
Abstract
Background:
This study aims to evaluate the therapeutic effects and potential mechanisms of p-cymene (CM) alone and in combination with quinine (Qu) against Plasmodium berghei-infected mice.
Methods:
A total of 108 BALB/c mice were randomly divided into nine groups included six infected groups, which received normal saline, Qu (10 mg/kg), CM 5 mg/kg, CM 10 mg/kg, CM (5 mg/kg) + Qu (10 mg/kg), and CM (10 mg/kg) + Qu (10 mg/kg) as well as three noninfected groups, which received normal saline, CM 5 mg/kg, and CM 10 mg/kg. Mice were intraperitoneally infected by 1 × 106 P. berghei malaria-infected erythrocytes. Infected mice were orally treated daily over a period of 4 days. Then parasite growth suppression (PGS), survival rate, the level of oxidant and antioxidant markers, and analysis of immune response-related genes were also evaluated.
Results:
The highest survival rate of 100% was observed in infected mice treated with a combination of CM and Qu, which also demonstrated a PGR value of 100% (p < 0.001). The combination of CM and Qu resulted in the most significant reductions in tissue concentrations of malondialdehyde and nitric oxide, while upregulating the expression of the superoxide dismutase, glutathione peroxidase, and interleukin-(IL)10 (>fourfold change) genes resulted in a reduction in the expression level of the tumor necrosis factor (<1.3-fold-change) and IL-1β (<1.4-fold change) genes. The combination of CM and Qu also caused significant modulation of serum levels of liver and kidney markers in malaria-infected mice.
Conclusion:
The results of this survey indicate that the combination therapy of CM with Qu demonstrates significant effectiveness in treating malaria-infected mice by regulating oxidative stress, enhancing antioxidant enzyme activity, and modulating inflammatory responses. However, to further validate the therapeutic potential of this compound, it is essential to conduct clinical trials that evaluate both its toxicity and therapeutic efficacy.
Introduction
Malaria is a severe tropical disease caused by protozoan parasites belonging to the genus Plasmodium (Cowman et al., 2016). Among the various Plasmodium species that infect humans, Plasmodium falciparum is recognized as the most lethal and virulent (Cowman et al., 2016). The World Health Organization’s Malaria Report indicates that in 2020, there were approximately 241 million cases of malaria globally, resulting in 627,000 fatalities (World Health Organization, 2020). Currently, the primary approach to malaria prevention and treatment involves the administration of chemical agents, including artemisinin, primaquine, chloroquine, and mefloquine (Bruxvoort et al., 2014). However, the emergence of resistance to antimalarial medications, particularly chloroquine in P. falciparum, has posed a significant public health challenge in developing nations (Yeung, 2018). The adverse effects associated with these pharmacological treatments, such as gastrointestinal disturbances, nausea, vomiting, headaches, dizziness, alopecia, anxiety, and sleep disturbances, further complicate their efficacy in malaria management (Hanboonkunupakarn and White, 2022). Consequently, the identification and development of new antimalarial agents that demonstrate high efficacy and minimal side effects are of paramount importance for improving malaria treatment outcomes.
Herbs and their derivatives are increasingly acknowledged as vital sources of a wide array of therapeutic compounds, primarily due to their availability, affordability, low toxicity, and significant efficacy (Cheraghipour et al., 2021). Monoterpenes, which are characterized by their hydrocarbon structures, constitute the largest group of plant secondary metabolites and are commonly found in essential oils (Wojtunik‐Kulesza et al., 2019). p-Cymene (CM, C10H14), also known as p-isopropyltoluene, is an alkyl-substituted aromatic compound that naturally occurs in the essential oils of numerous aromatic plants, including those from the genera Artemisia, Ocimum, Eucalyptus, and Thymus. Investigations in the biological sciences have indicated that CM exhibits a wide range of pharmacological properties, including anti-inflammatory, antioxidant, anticancer, antidiabetic, and antimicrobial effects (Balahbib et al., 2021; Marchese et al., 2017).
Although a considerable volume of research has demonstrated the effectiveness of various herbs and their derivatives in the treatment of malaria (Marques-Santos et al., 2024). Nevertheless, the interpretations of these studies are limited by several factors, including a lack of comprehensive understanding of the underlying mechanisms and concerns regarding potential toxicity. Considering the various limitations, adverse effects, and documented instances of drug resistance associated with synthetic drugs in malaria treatment, alongside the therapeutic potential of natural products—particularly monoterpene compounds derived from plants such as CM—this study aims to explore, for the first time, the potential therapeutic effects of CM in mice infected with Plasmodium berghei through assessing its impact on parasitic load, antioxidant activity, and anti-inflammatory properties.
Materials and Methods
Chemicals
CM (C10H14, purity >99%, No. 121452) was obtained from Sigma-Aldrich, Germany
Animals
One hundred and eight male BALB/c mice, each weighing between 25 and 30 grams and aged between 6 and 8 weeks, were procured from the Animal House at Imam Abdurrahman Bin Faisal University in Saudi Arabia. The mice were maintained under controlled environmental conditions, specifically at a temperature of 24 ± 1°C, with a 12-h light/dark cycle and relative humidity levels ranging from 40% to 70%. They were given unrestricted access to both water and food.
Establishment of a murine model of malaria
The P. berghei NK65 strain, obtained from Shaqra University in Saudi Arabia, was maintained through a weekly serial passage of blood from infected mice to noninfected counterparts. Mice exhibiting a parasitemia level of 20%–30% were selected for the purpose of parasite inoculation. Following the collection of infected blood from the donor mice via cardiac puncture into a heparinized vacutainer tube, the experimental mice were intraperitoneally inoculated with 200 µL of the infected blood, which contained approximately 1 × 106 P. berghei-parasitized erythrocytes (Kweyamba et al., 2019).
Study design and treatment
The study design involved the initiation of treatment 3 h postinoculation of the parasite in mice on day 1. This treatment was then orally administered daily over a period of 4 days, extending from day 1 to day 5. BALB/c mice were randomly divided into six infected groups, each consisting of 12 mice (Alkhaibari et al., 2024). In addition, three noninfected groups were included to assess the toxicity of the tested doses of CM on healthy mice. The selection of the doses of CM (5 and 10 mg/kg) was based on the previous reports, which indicated that CM at these doses did not lead to any mortality or abnormal clinical manifestations in animals (Kuwagata et al., 2024; Santos et al., 2022): Noninfected and nontreated mice Noninfected received CM 5 mg/kg Noninfected received CM 10 mg/kg Infected received normal saline (negative control) Infected received quinine (Qu) 10 mg/kg (positive control) Infected received CM 5 mg/kg Infected received CM 10 mg/kg Infected received Qu 10 mg/kg + CM 5 mg/kg Infected received + Qu 10 mg/kg + CM 10 mg/kg
Effects of CM on parasite growth suppression
The mortality rate of the mice was monitored on a daily basis for a duration of 4 weeks subsequent to the induction of the murine malaria model. On the fifth day postinfection, thin blood smears were prepared from the tails of the mice and subsequently stained with Giemsa to assess parasitemia and to evaluate the suppression of parasite growth using the formula established by Waako et al. (2005).
Evaluating the oxidative stress marker
On the fifth day following infection, six mice from each experimental group were subjected to deep euthanasia using a ketamine-xylazine combination (100:10 mg/kg). Subsequent to the extraction of brain tissues, the homogenates were employed to assess the expression levels of antioxidant and proinflammatory genes. The levels of lipid peroxidation (LPO) and nitric oxide (NO) were measured using commercial kits from Abcam (Abcam, USA), in accordance with the manufacturer’s instructions.
Evaluating the proinflammatory and antioxidant-related genes
Quantitative reverse transcription polymerase chain reaction (qRT-PCR) was utilized to assess the variations in the relative expression of genes implicated in inflammation (tumor necrosis factor [TNF-α], interleukin 1 beta [IL-1β], and IL-10), antioxidant (glutathione peroxidase [GPx]), and superoxide dismutase (SOD). Total RNA was extracted from the brain tissue of the treated mice using the Qiagen RNA kit, Germany, following the manufacturer’s established protocol for RNA isolation. The extracted RNA was subsequently transcribed using the Qiagen kit, Germany. The amplification products were quantified through SYBR Green Real-time PCR, allowing for the comparison of expression levels of the target genes relative to the housekeeping gene β-actin (Table 1). This was conducted using the StepOnePlus™ RT-PCR system (Thermo Fisher Scientific, USA) in conjunction with the Qiagen SYBR Green RT-PCR kit. The qRT-PCR protocol commenced with an initial denaturation step at 96°C for 9 min, followed by 40 cycles consisting of a denaturation phase at 96°C for 10 s and an annealing and extension phase at 64°C for 28 s. The 2−ΔΔCt values were subsequently calculated using the Bio-Rad iQ5 Optical System Software (USA) (Alkhaibari et al., 2024).
The Primers Were Applied for Real-Time Polymerase Chain Reaction
GPx, glutathione peroxidase; IL-1β, Interleukin 1 beta; SOD, superoxide dismutase; TNF-α, tumor necrosis factor.
Biochemical analysis and safety
A biochemical and toxicological evaluation of essential organs was conducted across all experimental groups. One day following the treatment, the mice were anesthetized with a combination of ketamine and xylazine (Sigma-Aldrich, Germany) at a dosage ratio of 100:10 mg/kg. Subsequent to anesthesia, serum samples were obtained from the cardiac blood of the mice and analyzed using commercial kits from Roche (Roche – Hitachi MODULAR Analytics, Japan) to determine serum levels of indicators of kidney function, specifically blood urea nitrogen (BUN) and creatinine (Cr), as well as liver function markers, including alanine transaminase (ALT) and aspartate transaminase (AST) (Mahmoudvand et al., 2016; Mahmoudvand et al., 2017).
Statistical analysis
Subsequent to the data collection phase, descriptive statistical techniques were utilized to delineate the characteristics of the dataset, which encompassed the computation of measures of central tendency and variability. The results were articulated as mean ± standard deviation, accompanied by the relevant p values, to facilitate a comprehensive statistical evaluation. To further explore the dataset, an analysis of variance (ANOVA) was executed, augmented by Tukey’s and post hoc tests. All statistical analyses were conducted using SPSS version 26 software, with a two-tailed significance level established at p < 0.05.
Results
In vivo antimalarial effects of LZNPs
The results demonstrated that mice infected with the P. berghei NK65 strain and treated with CM at dosages of 5 and 10 mg/kg, mainly in combination with Qu (10 mg/kg), exhibited a significant increase (p < 0.001) in survival rates and a reduction in parasite growth over the 28-day observation period, in comparison to the control group that received normal saline by ANOVA analysis (Fig. 1). The highest survival rate was observed 100% after treatment of the infected mice with the combination of the CM (5 and 10 mg/kg) with Qu (10 mg/kg) (p < 0.05, post hoc analysis). Considering the parasite growth suppression rate, the values for CM 5 mg/kg, CM 10 mg/ kg, Qu (10 mg/kg)+ CM 5 mg/kg, and Qu (10 mg/kg) + CM 10 mg/kg were 46.8%, 68.8%, 100%, and 100%, respectively, indicated the significant suppression in comparison with the infected mice received normal saline (p < 0.001).
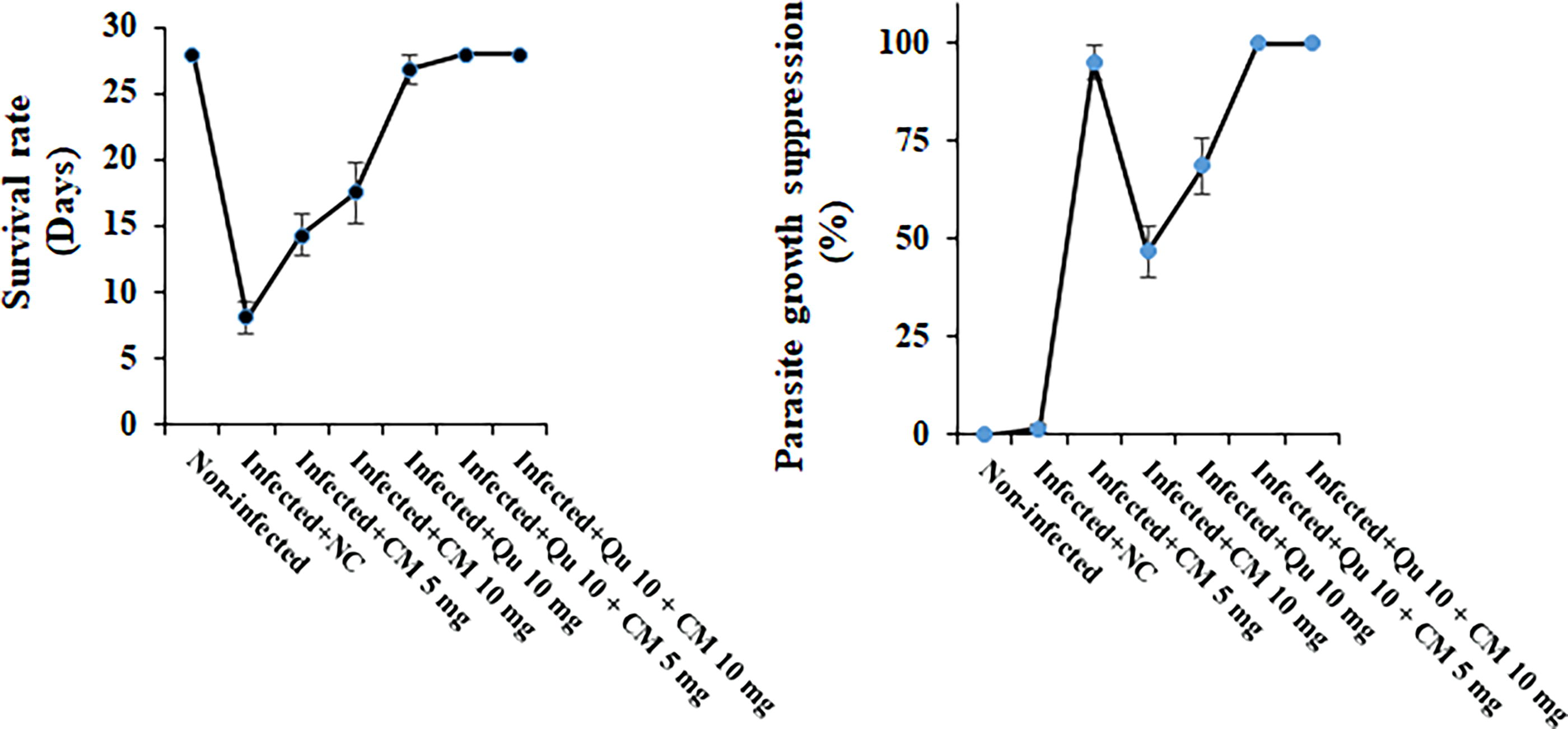
In vivo antimalarial effects of CM and Qu alone and in combination against strains of Plasmodium berghei-infected mice. Statistical analysis was conducted utilizing one-way ANOVA, accompanied by Tukey’s post hoc test. The results are presented as mean ± standard deviation (N = 12). ANOVA, analysis of variance; CM, p-cymene; NC, normal saline; Qu, quinine.
Reducing the oxidative stress marker
Figure 2 presents data indicating that the tissue levels of MDA and NO in the malaria-infected mice were significantly elevated compared to those in noninfected mice (p < 0.001). Conversely, the treatment involving CM, principally the combination therapy of CM at 5 mg/kg and Qu at 10 mg/kg (MDA = 1.69 nmol/mg tissue, NO = 1.34 nmol/mg tissue) as well as CM at 10 mg/kg and Qu at 10 mg/kg (MDA = 1.27 nmol/mg tissue, NO = 1.12 nmol/mg tissue) exhibited the most pronounced reduction in the tissue levels of MDA and NO in the malaria-infected mice (p < 0.01), as determined by post hoc analysis.
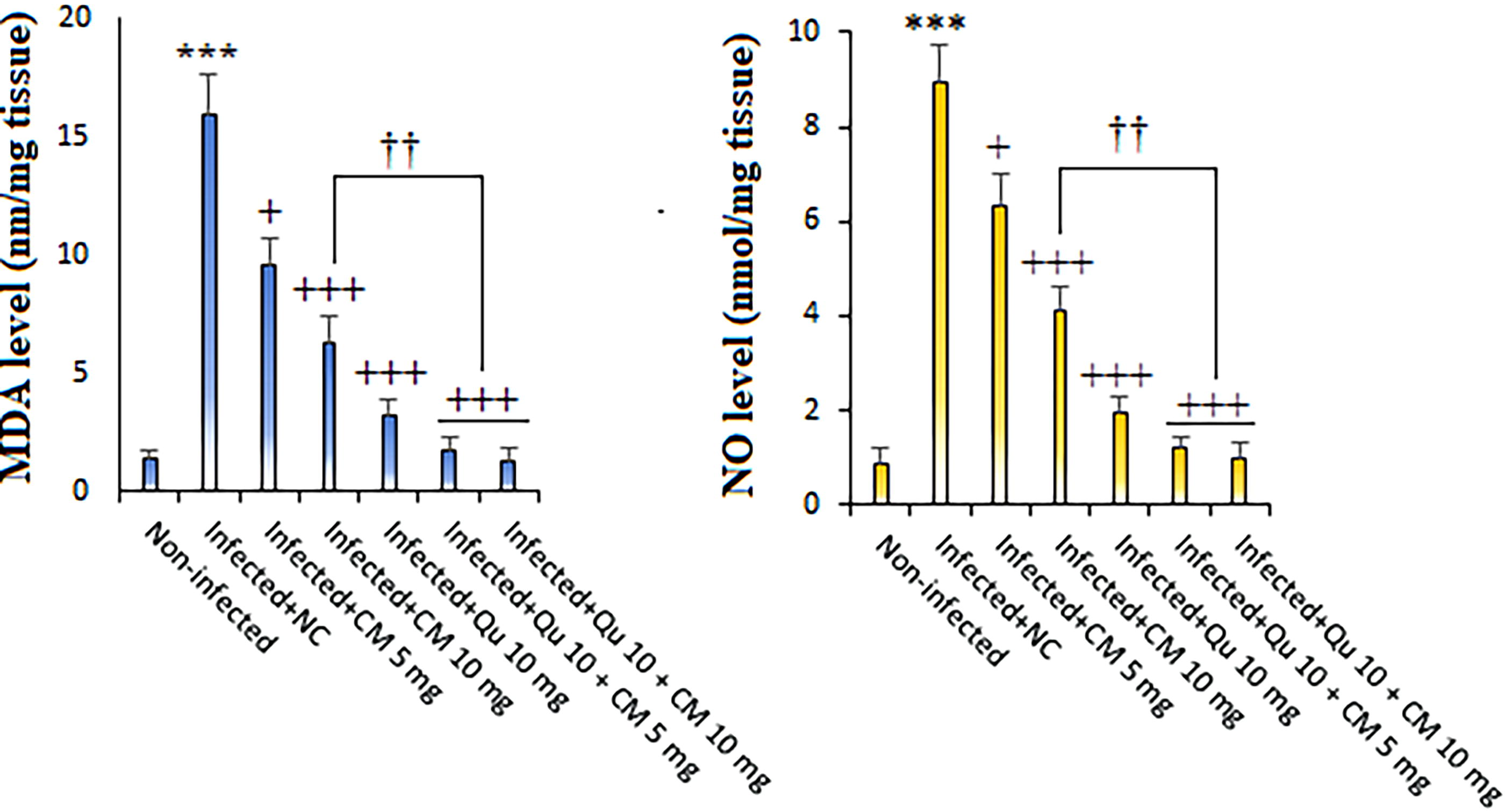
The effects of CM) and Qu alone and in combination on the tissue level of MDA and NO in the Plasmodium berghei-infected mice. Statistical analysis was conducted utilizing one-way ANOVA, accompanied by Tukey’s post hoc test. The results are presented as mean ± standard deviation (N = 12). *** p < 0.001 significant difference compared to noninfected mice; + p < 0.05 and +++ p < 0.001 compared to the infected mice received NC. †† p < 0.01 compared to the infected mice received Qu. MDA, malondialdehyde; NO, nitric oxide.
Evaluating the expression level of antioxidant genes
Figure 3 depicts a considerable decrease in the expression levels of the SOD and GPx genes in mice infected with malaria when compared to noninfected control subjects (ANOVA, p < 0.001). The treatment regimen involving CM, specifically the combination of Qu at 10 mg/kg + CM at 5 mg/kg (SOD = 4.66-fold change and GPx = 3.64-fold change), as well as Qu at 10 mg/kg + CM at 10 mg/kg (SOD = 4.98-fold change and GPx = 5.21-fold change), led to a significant upregulation of the SOD and GPx genes, as determined by post hoc analysis.
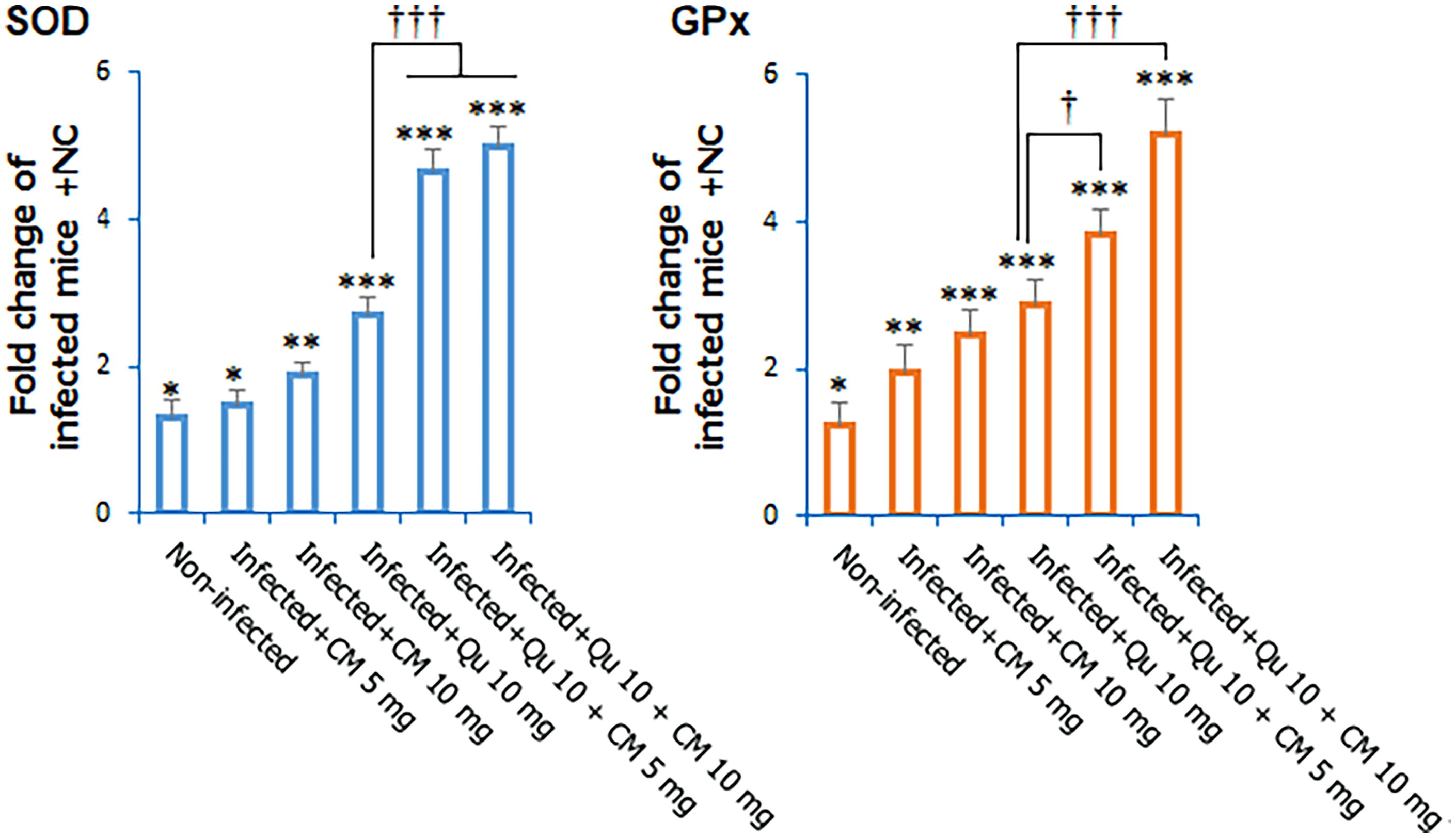
The effects of CM and Qu alone and in combination on the expression level of GPx and SOD genes in the Plasmodium berghei-infected mice. Statistical analysis was conducted utilizing one-way ANOVA, accompanied by Tukey’s post hoc test. The results are presented as mean ± standard deviation (N = 12). * p < 0.05, ** p < 0.01, and *** p < 0.001 significant difference compared to infected mice received NC; + p < 0.05 and +++ p < 0.001 compared to the infected mice received NC. † p < 0.05, ††† p < 0.001 significant difference compared to the infected mice received Qu. GPx, glutathione peroxidase; SOD, superoxide dismutase.
Modulating the expression level of proinflammatory genes
The results obtained from real-time PCR analysis utilizing the 2−ΔΔCT method indicated that mice infected with P. berghei NK65 demonstrated significantly increased expression levels of the TNF-α and IL-1β genes, alongside decreased expression levels of the IL-10 gene, in comparison to the noninfected control group (ANOVA, p < 0.001) (Fig. 4). Followed by treatment protocol utilizing CM, particularly the combination of Qu at 10 mg/kg + CM at 5 mg/kg (TNF-α = 1.28-fold change, IL-1β = 1.31-fold change, and IL-10 = 4.29-fold change), as well as Qu at 10 mg/kg + CM at 10 mg/kg (TNF-α = 1.09-fold change IL-1β = 1.12-fold change, and IL-10 = 4.77-fold change), the expression of TNF-α and IL-1β genes was found to be reduced, while the expression levels of IL-10 genes were elevated, as confirmed by post hoc analysis (p < 0.05).
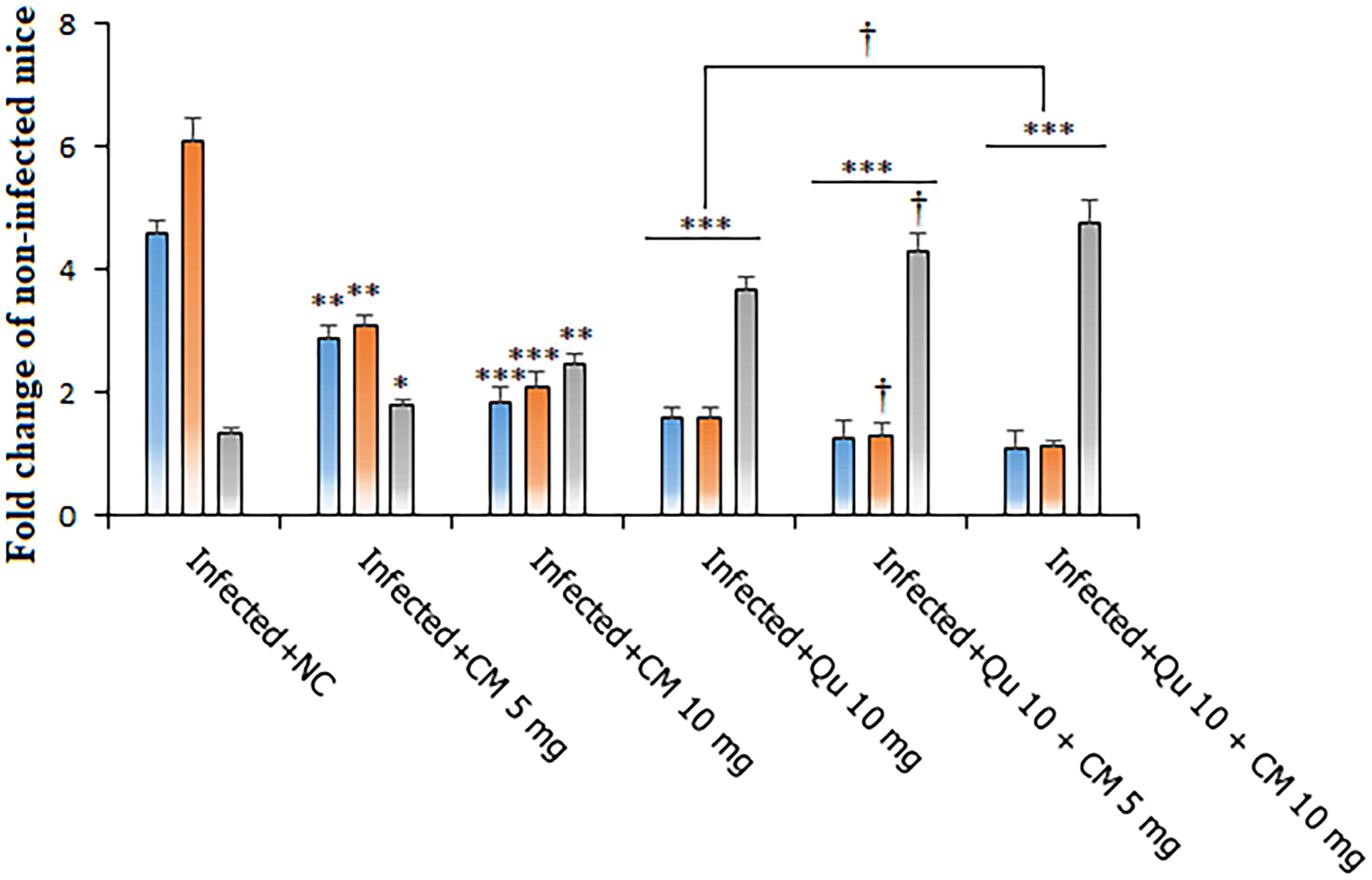
The effects of CM and Qu alone and in combination on the expression level of TNF-α, Interleukin 1 beta (IL-1β), IL-10 genes in the Plasmodium berghei-infected mice. Statistical analysis was conducted utilizing one-way ANOVA, accompanied by Tukey’s post hoc test. The results are presented as mean ± standard deviation (N = 12). * p < 0.05, ** p < 0.01, and *** p < 0.001 significant difference compared to infected mice received NC; † p < 0.05, significant difference compared to the infected mice received Qu. IL-1β, Interleukin 1 beta; TNF-α, tumor necrosis factor.
Safety and biochemical analysis
The biochemical assessments demonstrated a notable increase in serum concentrations of biomarkers associated with liver and kidney function in malaria-infected mice (p < 0.001) in comparison with noninfected mice. Subsequent treatment with CM, particularly the combination of Qu at 10 mg/kg + CM at 5 mg/kg (p < 0.001), as well as Qu at 10 mg/kg + CM at 10 mg/kg (p < 0.001) over a 5-day duration, resulted in a significant modulation of serum levels of AST, ALT, BUN, and Cr in the malaria-infected mice. Post hoc analysis also revealed no significant differences in the serum levels of liver and kidney function biomarkers between healthy mice that received CM at doses of 5 and 10 mg/kg and the control group of healthy mice that received normal saline (Fig. 5).
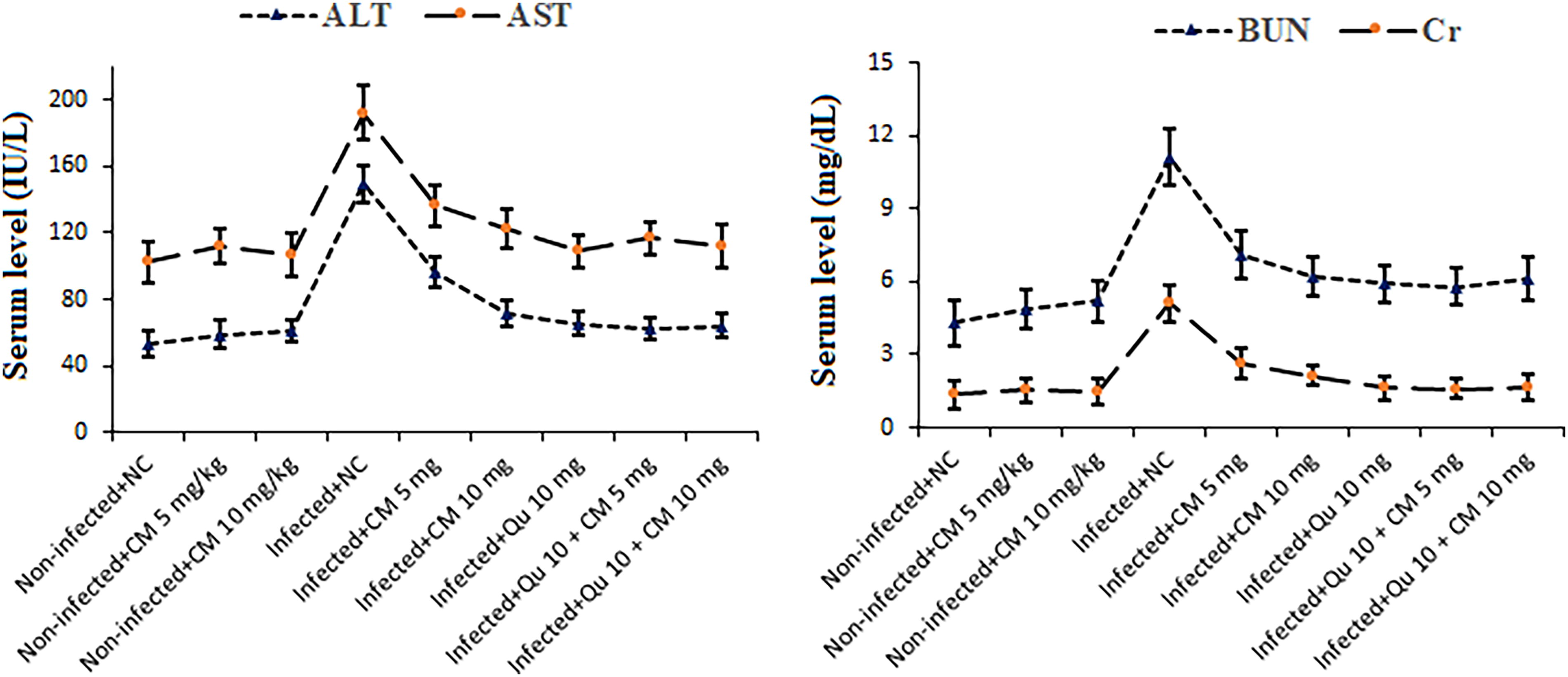
The effects of CM and Qu alone and in combination on the serum level of the liver functional factors of ALT and AST as well as kidney functional factors BUN and Cr in the noninfected mice and the Plasmodium berghei-infected mice. Statistical analysis was conducted utilizing one-way ANOVA, accompanied by Tukey’s post hoc test. The results are presented as mean ± standard deviation (N = 12). ALT, aspartate transaminase; AST, alanine transaminase; BUN, blood urea nitrogen; Cr, creatinine.
Discussion
The emergence and spread of resistance among malaria parasites to specific antimalarial medications have become a significant challenge in the effective management of malaria in endemic regions. This resistance is likely a contributing factor to the recent resurgence of malaria infections and the increase in malaria-related mortality (Yeung, 2018). Additionally, the adverse effects associated with chemical antimalarial drugs—such as gastrointestinal disturbances, nausea, vomiting, headaches, dizziness, alopecia, anxiety, and sleep disorders—have further complicated their therapeutic efficacy (Hanboonkunupakarn and White, 2022). Consequently, the identification and development of new antimalarial agents that exhibit high potency and minimal side effects could substantially enhance the treatment of malaria. Given the pharmacological properties of CM, the present experimental investigation was undertaken to evaluate the therapeutic efficacy of CM in a murine model of malaria.
The current results demonstrated that mice infected with the P. berghei NK65 strain and treated with CM at dosages of 5 and 10 mg/kg mainly in combination with Qu (10 mg/kg) exhibited a significant increase (p < 0.001) in survival rates and a reduction in parasite growth over the 28-day observation period, in comparison to the control group that received normal saline by ANOVA analysis.
Combination therapy by two or more beneficial drugs is considered as one of the top strategies that is now applied for treating malaria (Fu et al., 2023). Considering the benefits of combination therapy with natural products, prior investigations described that natural small molecules can serve as drug delivery systems to improve bioavailability, reduce toxicity, increase drug concentration, and prolong systemic circulation. This can improve the effectiveness of conventional drugs (Fu et al., 2023).
Recent investigations have provided substantial evidence regarding the antimicrobial properties of CM and the herbal essential oils it encompasses, demonstrating their effectiveness against a diverse array of microorganisms. This includes various bacterial strains such as Salmonella spp., Staphylococcus spp., Streptococcus spp., and Pseudomonas spp., as well as fungal species like Candida spp. and Aspergillus spp. (Balahbib et al., 2021; Marchese et al., 2017). In terms of the antiparasitic effects of CM, a study conducted by Glans et al. (2012) assessed the in vitro antiparasitic efficacy of a series of chelating cationic ruthenium (II)-arene complexes containing CM against two strains of P. falciparum, one of which was resistant to chloroquine, as well as a strain of Trichomonas vaginalis. Their results indicated that the complexes incorporating CM exhibited superior activity compared to those containing benzene. Subsequently, in 2014, Kpadonou Kpoviessi et al. (2014) investigated the in vitro antitrypanosomal and antiplasmodial properties of CM-rich essential oil derived from Ocimum gratissimum, targeting Trypanosoma brucei brucei and P. falciparum. Notably, CM constituted approximately 31.53% of the essential oil, which demonstrated antiparasitic activity, as evidenced by a concentration that resulted in the mortality of 50% of the parasites, with a half maximal inhibitory concentration (IC50) of 76.32 μg/mL. Furthermore, Santana et al. (2020) reported the potential antileishmanial properties of 1,8-cineole, α-pinene, and CM against Leishmania amazonensis amastigotes, with observed IC50 values of 48.4 μg/mL, 37 μg/mL, and 46 μg/mL, respectively. The variations observed in these findings can be attributed to several factors, including the specific species of parasite, the particular plant utilized, the methodologies employed, and the concentrations administered. Monoterpenes and sesquiterpenes have consistently demonstrated significant antimalarial efficacy against various Plasmodium species, including P. falciparum and P. berghei (Gabriel et al., 2018). In a study by Santos et al. (2022), 1,8-cineole exhibited a strong in vitro schizonticidal effect against P. falciparum, with an IC50 value of 1045.53 μM. Furthermore, the administration of 1,8-cineole (100 mg/kg) to mice infected with the P. berghei ANKA strain over a period of 6 days resulted in a substantial reduction in cerebral edema and a 50% decrease in parasitemia.
Current research indicates that the increase in oxidative stress associated with malaria contributes to oxidative damage throughout the body, particularly affecting vital organs such as the brain and liver (Gomes et al., 2022). This disruption in redox homeostasis has been correlated with severe clinical manifestations, including cerebral malaria (Gomes et al., 2022). In response to oxidative damage, the body employs a defense mechanism comprising antioxidants, which include both enzymatic and small molecular compounds, to neutralize the free radicals produced during infection (Nandi et al., 2022). Nevertheless, these endogenous defenses frequently prove inadequate to fulfill the body’s requirements. Consequently, researchers have explored supplementary antioxidant therapies aimed at enhancing the body’s antioxidant defense system (Nandi et al., 2022). The present study revealed that the therapeutic regimen incorporating combination therapy with CM, specifically at dosages of 5 mg/kg in conjunction with Qu at 10 mg/kg, as well as CM at 10 mg/kg paired with Qu at 10 mg/kg, demonstrated the most significant decrease in tissue concentrations of MDA and NO. Furthermore, this treatment resulted in a notable upregulation of the SOD and GPx genes in mice infected with malaria. De Oliveira et al. (2015) provided evidence that the administration of CM at dosages of 10 and 15 mg/kg resulted in a significant reduction of LPO and an enhancement of SOD and catalase activities in the hippocampus of infected mice. This was substantiated by the quantification of thiobarbituric acid levels. Investigations reported that CM exhibited its antioxidant effects through various mechanisms such as scavenging free radicals, inhibiting lipid peroxidation products and nitrite content, and increasing the activity of antioxidant enzymes like SOD and CAT. It has been proven that the metal ions can catalyze the production of harmful free radicals, whereas CM can bind to these metal ions, preventing them from participating in these harmful reactions (de Oliveira et al., 2015; Balahbib et al., 2021). These results underscore the considerable antioxidant properties of CM in the control of malaria in the infected mice.
It has been established that the primary objective of any prospective antimalarial agent is the elimination of the malaria-causing parasite. However, the most critical manifestation of the disease, referred to as cerebral malaria, is associated with elevated levels of inflammatory cytokines (Dunst et al., 2017). Severe malaria cases pose significant health risks and require effective therapeutic interventions. Consequently, the development of a novel compound that possesses dual functionalities—targeting the parasitic Plasmodium spp. while simultaneously mitigating the detrimental inflammation induced by the infection—represents a promising approach in contemporary pharmaceutical research (Miranda et al., 2013). The current study showed that followed by treatment protocol utilizing CM, particularly the combination of Qu at 10 mg/kg and CM at 5 mg/kg, as well as Qu at 10 mg/kg and CM at 10 mg/kg, the expression of TNF-α and IL-1β genes was found to be reduced, while the expression levels of IL-10 genes were elevated, indicating that the findings of this study corroborate that CM mainly combined with Qu possesses the capability to effectively control and treat malaria through the inhibition of specific inflammatory cytokines. Studies showed that CM displays anti-inflammatory effects through some mechanisms such as, by moderating the release of proinflammatory cytokines, constraining the main enzymes involved in inflammation such as cyclooxygenase and lipoxygenase, and affecting the stimulation of nuclear factor-κB and mitogen-activated protein kinases signaling pathways (Balahbib et al., 2021).
Currently, it is considered a standard procedure to investigate the toxicological effects of novel pharmaceutical therapies prior to their commercialization (Fielden and Kolaja, 2008). In this study, we examined the safety profile and protective effects of CM on hepatic and renal function in healthy murine models. The findings indicated that subsequent administration of CM, specifically the combination of Qu at 10 mg/kg and CM at 5 mg/kg (p < 0.001), as well as Qu at 10 mg/kg and CM at 10 mg/kg (p < 0.001) over a 5-day period, led to a significant alteration in serum levels of AST, ALT, BUN, and Cr in malaria-infected mice. Moreover, there were no statistically significant differences observed in the serum levels of biomarkers associated with liver and kidney function between healthy mice administered CM at doses of 5 and 10 mg/kg and the control group of healthy mice that were treated with normal saline. In alignment with our findings, Kuwagata et al. (2024) reported that the oral administration of CM at dosages of 2.4 and 12 mg/kg/day over a duration of 90 days did not lead to any mortality or abnormal clinical manifestations in the tested rats. Additionally, the no-observed-adverse-effect level for CM was established at 12 mg/kg/day for both male and female rats within the context of the present study, considering the alterations noted in urinalysis, hematological parameters, clinical biochemistry, and histopathological evaluations. In our recent research, we observed the significant efficacy of CM in treating malaria in mice. However, further investigation is required to examine the pharmacodynamic effects of CM on diverse biological targets. Additionally, pharmacokinetic studies are imperative to evaluate the absorption, bioavailability, and metabolic pathways related to this compound.
Conclusion
The results of this survey reveal that the combination therapy of CM with Qu exhibits considerable effectiveness in the treatment of malaria-infected mice. The findings imply that possible mechanisms for the management of malaria may involve the regulation of oxidative stress, the enhancement of antioxidant enzyme activity, and the modulation of inflammatory responses, all while preserving the functionality of essential organs without inducing toxicity. To further validate the therapeutic potential of this compound, it is essential to undertake clinical trials that assess both its toxicity and therapeutic efficacy.
Footnotes
Acknowledgment
The author would like to thank the Deanship of Scientific Research at Tabuk University for supporting this work.
Author’s Contributions
A.F.S. has planned and performed tests as well as analyzed and written the article. A.F.S. has also reviewed and approved the final version of the article for publication.
Ethical Statement
The animals were also handled according to Guides for the Care and Use of Laboratory Animals and ARRIVE guidelines checklists. The experimental protocol was designed and approved by the animal ethics committee of Almaarefa University in Saudi Arabia (IRB07-18052022-46) in Spring 2022.
Data Availability Statement
The dataset presented in the study is available on request from the corresponding author during submission or after its publication.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.