.
Discussion
Atherosclerosis is a serious threat to human health, and the dysfunction of VSMCs has been documented to take part in the formation of AS. 20 In this study, VSMCs were treated with TNF-α to establish the cell model for AS according to previous literatures. 21 A previous study has revealed that circ-Sirt1, which is induced by TNF-α in VSMCs, inhibits the development of AS. 22 Similarly, our study also investigated the function of a circRNA in AS. We found that the expression of hsa_circ_0030042 was elevated in TNF-α-induced VSMCs. Knockdown of hsa_circ_0030042 restrained the proliferation and migration of TNF-α-induced VSMCs and promoted cell apoptosis. Taken together, these results suggested that hsa_circ_0030042 contributed to AS progression and might be a potential target for AS treatment.
Emerging evidence has pointed that circRNAs exert their functions by cooperating with their host genes. 23 The results of our study showed that hsa_circ_0030042 positively regulated its host gene FOXO1. Mohammad Babaee et al have discovered that FOXO expression is increased in patients with coronary artery disease, and its expression was significantly related to the severity of stenosis. 24 Moreover, Li Qiang and his colleague have proposed that increased FOXO1 expression plays an important role in AS. 25 In this study, we found that FOXO1 was upregulated in VSMCs after TNF-α treatment, and silencing of FOXO1 obviously inhibited VSMC proliferation and migration whereas it promoted apoptosis after TNF-α treatment.
CircRNAs can target miRNA to modulate mRNA expression. 26 To understand the potential mechanism of FOXO1 regulated by hsa_circ_0030042 in AS, the miRNAs of hsa_circ_0030042 and FOXO1 were predicted, and miR-514a-3p was finally identified to combine with hsa_circ_0030042 and FOXO1 simultaneously. MiR-514a-3p has been reported to function as a tumor suppressor in many cancers.27,28 In this study, we proved that overexpression of miR-514a-3p exerted repressive effects on the proliferation and migration of TNF-α-induced VSMCs. In-depth rescue experiments suggested that hsa_circ_0030042 influenced the progression of AS by targeting miR-514a-3p to upregulate FOXO1. Moreover, it is indicated by IF and Western blot analyses that hsa_circ_0030042 modulates NF-kB pathway through P65 translocation.
There were some limitations in this study. Previous studies have indicated that FOXO1 can be involved in the development of disease via regulating various signaling pathways, such as NF-κB pathway 29 and Wnt/β-catenin signaling. 30 We have conducted assays to explore the effect of hsa_circ_0030042 on NF-kB pathway. However, whether the hsa_circ_0030042/miR-514a-3p/FOXO1 axis regulates AS progression via this pathway requires to be further investigated. In addition, the role of the hsa_circ_0030042/miR-514a-3p/FOXO1 axis in AS in vivo has not been explored. These issues will become the focus of our future study.
In summary, our study illustrated the overexpression of hsa_circ_0030042 in TNF-α-induced VSMCs, and its aberrant expression facilitated the proliferation and migration. Mechanically, the interaction of hsa_circ_0030042, miR-514a-3p and FOXO1 was elucidated in detail. Collectively, our study might provide a profound understanding about hsa_circ_0030042 in AS.
Supplemental Material
sj-tif-1-jet-10.1177_15266028211057086 – Supplemental material for Hsa_circ_0030042 Facilitates the Proliferation and Migration of Vascular Smooth Muscle Cells via the miR-514a-3p/FOXO1 Axis
Supplemental material, sj-tif-1-jet-10.1177_15266028211057086 for Hsa_circ_0030042 Facilitates the Proliferation and Migration of Vascular Smooth Muscle Cells via the miR-514a-3p/FOXO1 Axis by Ji Ma, Jia Liu, Tengfei Li and Jianzhuang Ren in Journal of Endovascular Therapy
Supplemental Material
sj-tif-2-jet-10.1177_15266028211057086 – Supplemental material for Hsa_circ_0030042 Facilitates the Proliferation and Migration of Vascular Smooth Muscle Cells via the miR-514a-3p/FOXO1 Axis
Supplemental material, sj-tif-2-jet-10.1177_15266028211057086 for Hsa_circ_0030042 Facilitates the Proliferation and Migration of Vascular Smooth Muscle Cells via the miR-514a-3p/FOXO1 Axis by Ji Ma, Jia Liu, Tengfei Li and Jianzhuang Ren in Journal of Endovascular Therapy
Supplemental Material
sj-tif-3-jet-10.1177_15266028211057086 – Supplemental material for Hsa_circ_0030042 Facilitates the Proliferation and Migration of Vascular Smooth Muscle Cells via the miR-514a-3p/FOXO1 Axis
Supplemental material, sj-tif-3-jet-10.1177_15266028211057086 for Hsa_circ_0030042 Facilitates the Proliferation and Migration of Vascular Smooth Muscle Cells via the miR-514a-3p/FOXO1 Axis by Ji Ma, Jia Liu, Tengfei Li and Jianzhuang Ren in Journal of Endovascular Therapy
Supplemental Material
sj-tif-4-jet-10.1177_15266028211057086 – Supplemental material for Hsa_circ_0030042 Facilitates the Proliferation and Migration of Vascular Smooth Muscle Cells via the miR-514a-3p/FOXO1 Axis
Supplemental material, sj-tif-4-jet-10.1177_15266028211057086 for Hsa_circ_0030042 Facilitates the Proliferation and Migration of Vascular Smooth Muscle Cells via the miR-514a-3p/FOXO1 Axis by Ji Ma, Jia Liu, Tengfei Li and Jianzhuang Ren in Journal of Endovascular Therapy
Footnotes
Acknowledgements
We appreciate for all authors.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.