
Abstract
Tularemia, caused by Francisella tularensis, is a zoonotic disease with various sources, transmission routes, and geographically different clinical signs. Diagnosis is challenging due to nonspecific symptoms, highlighting the importance of laboratory testing for accurate detection, effective outbreak management, and targeted treatment. Recent improvements have enhanced the accuracy of sensitive and specific immunological and molecular methods. Techniques such as enzyme-linked immunosorbent assay, immunochromatography, microagglutination tests, indirect immunofluorescence assays, and PCR continue to be key detection tools. Molecular typing methods such as whole-genome sequencing, single-nucleotide polymorphism analysis, multiple-locus variable-number tandem repeat analysis, pulsed-field gel electrophoresis, and matrix-assisted laser desorption ionization-time of flight enable precise genetic characterization of F. tularensis strains. These methods improve understanding of phylogeny, strain diversity, and transmission routes, while supporting outbreak investigations. This review aims to provide a comprehensive overview of current laboratory methods for diagnosing and typing tularemia as well as their application in clinical and research settings.
Introduction
Francisella tularensis is a Gram-negative intracellular bacterium that causes the zoonotic infection tularemia (Maurin and Gyuranecz, 2016). F. tularensis comprises three main subspecies: subsp. tularensis (Type A), the most virulent and mainly found in North America; subsp. holarctica (Type B), with the widest geographic distribution; and subsp. mediasiatica, primarily reported from Central Asia. F. novicida, previously classified within F. tularensis, is now often considered a separate species, although its taxonomic status remains debated, and it is generally not regarded as an agent of tularemia. (Sjöstedt, 2007; Muller et al., 2025). F. tularensis persists in nature through various reservoirs, mainly small mammals such as rodents and lagomorphs. Humans acquire infection via arthropod bites, contact with infected animals, contaminated water or soil, inhalation of aerosols, or ingestion of contaminated materials, while human-to-human transmission is very rare (World Health Organization, 2007; Maurin and Gyuranecz, 2016; Zellner and Huntley, 2019; Yeni et al., 2021). Symptoms typically develop within 3–5 days postexposure, often resembling influenza with fever, rash, lymphadenopathy, or soft-tissue lesions. The nonspecific nature of these symptoms can complicate diagnosis (Pal et al., 2021). Depending on the strain and route of infection, tularemia presents in six clinical forms: ulceroglandular, glandular, oculoglandular, oropharyngeal, typhoidal, and pneumonic, each showing distinct manifestations that reflect the bacterium’s diverse pathogenic potential (Maurin and Gyuranecz, 2016). Complications of tularemia may include suppurative lymphadenitis, meningitis, otitis media, myocarditis, brain abscess, encephalitis, and infections of the heart, bone, or soft tissues (Maurin, 2020; Makdasi et al., 2022). Tularemia occurs sporadically or in outbreaks, influencing the disease’s epidemiological patterns across regions (Sjöstedt, 2007). Accurate diagnosis of tularemia remains challenging due to the nonspecific nature of clinical symptoms and the intracellular lifestyle of the pathogen. Diagnosis of F. tularensis relies on culture, serology, and molecular methods, with serological tests such as Microagglutination Test (MAT), Immunofluorescence Assay (IFA), and Enzyme-Linked Immunosorbent Assay (ELISA) being the most commonly. However, interpretation of serological results is complicated by factors such as delayed antibody responses, persistence of antibodies, and potential cross-reactivity, while cell-mediated immunity may persist even in the absence of detectable antibody levels. Culture-based methods, although specific, are limited by low sensitivity and biosafety constraints, whereas molecular techniques provide rapid and sensitive detection but require specialized infrastructure (Wawszczak et al., 2022). Early and reliable diagnosis is essential for timely initiation of appropriate antimicrobial therapy, reduction of disease severity, and prevention of complications. In addition, prompt identification of cases plays a critical role in public health surveillance, outbreak detection, and implementation of control strategies (Johansson et al., 2004). Monitoring tularemia is crucial for tracking prevalence, guiding prevention efforts, and identifying high-risk areas. High-exposure groups such as wildlife workers, hunters, and laboratory personnel benefit from early detection. At the same time, rapid diagnosis in endemic regions or outbreaks is vital to control transmission and ensure timely care (Porsch-Özcürümez et al., 2004; Splettstoesser et al., 2005). Despite the availability of established diagnostic guidelines, including those provided by international organizations, several challenges remain unresolved, such as variability in diagnostic performance, lack of standardization across assays, and limitations in distinguishing closely related subspecies or cross-reactive infections. Furthermore, recent advances in immunological and molecular techniques have not been comprehensively integrated into existing frameworks. Therefore, this review aims to critically evaluate current laboratory methods for the diagnosis and typing of tularemia, with a particular focus on their limitations, recent improvements, and potential applications in clinical and research settings.
Diagnostic Criteria for Tularemia
Tularemia diagnosis integrates clinical signs, patient history, and laboratory tests (World Health Organization, 2007). The World Health Organization (WHO) defines a presumptive tularemia case as a patient exhibiting suggestive clinical symptoms, along with positive laboratory evidence, such as elevated serum antibodies to F. tularensis without documented fourfold change, detection of the bacterium in clinical specimens by fluorescent assay, or nucleic acid testing including PCR. A confirmed case requires both clinical signs and laboratory confirmation, which can be achieved by either isolating F. tularensis or demonstrating a fourfold or greater rise in antibody titers between the acute and convalescent phases of the disease. These criteria complement microbiological, immunological, and PCR-based methods to ensure accurate diagnosis (World Health Organization, 2007; Morse and Henkel, 2018; Wawszczak et al., 2022).
Hematological Findings in Tularemia
In tularemia, the white blood cell count is usually normal, though relative lymphocytosis with mild increases in C-reactive protein (CRP) and liver enzymes may occur. CRP elevation is generally modest, and the erythrocyte sedimentation rate often rises during the first month of infection (Tärnvik and Berglund, 2003).
Specimen Sampling for Tularemia Detection
Optimal sample collection should occur before antibiotic treatment, with serum preferred for most forms of tularemia. F. tularensis can be cultured from blood, swabs, tissue aspirates, or respiratory samples, while cerebrospinal or pleural fluids are rarely used. Invasive biopsies of lymph nodes should be avoided during the acute phase due to the risk of infection. Proper sampling prevents contamination and ensures an adequate specimen volume, reducing the risk of false-negative results. All procedures involving bacterial cultivation and animal experiments must be performed under biosafety level 3 (BSL-3) conditions, whereas routine specimen handling can be conducted under biosafety level 2 (BSL-2) conditions (Sewell, 2003).
Direct Identification of F. tularensis
Gram staining can be used in suspected cases of F. tularensis, potentially involving smears from fresh, unpreserved specimens such as cultures and tissues. Initial samples containing a high number of organisms are preferred for preparation. F. tularensis is a small, Gram-negative bacterium with a pleomorphic or coccoid morphology, typically measuring approximately 0.2–0.5 by 0.7–1.0 micrometers (World Health Organization, 2007). Due to its small size and weak Gram stain, conventional Gram stains have limited value for identifying F. tularensis. Suspected cases may show short, weakly stained Gram-negative rods or coccoid forms. Tularemia can be detected using the direct fluorescent antibody (DFA) assay, which enables rapid, presumptive, and confirmatory identification. Over 85% of confirmatory identifications in reference laboratories rely on DFA, using polyclonal or monoclonal fluorophore-labeled antibodies for fluorescence microscopy of primary samples (Splettstoesser et al., 2005; World Health Organization, 2007; Morse and Henkel, 2018).
Culture-Based Approaches for Tularemia Diagnosis
F. tularensis requires cysteine- and sulfhydryl-enriched media for growth. Commonly used media include cysteine-heart agar with 9% chocolate sheep blood and antibiotics, glucose-cysteine agar with thiamine and hemoglobin, chocolate agar with cystine, GC Agar II with isovitalex and hemoglobin, BCYE, Müller-Hinton agar with isovitalex, TGBA, Thayer-Martin, and heart-cysteine-blood agar. Colonies typically appear gray-white, smooth, and small after 48–72 h at 37°C with 5% CO2, though full growth on chocolate agar may take up to seven days (Hepburn and Simpson, 2008; Borgschulte et al., 2022; Ponderand et al., 2023). F. tularensis is highly infectious at low doses, especially via aerosols, underscoring its potential as a biological threat and the need for strict BSL-3 laboratory precautions. Despite optimal conditions, culture positivity from clinical specimens remains below 10% due to the bacterium’s fastidious growth, early antibiotic treatment, limited sample availability, and the need for specialized laboratory facilities (Johansson et al., 2004; Hepburn and Simpson, 2008; Yanes et al., 2018). In addition, F. tularensis bacteremia is often transient, making it challenging to isolate in blood cultures. While supplementing blood culture media with iron has shown promise in reducing growth periods and accelerating bacterial proliferation, the slow growth rate and stringent nutritional needs of the organism remain significant hurdles (Makdasi et al., 2022). Culture is generally not preferred for tularemia diagnosis but remains important in research to study F. tularensis growth and antimicrobial susceptibility. Biochemically, the bacterium is weakly catalase-positive, oxidase-negative, and H2S-positive, producing acid from glucose and maltose but not sucrose (Petersen et al., 2004; Glinšek Biškup et al., 2021).
Serological Diagnosis of Tularemia
Serological methods for detecting antibodies against F. tularensis primarily utilize antigens derived from bacterial lipopolysaccharide (LPS) and, to a lesser extent, outer membrane protein A (FopA). A key focus in these assays is the use of antigens derived from the live vaccine strain (LVS) of inactivated F. tularensis subsp. holarctica (Maurin and Gyuranecz, 2016; Maurin, 2020; Nagaratnam et al., 2022). Common techniques used include tube agglutination, microagglutination, latex agglutination, complement fixation, and flow cytometry. Microagglutination, ELISA, and indirect immunofluorescence are the most widely used methods in clinical practice (Ohara et al., 1974; Maurin, 2020). Serological tests confirm tularemia by detecting seroconversion or a fourfold rise in antibody titers between acute and convalescent sera collected at least 2 weeks apart. Early testing and prompt treatment enable rapid diagnosis, reducing the need for surgery and prolonged hospitalization (Schöbi et al., 2022). A major limitation of serological diagnosis is antigenic cross-reactivity. Due to structural similarities of LPS among Gram-negative bacteria, particularly Brucella spp. and Yersinia enterocolitica, false-positive reactions may occur in serological assays. Although immunoblotting (Western blot) can enhance diagnostic resolution by demonstrating characteristic LPS banding patterns, it does not fully eliminate cross-reactivity-related limitations. Furthermore, variability in cut-off titers and the absence of fully standardized protocols contribute to interlaboratory differences in diagnostic performance (Maurin, 2020; Pohanka, 2026). According to WHO guidelines, serological confirmation of tularemia is based on compatible clinical presentation together with a positive MAT. However, these recommendations should be interpreted in light of methodological constraints and subsequent advances in diagnostic immunology. In particular, recent studies have highlighted the use of alternative antigenic targets, including highly purified and recombinant proteins such as FopA and other immunodominant antigens, which have demonstrated improved specificity and reduced cross-reactivity compared with LPS-based assays (Johansson et al., 2004; Maurin and Gyuranecz, 2016; Nagaratnam et al., 2022). A major limitation of serological tests is the lack of standardized protocols, leading to variable sensitivity, specificity, and accuracy. The absence of a universally accepted gold standard complicates their use as definitive reference methods (Maurin, 2020). A timeline of the relevant diagnostic tests for tularemia is shown in Figure 1.
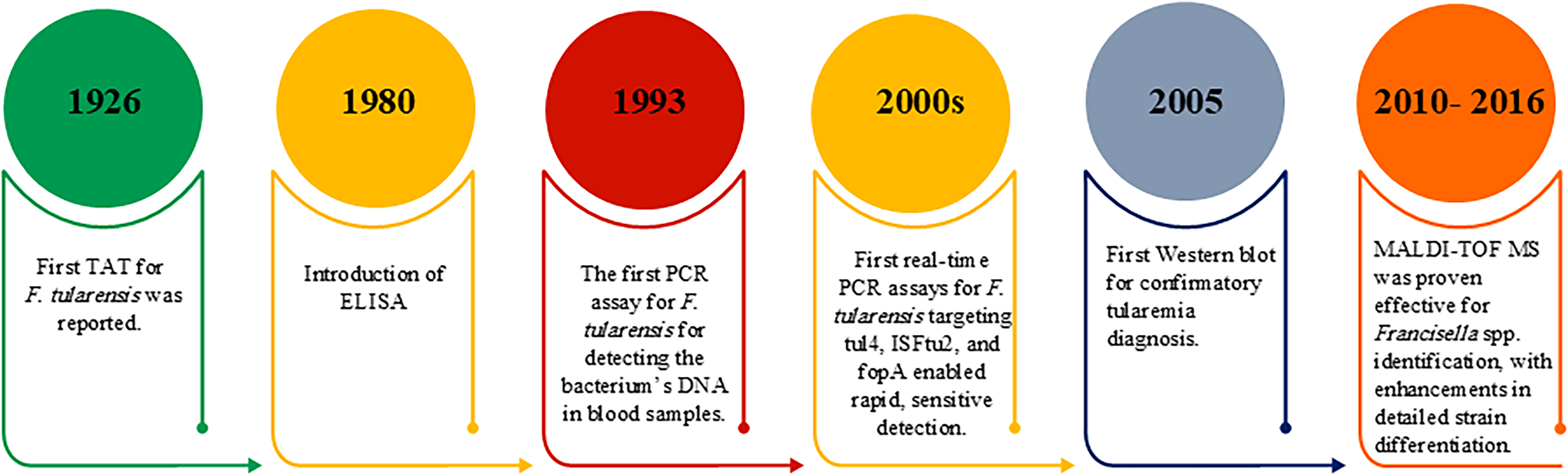
This timeline illustrates the progression of tularemia diagnostic tests from the 1926 until today. ELISA, enzyme-linked immunosorbent assay; MALDI-TOF MS, matrix-assisted laser desorption/ionization-time-of-flight mass spectrometry; PCR, polymerase chain reaction; TAT, tube agglutination test.
Agglutination
The tube agglutination test (TAT) and MAT is used to diagnose tularemia, with the MAT largely replacing the TAT due to its greater efficiency. MAT is faster (1–2 h versus a full day), simpler to perform and interpret, more sensitive in early diagnosis, and requires less antigen and serum than TAT (Table 1) (Brown et al., 1980; Maurin and Gyuranecz, 2016; Maurin, 2020). Studies by Yanes and Maurin showed microagglutination has high specificity (>98% at titers ≥80 to 160) but lower sensitivity (50–75%). At low titers (10–20), sensitivity is higher, though false positives increase due to cross-reactivity (Syrjälä et al., 1986; Yanes et al., 2018; Maurin, 2020). Another study revealed that when samples were obtained 7 days after the appearance of clinical symptoms, 100% sensitivity and specificity were demonstrated. However, cross-reactivity with Brucella and Yersinia enterocolitica spp., as well as Proteus spp., could potentially affect the accuracy of the results. Nevertheless, this issue can be partially addressed by considering the observed clinical manifestations (Porsch-Özcürümez et al., 2004).
Overall Advantages and Disadvantages of Methods of Identification of Tularemia

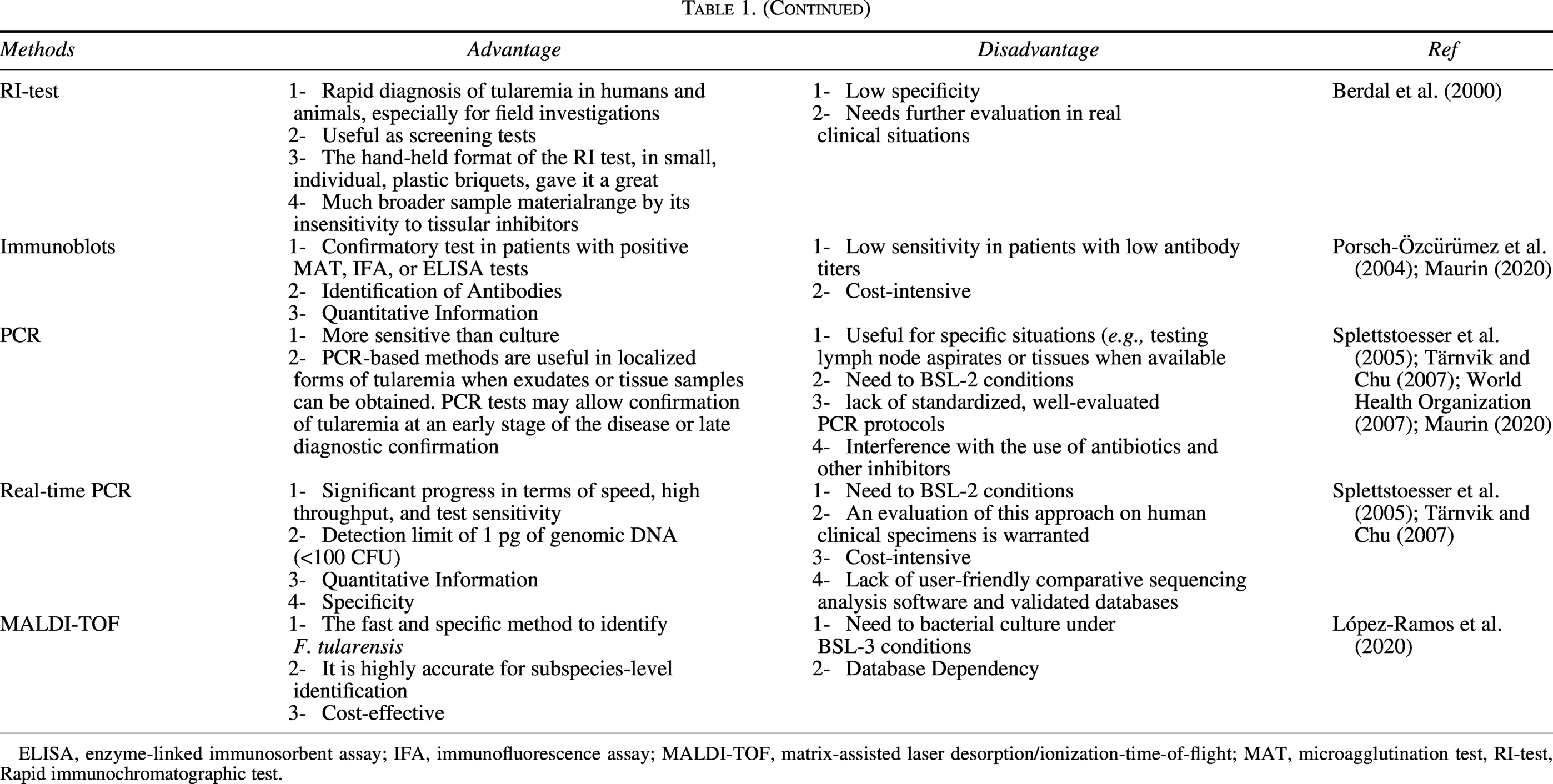
ELISA, enzyme-linked immunosorbent assay; IFA, immunofluorescence assay; MALDI-TOF, matrix-assisted laser desorption/ionization-time-of-flight; MAT, microagglutination test, RI-test, Rapid immunochromatographic test.
Indirect immunofluorescent assay
The process of antigen preparation for IFA is convenient and straightforward; however, challenges arise due to the requirement for a proficient technician. When the fluorescence intensity is low, accurately determining the final positive dilution (endpoint titer) becomes more critical for interpreting the individual. Nevertheless, a more precise assessment of the antibody titer can be achieved by establishing a reverse titer at the maximum dilution, where basic fluorescence observation remains feasible. Similar to the MAT, the practical sensitivity of IFA in authentic clinical situations might be decreased, primarily attributed to the inability to detect antibodies in sera collected during the initial 2 weeks following disease onset (Porsch-Özcürümez et al., 2004; Maurin, 2020).
Enzyme-linked immunosorbent assay
ELISA is another diagnostic test used for tularemia detection. Various antigens are utilized for conducting this specific test, with LPS and bacterial outer membrane protein being two of the most frequently employed. Through the implementation of the ELISA technique, it is possible to detect the presence of the antigen in serum and urine samples, as well as in the organs of infected animals, namely the spleen, liver, and lungs. In addition, even environmental samples, such as water and soil, can be examined to detect the presence of the antigen (Berdal et al., 2000; Splettstoesser et al., 2005). Various strategies for antigen preparation have been employed to evaluate the distinct immune responses after natural infection or vaccination. Following a sequence of preparations, pure LPS, pure outer membrane antigen, and bacterial cells from LVS and F. tularensis subsp. tularensis str. SCHU S4 was used for agglutination, ELISA, and Western blot tests (Syrjälä et al., 1986; Porsch-Özcürümez et al., 2004).
Following infection with F. tularensis, specific antibodies may persist for prolonged periods despite a gradual decline in humoral responses. Ericsson et al. (1994) demonstrated that anti-F. tularensis antibodies, particularly IgG, can remain detectable for up to 25 years after natural infection, even in the context of decreasing agglutinin titers over time. In serological assays such as ELISA, IgM, IgG, and IgA can be differentiated; however, IgG and IgM are most frequently detected in clinical and epidemiological investigations (Ericsson et al., 1994; Porsch-Özcürümez et al., 2004). ELISA allows for the separate identification of IgM, IgG, and IgA; however, IgG and IgM are commonly detected. Various methods are employed in ELISA, including complement-fixing ELISA (CF-ELISA) and sonicate ELISA (S-ELISA) (Maurin, 2020). In 2000, a research study led by Grunow et al. compared the effectiveness of capture enzyme-linked immunosorbent assay (cELISA), PCR, and manual immunochromatography-based techniques. The findings revealed that the ELISA exhibited greater sensitivity and specificity in differentiating between F. tularensis and F. tularensis subsp. holarctica (Grunow et al., 2000).
A study highlighted that combining the ELISA test with the Western blot confirmatory test significantly enhances the accuracy of tularemia diagnosis. This combined approach achieved an impressive specificity of 98.1% and sensitivity of 98.0%, reflecting its exceptional diagnostic precision. In addition, the positive predictive value was 10.2%, underscoring that, despite the relatively low predictive value, the method remains a precise and sensitive tool (Porsch-Özcürümez et al., 2004).
Several studies have demonstrated that ELISA exhibits an average sensitivity and specificity of 94.8% and 96.8%, respectively, and has been found to possess the capability of swiftly identifying specific antibodies. This was highlighted in the research conducted, which revealed that ELISA can detect specific antibodies within 1 to 2 weeks after the manifestation of clinical symptoms, and it also detects an increase in the titer at an earlier stage. In contrast, indirect immunofluorescence and microagglutination methods can only detect antibodies 3 to 4 weeks after the onset of clinical symptoms. Consequently, ELISA is more suitable for seroepidemiological studies because it is highly sensitive and specific (Yanes et al., 2018; Maurin, 2020). In addition, the structural similarities between antibodies targeting Brucella, Yersinia enterocolitica, Proteus vulgaris, and F. tularensis can lead to cross-reactions. This overlap in antibody specificity may complicate the interpretation of serological test results, emphasizing the need for careful consideration and confirmation when diagnosing infections using these methods (Wawszczak et al., 2022).
Rapid immunochromatographic test
The rapid immunochromatographic test is a rapid and convenient screening method that utilizes bacterial LPS to detect IgM, IgG, and IgA antibodies, typically within the early phase of infection (approximately 4–7 days postonset) (Grunow et al., 2000). While it may exhibit lower sensitivity and specificity than techniques such as MAT and ELISA, it enables the early identification of antibodies and potential diagnosis of the disease (Grunow et al., 2000). Due to the limited sensitivity of immunochromatography, negative results do not rule out tularemia and require confirmation by ELISA or PCR. providing qualitative results within ∼15 min and enabling rapid antibody detection in clinically relevant concentrations (Grunow et al., 2000; Schöbi et al., 2022; Bäck et al., 2023). These kits use lyophilized mouse monoclonal IgG on polyester pads, targeting F. tularensis LPS and conjugated with 30 nm colloidal gold. A red line in the test area indicates antigen presence (Berdal et al., 2000).
Flow Cytometry
A study found elevated CD3+/CD4–/CD8 + levels in patients with tularemia, with an 8% cutoff yielding 95.3% sensitivity and 89.5% specificity. Flow cytometry may enable earlier detection, approximately 7–14 days before serological assays become positive. PCR and real-time PCR are highly effective on deep tissue or lymph node samples before antibiotics, while flow cytometry is a reliable alternative if such samples are unavailable (Table 1) (Chrdle et al., 2019). Flow cytometry outperforms ELISA and rivals Western blot and MAT, but its routine use is limited by costly, specialized equipment and restricted availability (Porsch-Özcürümez et al., 2004).
DNA-Based Tools for the Diagnosis of Tularemia
PCR assay
PCR rapidly and accurately detects F. tularensis by amplifying specific DNA targets, providing high sensitivity, specificity, and safety (Johansson et al., 2004). It is beneficial for early diagnosis before antibiotics are administered and for testing tissues, including lymph node biopsies and wound swabs (Hepburn and Simpson, 2008). Common PCR targets include 16S rRNA, TUL4, FopA, and lpnA, ensuring reliable identification of F. tularensis (Grunow et al., 2000; Johansson et al., 2004; Sjöstedt, 2007).
It is challenging to detect Francisella using culture and PCR techniques due to the brief bacteremia period, limited sampling time, potential antibiotic administration, sample quality issues, and the presence of inhibitors (Yanes et al., 2018).
PCR on various pathological samples (lymph nodes, spleen, liver, lungs, kidney, intestine, and meninges) detected F. tularensis DNA in 14 of 16 cases, indicating that these samples are suitable for molecular testing. Inappropriate sample selection can lead to false-negative results (Johansson et al., 2004; Bäck et al., 2023).
Real-time PCR
Real-time PCR detects F. tularensis in human, animal, and water samples, including viable but nonculturable forms or bacteria within amoebae. It is also used to detect the organism from blood, spleen, lymph nodes, and vectors such as rodents, hares, insects, and ticks (Fooladfar and Moradi, 2023). In some studies, real-time PCR has replaced conventional PCR, enabling faster nucleic acid amplification and identification. It is 10 times more sensitive, detecting 10–100 cells per milliliter, with an error rate of less than 1% and a correlation coefficient of –1, indicating high reliability (Johansson et al., 2004). This method is more specific and efficient than conventional PCR, allowing detection of organisms even at very low concentrations (Table 1). Real-time TaqMan PCR targets genes such as tul4, ISFtu2, 16S rDNA, and fopA, enabling rapid and precise detection of F. tularensis (Fooladfar and Moradi, 2023). Further validation with human samples is needed. Despite its high accuracy, real-time PCR is limited to reference laboratories due to technical complexity, high costs, the need for skilled personnel, and the lack of user-friendly software and validated databases (Clarridge, 2004; World Health Organization, 2007).
Typing of F. tularensis
Due to the need for high-containment laboratories, culture and biochemical typing are often avoided. Molecular methods such as Pulsed-field gel electrophoresis (PFGE), whole-genome sequencing (WGS), multiple-locus variable-number tandem repeat analysis (MLVA), INDELs, and single-nucleotide polymorphisms (SNPs) enable the safe characterization of nucleic acids, facilitating the study of species differences, gene order, subspecies, genetic subpopulations, natural reservoirs, and transmission routes (Gyuranecz et al., 2012; Glinšek Biškup et al., 2021).
Whole-genome sequencing
WGS has provided numerous Francisella genome sequences, enhancing understanding of species differences, gene organization, subspecies, and genetic subpopulations. WGS can be performed on cultured organisms or directly from patient samples, but culturing is hazardous and time-consuming, and direct WGS remains expensive (Glinšek Biškup et al., 2021).
Single-nucleotide polymorphism
SNPs are point mutations that are evolutionarily stable in clonally reproducing organisms such as F. tularensis. This stability is valuable for categorizing bacterial populations into distinct genetic groups, allowing for accurate inference of phylogenetic relationships among these groups (Vogler et al., 2009; Wahab et al., 2014).
Multiple-locus variable-number tandem repeat analysis
Developing a typing system for F. tularensis requires methods that can distinguish isolates despite highly conserved genomes. MLVA targets variable genomic regions, making it a highly discriminative method. It can be applied to both direct and genomic samples, enabling rapid molecular characterization from patient specimens and supporting outbreak investigations and epidemiological studies (Johansson et al., 2004).
Pulsed-field gel electrophoresis
PFGE effectively differentiates Francisella species and subspecies and supports outbreak analysis, epidemiological tracking, and control strategy development. It provides insights into tularemia transmission and geographic distribution, though its ability to distinguish subtypes with high precision is limited (Johansson et al., 2004; Wahab et al., 2014). PFGE identified four distinct type A genotypes (A1a, A1b, A2a, A2b) and type B, providing a robust method to define Francisella genetic profiles, support outbreak investigations, and study bacterial diversity (Kugeler et al., 2009).
Matrix-assisted laser desorption ionization–time of flight
Rapid molecular methods enable precise identification and subtyping of F. tularensis at species and subspecies levels. In clinical labs, matrix-assisted laser desorption ionization–time of flight (MALDI-TOF) enables identification from culture colonies in under 15 min, eliminating the need for protein extraction, and can generate dendrograms for geographic typing. For detailed bacterial typing, higher-quality, reproducible spectra are required, offering faster and more cost-effective results compared with conventional overamplification methods that take several days (Table 1). BSL-3 precautions are essential when using colony-based MALDI-TOF methods (López-Ramos et al., 2020; de Vries et al., 2023).
Limitations and Perspectives
Current serological diagnostics for tularemia face multiple methodological and epidemiological challenges that limit their reliability, comparability, and applicability. The absence of a universally accepted gold standard complicates the accurate interpretation of diagnostic performance in studies. The clinical and epidemiological features of tularemia are often nonspecific, while culture methods exhibit low sensitivity (<10%) and require stringent biosafety conditions due to the highly infectious nature of F. tularensis. Although useful, PCR assays are generally applicable only to specific sample types and suffer from limitations, including difficulty in distinguishing closely related subspecies due to high genetic similarity, risk of false positives from environmental Francisella species, and restricted multiplexing capabilities resulting from limited fluorescent channels. Complex and expensive instrumentation further restricts their widespread clinical adoption.
Serological tests, considered the most sensitive diagnostic tools available, are constrained by their reliance on paired sera to confirm infection through seroconversion or a fourfold rise in antibody titers. In routine practice, the collection of such paired samples is uncommon, substantially reducing diagnostic sensitivity, particularly when clinical suspicion is delayed. Moreover, the lack of standardization in antigen preparation, assay methodologies, and cut-off titers across laboratories results in wide variability in reported sensitivities and specificities, complicating the comparison of results and limiting their generalizability. The clinical role of emerging serological assays, whether as high-sensitivity screening tools or high-specificity confirmatory tests, is frequently undefined. Furthermore, the characterization of false-positive, false-negative, and cross-reactive results remains insufficient, hindering their integration into routine diagnostic workflows. Molecular diagnostics also face barriers, including limited multiplexing capacity, risks of nonspecific amplification or contamination, and restricted accessibility in resource-limited environments due to high costs and the need for specialized personnel. In addition, successful empirical antibiotic treatment before a confirmed diagnosis may lead to underdiagnosis. Future research should focus on developing standardized protocols for antigen preparation and serological assay validation across diverse epidemiological settings with clear definitions of clinical application contexts. Efforts to streamline whole-genome sequencing workflows and reduce costs, potentially through hybrid short- and long-read sequencing approaches, as well as to create automated, user-friendly bioinformatics pipelines tailored for F. tularensis data, will facilitate wider adoption. Advances in portable, cost-effective real-time PCR platforms, such as microfluidic or CRISPR-based diagnostics, may improve accessibility in field- and resource-limited settings. Finally, the development of multiplex serological and molecular assays capable of simultaneously detecting F. tularensis alongside clinically similar pathogens will enhance diagnostic efficiency and accuracy. The integration of serological and molecular data with clinical and epidemiological information into robust diagnostic algorithms has the potential to improve case detection and public health responses.
Conclusion
Tularemia incidence has recently increased, even in previously unaffected areas, due to greater awareness and advancements in diagnostics. Natural reservoirs and human exposure risks influenced this rise. Diagnosing tularemia is challenging because its symptoms resemble those of other bacterial infections. While several serological assays exist, they often lack standardization and show variable sensitivity and specificity. Combining tests can enhance specificity but may decrease sensitivity. ELISA is particularly noted for its high sensitivity and early detection capabilities. Accurate diagnosis requires integrating serological and molecular techniques with clinical symptoms, patient history, travel background, and potential exposure to reservoirs. The integration of advanced biotechnological approaches, such as the production of high-affinity monoclonal antibodies and the use of recombinant antigens, has laid the foundation for a new generation of diagnostic tools, such as MALDI-TOF and biosensors. It has also led to advances in conventional techniques. While challenges related to cross-reactivity with pathogens such as Yersinia and Brucella, the development of antigen/antibody cocktails, and dual-mode detection systems continues to help improve diagnostic specificity. Developing more precise and standardized diagnostic assays is essential for confirming atypical cases and improving the consistency of clinical and epidemiological data worldwide.
Footnotes
Author Disclosure Statement
The authors declare that they have no conflicts of interest with each other.
Funding Information
This research received no external funding.