
Abstract
Background:
Tick-borne diseases caused by spotted fever group (SFG) Rickettsia and members of the family Anaplasmataceae are important public health concerns. Given that previous studies on SFG in Hokkaido, the northernmost prefecture of Japan, have been limited by small sample sizes or limited to specific pathogens or tick species, the epidemiology of tick-borne bacteria remains unclear. This study investigated the prevalence of SFG Rickettsia and Anaplasmataceae bacteria in multiple tick species in Hokkaido.
Methods:
DNA was extracted from 1,115 questing ticks representing six species (548 Ixodes ovatus, 368 Ixodes persulcatus, 95 Haemaphysalis concinna, 74 Haemaphysalis japonica, 28 Haemaphysalis megaspinosa, and 2 Haemaphysalis longicornis) collected from six districts in Hokkaido. Samples were screened using PCR assays targeting the gltA gene of SFG Rickettsia species and the groEL gene of Anaplasmataceae. Amplicons were sequenced for species identification.
Results:
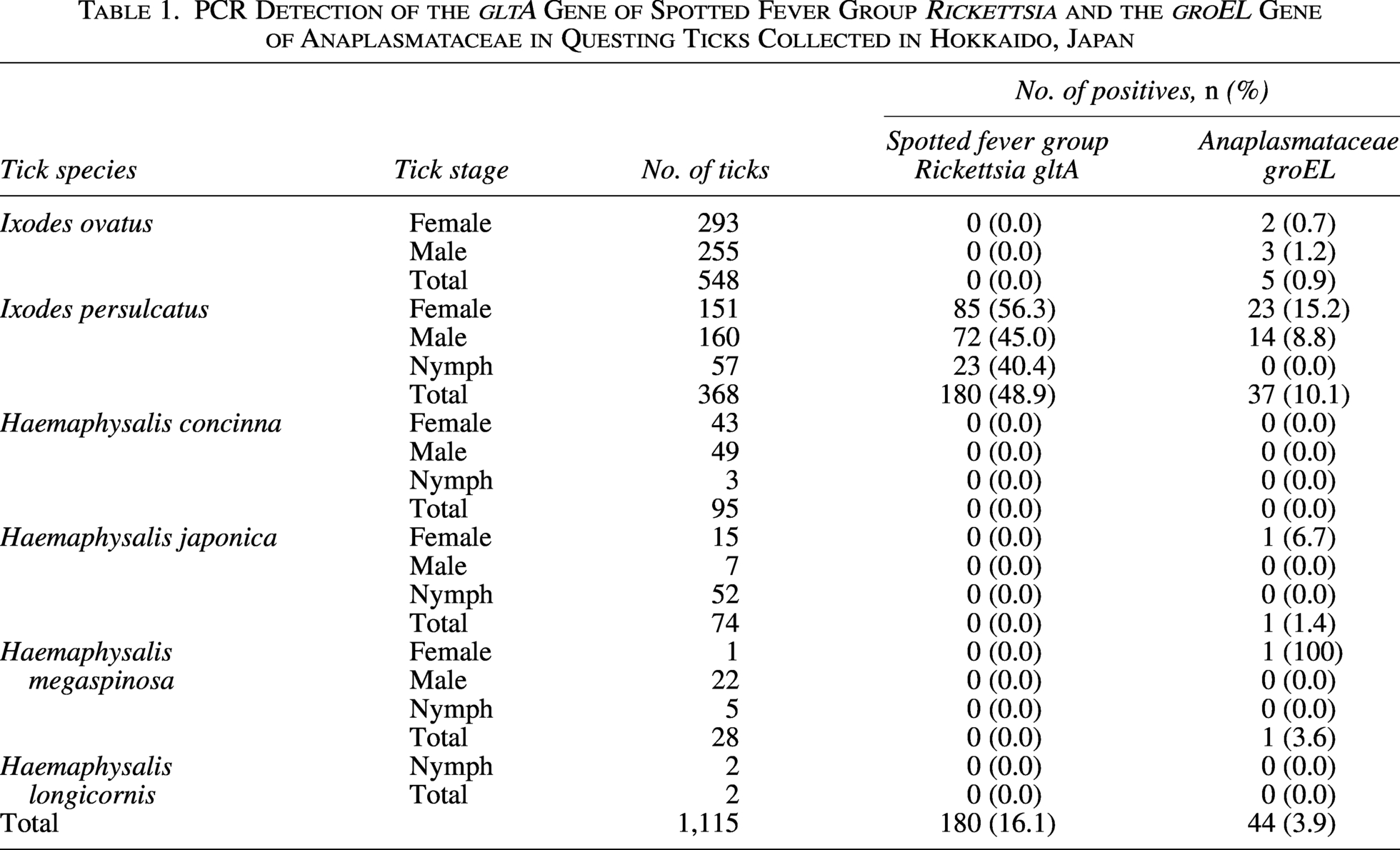
SFG Rickettsia DNA was found only in I. persulcatus (48.9%, 180/368), including R. helvetica (32.1%) and Candidatus R. tarasevichiae (16.8%). Anaplasmataceae DNA was detected in 3.9% (44/1,115) of ticks, including I. persulcatus, I. ovatus, H. japonica, and H. megaspinosa. In I. persulcatus, Ehrlichia muris, Candidatus Neoehrlichia mikurensis, and Anaplasma phagocytophilum were detected in 4.6%, 3.3%, and 0.3%, respectively. Ehrlichia japonica was found in I. ovatus (0.9%), while unclassified Ehrlichia spp. were detected in H. japonica (1.4%) and H. megaspinosa (3.6%). Seventeen I. persulcatus (4.6%) carried both SFG Rickettsia and Anaplasmataceae.
Conclusions:
The findings showed that questing ticks in Hokkaido harbor zoonotic bacteria. While I. persulcatus appears to be the principal vector, the detection of pathogens in I. ovatus, H. japonica, and H. megaspinosa suggests the potential for additional vectors in the region. These findings underscore the importance of continuous surveillance and heightened clinical awareness of tick-borne diseases in northern Japan.
Introduction
Tick-borne diseases, including rickettsioses, ehrlichiosis, anaplasmosis, and neoehrlichiosis, which are caused by obligate intracellular bacteria within the families Rickettsiaceae and Anaplasmataceae, pose a considerable threat to public health (Rochlin and Toledo, 2020). In particular, spotted fever group (SFG) rickettsioses and diseases caused by Anaplasmataceae species are of significant concern, given their clinical presentations, which range from fever and rash to serious complications such as organ failure and coagulopathies (Biggs et al., 2016). Clinically important examples include Japanese spotted fever (JSF) caused by Rickettsia japonica, human granulocytic anaplasmosis caused by Anaplasma phagocytophilum, human monocytic ehrlichiosis caused by Ehrlichia chaffeensis, and neoehrlichiosis caused by Candidatus Neoehrlichia mikurensis (Biggs et al., 2016; Dumler et al., 2007; Parola and Raoult, 2001; Wennerås, 2015).
R. japonica, A. phagocytophilum, E. chaffeensis, and Ca. N. mikurensis are primarily transmitted by hard ticks of the genera Ixodes, Haemaphysalis, and Amblyomma (Biggs et al., 2016; Parola and Raoult, 2001; Wennerås, 2015). These pathogens have been increasingly reported across Asia, Europe, and North America, underscoring the need for continuous vector surveillance to monitor their prevalence, detect emerging species, and reduce public health risks (de la Fuente et al., 2023; Parola and Raoult, 2001; Rochlin and Toledo, 2020).
In Japan, JSF caused by R. japonica is the most common clinical tick-borne rickettsiosis, especially in the western and coastal regions of the country (Mahara, 1997; Mahara et al., 1985). Other SFG Rickettsia species, including R. helvetica and Ca. R. tarasevichiae, have also been detected in ticks (Okado et al., 2021). In addition, several Anaplasmataceae bacteria, including Ehrlichia muris, E. japonica, Ca. N. mikurensis, and A. phagocytophilum, have been detected in both ticks and wild animals (Kawahara et al., 1993, 2004; Ohashi et al., 2005; Taira et al., 2019; Wu et al., 2015; Yoshimoto et al., 2010). Importantly, recent studies in Japan have provided serological and molecular evidence of human infections with these Anaplasmataceae bacteria (Ohashi et al., 2013; Su et al., 2022).
To date, no confirmed human cases of anaplasmatiosis or SFG rickettsioses have been reported from Hokkaido, despite the presence of candidate tick vectors in the region. Hokkaido, the northernmost island of Japan, has a distinct tick fauna shaped by its cold climate. Although previous studies provided valuable insights into Rickettsiaceae and Anaplasmataceae in Hokkaido, investigations conducted more than a decade ago detected R. helvetica and A. phagocytophilum in ticks from the region, but these studies were based on relatively small sample sizes (Ybañez et al., 2012; Ybañez and Inokuma, 2016; Yoshimoto et al., 2010). A more recent survey of Rickettsia species in ticks from eastern Hokkaido identified Ixodes persulcatus as a potential primary vector (Okado et al., 2021). However, Anaplasmataceae bacteria were not included in that study. Therefore, the present study investigated the prevalence of SFG Rickettsia and Anaplasmataceae species in questing ticks collected from Hokkaido.
Materials and Methods
Questing ticks and their DNAs
The present study used DNA samples extracted from a total of 1,115 questing ticks collected between 2008 and 2022 in Hokkaido, Japan (Ma et al., 2026). Briefly, ticks were collected from six districts: Ashoro (n = 156), Shikaoi (n = 128), Otofuke (n = 42), Memuro (n = 501), Taiki (n = 95), and Shibecha (n = 193) (Supplementary Table S1). Based on morphological keys (Yamaguti et al., 1971), the ticks were classified into six species: Ixodes ovatus (n = 548), I. persulcatus(n = 368), Haemaphysalis concinna (n = 95), Haemaphysalis japonica (n = 74), Haemaphysalis megaspinosa (n = 28), and Haemaphysalis longicornis (n = 2) (Table 1, Supplementary Table S1). I. persulcatus was collected from all six districts, I. ovatus from four districts (Ashoro, Shikaoi, Memuro, and Shibecha), and H. japonica from five districts except for Memuro. H. megaspinosa was collected from two districts (Shikaoi and Shibecha), while H. concinna and H. longicornis were each collected only from Shibecha (Supplementary Table S1). DNA was extracted from each tick using an ISOHAIR Kit (Nippon Gene, Toyama, Japan) following the manufacturer’s instructions and stored at −20°C until use.
PCR Detection of the gltA Gene of Spotted Fever Group Rickettsia and the groEL Gene of Anaplasmataceae in Questing Ticks Collected in Hokkaido, Japan
PCR amplification of the gltA gene in SFG Rickettsia and groEL gene in Anaplasmataceae
Tick DNA samples were subjected to two PCR assays targeting conserved regions of the gltA gene of SFG Rickettsia and the groEL gene of Anaplasmataceae. For gltA, the primers Cs2d (5′-ATGACCAATGAAAATAATAAT-3′) and CsEndr (5′-CTTATACTCTCTATGTACA-3′) were used (Mediannikov et al., 2004), while for groEL, the primers gro607F (5′-GAAGATGCWGTWGGWTGTACKGC-3′) and gro1294R (5′-AGMGCTTCWCCTTCWACRTCYTC-3′) were used (Tabara et al., 2007). For each PCR amplification, a 25 μL reaction mixture was prepared, incorporating 12.5 μL of 2× TaKaRa Ex Premier™ DNA Polymerase (Takara, Otsu, Japan), 0.2 μM of each primer, 2.0 μL of tick DNA, and 10.25 μL of distilled water, in accordance with the manufacturer’s instructions. Reactions were run on a VeritiPro thermal cycler (Applied Biosystems, Waltham, MA) under the following conditions: initial denaturation at 94°C for 1 min, followed by 40 cycles of 98°C for 10 s, 52°C (gltA) or 57°C (groEL) for 30 s, and 68°C for 1 min.
Following amplification, the PCR products were subjected to electrophoresis on a 2% agarose gel for 30 min, stained with Midori Green (Nippon Genetics, Tokyo, Japan) for 20 min, and observed under ultraviolet transillumination. Amplicons of approximately 1,290-bp (gltA) and 670-bp (groEL) were considered positive.
Sequencing and phylogenetic analyses
The nucleotide sequences of gltA and groEL amplicons were determined by Sanger sequencing at a commercial laboratory (Eurofins Genomics K.K., Tokyo, Japan). The resulting sequences were deposited in the DNA Data Bank of Japan (DDBJ).
The obtained gltA and groEL sequences, together with reference sequences from GenBank, were aligned using MEGA11 software (Tamura et al., 2021). For each dataset, the best-fitting substitution model was selected in MEGA11, and neighbor-joining phylogenetic trees were constructed using the Kimura two-parameter substitution model. Node support was assessed with 1,000 bootstrap replications.
Statistical analysis
Statistical analyses were performed to evaluate regional differences in pathogen prevalence. Chi-square or Fisher’s exact tests were applied as appropriate in R version 4.3.0 (R Core Team, Vienna, Austria). A p value of <0.05 was considered statistically significant.
Results
PCR detection of the gltA gene of SFG Rickettsia, sequencing, and phylogenetic analysis
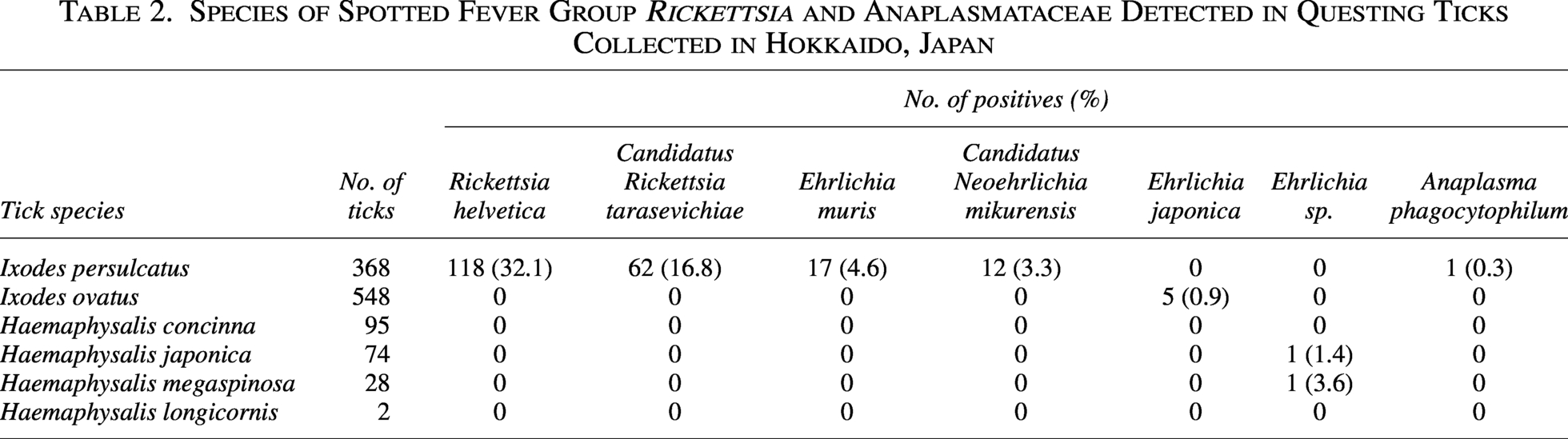
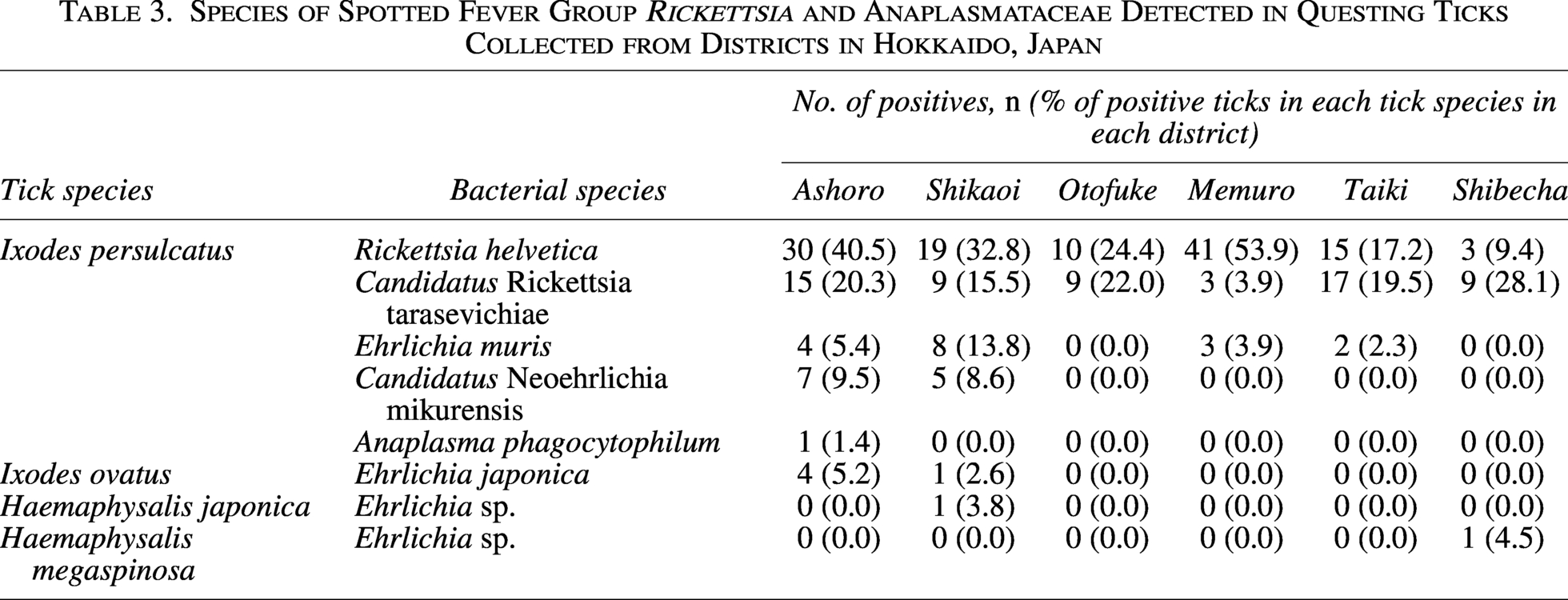
The SFG Rickettsia gltA gene was detected in 180 I. persulcatus ticks collected from all six surveyed districts, whereas all other tick species tested negative (Table 1, Supplementary Table S1). All 180 amplicons were successfully sequenced. Basic local alignment search tool (BLAST) analysis revealed that the obtained sequences shared high identity with those of two SFG Rickettsia species, R. helvetica and Ca. R. tarasevichiae (Table 2). A total of 118 sequences (32.1% of I. persulcatus samples) were identical to R. helvetica gltA gene sequences previously detected in I. persulcatus from Russia (accession number: KU310588) and Ixodes ricinus from the Netherlands (OY974117). The prevalence of R. helvetica varied among the districts, ranging from 9.4% to 53.9% (Table 3), and this difference was statistically significant (χ2 test, p < 0.05). The remaining 62 sequences (16.8% of I. persulcatus) were identical to gltA gene sequences of Ca. R. tarasevichiae previously reported in I. persulcatus from Russia (DQ168981). The prevalence of Ca. R. tarasevichiae ranged from 3.9% to 28.1% (Table 3), also showing significant regional variation (χ2 test, p < 0.05).
Species of Spotted Fever Group Rickettsia and Anaplasmataceae Detected in Questing Ticks Collected in Hokkaido, Japan
Species of Spotted Fever Group Rickettsia and Anaplasmataceae Detected in Questing Ticks Collected from Districts in Hokkaido, Japan
Representative sequences of R. helvetica (n = 6, LC872519–LC872523, LC872530) and Ca. R. tarasevichiae (n = 6, LC872524–LC872529) from each district were also deposited in the DDBJ. Phylogenetic analysis placed all sequences into two well-supported clades corresponding to R. helvetica and Ca. R. tarasevichiae (Fig. 1).

Phylogenetic tree of the gltA gene of spotted fever group Rickettsia. The gltA gene sequences generated in the present study, together with reference sequences from GenBank, were used to construct a neighbor-joining phylogenetic tree. The newly obtained sequences (indicated by black dots) clustered within clades corresponding to Rickettsia helvetica and Candidatus Rickettsia tarasevichiae.
PCR detection of the Anaplasmataceae groEL gene, sequencing, and phylogenetic analysis
The groEL gene of Anaplasmataceae was detected in a total of 44 ticks, including I. persulcatus, I. ovatus, H. japonica, and H. megaspinosa, but not in H. concinna and H. longicornis (Table 1). The highest detection rate was observed in I. persulcatus (10.1%, 37/368). Positive ticks were found in five of the six sampled districts, except Otofuke (Supplementary Table S1). Of the 44 PCR amplicons, 39 were successfully sequenced, while five failed due to insufficient DNA quality. BLAST analysis of the 39 sequences revealed that the obtained groEL sequences were derived from five species: E. muris, E. japonica, an unclassified Ehrlichia sp., Ca. N. mikurensis, and A. phagocytophilum. Of the 37 sequences determined, 11 representative sequences (LC872531–LC872541) were submitted to the DDBJ. The groEL sequences of E. muris (LC872531–LC872534), E. japonica (LC872537 and LC872538), and Ca. N. mikurensis (LC872535 and LC872536) were identical to sequences previously reported from a wild mouse in Japan (AF210459), I. ovatus in Fukushima, Japan (CP007474), and I. persulcatus in Russia (MG182157, FJ966359), respectively. The A. phagocytophilum sequence (LC872539) shared 99.6% identity with a sequence from I. persulcatus tick in Russia (MN989865). By contrast, the unclassified Ehrlichia sp. sequences from H. japonica and H. megaspinosa shared 98.7% and 98.6% identity with a sequence from an unclassified Ehrlichia sp. detected in a tick in China (PQ249203).
Ehrlichia muris, Ca. N. mikurensis, and A. phagocytophilum were detected in 17 (4.6%), 12 (3.3%), and 1 (0.3%) I. persulcatus ticks, respectively. While E. japonica was detected in five (0.9%) I. ovatus ticks, and the unclassified Ehrlichia sp. was detected in one (1.4%) H. japonica and one (3.6%) H. megaspinosa (Table 2). The prevalence of E. muris and Ca. N. mikurensis ranged from 2.3% to 13.8% and 8.6% to 9.5%, respectively, among the sampled districts (Table 3), and these regional differences were statistically significant (χ2 test, p < 0.05).
In the phylogenetic analysis, the E. muris, E. japonica, Ca. N. mikurensis, and A. phagocytophilum sequences determined in the present study clustered within their corresponding clades. The newly generated Ehrlichia sp. sequences grouped within a clade containing several unclassified Ehrlichia spp. (Fig. 2).

Phylogenetic tree of the groEL gene of Anaplasmataceae. The groEL sequences generated in the present study, together with reference sequences from GenBank, were used to construct a neighbor-joining phylogenetic tree. The newly obtained sequences (indicated by black dots) clustered within clades corresponding to Ehrlichia muris, Ehrlichia japonica, several unclassified Ehrlichia spp., Candidatus Neoehrlichia mikurensis, and Anaplasma phagocytophilum.
Coinfection of SFG Rickettsia and Ehrlichia in I. persulcatus
A total of 17 I. persulcatus ticks from Ashoro, Shikaoi, and Memuro were coinfected with SFG Rickettsia and Anaplasmataceae (Supplementary Table S2). Three ticks from Ashoro and two from Memuro were coinfected with R. helvetica and E. muris. Five ticks from Ashoro and three from Shikaoi were coinfected with R. helvetica and Ca. N. mikurensis. Additionally, one tick from Ashoro and three from Shikaoi were coinfected with Ca. R. tarasevichiae and E. muris.
Discussion
The present study detected R. helvetica, Ca. R. tarasevichiae, E. muris, Ca. N. mikurensis, and A. phagocytophilum in I. persulcatus and E. japonica in I. ovatus. By contrast, all H. concinna and H. longicornis ticks tested negative, which may reflect the small sample sizes or low infection rates in these species. In addition to known pathogens, unclassified Ehrlichia spp. not previously reported in Japan were detected in H. megaspinosa and H. japonica. The groEL sequences from these Ehrlichia spp. shared high identity with strains reported in ticks and dogs from China (Zhang et al., 2023). Further studies are needed to determine the pathogenic potential of these Ehrlichia strains.
Several limitations of this study should be noted. Species identification was based on single-locus sequencing (gltA for SFG Rickettsia and groEL for Anaplasmataceae), which provides useful taxonomic information but may not fully resolve closely related species. In addition, detection of Anaplasmataceae DNA in ticks does not necessarily indicate pathogenicity to humans, as groEL sequences alone are insufficient to assess clinical relevance. Further multilocus, genomic, and epidemiological studies are needed to clarify the diversity and public health significance of these bacteria. Although several Rickettsia and Anaplasmataceae species have been detected in humans, definitive assessment of their virulence requires phenotypic characterization using isolated strains. Therefore, further studies involving bacterial isolation and experimental infection are necessary to clarify their pathogenic potential.
Significant regional differences were also observed in the prevalence of R. helvetica, Ca. R. tarasevichiae, E. muris, and Ca. N. mikurensis. This regional heterogeneity in prevalence may be influenced by ecological and environmental factors, including local variation in tick density, host animal distribution, and habitat characteristics. The observed differences could also reflect distinct host–vector–pathogen interactions in each district. Similar findings of diverse prevalence patterns across sites in close geographical proximity have been reported for Borrelia burgdorferi sensu lato in I. ricinus ticks across Europe (Rauter and Hartung, 2005) and in Switzerland, where forest-dwelling I. ricinus populations exhibited pronounced local differences in pathogen prevalence (Lommano et al., 2012). These findings suggest that even small-scale ecological and environmental differences can result in significant regional variation in pathogen prevalence, underscoring the need for multi-district surveillance in Hokkaido.
With respect to SFG Rickettsia, our findings highlight the predominance of R. helvetica (32.1%) and Ca. R. tarasevichiae (16.8%) in I. persulcatus in Hokkaido. This contrasts with western Japan, where R. japonica is the primary causative agent of JSF and the prevalence of R. helvetica in ticks has been reported to be considerably lower (Mahara, 1997; Okado et al., 2021; Sando et al., 2018). In Europe, including Switzerland, France, and the Netherlands, R. helvetica prevalence in I. ricinus has been reported to range from 5% to 15% (Pritt et al., 2017; Welinder-Olsson et al., 2010), which is lower than that observed in the present study. Ca. R. tarasevichiae prevalence of 10–25% has been reported in I. persulcatus from Russia and northeastern China (Mediannikov et al., 2004; Zhang et al., 2023), which is comparable to the 16.8% detected in our study. These findings suggest that the rickettsial fauna and prevalence in Hokkaido are more consistent with those of northern Eurasia than with other regions of Japan.
Regarding Anaplasmataceae, the prevalence of E. muris (4.6%) and Ca. N. mikurensis (3.3%) in Hokkaido appears higher than that reported from Honshu, where detection in ticks was sporadic and typically <2% (Taira et al., 2019; Yoshimoto et al., 2010). Internationally, E. muris has been detected in I. persulcatus in Russia at prevalence levels of 3–7% and in ticks from the United States at comparable rates (Pritt et al., 2017). Ca. N. mikurensis is recognized as an emerging human pathogen, with prevalences of 2–8% in I. ricinus ticks in Europe and China (Welinder-Olsson et al., 2010; Wennerås, 2015), which is consistent with the findings of the present study. In contrast, A. phagocytophilum was rare in Hokkaido (0.3%), similar to previous reports from Japan (<1%) (Ohashi et al., 2005; Su et al., 2022), and markedly lower than prevalences reported in Europe and North America, where detection in ticks often exceeds 5–10% (Schudel et al., 2024). These comparisons indicate that although the distribution of Anaplasmataceae in Hokkaido shares similarities with international trends, the overall prevalence reflects a distinct epidemiological profile that warrants further investigation.
It is also noteworthy that no Anaplasmataceae DNA was detected in nymphal ticks. This absence may be due to the limited number of nymphs examined in the present study, but it could also reflect stage-specific pathogen dynamics, such as lower acquisition rates during earlier blood meals or reduced transstadial transmission efficiency. In addition, the lack of detection in nymphs may be related to the absence of transovarial transmission in these bacteria, as larvae are not infected at hatching and must acquire pathogens from infected hosts during feeding (Kocan et al., 2015; Parola and Raoult, 2001; Stich et al., 1989). Consequently, the likelihood of detection in nymphs is expected to be lower than in adults, which have already undergone one or more blood meals. Further studies with larger sample sizes are needed to clarify whether the absence of Anaplasmataceae in nymphs represents a true biological phenomenon or a sampling artifact.
Human infections caused by some of the pathogens detected in the present study have been documented in Japan, including cases of R. helvetica (Takada et al., 2006) and A. phagocytophilum (Ohashi et al., 2013). However, no human cases caused by these pathogens have been reported from Hokkaido to date. Outside Japan, infections with R. helvetica, Ca. R. tarasevichiae, E. muris, Ca. N. mikurensis, and A. phagocytophilum have been reported in several countries, such as Switzerland, France, Russia, China, and the United States (Mediannikov et al., 2004; Pritt et al., 2017; Welinder-Olsson et al., 2010; Wennerås, 2015). Several factors might account for this discrepancy. Another factor contributing to the scarcity of reported human cases is that infections caused by these bacteria are often mild or nonspecific and may not prompt medical consultation. First, since these pathogens are susceptible to doxycycline (Maurin et al., 2003; Schudel et al., 2024), undiagnosed infections may have been empirically treated with doxycycline as tick-borne infections. Second, unlike R. japonica or Orientia tsutsugamushi, which are notifiable diseases in Japan (Ministry of Health, Labour and Welfare, Japan, 2024), the pathogens identified in the present study are not subject to mandatory reporting, likely contributing to underreporting. Third, the prevailing assumption that rickettsial diseases are confined to western Japan (Mahara, 1997; Sando et al., 2018) may have led to these pathogens being overlooked during clinical diagnosis. Together, these factors emphasize the need for heightened clinical awareness and expanded surveillance of tick-borne bacterial diseases in Hokkaido.
Our findings suggest that tick-borne SFG Rickettsia and Anaplasmataceae bacteria may pose a public health threat in Hokkaido. Although I. persulcatus appears to be the primary vector, our results indicate that other tick species reported to bite humans (Natsuaki, 2021) may play a role in pathogen transmission. The detection of multiple pathogens in several tick species increases the likelihood of human exposure. Moreover, the presence of coinfected ticks raises the possibility of simultaneous transmission of multiple bacterial agents, which could complicate clinical diagnosis and management (Sánchez-Vicente et al., 2023; Stinco et al., 2016).
In conclusion, our study demonstrates that ticks in Hokkaido harbor multiple species of SFG Rickettsia and Anaplasmataceae with zoonotic potential. These findings underscore the importance of continuous vector surveillance and greater awareness among medical doctors and public health authorities to effectively manage the risk of tick-borne diseases in Hokkaido, Japan.
Authors’ Contributions
M.T.: Conceptualization, methodology, investigation, formal analysis, data curation, and writing—original draft. T.S., Y.M., and R.U.-S.: Methodology, investigation, formal analysis, data curation, and writing—review and editing. N.Y.: Conceptualization, investigation, writing—review and editing, resources, supervision, project administration, and funding acquisition. K.M.: Conceptualization, investigation, writing—review and editing, resources, supervision, and project administration.
Footnotes
Acknowledgments
The authors thank Ms. Hiroko Yamamoto of Obihiro University of Agriculture and Veterinary Medicine for her excellent technical assistance.
Author Disclosure Statement
All authors declare no conflicts of interest related to this study.
Funding Information
This work was supported by a grant from the Japan Agency for Medical Research and Development (Grant number: JP23wm0225031).
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.