
Abstract
Objective:
This study aimed to explore the therapeutic effect of resveratrol (RES) against abdominal aortic aneurysm (AAA) and the role of HMOX1 underlying this effect.
Methods:
Vascular smooth muscle cells (VSMCs) were induced by angiotensin II (Ang II) to construct the microenvironment of AAA. HMOX1 expression was downregulated by the short hairpin ribonucleic acid (RNA) specific to HMOX1 in RES-pretreated VSMCs. The levels of matrix metalloproteinase (MMP)-2, MMP-9, and elastin were measured by quantitative reverse transcription polymerase chain reaction (qRT-PCR) and western blot. Apoptosis rate was detected. The levels of apoptosis-related proteins (caspase-3 and Bax/Bcl-2), inflammatory cytokines (interleukin [IL]-6, tumor necrosis factor [TNF]-α, and IL-1β), and autophagy-related proteins (Beclin 1, light chain 3 [LC3] II/I, and p62) were detected by western blot. The secretion of inflammatory factors in cell supernatant was detected by enzyme-linked immunosorbent assay (ELISA). The number of autophagic vesicles in VSMCs was observed and analyzed by transmission electron microscopy. A rat model of pancreatic elastase-induced AAA was established to verify the effect and action mechanism of RES.
Results:
Stimulation of Ang II increased the messenger RNA (mRNA) and protein levels of MMP-2 and MMP-9, decreased elastin expression, and enhanced apoptosis, secretion of inflammatory factors, and autophagy in VSMCs, whereas RES pretreatment ameliorated Ang II-induced VSMC dysfunction. In addition, HMOX1 mRNA and heme oxygenase-1 (HO-1) protein levels were significantly increased in VSMCs pretreated with RES compared with Ang II treatment alone. Silencing of HMOX1 abolished the effects of RES on VSMC dysfunction. Consistently, RES suppressed the development of AAA in rats by increasing the expression of HMOX1.
Conclusion:
Resveratrol protects against AAA by inhibiting extracellular matrix degradation, apoptosis, autophagy, and inflammation of VSMCs via HMOX1 upregulation.
Clinical Impact
Our study found that angiotensin II (Ang II) stimulated increased the levels of MMP-2 and MMP-9 in vascular smooth muscle cells (VSMCs), decreased elastin expression, and promoted apoptosis, autophagy occurrence, and secretion of inflammatory factors, while resveratrol (RES) pretreatment improved this effect. In addition, downregulation of HMOX1 expression eliminated the effect of RES on the function of VSMCs. Our study elucidates that RES improves AAA progression through HMOX1 at both cellular and animal levels. This work can help doctors better understand the pathological mechanism of the occurrence and development of AAA, and provide a theoretical basis for clinicians to find better treatment options.
Introduction
Abdominal aortic aneurysm is a pathologic aortic dilatation defined by a maximal infrarenal abdominal aortic diameter greater than 3 cm, or at least 1.5 times the diameter of the adjacent normal artery.1–3 Abdominal aortic aneurysm is typically asymptomatic but at increased risk of aortic rupture. 4 Accumulative documents have revealed that the dysfunction of VSMCs is evidenced as a pathological feature in AAA, and VSMC apoptosis can elevate elastin degradation and AAA formation. 5 The phenotypic change of VSMCs from a contractile to a pro-inflammatory/promatrix remodeling phenotype in the aneurysm is associated with autophagy, but the exact beneficial or deleterious role of autophagy has not been identified. 6 Histopathologically, aneurysm tissue also exhibits degradation of extracellular matrix (ECM), which regulates the proliferation and metabolism of VSMCs by imparting mechanical cues to the VSMCs. 7 Matrix metalloproteinases are intrinsically capable of degrading ECM components and remodeling ECM and could be synthesized and released into the bloodstream by VSMCs, macrophages, and fibroblasts. 8 However, the molecular mechanisms behind the development of VSMC injury and ECM degradation in AAA are largely undefined.
Heme oxygenase-1 (gene HMOX1, protein HO-1), a stress-induced protein, can catalyze rate-limiting reactions in heme catabolism, thereby producing biliverdin, carbon monoxide, and iron with powerful antioxidant, anti-inflammatory, and signaling properties.9,10 Also, it has protective effects in the vessel wall. 11 As suggested by Qiu et al, betanin reduced the levels of tissue-reactive oxygen species by stimulating the nuclear factor-E2-related factor 2/HO-1 pathway, thus playing a preventive effect on AAA. 12 Reportedly, HO-1 induced autophagy and protected against IL-1β-mediated apoptosis in human nucleus pulposus cells by inhibiting nuclear factor kappa-B (NF-κB). 13 A previous study showed that HO-1 induction by heme slows down AAA progression in the porcine pancreatic elastase murine AAA models. 14 In vivo, mice lacking HO-1 had severe aortic dilatation in response to Ang II-induced AAA, along with enhanced macrophage infiltration, increased elastin degradation, and elevated MMP-9 level in the aneurysmal aortic wall. 15 Also, HMOX1 is a cytoprotective molecule that contributes to cellular homeostasis, and loss of HMOX1 results in increased inflammation and aggravated neuronal damage in subarachnoid hemorrhage mouse models. 16 Thus, searching for a candidate drug that can activate HMOX1/HO-1 may be beneficial for AAA treatment.
Resveratrol, 3,5,4′-trihydroxy-trans-stilbene, is a natural polyphenol found in the skin of plants including grapes and berries. 17 Reportedly, RES possesses vital biological activities including apoptosis induction, autophagy, anti-inflammation, 18 and anti-cancer. 19 Recent evidence revealed that RES had a protective effect in AAA rupture-induced liver injury by repressing oxidative stress and apoptosis in hepatocytes. 20 In high-fat chow-fed mice supplying with continuous Ang II, dietary RES supplementation substantially restrained suprarenal aorta expansion. 21 Moreover, RES has been shown to prevent AAA development in rat models through attenuation of macrophage infiltration, neovascularization, and oxidative stress.22,23 Hibender S et al pointed out RES repressed aortic root dilatation in Marfan syndrome mice by mechanically downregulating the aneurysm-related microRNA-29b expression in the aorta. 24 Another study revealed that RES-initiated SIRT1 activation could relieve β-aminopropionitrile monofumarate-induced acute aortic dissection symptoms. 25 On the basis of these data, RES was proved to be closely associated with AAA. Previously, RES was confirmed to protect against ischemia-induced renal injury by downregulating MMP-9 and upregulating HO-1. 26 Increased HO-1 expression was associated with RES-mediated protection in Aβ-induced neurotoxicity. 27 Therefore, this work attempted to investigate the role and underlying mechanisms of RES in AAA in relation to HMOX1/HO-1.
Materials and Methods
Cell Culture
Vascular smooth muscle cells from human aorta were acquired from American Type Culture Collection (Rockville, MD). The obtained cells were fostered in Dulbecco’s modified Eagle’s medium (HyClone, Invitrogen, Camarillo, CA) containing 10% fetal bovine serum (FBS; HyClone) at 37°C in a humidified incubator with 5% CO2.
Cell Transfection
Cell transfection was performed using Lipofectamine 2000 kit (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Vascular smooth muscle cells were separately treated with sh-HMOX1, pcDNA3.1-HMOX1, sh-NC, and pcDNA3.1-NC. The plasmids at a final concentration of 2 μg were used for the transfection. The VSMCs were pretreated with 10 μM of RES (Sigma-Aldrich, St. Louis, MO, USA) for 24 hours before the transfection. After transfection for 48 hours, the cells were stimulated with 50 nM of Ang II (Sigma-Aldrich) for 48 hours. In addition, the cells did not undergo RES treatment during transfection and Ang II stimulation.
Quantitative Reverse Transcription Polymerase Chain Reaction
Total RNA was extracted using TRIzol reagent (Invitrogen). Reverse transcription reactions were performed using the reverse transcription kit (TaKaRa, Tokyo, Japan), and all operations were performed following the manufacturer’s protocol. Gene expression was detected by the LightCycler 480 fluorescence quantitative polymerase chain reaction (qPCR) instrument (Roche, Indianapolis, IN), and reaction condition was performed using the fluorescent qRT-PCR kit (SYBR Green Mix, Roche). The thermocycling program included 5 minutes at 95°C; 45 cycles of 5 seconds at 95°C, 10 seconds at 60°C, and 10 seconds at 72°C; and 5 minutes at 72°C. Each reaction of qPCR was performed in triplicate. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the internal reference. Data were calculated using 2−ΔΔCt method, and the formula was as follows: ΔΔCt = (Ct [target gene] - Ct [reference gene]) experimental group - (Ct [target gene] - Ct [reference gene]) control group. The primer sequences for all target genes are shown in Table 1.
Primer Sequences for qRT-PCR to Determine the Expression Levels of HMOX1, MMP-2, MMP-9, and Elastin.
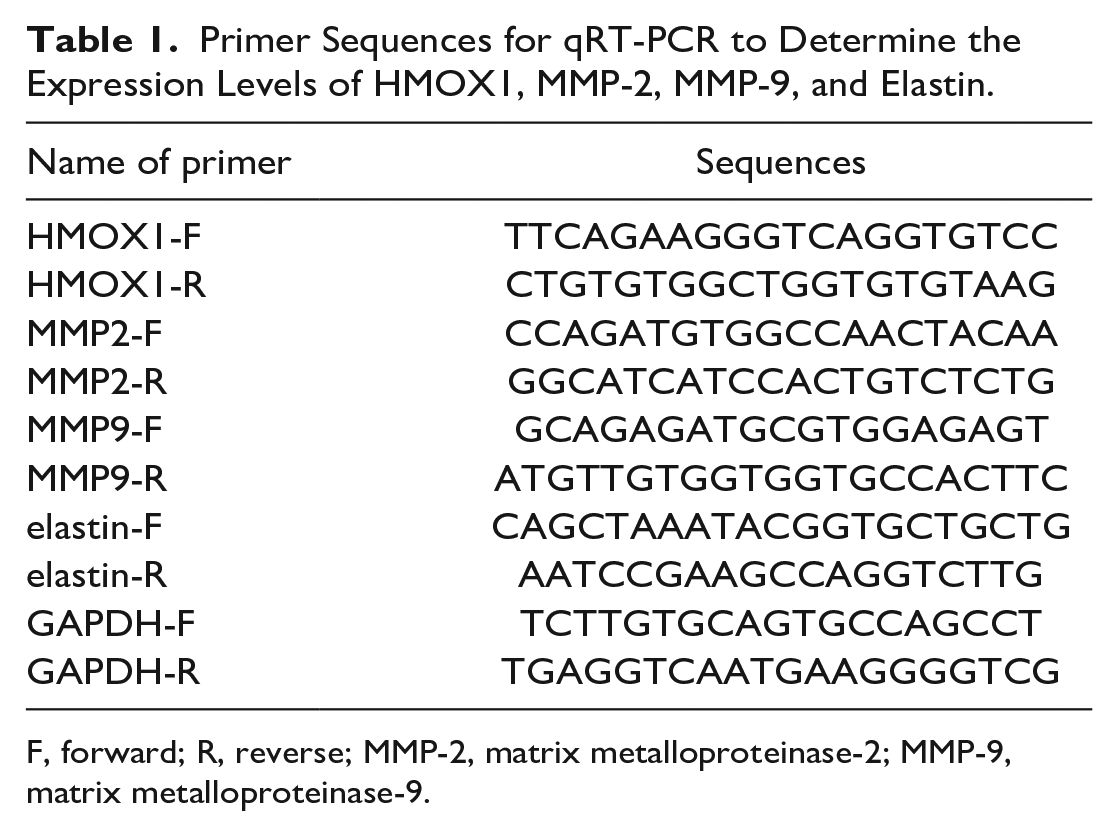
F, forward; R, reverse; MMP-2, matrix metalloproteinase-2; MMP-9, matrix metalloproteinase-9.
Western Blot
Vascular smooth muscle cells were lysed by radio-immunoprecipitation assay (RIPA) buffer to obtain protein samples. Thereafter, protein concentration was detected by a bicinchoninic acid assay (BCA) kit, after which the corresponding volume of protein was mixed into the loading buffer and then heated in boiling water for 3 minutes for protein degeneration. The RIPA buffer, BCA kit, and loading buffer were purchased from Beyotime Institute of Biotechnology. Electrophoresis was firstly conducted at 80 V for 30 minutes and then at 120 V for 1 to 2 hours. The protein was transferred onto membranes for 60 minutes at 300 mA in an ice bath. Afterward, the membrane was rinsed for 1 to 2 minutes and then blocked at room temperature for 60 minutes, or at 4°C overnight. Primary antibody of GAPDH (ab9485, 1:5000), HO-1 (ab13243, 1:2000), MMP-2 (ab97779, 1:2000), MMP-9 (ab38898, 1:1000), caspase-3 (ab32351, 1:4000), Bax (ab104156, 1:1000), Bcl-2 (ab196495, 1:2000), IL-6 (ab6672, 1:1000), TNF-α (ab6671, 1:1000), IL-1β (ab9722, 1:2000), elastin (ab217356, 1:500), light chain (LC3, (ab128025, 1:1000), Beclin1 (ab62557, 1:2000) (all from Abcam, Cambridge, UK), or P62 (#88588, 1:1000, Cell Signaling Technology, Beverly, MA, USA) was cultured on a shaker for 2 hours (room temperature) before washing 3 times for 10 minutes each. Then, the membranes were incubated with secondary antibody (horseradish peroxidase-labeled goat anti-rabbit, 1:5000; Beijing ComWin Biotech Co., Ltd., Beijing, China) for 1 hour (room temperature) followed by washings (3 × 10 min). Last, the membrane was visualized using chemiluminescence detection and imaging system (Bio-Rad Laboratories).
Cell counting kit-8 (CCK-8) Assay
Cell suspensions (100 μL) were separately seeded into the 96-well plates with 3 replicates per group. After the cells were cultured for 48 hours, 10 μL of CCK-8 solution (Dojindo Molecular Technology, Tokyo, Japan) was added to each well. Finally, the absorbance was determined at 450 nm after the cells were cultured for another 1 to 4 hours.
Flow Cytometry
Cells of each group were prepared into single-cell suspensions and centrifuged at 2000 r/minutes before twice washing in phosphate-buffered saline (PBS). The cells were resuspended in binding buffer, and 5 μL of Annexin-V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) were mixed into 195 μL cell suspension (containing 105 cells). After 10 minutes of incubation away from light, cell apoptosis was measured by flow cytometry (FCM) on a fluorescence activated cell sorting (FACS) Canto II 488N cytometer (BD Biosciences, USA). Each experiment was repeated 3 times.
Transmission Electron Microscopy
After washing with PBS 3 times, the cells were digested with 0.5% trypsin and then transferred into a 1.5-mL microcentrifuge tube. Thereafter, the cells were treated with 2.5% glutaraldehyde in PBS and then fixed with 1% osmium tetroxide, followed by dehydration with a graded series of ethanol (30%–90%) and embedding in 812 epoxy resin. Ultrathin sections (80 nm) were stained with 2% uranyl acetate and lead citrate, which were observed using an H-7650 transmission electron microscopy (TEM) (Hitachi, Japan).
Enzyme-Linked Immunosorbent Assay
The levels of proinflammatory cytokines (IL-6, TNF-α, and IL-1β) in the supernatants were measured using ELISA kits (Abcam) according to the manufacturer’s instructions. Malondialdehyde level in rat aortic tissue was detected using Malondialdehyde (MDA) kit (Abcam).
Immunofluorescence Staining
Cells were cultured on cover slides and fixed in 4% paraformaldehyde at room temperature, after which the cells were permeabilized with 0.5% Triton X100 (Shanghai Solarbio Bioscience & Technology Co., Ltd., Shanghai, China) for 15 minutes, and incubated with 3% BSA for 15 minutes for antigen blocking. Heme oxygenase-1 antibody (ab13243, 1:200, Abcam) was incubated with the cells at 4°C overnight before washing with PBS 3 times. Thereafter, the cells were incubated with Alexa Fluor488 Goat-anti-Rabbit IgG (H + L) (ab150077, 1:200, Abcam) at 37°C for 1 hour. The sections were visualized using a Leica DM3000 microscope (Leica Microsystems GmbH).
Induction of Abdominal Aortic Aneurysm in Rats
Forty healthy male Sprague-Dawley rats (weighing 250–300 g) were purchased from the Department of Animal Experimentation, Central South University. All rats were housed in individual cages, under the supervision of special personnel, with 12/12-hour day-night cycles and free access to water and food. All animal experiments in this study were carried out in accordance with the “Guidelines for the Care and Use of Laboratory Animals,” under the approval of the Animal Ethics Committee of the First Affiliated Hospital of Chongqing Medical University.
Rats were divided into sham, AAA, RES, and RES + sh-HMOX1 groups. Rats in the sham and AAA groups were given normal saline. For the RES group, 10 mg/kg RES was administered through an intraperitoneal route every day from the day of AAA operation for 14 days. For the RES + sh-HMOX1 group, HMOX1-interfering adenovirus (titer: 109 Pfu/mL; Hanbio, Shanghai, China) was injected via tail vein on the day of AAA surgery, and daily RES (10 mg/kg) was administered intraperitoneally from the day of AAA operation for 14 days. Abdominal aortic aneurysm was induced in rats by peri-aortic application of pancreatic elastase as previously described. 9 Briefly, rats were anesthetized with isoflurane and fixed on a 37°C thermostatic plate. The lower abdomen was bluntly dissected along the midline to expose the abdominal aorta (about 1 cm in length) between the renal arteries and the iliac arteries. The abdominal aorta was separated from other tissues and wrapped with a polyethylene sponge cloth previously immersed in 30 μL of elastase solution (elastase in saline). After 40 minutes, the sponge cloth was removed, and the incision was sutured. The sham group was treated with a saline-soaked sponge cloth, and other steps were the same as in the model group. After 14 days, the incidence of AAA in each group was observed, and the diameter expansion rate of abdominal aorta was calculated.
Statistical Analysis
Statistical analysis was analyzed by GraphPad Prism 8.0 software. Data are presented as mean ± standard deviation. Two sets of data were evaluated by t test, while the differences among multiple groups were detected by 1-way analysis of variance (ANOVA), followed by post hoc analysis with Tukey’s multiple comparison test. p<0.05 was considered to indicate a statistically significant difference.
Results
Resveratrol Represses Extracellular Matrix Degradation of Vascular Smooth Muscle Cells
To explore the role of RES in AAA progression, we investigated the effects of RES on ECM degradation, apoptosis, autophagy, and inflammation of VSMCs with VSMCs as research subjects. Vascular smooth muscle cells were pretreated with various concentrations of RES (0, 10, 20, or 40 µm) for 24 hours after which cell viability was detected using CCK-8 assay to determine the appropriate concentration of RES for VSMC treatment. The results showed significant cytotoxic effects of RES on VSMCs at 20 and 40 µM concentrations compared to 0 µM treatment (Figure 1A). Therefore, 10 µM RES was selected for subsequent experiments.
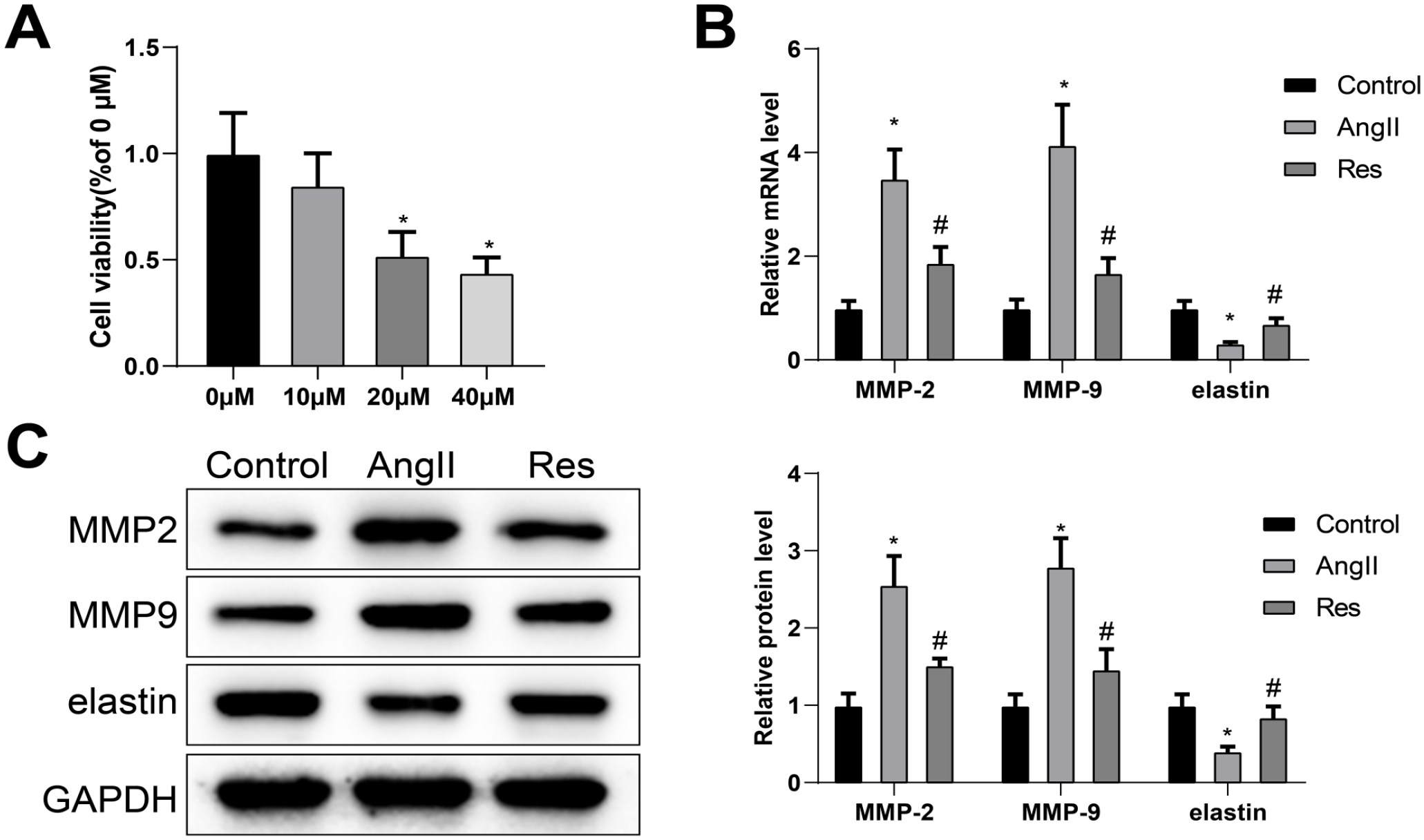
RES hinders ECM degradation of VSMCs. Cell viability was determined by CCK-8 assay (A). Differential expression of MMP-2, MMP-9, and elastin was measured by qRT-PCR and western blot analyses (B) and (C). Statistical significance of differences among multiple groups was determined using 1-way ANOVA and post hoc analysis with Tukey’s test. Each experiment was repeated 3 times; *p<0.05, vs μm or control group; #p<0.05, vs Ang II group. RES, resveratrol; ECM, extracellular matrix; VSMCs, vascular smooth muscle cells; MMP-2, matrix metalloproteinase-2; MMP-9, matrix metalloproteinase-9; Ang II, angiotensin II; ANOVA, analysis of variance; qRT-PCR, quantitative reverse transcription polymerase chain reaction.
After pretreatment with 10 µM RES for 24 hours, VSMCs were stimulated by Ang II for 48 hours to establish an in vitro aneurysm microenvironment. As shown in Figure 1B and C, compared with the control group, Ang II stimulation effectively promoted messenger RNA (mRNA) and protein levels of MMP-2 and MMP-9, while obviously hindered the levels of elastin in VSMCs. Also, analyses of qRT-PCR and western blot identified that the combination of Ang II and RES worked synergistically in reducing levels of MMP-2 and MMP-9 and increasing levels of elastin compared with VSMCs stimulated with Ang II alone, suggesting that RES treatment suppressed ECM degradation of VSMCs.
Resveratrol Suppresses Angiotensin II-Induced Vascular Smooth Muscle Cell Apoptosis and Inflammation
To investigate whether RES inhibits AAA progression by affecting VSMC function, we analyzed the effects of RES on Ang II-induced apoptosis and inflammation of VSMCs. Flow cytometry showed that the apoptosis rate was facilitated significantly in the Ang II group (vs control group) while refrained considerably in the RES group (vs Ang II group) (Figure 2A). Furthermore, western blot analysis showed that Ang II stimulation significantly promoted the protein levels of caspase-3 and Bax/Bcl-2, while RES reversed the effect of Ang II on the levels of caspase-3 and Bax/Bcl-2 (Figure 2B).
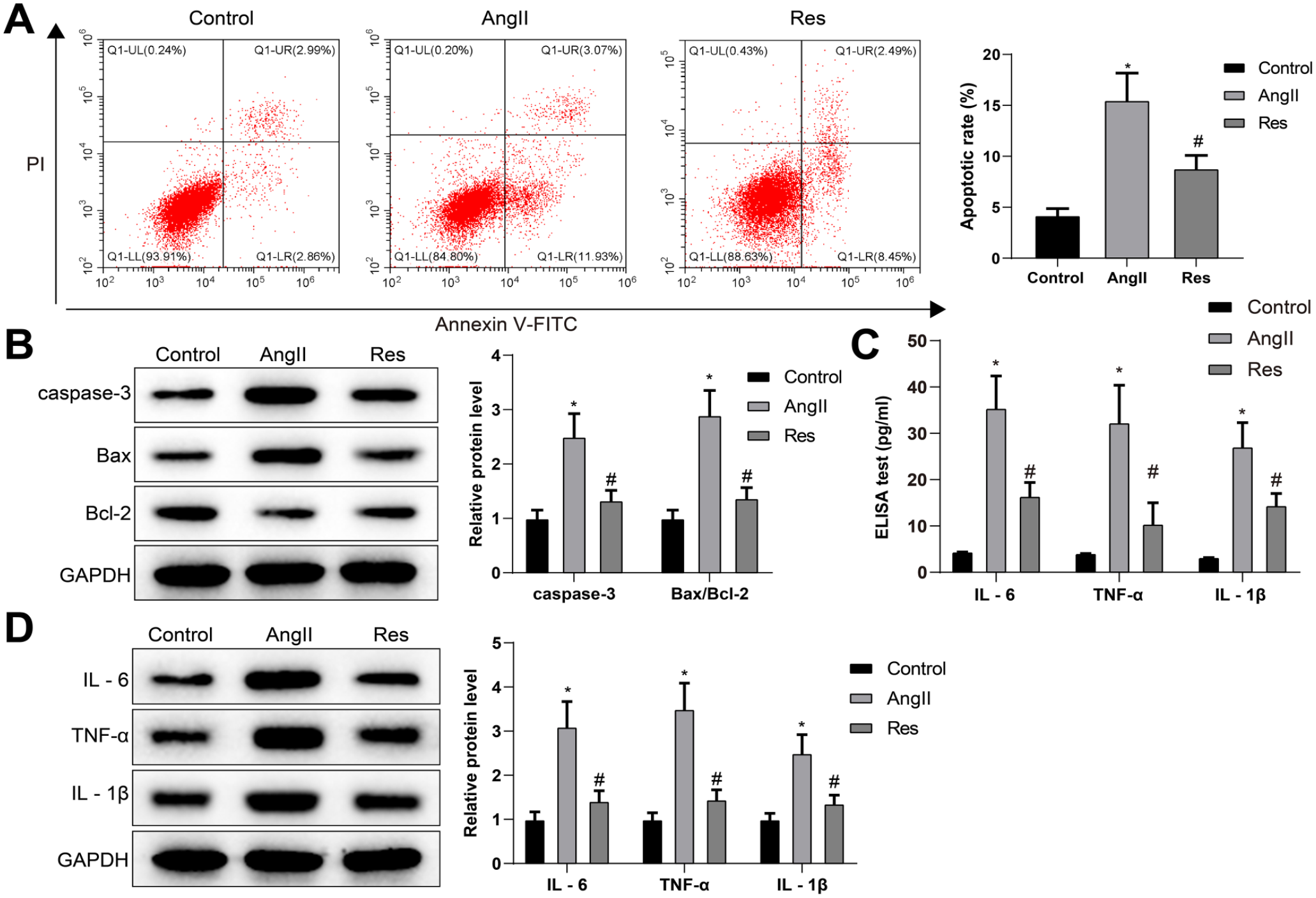
RES inhibits Ang II-induced VSMC apoptosis and inflammation. The apoptosis rate was determined by FCM (A). Expression levels of caspase-3 and Bax/Bcl-2 were evaluated by western blot (B). The levels of IL-6, TNF-α, and IL-1β were determined by ELISA (C) and western blot (D) analysis. One-way ANOVA and Tukey’s multiple comparison test were used to analyze the data among multiple groups. Each experiment was repeated 3 times; *p<0.05, vs control group; #p<0.05, vs Ang II group; RES, resveratrol; Ang II, angiotensin II; VSMC, vascular smooth muscle cell; FCM, flow cytometry; ANOVA, analysis of variance; ELISA, enzyme-linked immunosorbent assay; TNF, tumor necrosis factor; IL, interleukin; V-FITC, V-fluorescein isothiocyanate.
Then, the secretion of inflammatory cytokines in supernatants was examined by ELISA (Figure 2C) and confirmed by western blot analysis (Figure 2D). The results noted that Ang II group had augmented levels of inflammatory factors (IL-6, TNF-α, and IL-1β) in VSMCs (vs control group), but RES group had reduced levels of the inflammatory factors (vs Ang II group). Together, these data suggested that RES can inhibit Ang II-induced apoptosis and inflammatory responses of VSMCs.
Resveratrol Facilitates HMOX1 Expression in Vascular Smooth Muscle Cells
We next deciphered the effects of HMOX1 mRNA expression in VSMC of each group by qRT-PCR and HO-1 expression by western blot. The results showed that HMOX1 mRNA level and HO-1 protein level were significantly dropped in the Ang II group (vs control group), while RES treatment strikingly reversed the effect of Ang II on HMOX1 and HO-1 levels (Figure 3A and B). Also, the effect of RES on HO-1 expression was confirmed by immunofluorescence staining (Figure 3C).
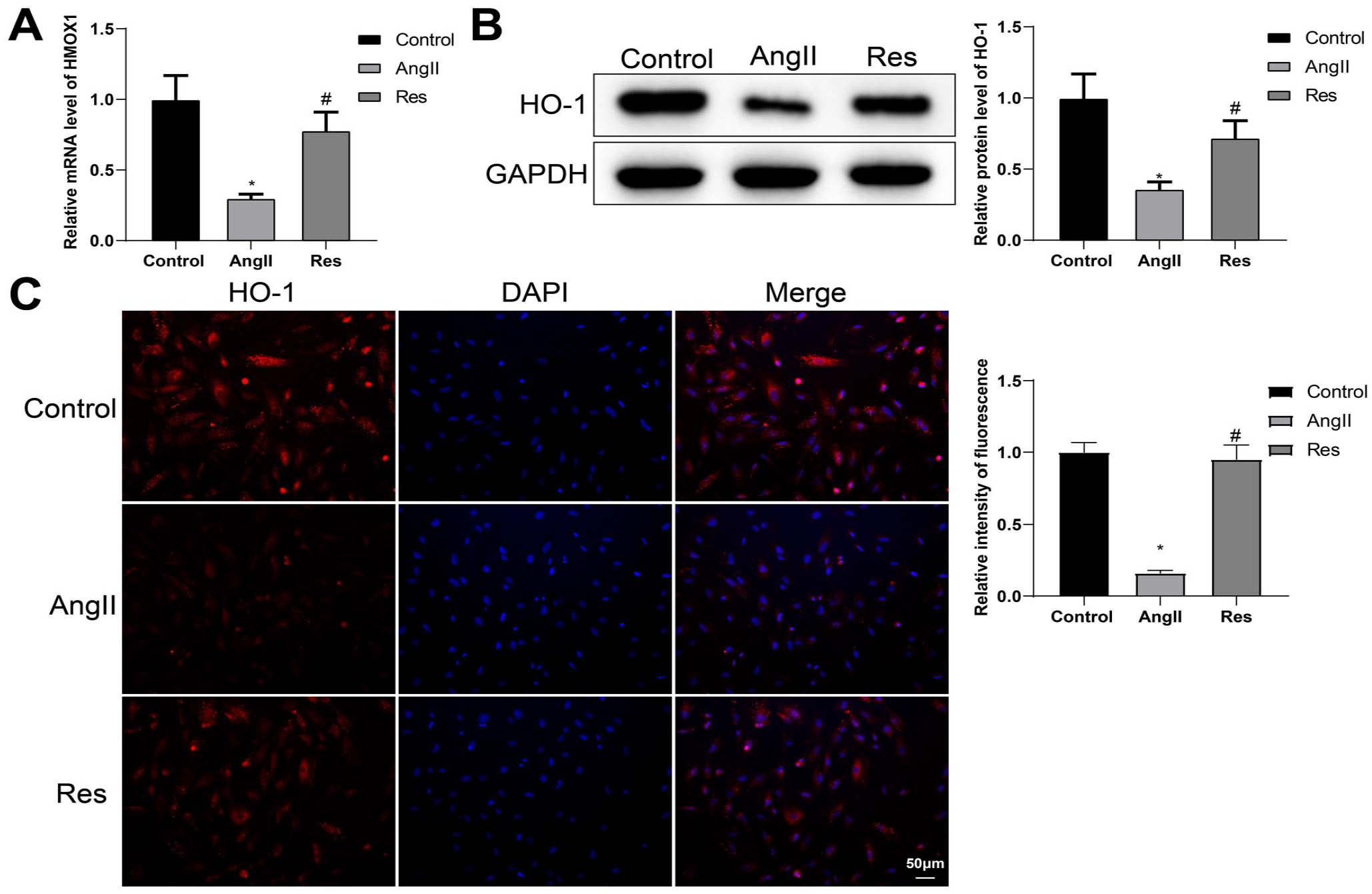
RES promotes HMOX1 expression in VSMCs. qRT-PCR was applied to determine the mRNA of HMOX1 (A). The protein level of HO-1 was assessed by western blot (B). HO-1 expression was evaluated by immunofluorescence staining (C). Differences among multiple groups were analyzed using 1-way ANOVA and post hoc analysis with Tukey’s test. Experiments were repeated in triplicate; *p<0.05, vs control group; #p<0.05, vs Ang II group; RES, resveratrol; VSMCs, vascular smooth muscle cells; HO-1, heme oxygenase-1; Ang II, angiotensin II; ANOVA, analysis of variance; qRT-PCR, quantitative reverse transcription polymerase chain reaction.
HMOX1 Regulates Vascular Smooth Muscle Cell Extracellular Matrix Degradation, Apoptosis, Autophagy, and Inflammation
Vascular smooth muscle cells were transfected with sh-HMOX1 or pcDNA3.1-HMOX1 before Ang II treatment to investigate the function of HMOX1 in VSMCs. The results illuminated that mRNA level of HMOX1 and protein level of HO-1 were dampened in the sh-HMOX1 group (vs sh-NC group), but enhanced in the pcDNA3.1-HMOX1 group (vs pcDNA3.1-NC group), indicative of satisfactory efficiency of sh-HMOX1 and pcDNA3.1-HMOX1 (Figure 4A and B).
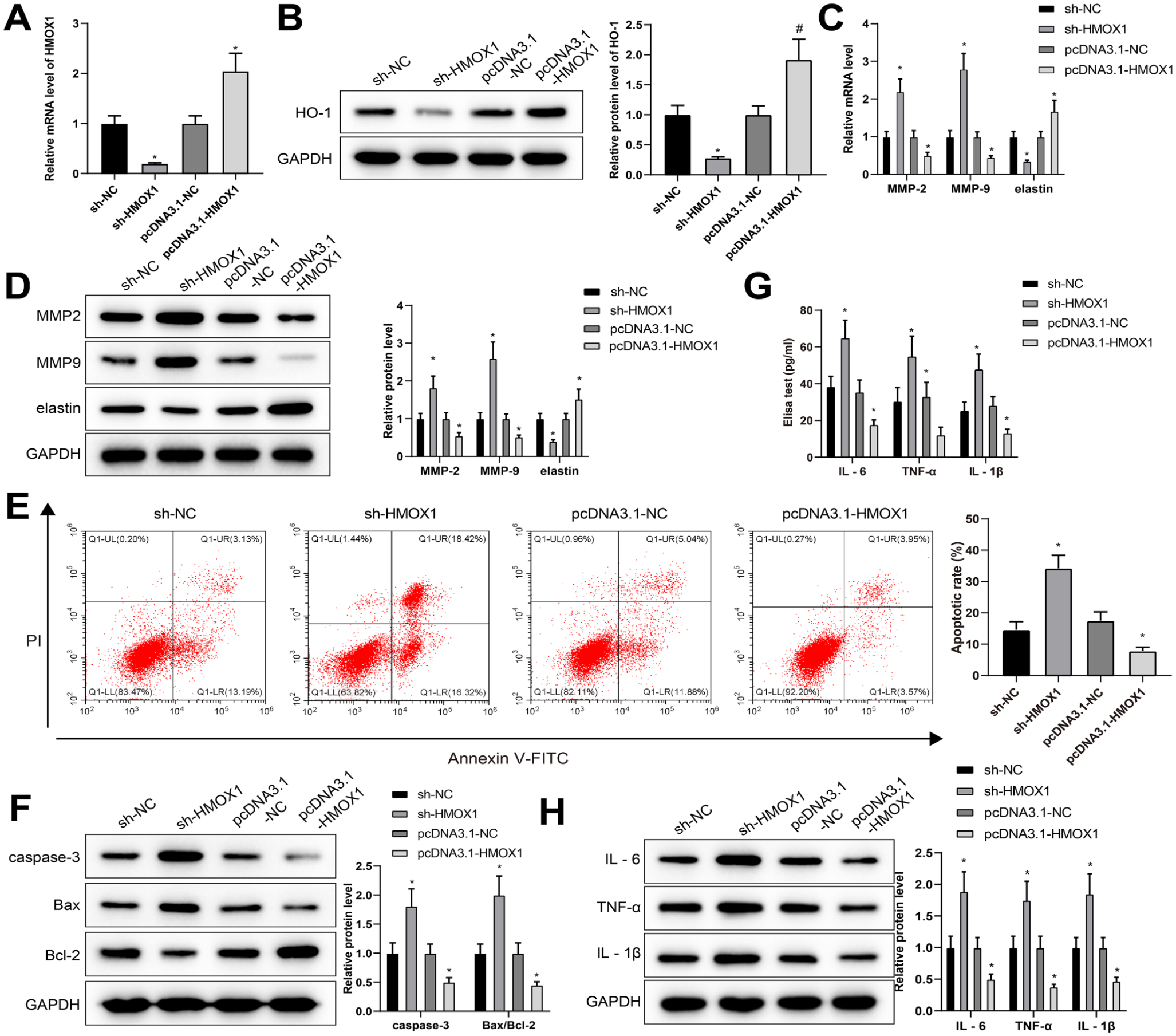
HMOX1 regulates VSMC ECM degradation, apoptosis, autophagy, and inflammation. After VSMCs were transfected with sh-HMOX1 and then treated with Ang II, the mRNA level of HMOX1 and protein level of HO-1 were separately detected by qRT-PCR and western blot (A) and (B). The levels of MMP-2, MMP-9, and elastin were measured by qRT-PCR and western blot (C) and (D). The apoptosis rate was detected by FCM (E). Western blot was used to detect the protein levels of caspase-3 and Bax/Bcl-2 (F). The levels of IL-6, TNF-α, and IL-1β were monitored by ELISA (G) and western blot (H) analyses. Differences among multiple groups were determined with 1-way ANOVA and post hoc analysis with Tukey’s test. Each experiment was repeated 3 times; *p<0.05, vs sh-NC group or pcDNA3.1-NC group; VSMC, vascular smooth muscle cell; Ang II, angiotensin II; HO-1, heme oxygenase-1; FCM, flow cytometry; ANOVA, analysis of variance; MMPs, matrix metalloproteinases; ECM, extracellular matrix; ELISA, enzyme-linked immunosorbent assay; qRT-PCR, quantitative reverse transcription polymerase chain reaction; TNF, tumor necrosis factor; IL, interleukin.
The detection of qRT-PCR and western blot described that the mRNA and protein levels of MMP-2 and MMP-9 in VSMCs treated with Ang II after HMOX1 knockdown were facilitated significantly, while the levels of elastin were refrained considerably (Figure 4C and D). Furthermore, FCM and western blot showed that apoptosis (Figure 4E) and the protein levels of caspase-3 and Bax/Bcl-2 (Figure 4F) were significantly increased in the sh-HMOX1 group (vs sh-NC group). Also, ELISA showed that downregulation of HMOX1 promoted inflammatory responses (Figure 4G), and the results were confirmed by western blot analysis (Figure 4H). On the contrary, enforced HMOX1 expression significantly inhibited Ang II-induced VSMC dysfunction (vs pcDNA3.1-NC group) (Figure 4C–H). These findings indicated that HMOX1 regulated ECM degradation, apoptosis, autophagy, and inflammation of VSMCs.
Resveratrol Inhibits Angiotensin II-Induced Autophagy in Vascular Smooth Muscle Cell
We next investigated whether RES plays a role in VSMC autophagy during AAA. Western blot was performed to test the expression of autophagy markers LC3 II/I, Beclin1, and p62. The detection showed that Beclin1 and LC3 II/I were overexpressed while p62 was downregulated in the Ang II group (vs control group). Reversed expression patterns were found in the RES group (vs Ang II group) (Figure 5A), indicating RES treatment significantly reduced the level of autophagy. These data were confirmed by TEM analysis showing an enhanced number of autophagic vesicles in VSMCs of Ang II group compared with the control group, while the declined number of autophagic vesicles was noted in the RES group (vs Ang II group) (Figure 5B). The aforementioned information suggested that RES may regulate Ang II-induced VSMC autophagy in the context of AAA.
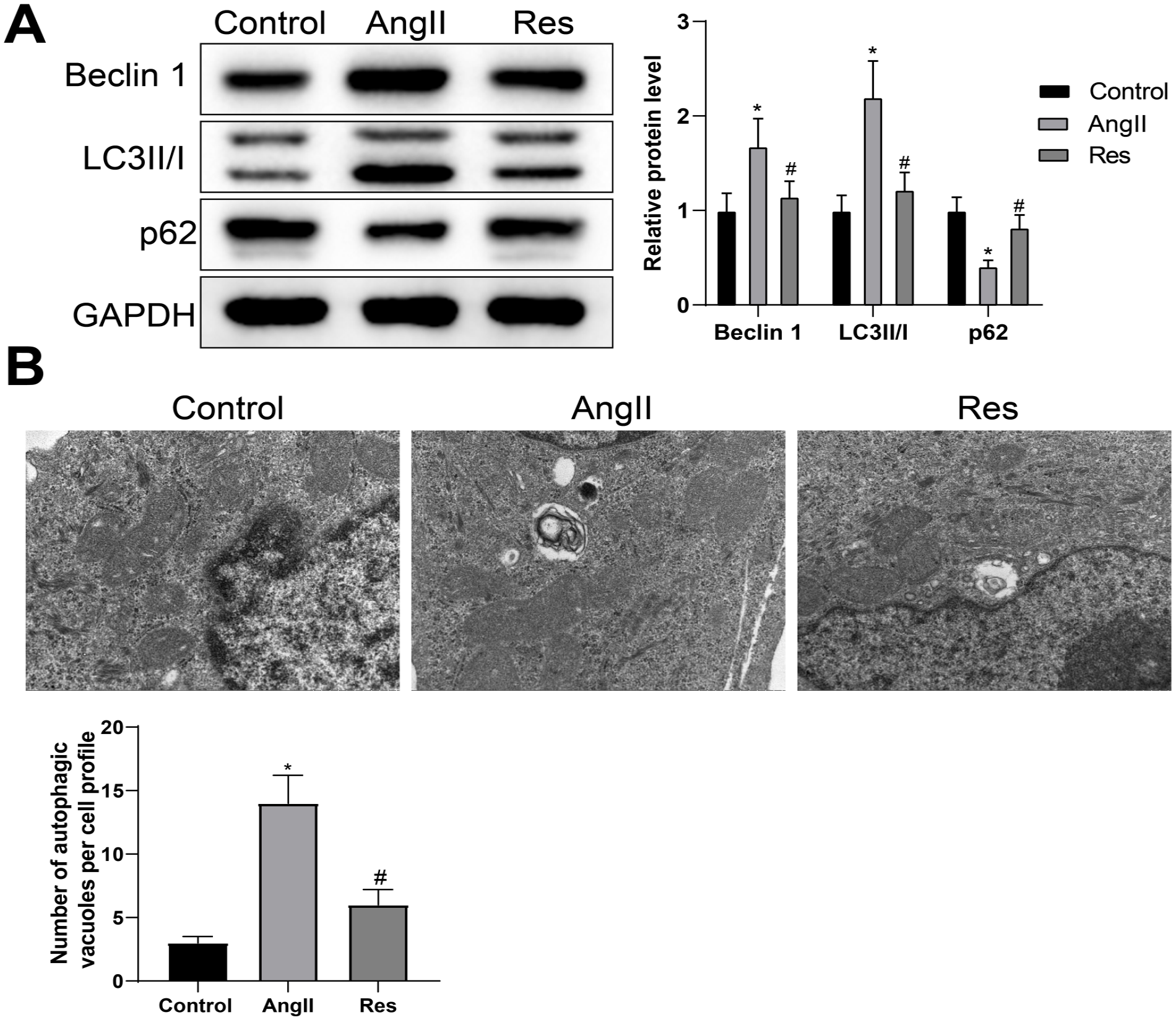
RES represses Ang II-induced autophagy in VSMC. The protein levels of Beclin 1, LC3 II/I, and p62 were measured by western blot (A). The numbers of autophagic vesicles were observed by TEM (B). One-way ANOVA and Tukey’s test were used to analyze the data among multiple groups. Experiments were repeated in triplicate; *p<0.05, vs control group; #p<0.05, vs Ang II group; RES, resveratrol; Ang II, angiotensin II; VSMC, vascular smooth muscle cell; TEM, transmission electron microscopy; ANOVA, analysis of variance.
Resveratrol Restrains Vascular Smooth Muscle Cell Extracellular Matrix Degradation, Apoptosis, Autophagy, and Inflammation via HMOX1 Upregulation in Abdominal Aortic Aneurysm
To further investigate whether HMOX1 mediates the protective effect of RES against AAA, sh-HMOX1 or sh-NC was transfected into RES-pretreated VSMCs before Ang II stimulation, which were separately identified as the Res-sh-HMOX1 group and Res-sh-NC group. Quantitative reverse transcription polymerase chain reaction and western blot showed that the Res-sh-HMOX1 group had attenuated HMOX1/HO-1 levels (vs Res-sh-NC group) (Figure 6A and B), along with elevated levels of MMP-2 and MMP-9, and decreased level of elastin (vs Res-sh-NC group) (Figure 6C and D).
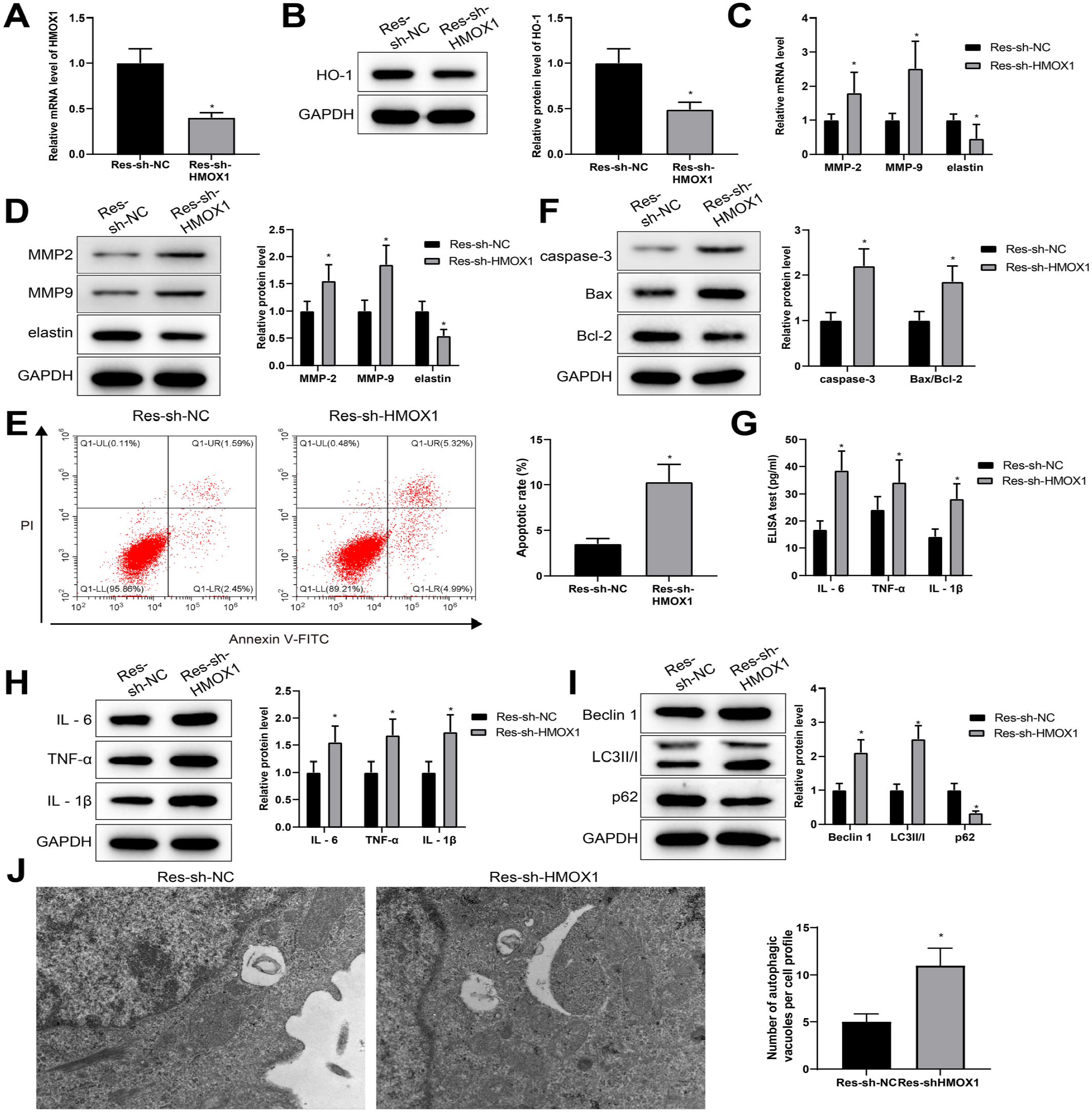
RES protects against VSMC ECM degradation, apoptosis, autophagy, and inflammation in AAA via upregulating HMOX1. After RES-pretreated VSMC was transfected with sh-HMOX1 and then stimulated by Ang II, HMOX1 mRNA and HO-1 protein levels were measured (A) and (B). mRNA and protein levels of MMP-2, MMP-9, and elastin were detected (C) and (D). The apoptosis rate was monitored by FCM (E). Western blot was applied to measure the levels of caspase-3 and Bax/Bcl-2 (F). The levels of IL-6, TNF-α, and IL-1β were detected by ELISA and western blot (G) and (H). The protein levels of Beclin 1, LC3 II/I, and p62 were examined by western blot (I). The numbers of autophagic vesicles were evaluated by TEM (J). The comparison between 2 groups was performed by t test, and each experiment was repeated 3 times; *p<0.05, vs sh-NC group; RES, resveratrol; AAA, abdominal aortic aneurysm; VSMC, vascular smooth muscle cell; Ang II, angiotensin II; HO-1, heme oxygenase-1; FCM, flow cytometry; TEM, transmission electron microscopy; ECM, extracellular matrix; MMP-2, matrix metalloproteinase-2; MMP-9, matrix metalloproteinase-9; ELISA, enzyme-linked immunosorbent assay; TNF, tumor necrosis factor; IL, interleukin.
Flow cytometry and western blot showed that knockdown of HMOX1 eliminated the inhibition of RES pretreatment on apoptosis. Also, the Res-sh-HMOX1 group had increased apoptosis rate, in parallel with the elevated protein levels of caspase-3 and Bax/Bcl-2 (vs Res-sh-NC group) (Figure 6E and F). Both ELISA and western blot showed that attenuation of HMOX1 eliminated the inhibition of RES pretreatment on inflammation (Figure 6G and H). Western blot and TEM revealed that downregulation of HMOX1 abolished the inhibition of RES pretreatment on autophagy (Figure 6I and J). Combining with the above findings, RES upregulated HMOX1 expression in VSMCs, possibly contributing to the prevention of AAA.
Resveratrol Inhibits Abdominal Aortic Aneurysm Development in Rats by Upregulating HMOX1
The anti-AAA effect of RES was further tested in a rat model of AAA. The incidence of AAA and the abdominal aorta diameter of the rats in each group are shown in Figure 7A and B. No aneurysm was found in the sham group. The incidence of AAA in the AAA group was 90%, and the diameter of the abdominal aorta was significantly increased. Resveratrol treatment decreased the incidence of AAA and the diameter of the abdominal aorta; however, sh-HMOX1 transfection reduced the therapeutic effect of RES. The detection of qRT-PCR and western blot revealed that HMOX1 mRNA and HO-1 protein were downregulated in the abdominal aorta tissues of the AAA group. The levels of HMOX1 mRNA and HO-1 protein were rescued by RES; however, they were downregulated in rats co-treated with sh-HMOX1 and RES (Figure 7C and D). The western blot analysis also showed that the expression trends of MMP-2, MMP-9, IL-6, TNF-α, IL-1β, caspase-3, Bax, Beclin 1, and LC3 II/I were opposite to that of HO-1, whereas the expression trends of elastin, Bcl-2, and p62 were consistent with that of HO-1 (Figure 7E and H). Besides, MDA level in the aortic tissues of rats was detected using ELISA kit to explore whether RES mediated the antioxidant effect of AAA rats through HO-1. Results displayed that the AAA group had higher MDA level than the sham group, which could be reversed by RES treatment; however, sh-HMOX1 transfection elevated the level of MDA (Figure 7I). These data indicated that RES inhibited the progression of AAA in rats by upregulating HMOX1.
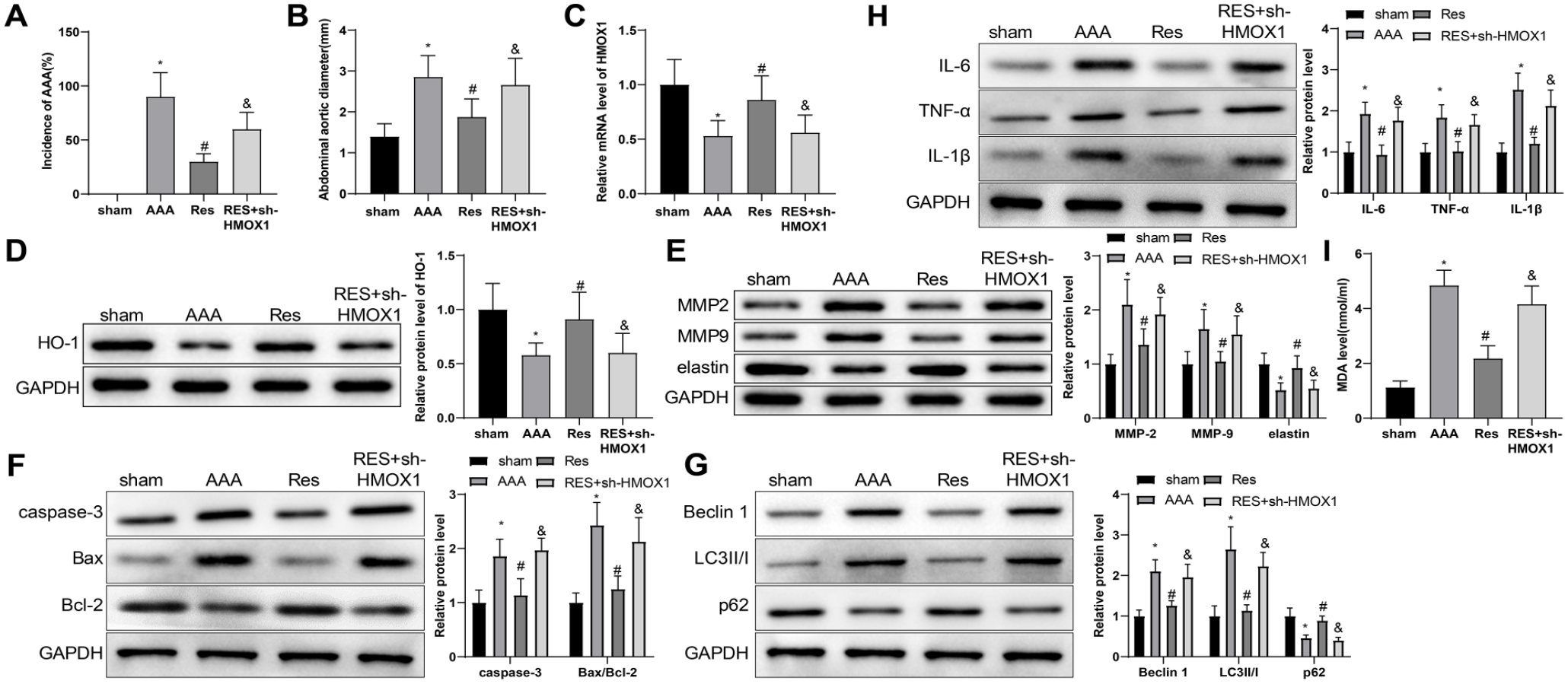
RES inhibits AAA development in rats by upregulating HMOX1. The incidence of elastase-induced AAA in rats (A). The abdominal aorta diameter of the rats in each group (B). qRT-PCR and western blot were used to detect the expression of HMOX1 mRNA and HO-1 protein (C) and (D). Western blot was used to detect the expression of MMP-2, MMP-9, and elastin (E). Western blot was used to measure the protein expression of caspase-3, Bax, and Bcl-2 (F). Western blot was used to evaluate the protein expression of Beclin 1, LC3 II/I, and p62 (G). Western blot was used to evaluate the protein expression of IL-6, TNF-α, and IL-1β (H). ELISA was used to test the level of MDA (I). One-way analysis of variance was used for comparison between multiple groups, followed by Tukey’s multiple comparisons test. Ten rats in each group. *p<0.05, vs sham group; #p<0.05, vs AAA group; &p<0.05, vs RES group. RES, resveratrol; AAA, abdominal aortic aneurysm; MMP-2, matrix metalloproteinase-2; MMP-9, matrix metalloproteinase-9; HO-1, heme oxygenase-1; ELISA, enzyme-linked immunosorbent assay; MDA, Malondialdehyde; TNF, tumor necrosis factor; IL, interleukin; qRT-PCR, quantitative reverse transcription polymerase chain reaction.
Discussion
Abdominal aortic aneurysm is a disease that carries significant mortality in the context of lacking early detection and treatment. As the major cell type in the vessel wall, VSMCs could provide elastin to recover the elasticity of the aortic wall. 28 This study was designed to probe whether RES could reduce AAA-related VSMC apoptosis and ECM degradation. Hence, this study established a cellular and a rat model of AAA using Ang II and pancreatic elastase, respectively. In the present work, we disclosed that RES could reduce MMP-9/2 expression and restrain the apoptosis, inflammation, and autophagy in Ang II-induced VSMCs. Mechanistically, RES upregulated HMOX1 to function its anti-apoptosis and anti-inflammatory effects in Ang II-induced VSMCs. Also, RES inhibited the progression of AAA in rats by upregulating HMOX1.
Many studies have demonstrated the protective effects of RES in AAA,21,23,29 yet whether RES could regulate VSMC function (including ECM degradation, apoptosis, autophagy, and inflammation) in AAA progression has not been determined. Several lines of evidence have shown that oxygen-derived free radicals and other biomolecules from the inflammatory cells could induce VSMC apoptosis and phenotypic changes in AAA pathogenesis, and degradation of ECM, coupled with inflammation in the aortic wall, which are pathological hallmarks of AAA.30–32 Matrix metalloproteinases, a family of zinc-dependent endopeptidases, hold great importance in ECM degradation. Matrix metalloproteinases, especially MMP-2 and MMP-9, were deemed as key regulators in the formation and development of AAA. 33 Elastin is also a vital protein of ECM that provides elasticity and resilience to tissues and organs including large arteries.34,35 Elastin loss is a main pathological change of AAA. 36 Our work illustrated that the levels of MMP-2 and MMP-9 were upregulated, while elastin was inhibited in Ang II-stimulated VSMCs. However, 10 µM RES pretreatment could reverse the effects of Ang II treatment on the levels of MMP-2, MMP-9, and elastin. Here, we illustrated that RES could repress ECM degradation in VSMCs stimulated by Ang II. Previously, RES could preserve the contractile phenotype of VSMCs in response to subarachnoid hemorrhage. 37 Resveratrol also seems to play a suppression role in VSMC apoptosis and inflammation.38,39 Here, our results showed that RES strikingly reversed the aggravation of Ang II on the levels of apoptosis-related proteins (caspase-3 and Bax/Bcl-2) and inflammatory factors (IL-6, TNF-α, and IL-1β) in VSMCs. In addition, FoxO3a promotes AAA progression via the P62/LC3B II autophagy signaling pathway, indicating that the occurrence of AAA is associated with autophagy. 40 Our findings showed that Ang II treatment increased Beclin1 and LC3 II/I protein levels and decreased p62 level. On the contrary, RES treatment reversed the effects of Ang II on Beclin1, LC3 II/I, and p62 levels. Also, our results showed decreased number of autophagic vesicles after RES treatments, suggesting RES treatment could reduce Ang II-induced autophagy in VSMCs. Although we proved the protective effect of RES on restoring ECM degradation, apoptosis, autophagy, and inflammation in VSMCs in the presence of Ang II, the exact modulatory mechanisms remain to be discovered.
In coronary arterial endothelial cells, RES could increase HMOX1 expression in a concentration-dependent fashion. 41 Likewise, RES at a low concentration (1-10 µM) could concentration- and time-dependently induce HO-1 expression in human aortic smooth muscle cells. 42 Prior studies have shown that HMOX1 gene plays an important role in preventing the development of AAA.9,43 However, whether RES exerts anti-AAA effects in VSMCs through HMOX1 remains to be studied. Our work showed that RES treatment substantially abolished the inhibitory effect of Ang II on HMOX1 and HO-1 levels. Next, we evaluated whether HMOX1 could regulate ECM degradation and impact the apoptosis and inflammation of VSMCs. As a result, HMOX1 deficiency can promote mRNA and protein levels of MMP-2 and MMP-9 and repress elastin level in Ang II-stimulated VSMCs. Knockdown of HMOX1 increased apoptosis and protein levels of caspase-3 and Bax/Bcl-2. HMOX1 silencing accelerated inflammatory responses. Conversely, forced expression of HMOX1 could suppress Ang II-induced VSMC dysfunction. In this study, HMOX1 deficiency reversed the effects of RES pretreatment on decreasing MMP-2 and MMP-9 levels, elevating elastin level, and repressing apoptosis and inflammatory response in VSMCs, suggesting RES provides significant protection against Ang II-induced ECM degradation and VSMC apoptosis and inflammation by regulating HMOX1. Also, downregulation of HMOX1 eliminated the inhibition of RES pretreatment on VSMC autophagy. A previous study has reported that RES inhibits the development of experimental AAA by increasing the expression of angiotensin-converting enzyme 2 (ACE2), 21 suggesting that RES may exert its anti-AAA effects through multiple genes and pathways. It may be an interesting topic to future research that whether HMOX1 works in synergy with ACE2 in eliminating AAA.
Conclusions
Overall, this work provides a possible explanation for the action mechanism of RES regulating ECM degradation and affecting the apoptosis, inflammation, and autophagy of VSMCs in AAA by enhancing the expression of HMOX1. In this study, we used peri-aortic application of pancreatic elastase to induce rat models based on previous methods.9,44,45 Perhaps, we should try to use an Ang II-induced AAA rodent model to make the results more comparable, which is a limitation of our study. Because AAA research at the cellular and animal level is still in the development stage, more efforts should be carried out in vivo to determine whether RES may be beneficial in treating AAA and translate RES applications into clinical treatments.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.