
Abstract
Background:
Leishmania major and Leishmania infantum infection were evaluated in sand flies and rodents during a cutaneous leishmaniasis (CL) outbreak in two military camps in southern Israel (2022–2023). The efficacy of a novel intervention combining drone-based pesticide application and rodent bait stations was assessed.
Methods:
Sand flies and rodents were sampled and tested for Leishmania by PCR and DNA sequencing. In April 2023, precision spreading of pesticides via drones and bait stations targeting Meriones tristrami and Mus musculus domesticus was done in camp 2, while camp 1 served as control. Leishmania infection rates in vectors and reservoirs were compared before and after intervention, alongside CL incidence in soldiers.
Results:
A total of 7,204 female Phlebotomus papatasi were analyzed. The minimal infection rate of L. major in camp 2 declined from 1.23 in 2022 to 0.03 post-iinterventionin 2023 (p = 0.034). In camp 1, no significant change occurred (0.50 in 2022 vs. 0.22 in 2023, p = 0.622). Among 63 rodents, 8 (12.7%) were infected: five with L. infantum (2/11 M. musculus domesticus [18.2%] and 3/52 M. tristrami [6%]) and three M. tristrami (6%) with L. major. Overall rodent infection declined significantly from 24.1% in 2022 to 2.9% in 2023 (p = 0.032), with camp 2 showing the most pronounced reduction (31.6% vs. 3.3%, p = 0.02). Human CL incidence also dropped significantly: in camp 1, from 17.1 to 2.6/1000, and in camp 2, from 33.3 to 4.9/1000 (p < 0.001).
Conclusions:
Significant reductions in Leishmania infection were observed in vectors, reservoirs, and humans following the intervention. Although CL dynamics are influenced by multiple factors, these findings support the potential of drone-based and host-targeted approaches for integrated leishmaniasis control. Further studies are required to validate and optimize such technologies.
Keywords
Introduction
Cutaneous leishmaniasis (CL), caused primarily by Leishmania major and Leishmania tropica, is endemic in Israel and throughout the Middle East (Ruiz Postigo, 2010). The disease is transmitted by the bite of infected female Phlebotomus sand flies and causes skin ulcers that may result in permanent scars (Mann et al., 2021). In Israel, Phlebotomus papatasi is the vector for L. major, while P. sergenti and P. arabicus transmit L. tropica (Jaffe et al., 2004). Reservoir hosts for L. major in Israel include rodents such as Psammomys obesus, Meriones crassus, Gerbillus dasyurus, and Meriones tristrami (Jaffe et al., 2004; Faiman et al., 2013). The primary reservoir for L. tropica is the rock hyrax (Procavia capensis) (Talmi-Frank et al., 2010a). Leishmania infantum is the causative agent of visceral leishmaniasis in Israel, and dogs and wild canines are its main reservoirs (Baneth et al., 1998). L. tropica and L. infantum are present mainly in the central and northern parts of Israel, while L. major is prevalent in the arid southern parts of the country (Schlein et al., 1984; Jacobson et al., 2003; Jaffe et al., 2004; Schnur, et al., 2004).
CL is a notifiable disease both in the Israeli Ministry of Health (IMOH) and in the Israel Defense Forces (IDF). However, the IDF conducts active surveillance during outbreaks, and the IMOH system is based on passive reporting only (Mimouni et al., 2009). Incidence among IDF soldiers is historically much higher than in the civilian population due to being stationed in hyperendemic regions, their younger age, and being likely naïve to the parasite. Between 2008 and 2024, CL incidence among soldiers peaked at 242.4/100,000 (2017), later rising again to 178.3/100,000 by 2024 (latest data retrieved on January 02, 2025, information department of the Israeli Medical Corps [IMC], IDF). In contrast, civilian incidence peaked at 4.7/100,000 in 2014 and declined to 1.7/100,000 in 2024 (https://www.gov.il/he/Departments/DynamicCollectors/weekly-epidemiological-report?skip=0). The number of civilian cases reported to the IMOH is lower compared with reports obtained by clinically and laboratory-confirmed cases in Israeli hospitals during 2017–2022, underlining the extent of CL underreporting by passive surveillance (Avni et al., 2024).
In recent years, CL has reemerged in the northwestern Negev desert, particularly near Ofakim city and surrounding agricultural communities. Two epidemic waves occurred in this region’s military bases: the first between 2011 and 2015, and the second starting in 2021. Civilian outbreaks paralleled these trends (Ben-Shimol et al., 2016; Orshan et al., 2016; Kleinerman et al., 2023).
This study aimed to monitor the dynamics of Leishmania infection in sand fly vectors and rodent reservoirs, map the geographic distribution of L. major and its hosts, and assess the effectiveness of a drone-assisted intervention program targeting reservoir rodents over a 2-year period in two military camps in the northwestern Negev. Additionally, monthly CL case reports were analyzed to explore potential associations between vector dynamics and human infections.
Materials and Methods
Study area
The study was conducted in two adjacent military camps located near Kibbutz Urim in the northwestern Negev desert, a semiarid agricultural area with seasonal crops such as wheat, potatoes, peanuts, carrots, and melons. Local wildlife fauna includes Meriones tristrami, Mus musculus domesticus, Erinaceus concolor, Hemiechinus auritus, Herpestes ichneumon, Vulpes vulpes, Canis aureus, and Felis catus. In camp 1, only 22% of soldiers reside full time, while in camp 2, all are permanent residents, increasing their exposure to local infected sand fly populations.
Human CL incidence
Monthly and annual incidence rates of CL among soldiers in both camps from 2016 to 2024 were obtained from the IMC information department.
Sand fly collection and identification
Sand flies were collected monthly from May to November 2022 and June to November 2023 using modified Centers for Disease Control and Prevention (CDC) traps (Orshan et al., 2016). Traps were powered by AA batteries, baited with dry ice (∼1.5 kg), and set overnight at consistent locations near camp perimeters (next to agricultural fields). In 2022, 12 traps per camp were used initially, then reduced to 6–8 (each month). In 2023, 6–8 traps per camp were used monthly. A total of 200 traps were deployed at 38 sites across 26 nights.
In the IMOH, collected flies were chilled, counted, and sorted. Male specimens were identified morphologically by genitalia and pharynx (Abonnenc, 1972; Lewis, 1982). Females were identified molecularly using HRM-PCR targeting the cytochrome b gene (Studentsky et al., 2023). Female sand flies from large catches (>100 specimens) were pooled (≤20 specimens/pool) for Leishmania detection. Samples were stored at −20°C until testing.
Rodent trapping and sampling
Rodents were trapped during five field sessions between 07/2022 and 11/2023 using Sherman live traps (Tallahassee, FL) baited with peanut snacks (Bamba; Osem Ltd., Holon, Israel). Traps were placed 15–20 m apart near burrows inside and outside the camps. Only M. tristrami and M. musculus domesticus were retained; other species of animals, if trapped, were released. Euthanasia was performed following the American Veterinary Medical Association 2020 guidelines, using 10% isoflurane (Piramal, Bethlehem, PA) via the open-drop method (https://www.avma.org/sites/default/files/2020-02/Guidelines-on-Euthanasia-2020.pdf). Death was confirmed by a veterinarian.
Tissue samples were taken from the ear pinna, snout, and lesions. Blood was collected on FTA collection QIAcard (Qiagen, Venlo, The Netherlands), and exfoliating cells from the conjunctiva were taken with sterile swabs. If lesions were present, touch impressions were taken, stained (Diff-Quick; Kaltek srl, Padova, Italy), and examined microscopically (×400). Tissue was split for histology (in 4% paraformaldehyde) or frozen for DNA extraction. Rodent metadata (species, sex, age group, lesion presence) were recorded. Protocols were approved by the IDF animal ethical committee (Approval 02–2023).
DNA extraction and Leishmania detection
DNA was extracted from rodent blood (DNeasy; Qiagen), swabs (QuickDNA Miniprep Plus Kit; Zymo, Orange, CA), and tissue (Exgene Cell SV; GeneAll, Biotechnology Co., Ltd., Seoul, South Korea). Sand fly DNA was extracted using the QIAsymphony DNA Mini Kit and SP machine (Qiagen N.V., Venio, The Netherlands). Detection of Leishmania DNA targeted a 265-bp fragment of the ITS1 region using real-time PCR and High-Resolution Melting (HRM) analysis (Talmi-Frank et al., 2010b), with StepOnePlus (Applied Biosystems) for rodent DNA samples and LightCycler 96 (Roche) for sand fly DNA. Positive controls used L. infantum promastigote DNA; negative controls were nontemplate controls. Amplicons were purified (Exo-Sap, NEB; New England Biolabs, Inc., Ipswich, MA) and Sanger-sequenced at the Hebrew University. Identity was confirmed by Basic Local Alignment Search Tool (BLAST) analysis (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
Cytology and histopathology
Cytological evaluation of stained skin touch impressions was made using the Nikon Eclipse 200 microscope. Histopathology of rodent tissue was performed at the Weizmann Institute of Science Department of Veterinary Resources (Rehovot, Israel).
Rodent control intervention (camp 2 only)
In April 2023, rodent control in camp 2 combined drone-based pesticide application and bait station distribution. Drone delivery was done in several steps: first, agricultural areas surrounding the camp were scanned and mapped using a DJI Phantom 4 RTK drone (Shenzhen, China); second, burrow locations were identified and plotted using Tantos software (ALTA, Nahalal, Israel); finally, sodium fluoroacetate (Rosh 80®, Rimi, Israel) was dispersed directly over burrows using the Gaya® drone system (Regev et al., 2019), minimizing environmental impact. The entire process was supervised by the Plant Protection and Inspection Services of the Israeli Ministry of Agriculture. In addition, 95 bait stations containing brodifacoum 0.005% (Blokion Plus®; RPC Ltd, Nes Tziona, Israel) were distributed throughout the camp and replenished monthly following manufacturer instructions.
Data analysis
In order to evaluate the level of Leishmania infection in the sand flies, we calculated the minimal infection rate (MIR), which assumes one infected female sand fly per positive pool (Katholi and Unnasch, 2006), and was obtained as:
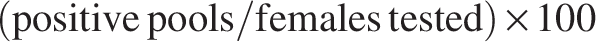
MIR was selected as a conservative surveillance metric consistent with national vector-borne disease monitoring practices, allowing comparison with historical datasets.
MIRs were compared by camp and year using the Mann–Whitney U test. In addition, the estimated risk of exposure index (EREI), assuming that only one vector sand fly species was present in the area (Kilpatrick et al., 2005), was calculated as:
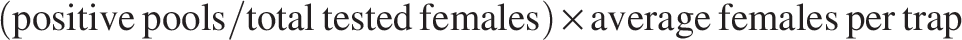
Associations between EREI, MIR, and monthly CL cases (lagged 1–6 months) were assessed using the Spearman correlation test. Rodent infection rates were compared between camps and years using the Pearson chi-squared test. CL incidence rates between 2022 and 2023 were compared using the Fisher exact test. Analyses were conducted in SPSS v29 (IBM, New York, NY).
Results
Human CL cases
Data on CL cases in the two military camps were collected per year and month between 2016 and 2024 (last retrieved January 02, 2025, from the IMC information department). Annual incidence ranged from 0 to 17.1/1000 in camp 1 and 0 to 33.3/1000 in camp 2. In camp 1, incidence rose from 2.1/1000 in 2016 to 17.1/1000 in 2022, then declined to 2.6/1000 in 2023 and slightly increased to 5.1/1000 in 2024. In camp 2, incidence increased from 0/1000 in 2016 to 33.3/1000 in 2022, dropped to 4.9/1000 in 2023, and rose again to 11.7/1000 in 2024 (Fig. 1). Across the study period, 61% of camp 1 cases and 67% of camp 2 cases were diagnosed between November and February. A significant reduction in incidence was observed from 2022 to 2023 in both camps, from 17.1 to 2.6/1000 in camp 1 and from 33.3 to 4.9/1000 in camp 2 (Fisher’s exact test, p < 0.001).
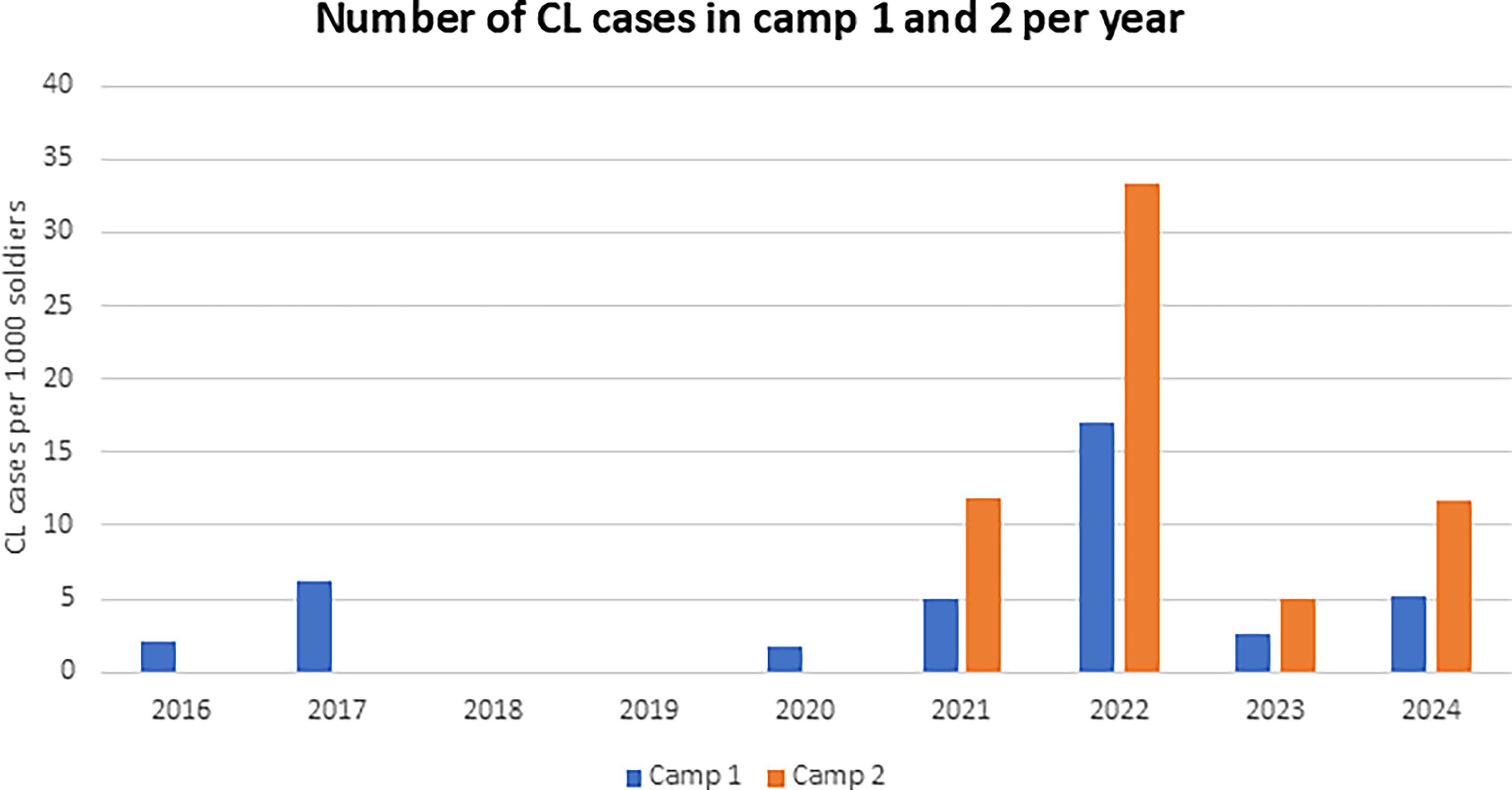
Number of annual human cutaneous leishmaniosis cases per 1,000 soldiers in camps 1 and 2 from 2016 to 2024.
Sand flies
A total of 12,083 sand flies (9,696 females, 2,387 males) were collected in 2022–2023 across 26 nights at 25 sites using 158 traps (including only trapping events yielding sand flies). Sampling occurred monthly from April to November 2022 and June to November 2023 (excluding October). Ph. papatasi was the only species detected. Of 9,696 females, 7,204 (74.3%) were PCR-tested, with L. major DNA identified in 37/424 pools from 14 sites.
Sand flies in camp 1
In 2022, a total of 6,313 Ph. papatasi sand flies (5,042 females and 1,271 males) were trapped over 8 nights at 11 sites using 52 traps. The number of specimens per catch ranged between 0 and 570. Of the 5,042 females collected, 3,470 were tested, and L. major was detected in 18 pools from six sites out of 232 pools analyzed. Female abundance peaked in August, whereas the maximum MIR occurred in October and the highest EREI in September (Table 1). During 2023, collections decreased markedly, with only 1,586 specimens (1,214 females and 372 males) obtained across 8 sites and 5 nights using 31 traps, ranging from 0 to 243 specimens per catch. Among 1,214 females, 1,117 were tested, and L. major was detected in only 3 pools from 2 sites out of 63 pools. Female abundance peaked in August; however, the highest MIR was recorded in June, while the EREI reached its maximum in September (Table 1).
Average Number of Phlebotomus papatasi Females Collected, Minimal Infection Rate, and Estimated Risk of Exposure Index per Month per Camp
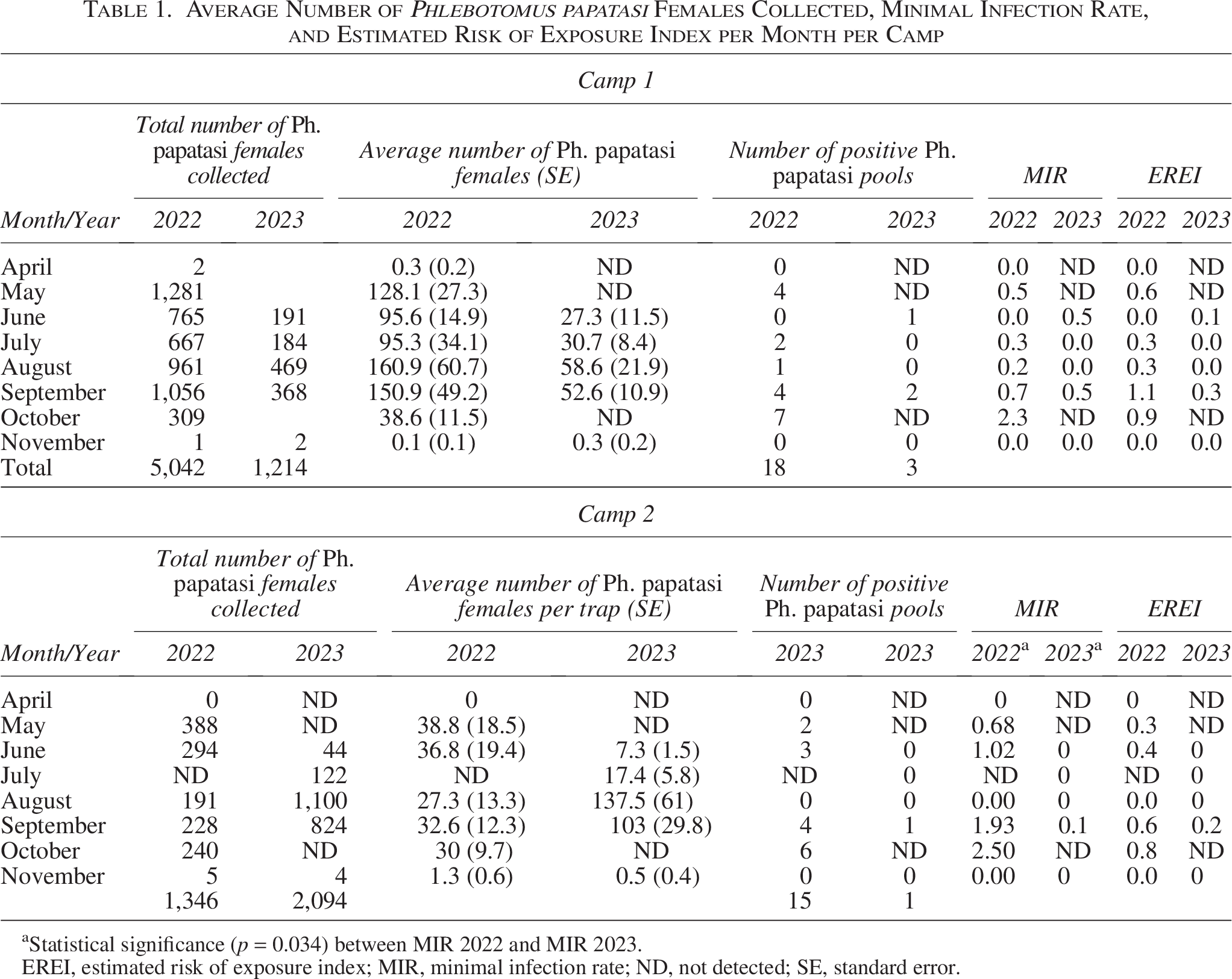
Statistical significance (p = 0.034) between MIR 2022 and MIR 2023.
EREI, estimated risk of exposure index; MIR, minimal infection rate; ND, not detected; SE, standard error.
Sand flies in camp 2
In 2022, 1,560 Ph. papatasi sand flies (1,346 females and 214 males) were collected during 8 nights at 14 sites using 44 traps, with catches ranging from 0 to 217. Out of the 1,346 females trapped, 1,226 were tested, and L. major was identified in 15 pools from 6 sites out of 49 pools tested. Female abundance was highest in May, while both MIR and EREI peaked in October (Table 1). In 2023, the number of Ph. papatassi sand flies increased to 2,624 specimens (2,094 females and 530 males) collected at 9 sites over 5 nights using 31 traps, ranging from 0 to 618 per catch. Of the 2,094 females, 1,391 were tested, and only a single pool was positive out of 80 pools analyzed. The corresponding MIR and EREI were 0.15 and 0.2, respectively. Abundance reached its peak in August, and the single positive pool was detected in September (Table 1).
Analysis of sand fly data
In camp 2, the average MIR declined significantly from 1.23 in 2022 to 0.03 in 2023 (Mann–Whitney U, p = 0.034). No significant change was seen in camp 1 (0.50 in 2022 vs. 0.22 in 2023, Mann–Whitney U p = 0.6). In camp 2, MIR (p = 0.045) and EREI (p = 0.023) correlated (Spearman correlation test) with soldier CL cases with a 3-month delay, but no correlation was observed in camp 1 (p > 0.05). Figures 2 and 3 show the geographical distribution of sand fly collections, positive sites, and their proximity to the camps. Most sand flies were trapped outside camp boundaries, at agricultural field margins.
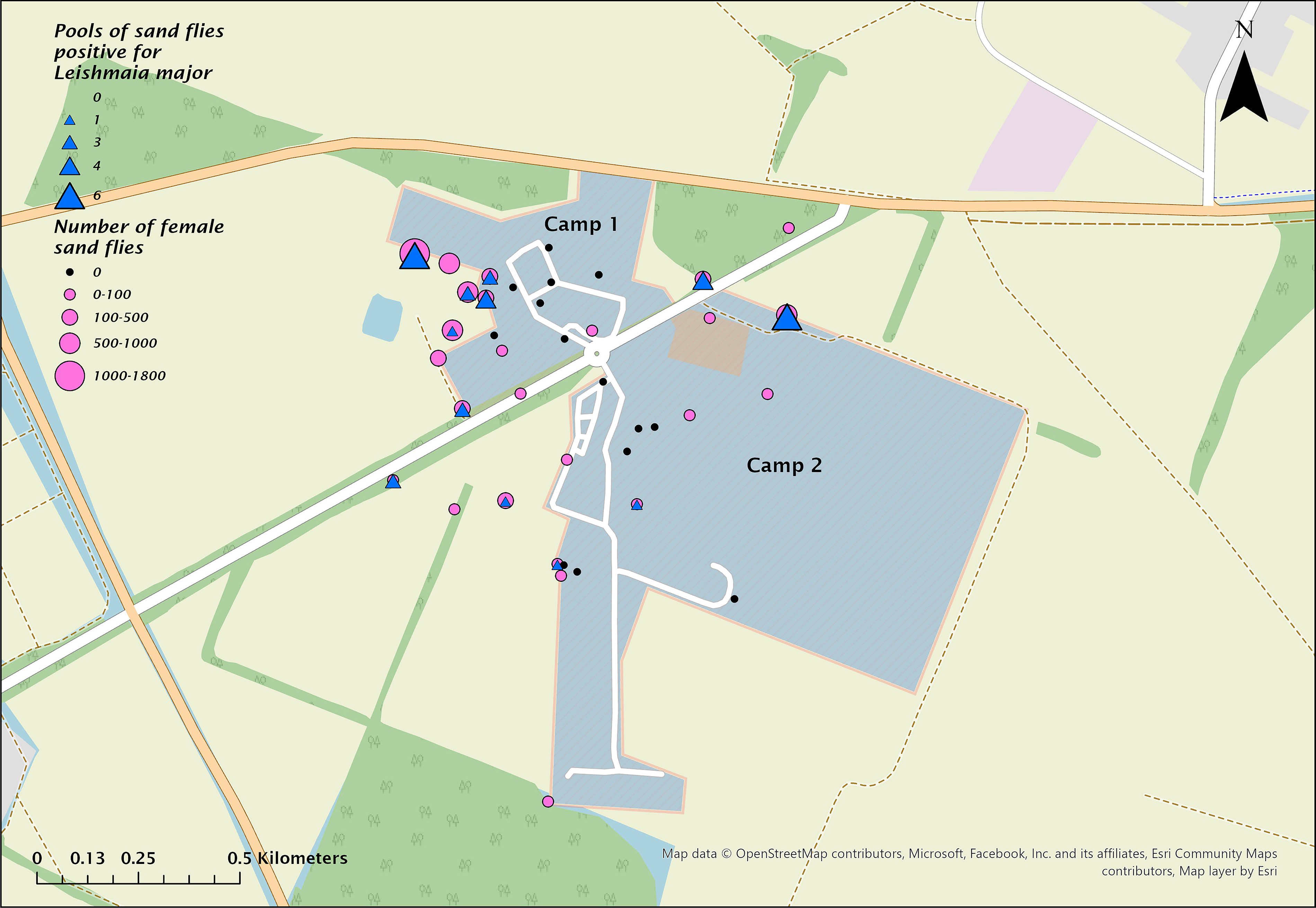
Map of camps 1 and 2 showing the dispersion of Phlebotomus papatasi sand flies trapped and the location of Leishmania major infected pools (blue triangles) during 2022.
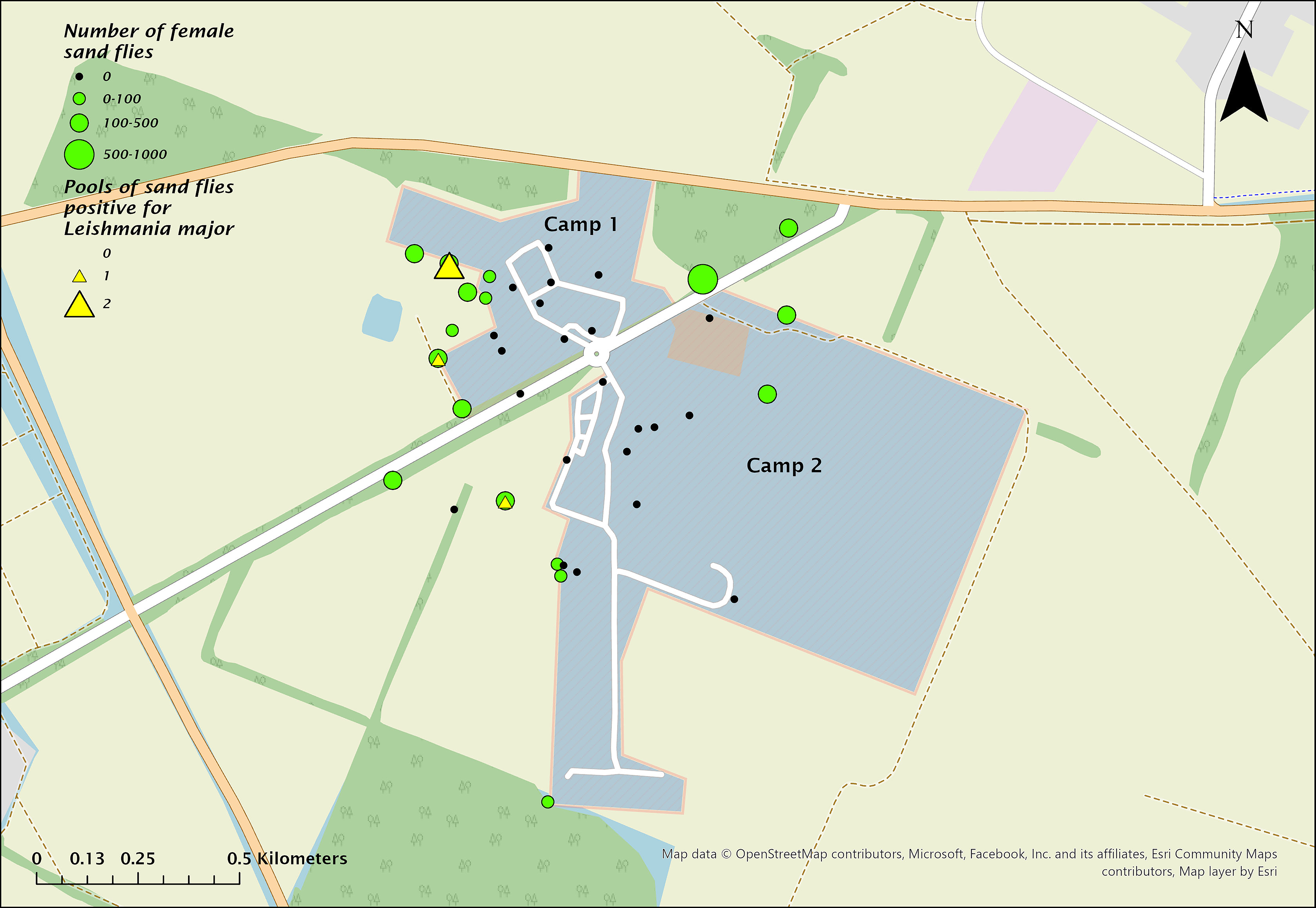
Map of camps 1 and 2 showing the dispersion of Phlebotomus papatasi sand flies trapped (green circles) and the location of Leishmania major infected pools (yellow triangles) during 2023.
Environmental intervention using drones and bait stations in camp 2
A total of 220 hectares of agricultural fields (potato, carrot, and other crops) surrounding camp 2 were surveyed and treated with sodium fluoroacetate (Rosh 80) in compliance with pesticide regulations under the supervision of the Plant Protection and Inspection Services, Ministry of Agriculture. Rodent burrows, paths, and crop damage were first identified (Supplementary Fig. S1), plot mapping and analysis were performed, and finally, bait distribution in each marked plot was done using the Gaya system drone (Supplementary Fig. S2).
In addition, 96 bait station traps containing brodifacoum 0.005% (Blokion Plus) were distributed inside camp 2. Rodent trapping data indicated that in camp 1, in 2022, 10 rodents were caught outside the limits of the camp, and in 2023, 3 were caught outside. In camp 2, in 2022, 18 rodents were trapped outside and 1 inside the camp. By contrast, in 2023, after the intervention, 31 rodents were trapped outside, but none were found inside the camp.
Rodents
A total of 63 rodents were trapped and sampled; the species found were M. tristrami (n = 52) and M. musculus domesticus (n = 11). Twenty-nine were sampled in 2022 and 34 in 2023. In camp 1, 10 were sampled in 2022 and 4 in 2023; in camp 2, 19 in 2022 and 30 in 2023 (Table 2).
Number, Species of Rodents, and Leishmania spp. Infection in Camps 1 and 2 in 2022 and 2023
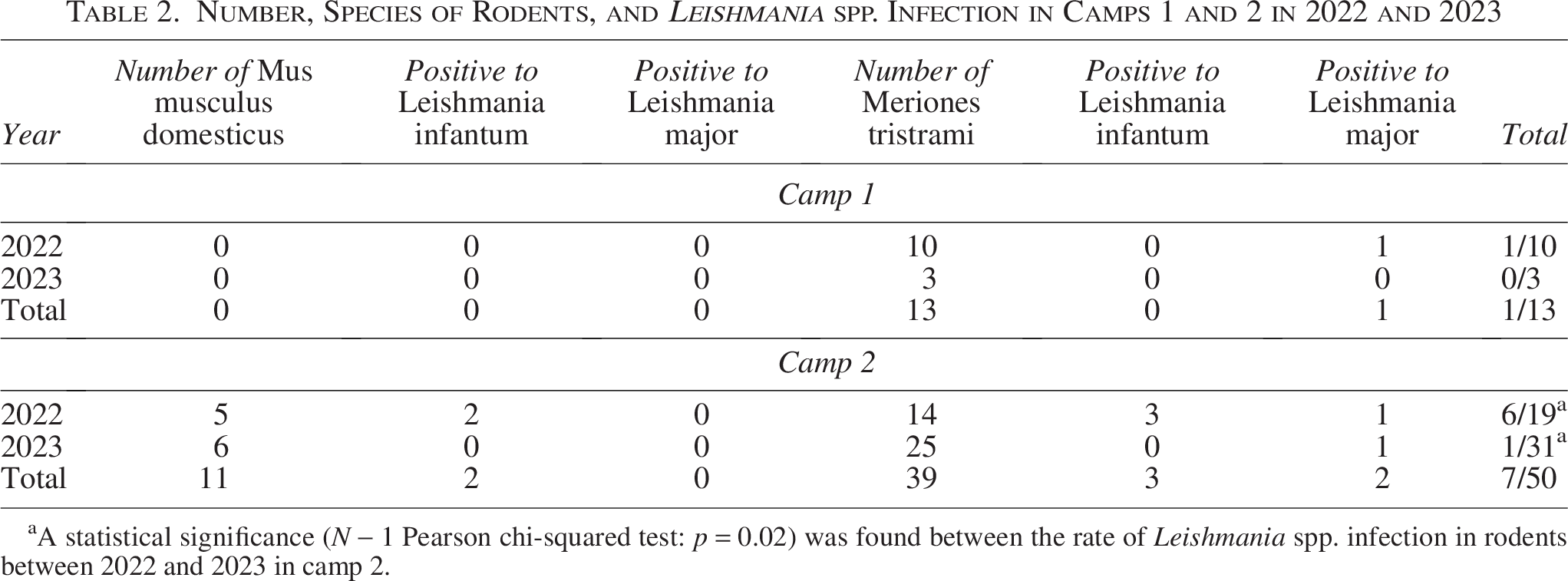
A statistical significance (N − 1 Pearson chi-squared test: p = 0.02) was found between the rate of Leishmania spp. infection in rodents between 2022 and 2023 in camp 2.
Leishmania DNA was detected in eight rodents (12.7%). Five carried L. infantum, including two M. musculus domesticus (18.2%) and three M. tristrami (5.8%), all sampled in 2022. L. major was found in three M. tristrami (5.8%): two in 2022 and one in 2023 (Table 2).
In total, 263 tissue samples were analyzed: 47 blood samples (whole blood or filter paper), 104 ear skin (51 animals had both ears, 2 had one), and 112 conjunctival swabs (55 had both eyes, two had one) (Table 2 and Supplementary Tables S1 and S2).
Leishmania infection in both camps analyzed together was higher in 2022 (7/29, 24.1%) compared with 2023 (1/34, 2.9%) (N − 1 Pearson χ2 = 4.6, p = 0.032). In camp 2, infection fell from 31.6% (2022) to 3.3% (2023) (χ2 = 5.4, p = 0.02), while no difference was observed in camp 1 (Fisher exact test, p = 1) (Table 2).
Rodent distribution and infection sites are shown in Figures 4 and 5. In 2022, most rodents were trapped outside camps in nearby agricultural plots. Only one M. tristrami was caught inside camp 2, infected with L. major. Another L. major–positive M. tristrami was trapped 240 m west of camp 1, near a water reservoir. L. infantum was detected in three M. tristrami and two M. musculus domesticus, all in 2022: one M. tristrami east of camp 2 (850 m), and two M. tristrami plus two M. musculus domesticus west of camp 2 (1,200 m). In 2023, a single L. major–positive M. tristrami was caught 1,200 m east of camp 2.
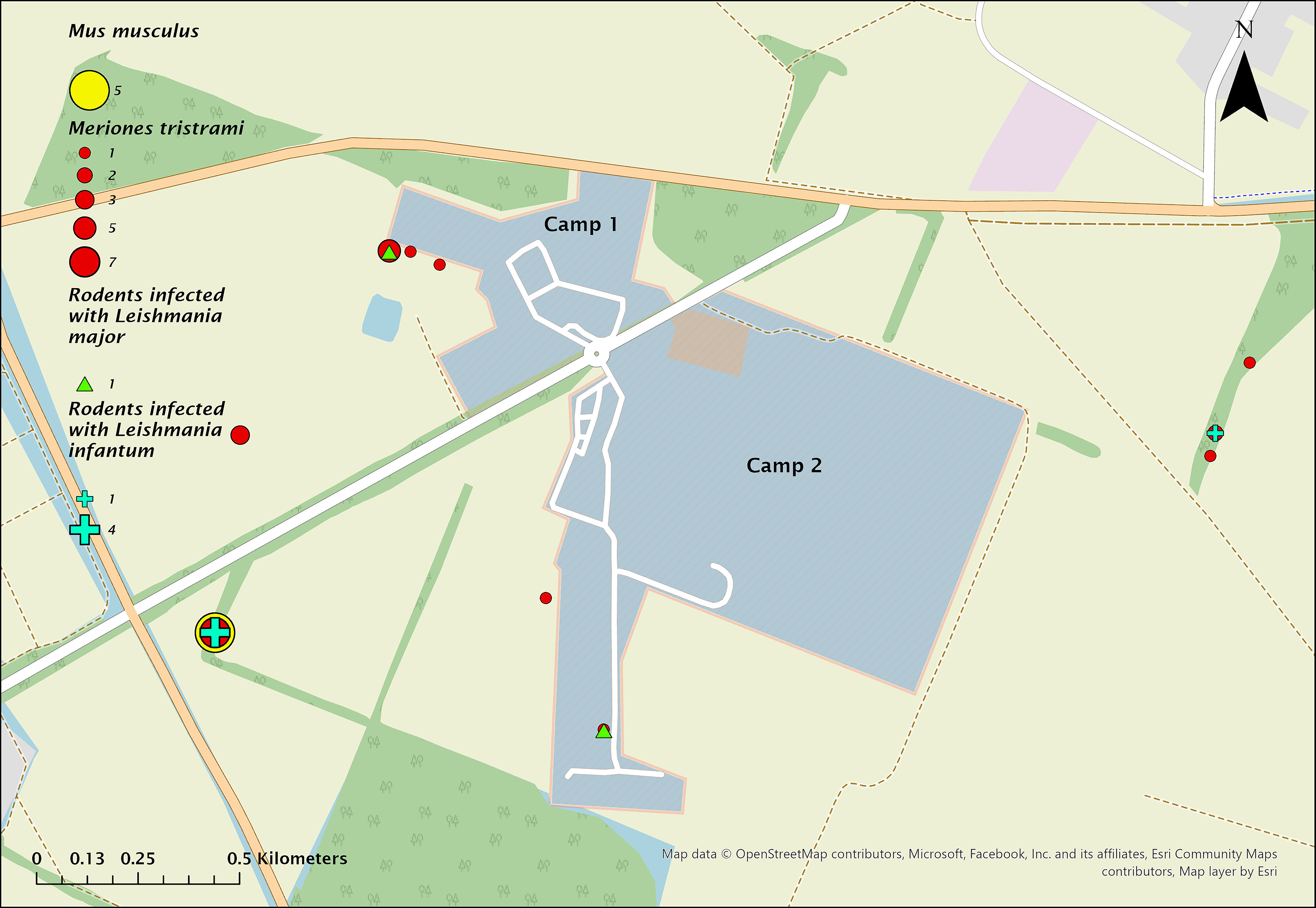
Map of camps 1 and 2 showing the dispersion of trapped Meriones tristrami (red circles) and Mus musculus domesticus (yellow circle) and the location of Leishmania major infection (green triangle) and Leishmania infantum (turquoise plus sign) during 2022.
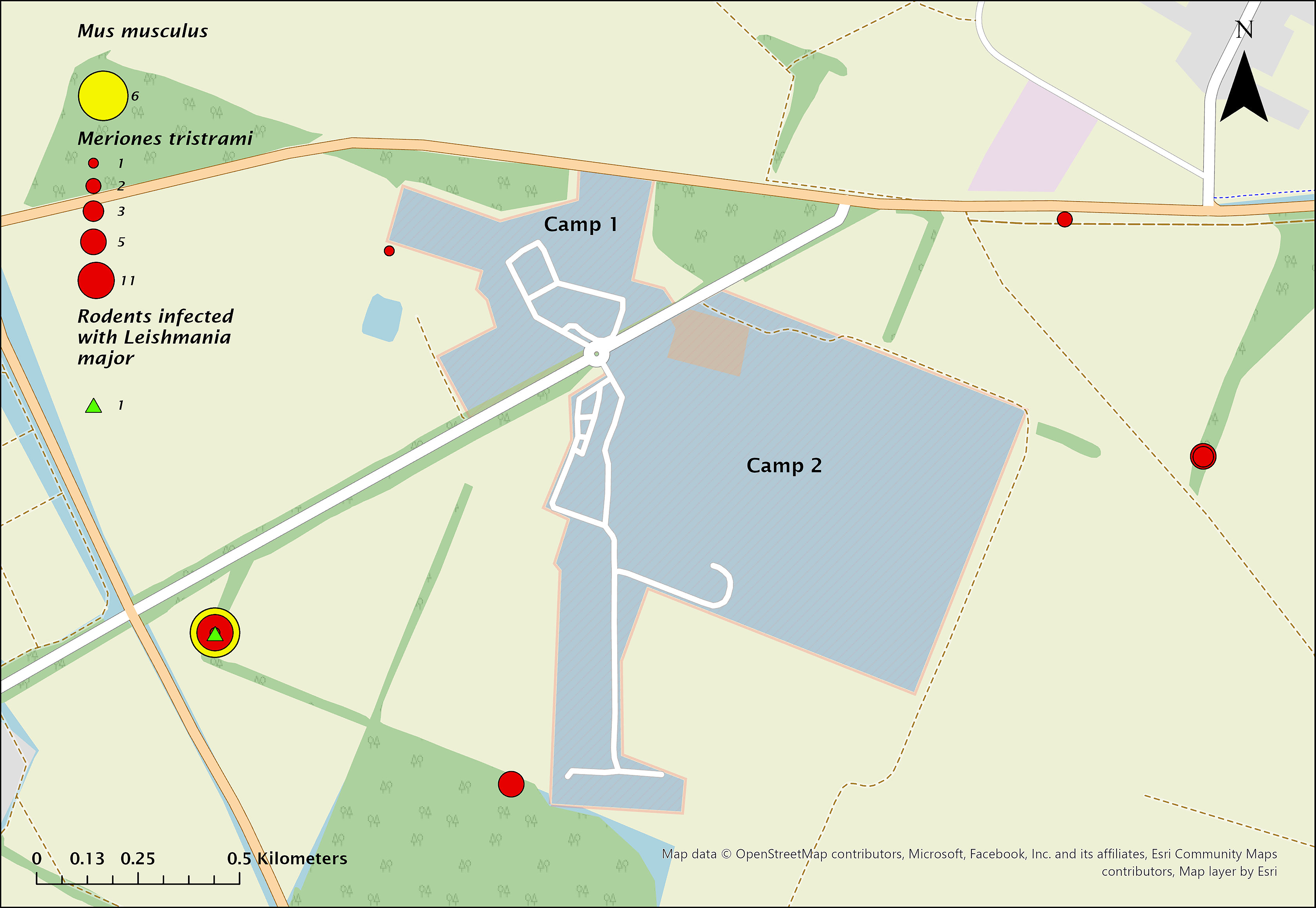
Map of camps 1 and 2 showing the dispersion of trapped Meriones tristrami (red circles) and Mus musculus domesticus (yellow circle) and the location of Leishmania major infection (green triangle) during 2023.
Cytology and histology
Three M. tristrami with ear lesions (hair loss, mild bruising) were examined by cytology and histopathology (Fig. 6). Amastigotes were detected in one animal, in both ears, by both methods (Figs. 7 and 8).
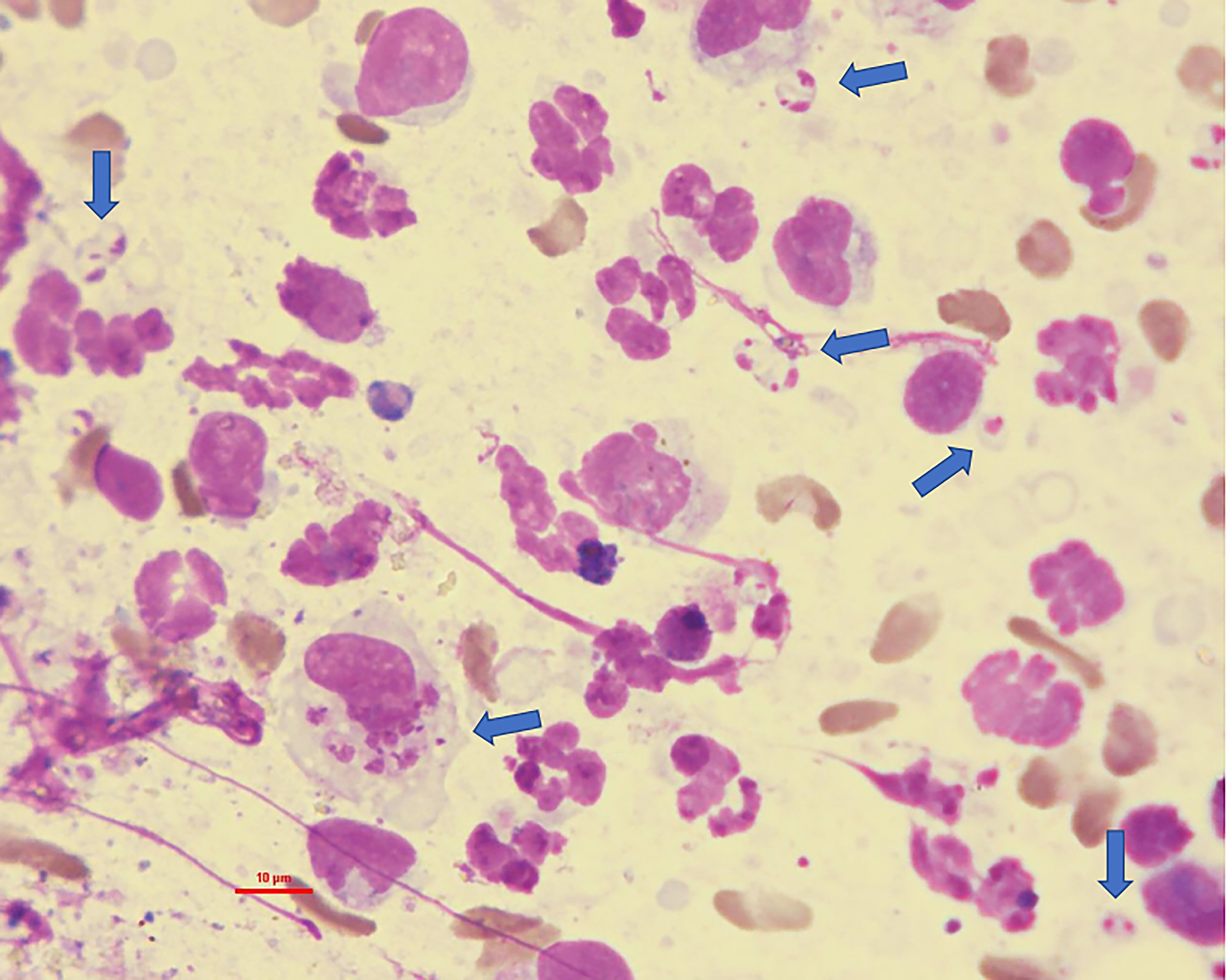
Cytology of a Leishmania major PCR-positive Meriones tristrami touch impression of the left ear skin showing amastigotes within and outside macrophages, marked with blue arrows.
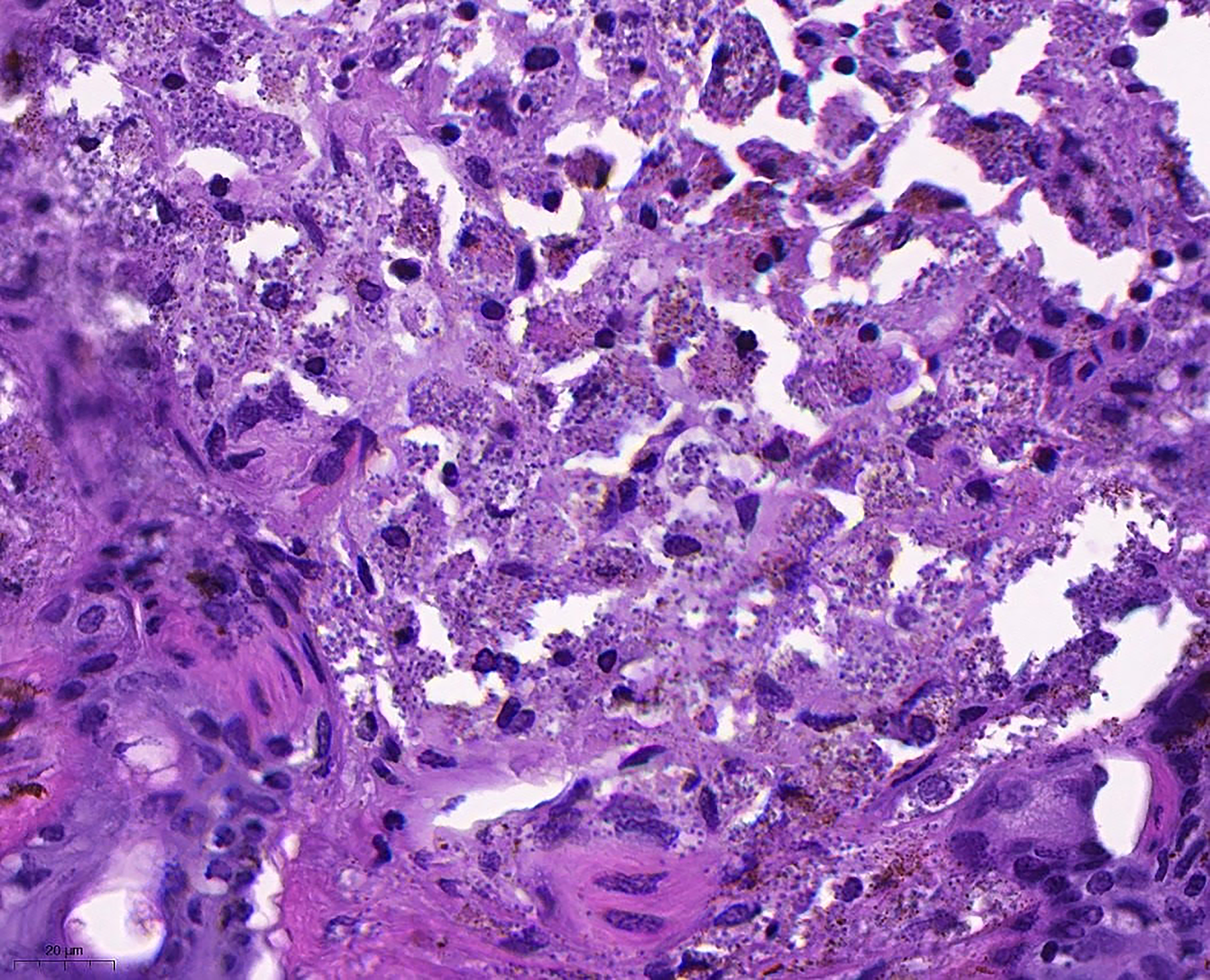
Histology of a Leishmania major PCR-positive Meriones tristrami ear showing macrophages laden with granular material consistent with amastigotes, visible as numerous small basophilic staining cytoplasmic inclusions.
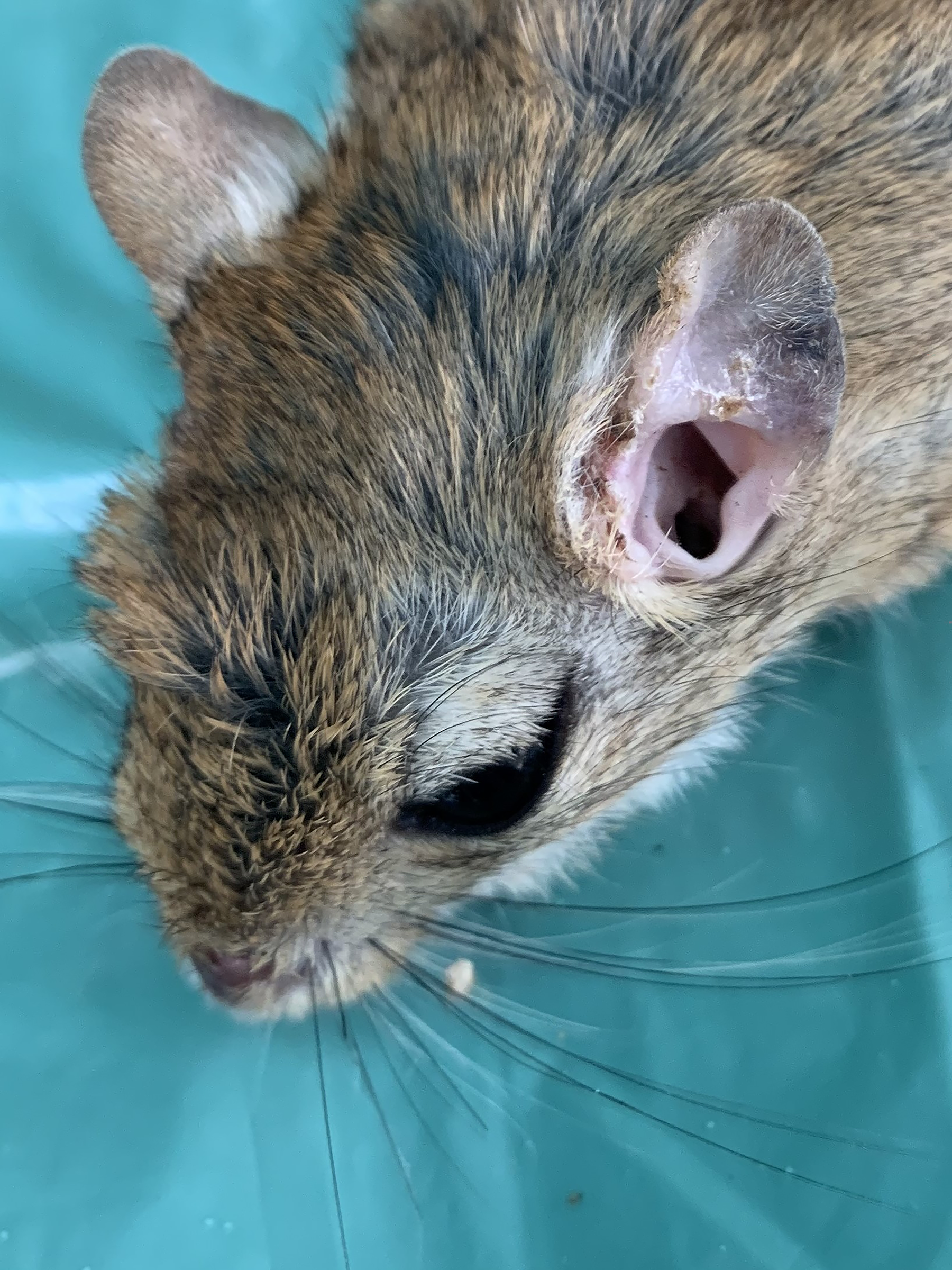
Meriones tristrami with lesions consisting of hair loss and mild skin scars in the left ear.
Discussion
This study provides an integrated, field-based assessment combining entomological surveillance in sand fly vectors, screening of rodent reservoirs, and analysis of human case data collected during 2022–2023 in two military bases located in a hyperendemic CL region of southern Israel. Human CL incidence correlated significantly with sand fly infection (MIR: p = 0.045, EREI: p = 0.023), with a 3-month lag consistent with the incubation period of L. major (Aoun et al., 2020). This study provides the first published observation of L. infantum infection in rodents in Israel and, to our knowledge, the first report in M. tristrami worldwide, consistent with previous reports of rodent infection in southern Europe and the Middle East (Papadogiannakis et al., 2010; Helhazar et al., 2013; Navea-Pérez et al., 2015; Pourmohammadi et al., 2017; Karakuş et al., 2020).
Importantly, Ph. papatasi is a recognized vector of L. major but is not considered a competent vector of L. infantum; consistent with this, L. infantum was not detected in Ph. papatasi in our study. Therefore, the detection of L. infantum in rodents is interpreted as incidental exposure rather than evidence of an active local transmission cycle involving Ph. papatasi.
We assessed an environmental intervention combining drone pesticide application with bait station traps targeting M. musculus domesticus and M. tristrami, known L. major reservoirs and agricultural pests (Faiman et al., 2013) in camp 2. Drone delivery, compared with crewed aircraft-dispersal, has been reported to provide better precision, improved environmental sustainability, lower costs, enhanced worker safety, and better access to remote areas (Morley et al., 2017; Regev et al., 2019). Drone delivery has been applied for rodent, mosquito, and zoonotic disease control (Yu et al., 2017; Regev et al., 2019; Carrasco-Escobar et al., 2022; Keshet et al., 2022; Morley et al., 2024).
Reports of CL in soldiers were analyzed from 2016 to 2024, and a clear outbreak was detected in 2021 and 2022 (Fig. 1). Human CL incidence declined significantly from 2022 to 2023: 33.3 to 4.9/1000 in camp 2 and 17.1 to 2.6/1000 in camp 1 (p < 0.001). Previous outbreaks of L. major in this region occurred in civilians in 2010–2012 and in military personnel in 2011–2015 (Gandacu et al., 2014; Orshan et al., 2016; Ben-Shimol et al., 2016; Kleinerman et al., 2023).
Ph. papatasi was the sole vector species found in the current study, confirming earlier surveys (Orshan et al., 2016; Schlein et al., 1984; Wasserberg et al., 2003). Sand fly abundance peaked in August in most cases, with year-to-year variation, as observed previously (Berger et al., 2014). Leishmania infection prevalence increased toward the late season, peaking in September and October 2022. Following rodent intervention in April 2023, only four positive pools were detected (three in camp 1 and one in camp 2). Although more sand flies were collected in camp 2 in 2023 compared with 2022 (2,624 vs. 1,560), the MIR declined significantly (1.23 vs. 0.03, p = 0.034). In camp 1, sand fly numbers fell (6,313 vs. 1,586), but the MIR did not differ (0.50 vs. 0.22, p = 0.59). These trends suggest a reduced host-to-vector transmission, though sand fly abundance likely persisted through alternative hosts or breeding sites (Tsurim et al., 2023). Most sand flies were trapped outside camp fences, consistent with previous findings (Boaz, 1991; Orshan et al., 2016), indicating probably higher soldier exposure near boundaries.
Among 63 rodents, eight were infected: L. infantum was found in 2/11 M. musculus domesticus (18.2%) and 3/52 M. tristrami (6%), and L. major in 3/52 M. tristrami (6%). L. major prevalence was lower than found in earlier Israeli reports (58–67%) (Faiman et al., 2013; Orshan et al., 2016), possibly reflecting a natural decline in the studied area. L. infantum was previously found in 2/75 Meriones persicus in Iran, a closely related species to M. tristrami (Fallah et al., 2006) and in 24.2–88.9% of house mice tested in southern Europe (Alcover et al., 2021). Our small M. musculus domesticus sample limits firm conclusions on their epidemiological role. Despite higher rodent capture in 2023, infection prevalence declined significantly overall (p = 0.032) and particularly in camp 2 (p = 0.02) compared with 2022.
In a previous study, cats living in these camps showed high prevalence of L. major and L. infantum and cutaneous clinical signs, and represented the first feline L. major infections reported in Israel (Kleinerman et al., 2023). Since Ph. papatasi feeds on multiple hosts, felines may also contribute to transmission cycles involving rodents and humans.
Surveillance in 2023 revealed reduced infection in sand flies, rodents, and humans, particularly in camp 2, suggesting an intervention effect. Yet CL is cyclic, with natural peaks roughly every decade (Mimouni et al., 2009). The last major outbreak locally was in 2011–2015 (Kleinerman et al., 2023). Thus, the 2023 decline may partly reflect natural dynamics. Nonetheless, the concurrent reduction in both vectors and reservoirs in camp 2 strongly supports intervention impact. Comparable host-targeted control has reduced CL elsewhere, such as zinc phosphide baits in Iran (Ershadi et al., 2005) and strychnine wheat baits in Morocco (Bennis et al., 2015). Our approach differs from these studies by combining bait stations with drone pesticide delivery in order to maximize efficacy and minimize environmental damage.
With regard to ethical considerations, drone-assisted rodent control is not part of a routine preventive practice. Its selective implementation represented an exploratory outbreak response rather than the withholding of established measures. Regarding preventive measures, standard personal protective practices were routinely recommended in both camps.
The study limitations included using only camp 1 as the reference site due to its proximity and similar ecology to camp 2. Although an external control would have strengthened the comparison, camp 1 provided a reasonable baseline for assessing the intervention impact. Regarding the trapped sand flies, as these were tested in pools, MIR provided a conservative measure suitable for comparing trends across camps and years but not for estimating absolute prevalence. While a larger rodent sample size could have enhanced the statistical power of the outcomes, the detection of L. major and L. infantum in two species offers novel insights into reservoir dynamics in the region. With regard to the environmental intervention, as both drone-based pesticide application and bait stations were deployed simultaneously, their individual effects could not be separated. Future studies using phased or factorial designs to assess the specific contribution of each approach are needed. Seasonal and behavioral variability likely also influenced sand fly and rodent populations; however, the post-iinterventiondeclines point to a tangible impact of targeted control measures.
In conclusion, Leishmania circulated in sand flies and rodents during a 2022–2023 outbreak in two southern Israeli bases. L. infantum and L. major infections were detected in rodents, complementing previous findings in local felines. Infection rates in vectors and reservoirs fell sharply in the camp subjected to drone-based pest control. However, this study should be interpreted as preliminary, field-based evidence, and a longitudinal, multisite study, which incorporates environmental covariates, is needed to unravel natural cycles from intervention outcomes. Nonetheless, these findings highlight the potential of drone technologies for integrated control of zoonotic vector-borne diseases.
Authors’ Contributions
G.K.: Conceptualization, data curation, investigation, and writing (lead). L.C. and L.O.: Conceptualization and supervision (equal). S.E.: Methodology and formal analysis (equal). S.G.: Data curation and supervision (equal). Y.M.: Data curation and investigation (equal). Y.N.-B.: Formal analysis (equal). O.D.K.: Writing—review and editing (supporting). S.M., S.G., N.M., Y.S., and M.T.: Investigation (equal). D.G. and Y.L.M.: Supervision (supporting). R.G.K.: Investigation (supporting). S.E.M.: Investigation (supporting). N.D.: Supervision and writing—review and editing (supporting). G.B.: Supervision and writing—review (lead). All authors read and approved the final article.
Footnotes
Acknowledgment
The authors thank Dr. Ori Brenner, from the Weizmann Institute of Sciences for his assistance with histopathology of the rodent ear tissues.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.