
Abstract
Background:
The COVID-19 pandemic, driven by the SARS-CoV-2 virus, has become an unprecedented global public health challenge. Comprehensive investigation of the virus transmission dynamics requires exploration of all potential routes, including the role of animal reservoirs. This study aimed to assess the presence of SARS-CoV-2 RNA and antibodies in urban rodents captured within two landscape units (low-income neighborhoods and green spaces) in the metropolitan area of Buenos Aires (MABA), Argentina.
Methods:
Between July 2020 and September 2022, 125 synanthropic rodents (68 Rattus norvegicus, 22 R. rattus, and 35 Mus musculus) from seven low-income neighborhoods and seven green spaces in the MABA were captured and examined for SARS-CoV-2 exposure and infection by indirect immunofluorescence serology and RT-qPCR, targeting the N, E, and ORF1ab genes.
Results:
Two M. musculus and six R. norvegicus from five different low-income neighborhoods tested positive for SARS-CoV-2 antibodies. No rodent with positive serology was detected in green spaces. All rodents tested negative for SARS-CoV-2 RNA when analyzed by RT-qPCR.
Conclusion:
The results of this study provide valuable insights into the ecological dynamics of SARS-CoV-2 in synanthropic rodents in two landscape units within a Latin American city, with seropositive rodents detected in low-income neighborhoods, suggesting a possible increased exposure risk in these settings. This information is critical for developing management strategies for potentially zoonotic viruses and underscores the importance of continued surveillance of SARS-CoV-2 in rodents as part of an integrated One Health approach.
Introduction
In recent decades, pandemics caused by zoonotic viral pathogens have had a significant impact on humanity (Bhadoria et al., 2021). The COVID-19 pandemic, driven by SARS-CoV-2 virus, represents an unprecedented public health emergency, which has severely impacted on health care systems, economies, and societies around the world (Cutler and Summers, 2020; Naseer et al., 2022). This crisis has highlighted the urgent need to increase our understanding of the mechanisms underlying infectious diseases, including transmission routes and animal reservoirs, particularly in the context of the potential zoonotic origins of SARS-CoV-2 (Ray and Bhattacharya, 2023; Yang et al., 2023). This is especially important in urban and peri-urban ecosystems because the high densities of livestock, wildlife, and companion animals living in very close contact with dense human populations, and the presence of urban parks, vacant lots, and zoos, could increase the risk of pathogen transmission between humans, wildlife, livestock, and companion animals (Dobigny and Morand, 2022).
Synanthropic rodents, such as the brown rat (Rattus norvegicus), the black rat (R. rattus), and the house mouse (Mus musculus), live in close association with humans (Feng and Himsworth, 2014). They are critical targets for surveillance due to their role as reservoirs for several pathogens, including some coronaviruses responsible for zoonotic diseases (Akhtar et al., 2023; Gravinatti et al., 2020; Himsworth et al., 2013).
Since the beginning of the COVID-19 pandemic, several studies have been carried out in different countries in Europe, Asia, and America. These studies have provided serological evidence and confirm the presence of SARS-CoV-2 in synanthropic rodents, reporting prevalence rates ranging from 0.5% to 16.5% (Fisher et al., 2023; Lee et al., 2024; Martínez-Hernández et al., 2024; Miot et al., 2022; Orlando et al., 2024; Robinson et al., 2023; Tan et al., 2024; Wang et al., 2023). In Latin America, evidence of SARS-CoV-2 circulation in synanthropic rodents is geographically heterogeneous. Confirmed infections have been reported in Mexico City and Guayaquil, Ecuador (Martínez-Hernández et al., 2024; Orlando et al., 2024), while a low seroprevalence (0.87%) was detected in rural rodents from Chile (Grisolia et al., 2025) and no exposure was found in Brazil (Almeida-Souza et al., 2025). Collectively, these findings indicate a variable pattern of exposure across countries and landscapes, underscoring the need for further studies in diverse urban contexts throughout the region. Such heterogeneity, shaped by ecological, host-related, and socioeconomic factors, directly affects infection risks and should be considered in disease prevention strategies (Rothenburger et al., 2017).
Latin American countries were among the hardest hit by the COVID-19 pandemic, mainly due to their densely populated cities, lack of health infrastructure, and economic, health, and social inequities (Schwalb et al., 2022). The metropolitan area of Buenos Aires (MABA) was among the regions with the highest incidence of COVID-19 cases in Argentina (Ministerio de Salud, 2024). With an area of 3,833 km2 and 13,971,006 inhabitants, the MABA is the most densely populated area in the country (DAMI, 2021). It includes both urban and rural zones, with buildings, houses, and paved streets interspersed with parks, green spaces, and low-income neighborhoods, as well as crop and livestock fields (Cavia et al., 2009; DAMI, 2021).
Green spaces and low-income neighborhoods are the landscape units with the highest abundance of synanthropic rodents in Buenos Aires (Cavia et al., 2009). Green spaces are defined as parks or nature reserves for recreational or conservation purposes of more than 10 hectares that include public recreational areas and may include a mosaic of different vegetation types, including spontaneous vegetation of native flora, groves, or ornamental grasses (Cavia et al., 2009; Hancke and Suarez, 2018 ). On the other hand, low-income neighborhoods refer to areas inhabited by a very low-income population that lives in precarious dwellings with an inadequate supply of basic urban services, such as garbage removal, sanitation networks, electricity, telephones, and plumbing (Fernández et al., 2007; Hancke and Suárez, 2018). The probability of rodent-human contact varies across these units, being much higher in low-income neighborhoods compared to green spaces. Furthermore, low-income neighborhoods were particularly affected by the SARS-CoV-2 pandemic due to factors such as overcrowded living conditions, lack of reliable access to water, and essential urban services such as sanitation and waste disposal (Carbajo et al., 2023). These conditions not only encourage the establishment and growth of synanthropic rodent populations but also potentially increase the risk of zoonotic transmission (Battersby et al., 2002; United Nations, 2018). Due to the close association between humans and rodents in these areas, it is crucial to investigate the role of these animal species in the COVID-19 disease. Therefore, this study aimed to assess the presence of SARS-CoV-2 virus and antibodies in synanthropic rodents captured within two specific landscape units (low-income neighborhoods and green spaces) in the MABA.
The findings from this research will offer essential information for planning integrated disease prevention strategies against potentially zoonotic viruses, such as SARS-CoV-2, in urban environments.
Materials and Methods
Rodent trapping
A total of 24 trapping sessions (approximately one per month) were conducted between July 2020 and September 2022. Rodent sampling was carried out at 14 sites of two landscape units (7 low-income neighborhoods and 7 green spaces) from the MABA (Fig. 1). Green space sites included public parks and nature reserves ranging from 10 ha (site 11) to 70 ha (site 13), whereas low-income neighborhoods exhibited population densities between 3,717.1 and 50,424.4 inhabitants/km2 (sites 2 and 4, respectively, (De Grande and Salvia, 2019)). Sites were chosen based on accessibility, resident participation, and previous knowledge of rodent presence. Each site was sampled between one and four times, depending on accessibility, which was occasionally restricted by COVID-19-related lockdowns.
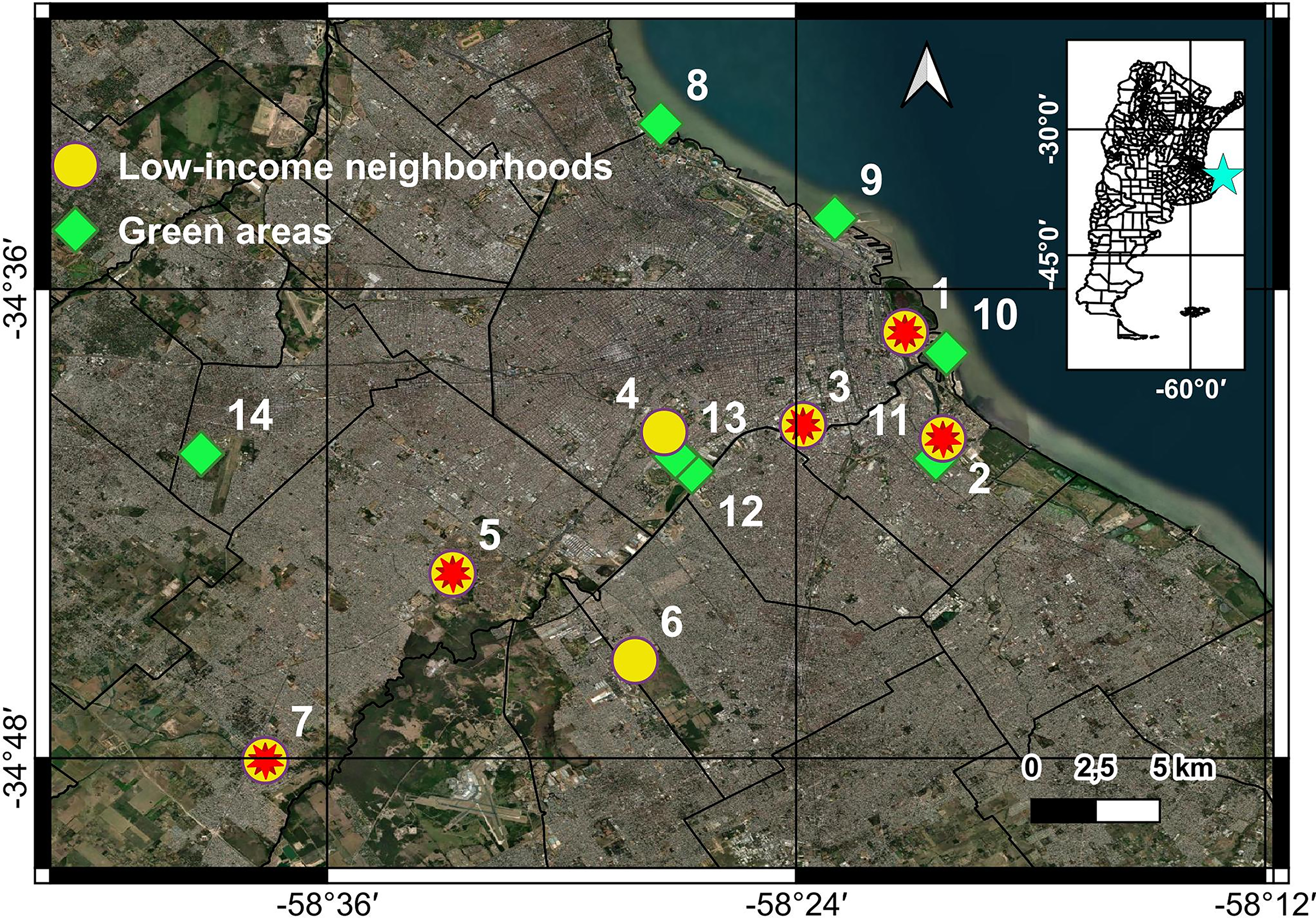
Location of the 14 sampling sites in the metropolitan area of Buenos Aires, Argentina. Green squares indicate green spaces, yellow circles represent low-income neighborhoods, and red stars mark the sites where rodents tested seropositive for SARS-CoV-2 antibodies. Basemap: Source: Esri, Maxar, Earthstar Geographics, CNES/Airbus DS, USDA, USGS, AeroGRID, IGN, and the GIS User Community.
At each site, single-capture live traps (Sherman and wire cage traps) were used, as described in Cavia et al. (2009). A 1:1 ratio of Sherman to cage traps was maintained across all sites. The total trapping effort per site varied depending on the area’s size and habitat complexity. In green spaces, traps were deployed along linear transects, spaced 5 meters apart, with an effort ranging from 232 to 556 trap-nights per site. In low-income neighborhoods, trapping effort ranged from 156 to 340 trap-nights per site. In these areas, one Sherman and one cage trap were placed per household, with the participation of residents who voluntarily agreed to the study. Sherman traps were set indoors (e.g., kitchens, bedrooms), and cage traps were set in adjacent outdoor spaces (e.g., yards or gardens). Sherman traps were baited with a mixture of peanut butter and beef fat, and cage traps with carrots and raw meat; they were monitored for four consecutive nights and checked every morning.
Trapped rodents were anesthetized using a combination of ketamine and acepromazine prior to sampling. Once sedated, rodents were weighed, measured, and assigned to species according to external characteristics (Gómez Villafañe et al., 2005). Serum, urine, and oropharyngeal and rectal swabs were collected. All samples were stored at −80°C until analysis. All synanthropic rodents captured were removed because tissues sampled were also collected for other parasitological studies. Rodents were handled strictly following biosafety norms and according to National Law No. 14346 of animal care (Protocol No. 156 approved by the Institutional Commission for the Care and Use of Laboratory Animals [CICUAL] of the Faculty of Exact and Natural Sciences, University of Buenos Aires).
RNA and RT-qPCR analysis of rodent’s tissue samples
RNA extraction and RT-qPCR assays were performed on urine and serum samples collected directly, as well as on oropharyngeal and rectal swabs collected using the Copan Universal Transport Medium (UTM-RT®) System. RNA was extracted with the QIAamp Viral RNA Mini Kit (QIAGEN), following the manufacturer’s instructions under biosafety level-2 (BSL-2) conditions. RT-qPCR was performed on each sample using the commercial Detection Kit for 2019 Novel Coronavirus (2019-nCoV) RNA (Da An Gene Co., Ltd., Sun Yat-sen University). The assay targets the N and ORF1ab genes of SARS-CoV-2, and the kit’s performance had been previously verified by the National Reference Laboratory for Influenza, SARS-CoV-2, and Other Respiratory Viruses (Servicio Virosis Respiratorias, INEI-ANLIS “Dr. Carlos G Malbrán,” Fellner et al., 2021). The positivity criteria were applied according to the manufacturer’s instructions.
Amplification of a β-actin gene fragment by in-house qPCR was used as an internal control for each individual sample to assess RNA integrity, using the primers and probes previously reported by Wakeley et al. (2005). Primers and probes were used at a working concentration of 10 µM with the qScript XLT One-Step RT-qPCR ToughMix enzyme (Quantabio). The reaction mix was prepared according to the manufacturer’s instructions. Amplification was performed on a CFX96 Touch Real-Time PCR Detection System (Bio-Rad) under the following cycling conditions: 1 minute at 95°C, followed by 45 cycles of 95°C for 10 s and 60°C for 35 s.
Serum samples with volumes less than 100 µL were reserved exclusively for serological testing.
Preparation of slides with SARS-CoV-2-infected cells
Slides containing SARS-CoV-2-infected cells of the ancestral Wuhan variant were prepared under BSL-3 laboratory conditions. The viral isolate was obtained using the Vero cell line (ATCC CCL-81), originally derived from a nasopharyngeal swab sample (CoV-19/Argentina/C121/2020/EPI_ISL_420600/2020-03-07), at a concentration of 1,000,000 cells/mL. The isolate was mixed in equal proportions with uninfected Vero cells with the same concentration, and 20 µL of the mixture were applied to an immunofluorescence microscope slide, air-dried, and fixed with cold glacial acetone to preserve cellular morphology and viral antigens.
Serology
Indirect immunofluorescence was performed to detect IgG antibodies against SARS-CoV-2 in rodent serum samples. Serum samples were diluted 1:100 and incubated on tissue imprints at 37°C for 30 min. After washing, a fluorescein isothiocyanate (FITC)-labeled secondary antibody, either antirat IgG or antimouse IgG (Sigma-Aldrich®), was applied at a 1:300 dilution and incubated again at 37°C for 30 min. The slides were then examined under a fluorescence microscope. Samples showing two or more fluorescent cells per 40X field were considered reactive for SARS-CoV-2 IgG antibodies (Fig. 2, Agusi et al., 2024; Perez et al., 2005).
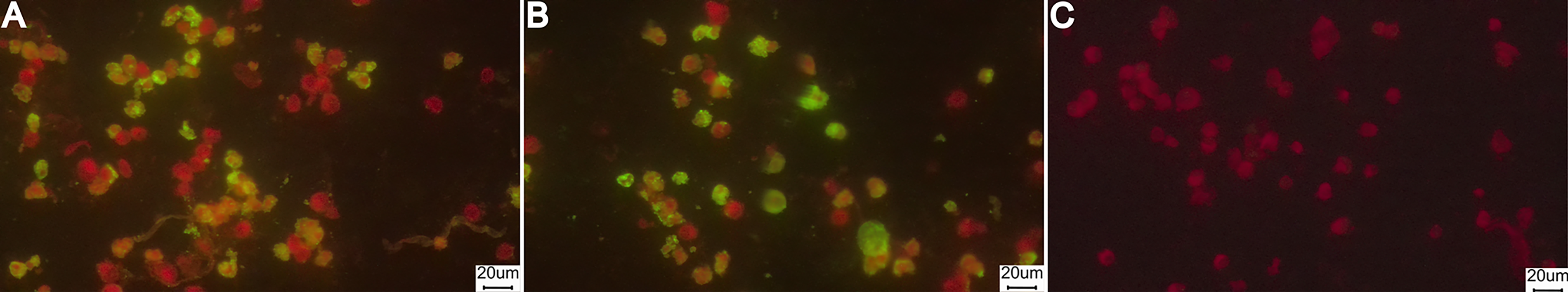
Representative images of indirect immunofluorescence assays for SARS-CoV-2 IgG detection in rodent serum samples.
To validate the assay’s sensitivity and specificity, a positive control (human serum reactive for SARS-CoV-2 IgG by Enzyme-Linked Immunosorbent Assay (ELISA)) and a negative control (human serum nonreactive for SARS-CoV-2 IgG by ELISA technique) were included using the same dilution conditions. A FITC-labeled secondary antibody against the Fc portion of human IgG was used to detect the presence of SARS-CoV-2 antibodies. Both controls demonstrated expected reactivity and nonreactivity respectively, confirming the assay’s accuracy.
In addition, serum samples from 34 urban rodents (9 M. musculus, 13 R. rattus, and 14 R. norvegicus) captured at six different sites in the same landscape units in the prepandemic period (between July 2018 and November 2019) were used as a negative control of the technique.
Statistical analysis
Seroprevalence was calculated as the proportion of rodents testing positive for IgG antibodies against SARS-CoV-2, and 95% confidence intervals (CIs) were estimated using the Clopper–Pearson exact method, appropriate for binomial data with small sample sizes. To evaluate differences in seroprevalence between landscape units and sampling periods, Fisher exact test was applied due to the presence of small cell counts and zero values in some groups. All statistical analyses were performed using R software (version 4.4.2), and exact confidence intervals were calculated using the GenBinomApps package version 1.2.1.
Results
We caught a total of 139 synanthropic rodents, corresponding to 75 R. norvegicus, 23 R. rattus, and 41 Mus musculus. Of these, 125 individuals were tested for SARS-CoV-2 IgG antibodies using indirect immunofluorescence; the remaining 14 could not be analyzed due to the absence of serum samples. Antibodies were detected (Fig. 2) in 8.8% (3.3–18.2%) of R. norvegicus and 5.7% (0.7–19.2%) of M. musculus analyzed. All seropositive animals were captured in 5 low-income neighborhoods over the 3-year study (Fig. 1, Table 1). No positive results for SARS-CoV-2 antibodies were observed in samples collected before the COVID-19 pandemic or in green spaces. Fisher exact test indicated a trend toward a higher probability of SARS-CoV-2 seropositivity in rodents from low-income neighborhoods compared to other groups (p-value = 0.060). In addition, all urine and serum samples, as well as oropharyngeal and rectal swabs from the 139 rodents, tested negative for SARS-CoV-2 virus RNA by RT-qPCR. The β-actin gene was detected in all tested samples, indicating acceptable sample quality.
Number of Rodents Captured and Seropositive for IgG Antibodies Against SARS-CoV-2, Detected by Indirect Immunofluorescence, by Year, Rodent Species, and Landscape Unit in the Metropolitan Area of Buenos Aires, Argentina
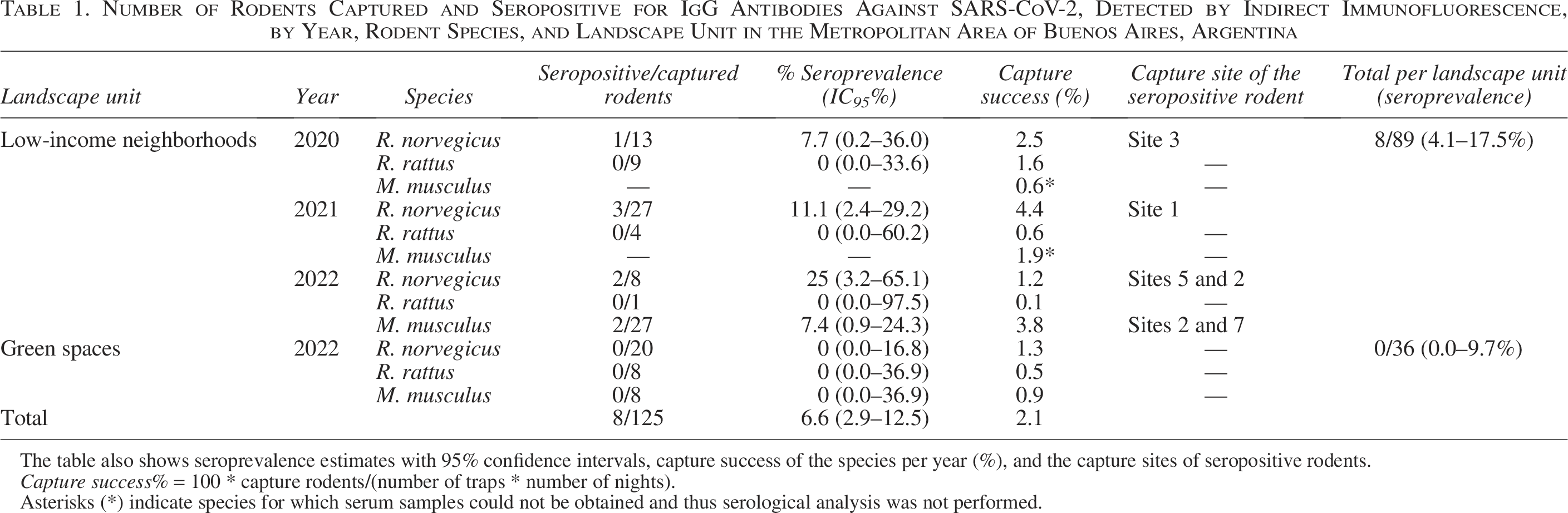
The table also shows seroprevalence estimates with 95% confidence intervals, capture success of the species per year (%), and the capture sites of seropositive rodents.
Capture success% = 100 * capture rodents/(number of traps * number of nights).
Asterisks (*) indicate species for which serum samples could not be obtained and thus serological analysis was not performed.
Discussion
This study provided valuable information on the exposure of synanthropic rodents to SARS-CoV-2 in two landscape units of a Latin American city, with seropositive animals detected in low-income neighborhoods but not in green spaces during our study. Although statistical support for differences between landscape units was marginal (Fisher exact test, p = 0.060) and should therefore be interpreted with caution, our findings point to possible spatial variation in human–rodent interactions and associated spillover risk across urban landscapes. A previous study conducted by Carbajo et al. (2023) in Buenos Aires City during the first half of 2020 indicated that 51.4% of COVID-19 cases were among residents of low-income neighborhoods. This spatial distribution of COVID-19 cases reflects the challenge of maintaining physical distancing in the densely populated conditions typical of these areas (Carbajo et al., 2023). In such environments, where human exposure to SARS-CoV-2 is high and rodents are commonly found near dwellings, transmission of the virus from humans to rodents could be facilitated through direct contact with infected individuals or indirect contact with contaminated environments (Bosco-Lauth et al., 2021; Franklin and Bevins, 2020; Gryseels et al., 2021; Hale et al., 2022; Orlando et al., 2024; Robinson et al., 2023). However, the specific routes of exposure to SARS-CoV-2 in these rodents remain unclear, and the possibility of transmission of the virus between rodents has not been established. Contaminated food waste, wastewater, urban runoff, and general household waste (such as used tissues, face masks, and other items contaminated with bodily fluids) may serve as potential transmission routes to synanthropic rodents, considering that SARS-CoV-2 can be transmitted via droplets, aerosols, fomites, and the fecal–oral route, among others (Alberto-Orlando et al., 2022; Liu et al., 2021; Orlando et al., 2024; Robinson et al., 2023; Sherchan et al., 2023). In turn, the lack of serological evidence of SARS-CoV-2 in green spaces could be explained by the lower human activity in these spaces, since the social gatherings and circulation of people in public areas were limited by the measures imposed by the government aimed at mitigating the effects of the pandemic and protecting public health (https://www.boletinoficial.gob.ar/detalleAviso/primera/241290/20210228). However, due to the small sample size and absence of statistical significance, cautious interpretation of this result is warranted. Notably, although capture success for Rattus spp. was comparable between green spaces and low-income neighborhoods in 2022, rodent population dynamics in green areas were likely influenced by pandemic-related lockdowns during 2020–2021. Reduced human activity and food waste availability during that period likely contributed to a marked decline in rodent abundance, thereby limiting sampling opportunities in green spaces during the early phase of the study.
The positivity rates for SARS-CoV-2 detection by serology found in this study were 12.5% in R. norvegicus and 7.4% in M. musculus from low-income neighborhoods. These results were consistent with previous research carried out in several cities in America, Europe, and Asia, which reported seroprevalences ranging from 0.5% to 16% in species such as R. norvegicus, R. rattus, M. musculus, and Sundamys muelleri (Almeida-Souza et al., 2025; Fisher et al., 2023; Grisolia et al., 2025; Lee et al., 2024; Martínez-Hernández et al., 2024; Miot et al., 2022; Orlando et al., 2024; Robinson et al., 2023; Tan et al., 2024; Wang et al., 2023). These findings suggested that murine rodent populations in different geographic regions have experienced similar levels of exposure to SARS-CoV-2. Some recent studies have suggested that the SARS-CoV-2 omicron variant might have adapted to mouse ACE2 receptors before infecting humans, suggesting a rodent origin of this variant (Wei et al., 2021; Zhang et al., 2022). Furthermore, recent experimental studies demonstrated that rats inoculated with SARS-CoV-2 developed specific antibodies, despite the absence of detectable viral replication (Beissat et al., 2025). These findings suggest that while rodents are capable of mounting an immune response upon exposure, their potential to maintain and transmit the virus under natural conditions remains uncertain. Therefore, further experimental and field investigations are required to clarify the role of urban rodent populations in the circulation and potential maintenance of SARS-CoV-2.
Although our positive serology findings could potentially be due to nonspecific antibody binding or cross-reactivity, the absence of seropositive results in rodents captured in the pre-COVID-19 period (2018–2019) and in areas with low human activity (green spaces) during the study period (COVID-19 pandemic) may suggest an association between seropositivity and areas of high human–rodent interaction during the pandemic. While we acknowledge that the sample sizes for these groups were relatively small and that the serological assay was validated using human control sera rather than rodent-specific positive controls, the observed spatial pattern of seropositivity (restricted to rodents from low-income neighborhoods), combined with consistent findings from studies in other countries, strengthens the interpretation that these rodents were indeed exposed to SARS-CoV-2 under natural conditions.
Circulation of different SARS-CoV-2 variants in Argentina during the study period could have influenced the exposure dynamics in rodents. The ancestral Wuhan strain predominated in 2020, followed by the emergence of Alpha and Gamma variants in early 2021, Delta in mid2021, and Omicron from late 2021 onward (https://www.argentina.gob.ar/salud/epidemiologia/vigilancia-epidemiologica/reportes-sobre-covid-19-en-argentina). While our serological assay was based on the ancestral strain, cross-reactivity with antibodies induced by other variants has been demonstrated (Vinzón et al., 2023), supporting its ability to detect responses to diverse viral lineages. In turn, the detection of seropositive rodents between 2020 and 2022 showed an increasing trend in seroprevalence over time: 4.5% in 2020, 9.7% in 2021, and 12.1% in 2022 in low-income neighborhoods. This temporal pattern appears to follow the overall increase in reported human cases of COVID-19 in the same region during those years (853,000 in 2020, 1,929,229 in 2021, and 3,133,558 cases in 2022), suggesting that rodent exposure may have occurred during periods of intense viral circulation and variant renewal in the human population. While this observation suggests a possible relationship between viral circulation in humans and rodent exposure, further studies are needed to fully evaluate this association.
The main limitation of this study was the nondetection of viral RNA in any of the rodents analyzed, so it was not possible to identify the specific SARS-CoV-2 variant to which these animals were exposed. Similar results had been reported in synanthropic rodents from urban areas in Spain, Japan, Germany, Belgium, Malaysia, Canada, and the United States, where tissue samples or swabs from rodents tested negative for SARS-CoV-2 RNA (Colombo et al., 2022; Fernández-Bastit et al., 2024; Kuroda et al., 2024; Lee et al., 2024; Robinson et al., 2023; Tan et al., 2024; Wernike et al., 2022). To date, only three studies have confirmed SARS-CoV-2 infection in rats from New York, Mexico City, and Guayaquil, reporting positivity rates for rats between 5% and 6% (Martínez-Hernández et al., 2024; Orlando et al., 2024; Wang et al., 2023). The RT-qPCR methodology used in our study, which targeted the N, E, and ORF1ab genes, was consistent with these investigations, assessing viral presence in animal fluids and swabs. Thus, the lack of detectable viral RNA in our study may be attributed to the timing of sample collection in relation to the progression of the infection or to variations in viral load in different tissues and fluids (Robinson et al., 2023). Moreover, the detection of the β-actin gene in all the samples and the lack of SARS-CoV-2 gene detection by Rt-qPCR could be considered true negative SARS-CoV-2 results because of the low detection limit of the RT-qPCR used for the detection of the viral genes, similar to the method recommended by WHO, 2020, and the confirmation of integrity of the rodent samples by the β-actin gene detection. These findings contribute to a broader understanding of SARS-CoV-2 dynamics in urban wildlife and highlight the importance of combining serological and molecular tools in zoonotic surveillance.
Our findings provide preliminary evidence of SARS-CoV-2 exposure in urban rodents from low-income neighborhoods in Buenos Aires. Broader studies are required to assess whether rodent exposure differs across other socio-economic and landscape contexts. The results of this study highlight the need for continued monitoring to detect the circulation of SARS-CoV-2 in urban rodent populations, especially since they could harbor new variants. This is critical in low- and middle-income countries, where low-income neighborhoods have reported an elevated number of COVID-19 cases along with high rodent abundance. Preventive measures, such as rodent control, waste management, and public health education in these areas, could mitigate the risk of transmission of zoonotic disease. Ultimately, an integrated One Health approach is vital to reducing the likelihood of future outbreaks and pandemics.
Authors’ Contributions
M.A.T.: Conceptualization, data curation, methodology, project administration, and writing—original draft, review, and editing. M.E.D.: Data curation, formal analysis, methodology, and writing—original draft, review, and editing. E.M.: Conceptualization, data curation, methodology, and writing—review, and editing. A.L.P.: Data curation, methodology, and writing—review and editing. M.D.M.: Data curation, methodology, and writing—review and editing. M.P.N.: Data curation, methodology, and writing—review and editing. D.H.: Conceptualization, data curation, methodology, and writing—review and editing. E.B.: Conceptualization, formal analysis, funding acquisition, methodology, and writing—review and editing. O.V.S.: Conceptualization, funding acquisition, formal analysis, methodology, project administration, and writing—review and editing.
Footnotes
Acknowledgments
The authors thank the team of the Laboratorio de Ecología de Roedores Urbanos for their assistance during the field sampling.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
Financial support was provided by the University of Buenos Aires, Gobierno de la Ciudad de Buenos Aires, Autoridad de la Cuenca Matanza Riachuelo (ACUMAR), and CONICET.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.