
Abstract
This article provides a discussion of the scientific, intellectual, and ideological frameworks that influenced Oparin’s formulation of the heterotrophic theory of the origin of life. Based on a Darwinian perspective, Oparin rejected the generally accepted idea that the first entities were photosynthetic microbes. He proposed instead that life had emerged through a gradual, stepwise, non-teleological process of prebiotic evolution that started with the abiotic synthesis and accumulation of organic compounds on primitive Earth. Influenced by Haeckel’s and Timiriazev’s evolutionary ideas and by biochemical oxidation processes proposed by Bakh, Oparin concluded that the first organisms were anaerobic heterotrophs that had evolved from colloidal aggregates such as gels and coacervate-like systems. In sharp contrast to proposals that explained the origin of life with the chance appearance of viruses or living molecules, Oparin’s theory connected the emergence of fermentative cells as the first life-forms with the early evolution of Earth. The construction of a stepwise, slow evolution of different stages suggested by Oparin with colloids and coacervate as models of precellular evolution separated the biochemical and chemical origin of life from the idea of spontaneous generation and led to the development of a multi- and interdisciplinary research program.
1. Introduction
A little over a century ago, Alexandr I. Oparin (1924) published a small book in Moscow entitled The Origin of Life. He was not the first to address this issue, but from the very beginning of his scientific career, Oparin placed the question of the origin of life within a Darwinian framework. In sharp contrast to those who assumed that the first organisms were naked genes or living molecules, he suggested that biological entities were the outcome not of the random emergence of a living molecule but of non-teleological historical processes of prebiological evolution that had depended on the self-assembly and evolution of sets of organic molecules of abiotic origin.
Although the discovery of ribozymes led to a series of proposals in which the idea of a “living molecule” was revived, most contemporary authors accept the possibility that the precursors of the first cells were polymolecular systems. Several candidates have been proposed, including liposomes, the so-called proteinoid microspheres, accumulation of organic compounds in mineral pores, and of course, coacervates, which Oparin incorporated in his 1936 book as models of precellular systems.
The purpose of this article is to review the intellectual and scientific environment that shaped this proposal. As summarized here, what began as models of cell precursors based on the ideas on cytoplasm prevalent in the first decades of the last century rapidly became an experimental alternative that was used by Oparin to avoid the risks of a reductionist perspective that assumed that the first biological entities had been living molecules that arose randomly by chance.
Ernst Haeckel: Monera and the Origin of Life
Despite the enormous admiration that The Origin of Species awoke in Ernst Haeckel, he was deeply disturbed by the suggestion of a divine act of creation with which Darwin concluded the second edition of his book. Although this statement was removed in later editions, Haeckel wrote in a lengthy footnote to his monograph Die Radiolarien that the chief defect of the Darwinian theory is that it throws no light on the origin of the primitive organism—probably a simple cell—from which all the others have descended. When Darwin assumes a special creative act for this first species, he is not consistent, and, I think, not quite sincere. (Haeckel, 1862)
Haeckel’s remarks reflect not only his unyielding secular commitment but also his strong adherence to cell theory, which he saw as an essential component of biology and which he completed with a Darwinian perspective. He had become acquainted with the proposal that cells are the basic unit of life through the writings of Schleiden (1847) who, like Schwann (1847), had concluded that cells are the basic unit of both animal and plant life. Together with Virchow, who famously wrote that omnis cellula ex cellula, that is, all cells come from pre-existing cells (Ribatti, 2018), Haeckel is recognized as one of the authors who shaped cell theory. His extraordinary understanding of microbial life led him to recognize the limitations of partitioning the living world into plants and animals, and he enlarged the existing taxonomic systems by proposing a third major biological kingdom, the Protista, in which he placed most microbial life-forms. He also included in the new kingdom the Monera, which were seen at the time as microscopic entities that for all purposes were mere small masses of non-nucleated protoplasm and that roughly correspond to what we now call prokaryotes (Reynolds, 2008; Richards, 2008).
As time went by, Haeckel varied the taxonomic boundaries of his newly proposed Protista kingdom, but he always kept within it the Monera. He consistently thought of the Protista as the group from which the ancestors of both plants and animals had evolved (Fig. 1) and, displaying enviable biological intuition, suggested that the nucleus was the abode of heredity. “Accordingly, insofar as we are able to regard the [proto]plasma chiefly as the nutritive component of the cell and, on the other hand as the reproductive component, we are justified in respect to the nucleus as the principal organ of inheritance and the plasma as the principal organ of adaptation,” he wrote in 1866 in the History of Creation. He added that in the case of Monera “where nucleus and plasma are not differentiated, we will have to regard the entire [proto]plasma as the common organ having both functions” (Haeckel, 1876).
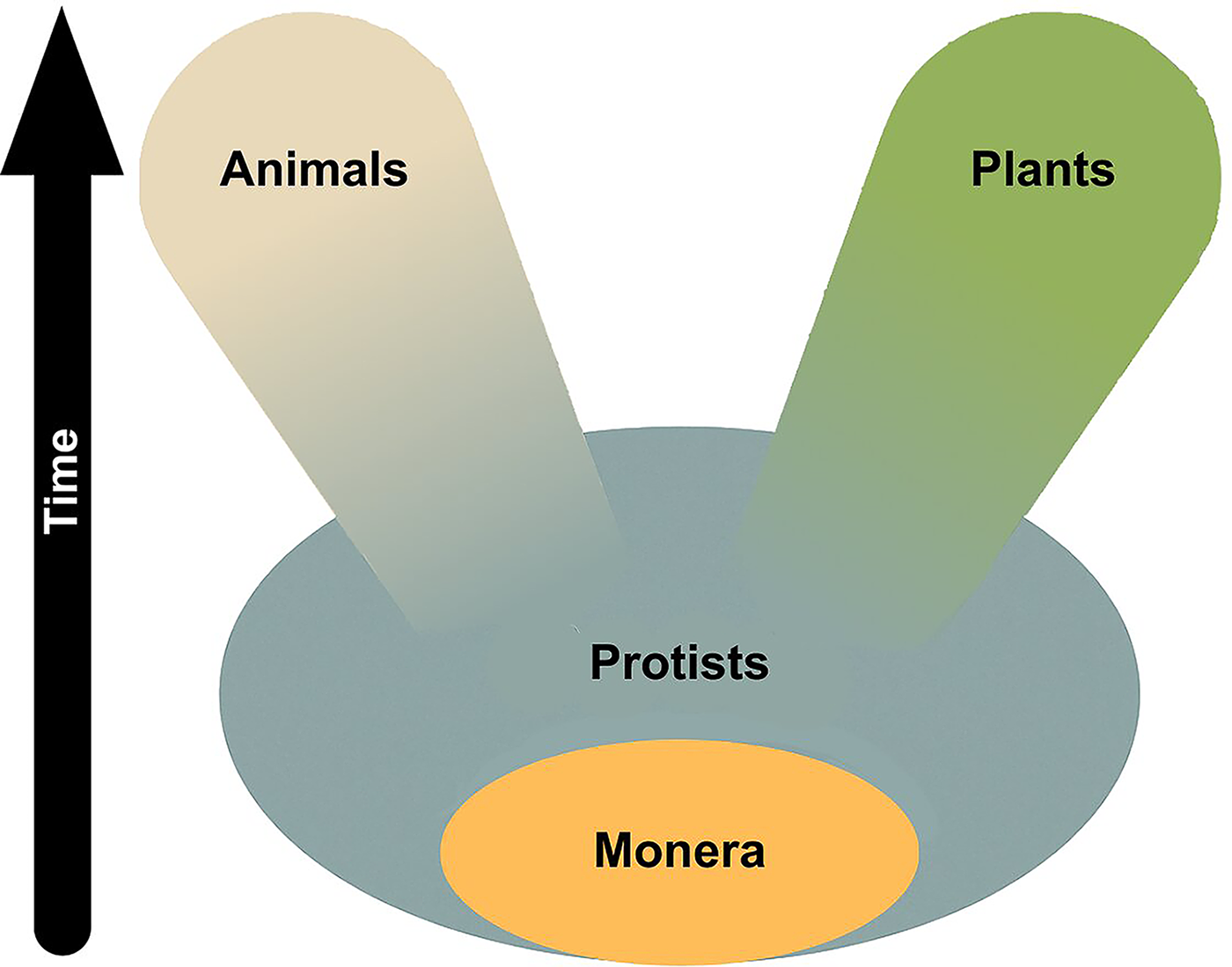
Schematic representation of Haeckel’s three-kingdom taxonomic system. The diagram shows Monera as a subgroup of the Protista and the evolutionary relationship of plants and animals with their protist ancestors.
Beginning with Kant’s nebular hypothesis on the origin of the solar system, Haeckel proposed in his writings an evolutionary scheme in which the first life-forms had been non-nucleated protein-rich blobs of protoplasm that appeared spontaneously (Fig. 2a). After describing in his History of the Creation the formation of Earth, he added that since water is a major component of protoplasm, “it was not until the terrestrial crust had so far cooled that the water had condensed into a fluid form, it was not till the hitherto dry crust of the Earth had for the first time become covered with liquid water, that the origin of the first organisms could take place” (Haeckel, 1876).
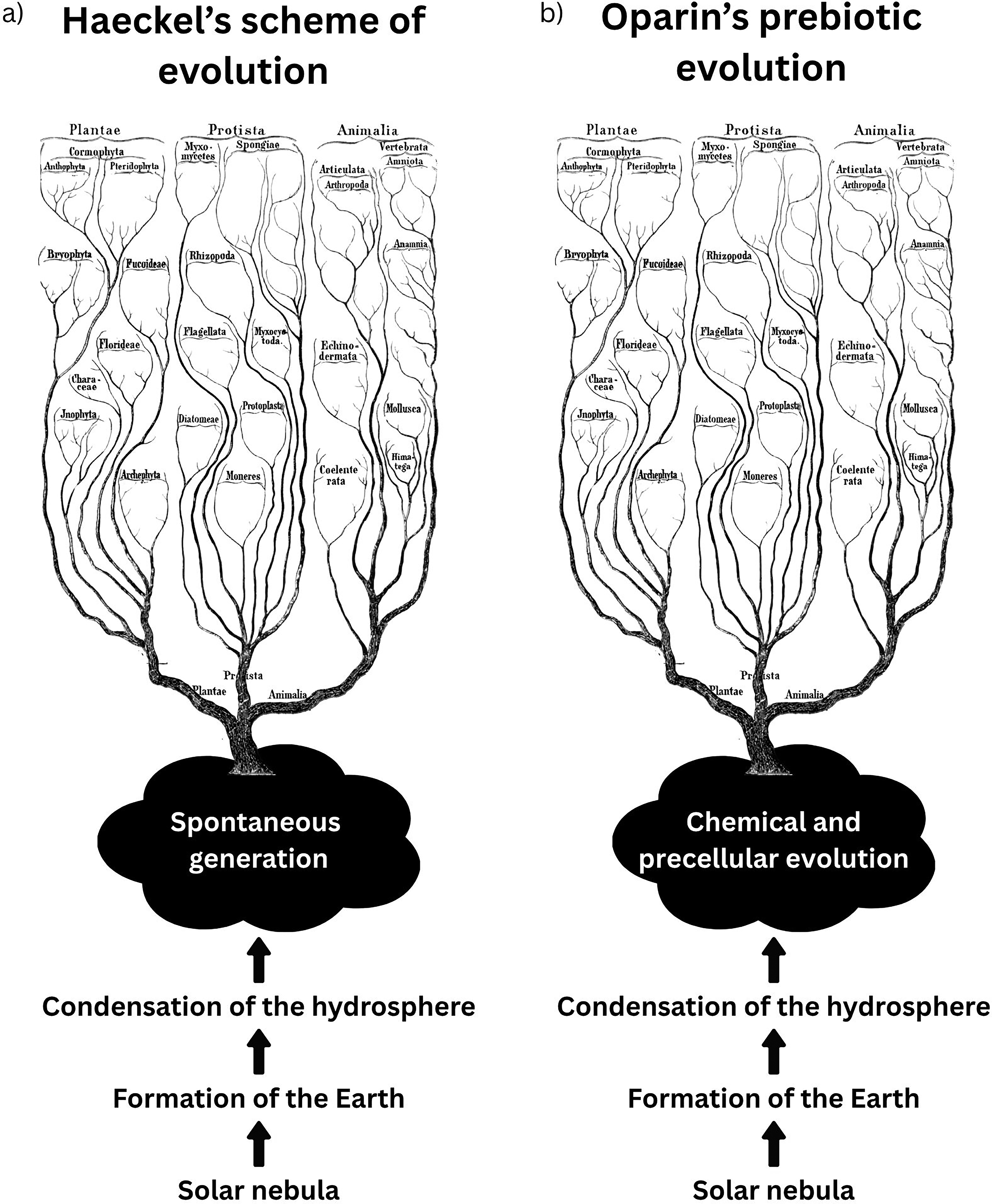
Haeckel’s books became the 19th-century benchmark of Darwinian evolutionary theory. His works were translated into many languages and read in many countries, and they became quite popular among Russian university professors and students (Vucinich, 1988). Like most of his fellow undergraduates, Oparin became familiar with his ideas. He adhered to the three kingdoms scheme until the very end of his life. Together with the teachings of his mentors Aleksei N. Bakh, a biochemist, and Kliment A. Timiriazev, a plant scientist, two internationally recognized researchers, Haeckel would remain a major influence in Oparin’s work and theories (Lazcano, 2016, 2024a, 2024b).
Well before the 1917 Soviet revolution, The Origin of Species had attracted the attention of Russian scientists and thinkers. Perhaps not surprisingly, Darwinism rapidly became a battle cry for many progressive and left-wing militants. Unfortunately, following the “rediscovery” of Mendel’s work in 1900, genetic mutations were considered by many as the fundamental source of evolutionary novelties in opposition to Darwin’s natural selection (Vucinich, 1988). This false dichotomy, which occurred long before Lysenko’s assault on genetics, was reflected in what became opposing scientific and political attitudes. By the time Oparin approached Bakh and Timiriazev, both had taken sides against Mendelian genetics.
Bakh’s youthful political activism had forced him to leave Russia, fleeing first to France and then to Switzerland, where he worked intensively in biochemistry of oxidation as the basis of metabolism and gained significant recognition. Following the downfall of the Menshevik government in 1917, he returned to Moscow, where he renewed his friendship with Lenin and, at his request, became involved in the reorganization of the Soviet scientific apparatus. Rightfully considered the founder of Soviet biochemistry, he organized a research group that was based at what became the Karpov Physical Chemistry Institute in Moscow.
Bakh had an extraordinarily dynamic vision of biological phenomena. He was convinced that the main characteristic of living beings is not their specific composition but the manifold of chemical transformations that continually take place in organisms. Driven by his interest in plant biochemistry, the 23-year-old Oparin joined his group, worked on photosynthesis and the biological chemistry of tea and beets, and rapidly became quite knowledgeable about not only the properties of enzymatic processes but also Bakh’s organizational and political skills.
Like Bakh, Timiriazev had also worked on the study of photosynthesis and the properties of chlorophyll. A highly educated researcher who had studied plant nutrition abroad, he was a strong adherent of evolutionary theory and became known as “Darwin’s Russian bulldog.” Along with other colleagues, he had resigned from Moscow State University in 1911 to protest against the repressive policies of the Czarist government, but he continued to give public lectures on Darwinism, plant biology, and the role of science in society.
Oparin became a regular follower of his talks, and his strong interest in evolutionary theory caught Timiriazev’s attention, who invited him to the weekly meetings he held with colleagues and other researchers in his Moscow flat. In that environment, the young Oparin was exposed to an extraordinary academic atmosphere that introduced him not only to the Darwinian literature available in the USSR but also, equally significant, to the techniques and methodologies of evolutionary analysis and the historical interpretation of life’s features (Lazcano, 2024a, 2024b). Very quickly, he began to view biochemical phenomena from an evolutionary perspective and discuss the application of Darwinian concepts at the subcellular level.
At the same time, Oparin’s understanding of biochemical mechanisms, metabolic pathways, and cell biology was deepening thanks to his close association with Bakh. He was keenly aware of the complexities of photosynthesis, which requires pigments, cellular membranes, and electron transporters, which he found difficult to imagine in the first living entities. This led him to suggest that the first living beings must have been heterotrophic anaerobes. This possibility was consistent with the universality of fermentative metabolism and its enzymes in both anaerobic and aerobic organisms, which, in Darwinian terms, implied that all present-day living beings descended from an anaerobic heterotrophic ancestor.
Oparin’s conclusion that the earliest life-forms were anaerobic heterotrophs represented a radical departure from the widely held view that the first organisms were photosynthetic autotrophs. Since his proposal required an abiotic source of organic compounds prior to the emergence of life-forms, he broke with Timiriazev’s view that the transformation of simple inorganic substances, carbonic acid and water, into the organic substance, starch, is the only natural process by which organic matter is formed on our planet. All organic substances, however diverse and wherever they are found, whether in plants, animals, or humans, originated in the leaves; they were formed from substances manufactured by the leaves. Nature possesses no other laboratory for the formation of organic matter except the leaves, or, more correctly, the chloroplast. In all other organs and organisms, organic matter is simply transformed; only here does it arise anew from inorganic matter (Timiriazev, 1912).
Oparin’s gratitude to Timiriazev never waned, but his proposal of a heterotrophic origin of life required a non-biological source of organic compounds. He did not perform any experiments but sought out and reinterpreted the rich tradition of 19th-century abiotic synthesis that had started with the laboratory formation of urea (Wöhler, 1828). Based on Mendeleev’s experiments and the work of other chemists who had achieved the laboratory synthesis of hydrocarbons and other organic compounds, Oparin proposed that abiotic processes had led to the prebiotic formation and accumulation of organic molecules in the primitive environment. His intellectual ability to compile and reinterpret isolated chemical phenomena is shown in his conclusion that prebiotic synthesis of organic compounds took place soon after Earth formed, based on astronomical observations of molecules such as HCN and CH+ in cometary spectra and reports of bitumen in chondritic meteorites (Oparin, 1924).
Prebiotic Syntheses and Precellular Systems
Oparin was not the only one to propose that the first living entities had been preceded by the abiotic synthesis of organic molecules. In 1924, the very same year in which he published his first book, his contemporaries Harvard microbiologist Richard B. Harvey (1924), Berkeley geochemist Charles B. Lipman (1924), and a few years later, distinguished evolutionist John B.S. Haldane (1929), who coined the lucky term “hot dilute soup,” argued independently that the first organisms were heterotrophs that had formed from a mixture of organic compounds present in the early environment.
However, there are important differences between these proposals. While it is true that Harvey (1924) and Lipman (1924) recognized the possibility that the first organisms were heterotrophic and therefore organic compounds were a prerequisite for the emergence of life, neither suggested a scheme of precellular evolution as detailed as Oparin’s nor a research program addressing the origin of the first life-forms.
Haldane’s case is quite different. Like Oparin, he profited from accumulated experimental results of research on the molecular mechanisms of CO2 fixation by plants, which many believed took place with H2CO as an intermediate. Haldane was well acquainted with the experiments of Edward Charles C. Baly, a University of Liverpool chemist whose interest in photosynthesis had led him to study the formation of sugars and amino acids from the ultraviolet irradiation of a CO2 solution. In his 1929 essay, Haldane reinterpreted these results and concluded that a similar process had given rise to the primitive soup, from which bacteriophages had formed and represented a transitional stage in the process leading to the first heterotrophic cells.
Haldane’s proposal of a virus-like intermediate stage in the evolutionary transition from a primitive soup to primordial heterotrophic cells (Haldane, 1929) can be understood in the context of the rediscovery not only of Mendel’s laws but also of Felix d’Herelle’s discoveries and Muller’s suggestion that perhaps viruses and genes were the same (Muller, 1926). The recognition of the role that genetic material played in the origin of life represented a significant advance in the field. Muller had been preceded by Troland (1914, 1917) and others who argued that the emergence of living systems was the outcome of the chance emergence of a living molecule; they confused the simple with the primitive and thought that the basic properties of life could be reduced to a single living molecule.
In sharp contrast with Muller, Oparin had a pre-Mendelian view of genetics and consistently adhered to Haeckel’s idea that both metabolic and hereditary processes resided in the protoplasm. For him, there was no such thing as a “living molecule” that had appeared by chance in primitive oceans. As he argued first in a lecture at the Moscow chapter of the Russian Botanical Society in 1922 (Kursanov, 1995) and then in his 1924 book, the first organisms were the outcome of a slow, stepwise, lengthy historical process of prebiotic evolution constrained by the laws of physics and chemistry.
From the very beginning, for Oparin, abiotic syntheses and accumulation of organic compounds were a necessary but not sufficient condition for the emergence of the first organisms. Protoplasm, he argued, had been preceded by gels and droplets of colloids formed by the interaction of proteins of abiotic origin and other molecules (cf. Lazcano, 2024a, 2024b). Muller was a staunch Mendelian mutationist, not a Darwinian evolutionist. His single-gene theory of the origin of life assumed that the first living being was a gene that appeared by chance in the primitive oceans and that from the very beginning was endowed with three defining traits: autocatalysis (i.e., replication), heterocatalysis (i.e., metabolism), and mutability, which he identified as evolvability.
During the first three decades of the 20th century, the frontier between chemistry and biology was constantly changing, but the prevailing view was that colloidal proteins were the basic component of protoplasm. From the outset, Oparin chose colloidal systems as models for the structures that had preceded the first cells. As he wrote in his first book, he based his work on the division of molecules into crystalloids and colloids that Thomas Graham had defined as early as 1861, although, as Oparin (1924) himself emphasized, over time it became clear that under certain conditions a substance can exhibit properties of one or the other.
“The study of the colloidal state is therefore of special importance, since the vast majority, if not all, of the substances from which protoplasm is made up have very large and complicated particles and therefore must give colloidal solutions” wrote Oparin (1924), adding that “colloidal substances do not give crystals, but they are still fairly easily precipitated as clots or lumps of mucus or jelly … the structure of coagulates is strikingly reminiscent of that of protoplasm.” By choosing colloids and gels formed by the synthesis and accumulation of hydrocarbons as models of precursors of the first cells, Oparin (1924) introduced the prebiotic syntheses and models of precellular evolution in his first book (Fig. 2b), avoiding the cul-de-sac spontaneous generation advocated by Haeckel’s scheme and the epistemological risks of the “living molecule” hypothesis.
The Philosophical Battleground: The 1924 and 1936 Period
Oparin developed his initial ideas and published his first book during the harsh times of the Civil War that followed the 1917 Bolshevik takeover of the government. He was a teaching assistant at the Natural Science Division of the Moscow State University during a period when heating, water, laboratory equipment, and substances for teaching and research were often lacking. In 1922, Oparin went abroad and completed an academic stay in Heidelberg, in Albrecht Kossel’s group. Kossel had received the Nobel Prize in 1910 “in recognition of the contributions he made to our knowledge of cell chemistry through his work on proteins, including nucleic acids” (Nobel Prize Outreach, 2025). He was known to welcome foreigners in his research group (Kohler, 1982).
Oparin did not succumb to the seduction of nucleic acids. Like the vast majority of his contemporaries, he continued to consider colloidal proteins as the metabolic and hereditary basis of life. Nevertheless, Oparin’s first book is an example of intellectual versatility and admirable fresh scholarship. It was published during a critical period of major social and political changes in the Union of Soviet Socialist Republics (USSR). Those years witnessed an enormous creativity and incomparable intellectual freedom; but, as reflected in the poignant pages of Boris Pasternak’s Doctor Zhivago and Walter Benjamin’s Moscow Diaries, this period came to a tragic end when Stalin took over the Bolshevik government in 1928.
That very same year, Stalin’s first Five-Year Plan began. It included a thorough restructuring of the scientific apparatus that forced researchers to support the Soviet regime. As Graham (1972) has described, during this period, many Bolsheviks began to occupy positions in the Academy of Sciences, which soon became the governing body of Soviet science. Driven by the Politburo, the ideologization of the USSR’s scientific apparatus imposed dialectical materialism as the official philosophy of the country. This forced writers, artists, and scientists to accept a Marxist framework. The mandatory philosophical premises and general laws on the development of nature shaped not only hypotheses and experimental work but also theories in which the rejection of mechanistic materialism that reduced biological phenomena to physics and chemistry was imposed (Joravsky, 1960; Graham, 1993).
Oparin joined this trend with considerable enthusiasm. Although he was never a card-carrying Communist Party member, from his youth, he had been intellectually and scientifically linked to Timiriazev and Bakh, who were part of the upper echelons of the Soviet establishment and who undoubtedly aided his political advancement. Like many others, both in the USSR and in other countries, Oparin was convinced that the non-reductive principles of dialectical materialism could shape the framework in which his work could be based. He dived into dialectical materialism, matured his original views, and abandoned the unsophisticated, simplistic, and crude materialism of his first book. Based on a new philosophical perspective, he described the origin of life as an emergent property that resulted from qualitative transformations in which physical and chemical processes gave way to biological processes defined by natural selection.
During this period, Oparin wrote several essays and popular articles in which he emphasized the role of Marxism in the formulation of an evolutionary perspective of living phenomena. More importantly, Oparin published in 1936 a second book, also titled The Origin of Life, which was translated into English 2 years later (Oparin, 1938). The new volume is an extraordinary example of Oparin’s intellectual and scientific maturity. Contrary to what has been stated (Muller, 1966; de Duve, 1991), it was not written at the request of the Soviet authorities. The book starts with a detailed historical description, from a Marxist perspective, of past theories and hypotheses on the origin and nature of life. However, the most significant contribution is the extraordinarily detailed evolutionary narrative that starts with an exhaustive analysis of the available astronomical literature, the reinterpretation of a large corpus of reports of organic chemistry syntheses, a discussion of colloidal systems, and a critical summary of biochemical data and microbial metabolisms, all of which constitute a refined multi- and interdisciplinary research program that shunned the reductionism risk of a “living molecule” concept.
It is striking that in spite of the rigidity of a government-imposed philosophy framework, Oparin found the intellectual freedom to develop non-reductive principles of natural law within a historical narrative that links the emergence of life to the environmental conditions of primitive Earth. He adopted Mendeleyev’s proposal on the origin of petroleum and argued that the iron carbides present in primitive Earth had reacted with water vapor to form hydrocarbons, which, when oxidized, produced alcohols, ketones, and aldehydes that, after reacting with ammonia, formed amines, amides, and ammonium salts. These compounds became precursors of proteins, which, when dissolved in the primitive hydrosphere, formed colloidal systems such as coacervates, in which the first metabolic pathways arose and gave origin to the first living beings (cf. Lazcano, 2016).
It is in this context that we must understand Oparin’s abandonment of the colloid droplets and gels discussed in his first book and his incorporation of coacervates as models of precellular systems. His second book is neither a theory of coacervates nor a proposal for the primacy of metabolism over hereditary material. It is a highly refined non-reductionist theory on the origin and early evolution of life. Like his mentors Timiriazev, Bakh, and many others, Oparin remained convinced of the incompatibility between Mendelism and Darwinian evolution and continued to follow Haeckel’s assumption that in the non-nucleated Monera, the protoplasm was the abode of both inheritance and metabolism, that is, biological inheritance could be explained in terms of the growth and division of the bacterial protoplasm. Unfortunately, in what is the least worthy period of his personal and scientific biography, Oparin ended up aligning himself quite willingly with Lysenko and hence with Stalin.
Chance and Necessity in the Origin of Life: The Conflict Between Monod and Oparin
The synthesis of amino acids and other organic compounds under possible prebiotic conditions reported by Miller (1953) is a major milestone in the modern study of the origin of life, and it was seen by many as proof of Oparin’s premises. It is frequently forgotten, however, that Miller’s results appeared 20 days after the publication of Watson and Crick’s (1953) double-helix DNA model. At first, the latter attracted little attention from the press, but the birth of molecular biology changed life sciences forever by imposing a perspective that, for many years, led to a reinforcement of a reductionist perspective that severely affected organismic biology and other areas of life sciences.
Inevitably, this led to the study of the origin of life being inscribed within the context of this new molecular framework of biological phenomena. It was not until the synthesis of adenine and other purines under possible primitive conditions reported by Oró (1960) that prebiotic chemistry and molecular biology finally met. During the first 20 years following Miller’s (1953) experiment, attempts to understand the origin of life were shaped to a considerable extent by unraveling the details of DNA replication and protein biosynthesis.
The recognition that RNA was the genetic polymer of a number of viruses, combined with extraordinary insights from Belozerskii (1959), Handler (1963), and Eakin (1963) on the key role of ribonucleotide cofactors in enzyme catalysis, led to proposals of an ancestral metabolism based on coenzymes. These hypotheses fit well with independent suggestions from Rich (1963), Woese (1967), Crick (1968), and Orgel (1968) that the first living entities were devoid both of DNA and proteins, a hypothetical possibility that would lead to the first formulations of the so-called RNA World hypothesis (Lazcano, 2012).
There are many definitions of the RNA World, including several contradictory ones. As summarized elsewhere (Negrón-Mendoza et al., 2025), one could say that it is an early, perhaps primordial, stage during which RNA molecules played a much more conspicuous role in heredity and metabolism. A comprehensive view of this hypothetical stage should add a wide range of molecules, including ribonucleotides and their derivatives, which play directly or indirectly manifold roles in different biological processes. Orgel (1968) emphasized the widespread presence of nucleotide coenzymes and modified ribonucleotides in metabolic pathways, which can be interpreted as an indication of an early intertwining of genetic material and biochemical catalysis.
With considerable insight, Haldane wrote in 1954 that the long-chain polymers found in living organisms have “back-bones,” composed of phosphate (i.e., nucleic acids), glycine or pentose residues. The first seem to be the most catalytically active, and may be the most primitive. The critical event which may have best be called the origin of life was the enclosure of several different self-reproducing polymers within a semipermeable membrane (Haldane, 1954).
As shown by the writings of Jacques Monod, not all shared this integrative scientific perspective.
After spending some time in the United States in the group led by Thomas H. Morgan, the great promoter of Mendelian genetics, Monod returned to the Sorbonne, where he quickly began working in enzymology and soon became a towering figure in molecular biology. He was a free spirit that had joined the Resistance, but he broke all ties with the pro-Soviet French Communist Party when he learned that in a session of the USSR Academy of Agricultural Sciences Lysenko had harshly condemned Mendelian genetics, accusing it of being reactionary and opposed to dialectical materialism. Monod was a talented writer, and in 1970, he published the first edition of his work Chance and Necessity (Monod, 1970), a beautifully written book that soon became a scientific classic and catalyzed a series of interpretations and controversies that reverberate to this day.
It was one of the heydays of molecular biology, and in his book, Monod emphasized the importance of the molecular description of the genetic code, which, as he wrote, allowed for the construction of a purely physical, material theory of inheritance, which he described as the “secret of life.” Monod wrote that it was usually believed that the discovery of the basic universal mechanisms for the essential properties of living beings would have helped solve the problem of the origin of life, but the new discoveries showed that the emergence of the first organisms is more difficult than it previously seemed.
When Monod wrote his book, Miller (1953), Oró (1960), and others had already demonstrated the abiotic synthesis of amino acids, adenine, sugars, fatty acids, and many other monomers. Moreover, analysis of the 4.6-billion-year-old Murchison meteorite had proven that a range of organic molecules had formed in the early solar system, which supported the possible existence of a primitive soup. It is unfortunate that Monod did not consider these results when arguing in his book that the origin of the first organisms was the result of mere chance. The material nature of life could be partly understood through the physical and chemical properties of the structures, functions, and interactions of nucleic acids and proteins, but at that time, the question of how these molecules appeared and how the genetic code emerged seemed unanswerable.
Oparin had incorporated DNA and protein synthesis since 1957 in his analysis of the origin of life and repeatedly rejected the possibility of a “living molecule” together with the teleological and reductionist views that characterized the work of many molecular biologists. Equally significant, he consistently refused to grant chance an ontological role. Monod’s position was the opposite, since for him, the emergence of life on Earth was a highly improbable accident that occurred thanks to a fortunate combination of cosmic dice that could hardly be repeated elsewhere in the universe.
This scientific antagonism must be seen in the context of the intense ideological struggle and political tensions that separated Monod from Lysenko and Oparin. With few exceptions, the evolutionary perspective was absent in molecular biology, which was dominated by a highly reductionist view. Many of its practitioners were convinced that the essential features of subcellular processes could only be explained by random mutations. As Mayr (1988) wrote, “Although Monod strongly opposes determinism, including the invocation of final causes, he completely ignores natural selection as a creative process and attributes all evolution to pure chance.”
The Prebiotic Soup: A Cold Gazpacho or a Boiling Broth?
The possibility that the first living beings had first evolved in the sea is deeply rooted in the intellectual traditions of Western naturalists. It is depicted in the splendid illustrations of the Carmelitan monk Francesco Maria Soldini (Barbagli, 2009), published before Lamarck’s 1809 Philosophie Zoologique, where the French evolutionist discussed the appearance of life in the warm waters of the ocean. The primitive ocean’s dimensions were reduced to a puddle when Darwin wrote of a warm little pond, but they recovered their planetary dimensions when Haeckel stressed that, given the abundance of water in the protoplasm, microbes could not have appeared until early Earth had cooled down and the hydrosphere could condense.
As underlined independently by Oparin and by Haldane, who coined the famous metaphor of a “hot dilute primitive soup,” the process of syntheses and accumulation of organic molecules had taken place in the waters of the primordial oceans (Lazcano, 2011). However, Bernal (1944) was perfectly aware that in an open ocean, the probability of encounters between different organic compounds of abiotic origin was very low, and he proposed that molecular interactions would have been facilitated in clays; thus, he introduced the concept of microenvironments on early Earth in prebiotic evolution.
Synonymous terms such as “primitive soup,” “primordial broth,” or “Darwin’s warm little pond” have led in some cases to major misunderstandings, including the simplistic image of a worldwide ocean, rich in self-replicating molecules and accompanied by all sorts of biochemical monomers (Lazcano, 2024a, 2024b). Nowadays, however, such terms would refer more precisely to parts of the prebiotic environment where the accumulation and interaction of the products of abiotic synthesis may have taken place, including oceanic sediments, intertidal zones, shallow ponds, membrane-bound systems, freshwater lakes, and lagoons undergoing wet-and-dry cycles. The soup may have been semi-frozen, and glacial ponds where evaporation, eutectic separations, or other physicochemical mechanisms such as the adherence of biochemical monomers to active surfaces could have raised local concentrations of organic molecules.
An eclectic approach to the conditions that led to the origin of life is probably the best alternative. Not all environments on early Earth would have facilitated the synthesis and accumulation of organic compounds or the formation of precellular systems. Quite surprisingly, the formation of membranous vesicles is affected by the properties of marine waters (e.g., salt and divalent cations). As reported by Milshteyn et al. (2018), there is difficulty in forming fatty acid vesicles in seawater due to high ionic concentrations, which disrupts the membrane’s stability.
Hydrothermal vents must have played a key role in the primitive terrestrial geochemical cycles, but in spite of their considerable popularity, it is unlikely that they met all the conditions to act as a cradle of life. Amino acids and other biomolecules rapidly decompose at timescales of minutes at the high temperatures (>350°C) associated with most vent discharges (Bernhardt et al., 1984; Miller and Bada, 1988; Bada et al., 1995), and at pH 7, the half-life of RNA is 2 min at 250°C for the hydrolysis of every phosphodiester bond. The possibility of forming liposomes and coacervates would also be significantly hampered at the pH, temperature, and ion concentrations present in hydrothermal vents. Liposome formation has been achieved in cool and fresh waters (Maurer, 2017) and in cycles of hydration and dehydration (Damer and Deamer, 2020) but not in experiments that simulate submerged hydrothermal vent conditions.
Coacervates and Precellular Evolution
Coacervation is a self-organizing phase separation process originally described by Bungenberg de Jong and Kruyt (1929) that rapidly became a model of protoplasm and, from Oparin (1936) onward, experimental systems that could model precellular systems. Why did Oparin use coacervate droplets instead of double lipid membrane-bounded liposomes? It is true that the earliest proposals of a lipidic cell membrane were published toward the end of the 19th century, but their existence was held in doubt as late as 1921 (Lombard, 2014). Liposomes were discovered in 1961 but not reported until 1964 (Bangham and Horne, 1964), and although soap-forming micelles had been described since 1913 (McBain, 1913), their composition clearly excluded them as models of precellular structures.
The popularity of coacervates as physicochemical models of cells was clearly linked to the idea of protoplasm, which was eventually displaced by cytoplasm, a concept that had been proposed in the 19th century but took a long time to be incorporated into mainstream biology. Although colloidal chemistry experienced a significant decline in the 1940s, as noted by Ling (1984), “in the Soviet Union, some biologists continued to describe living phenomena in terms of colloidal chemistry.”
The formation of coacervates can be easily understood in terms of the interaction of different macromolecules that lead to the self-assembly of liquid-phase droplets. Oparin used a variety of molecules of biological origin, which included proteins, polyamines, polysaccharides, and nucleic acids, including RNA, to experimentally demonstrate not only the concentration of different reactants but also the exchange of ions and small monomers with the surrounding media and other compartments (Oparin, 1968). He demonstrated a variety of reactions that occurred preferentially within the coacervates formed in a distilled aqueous environment. The presence of enzymes within the coacervates allowed for hydrolytic degradation of polysaccharides or polynucleotides, or oxidation–reduction reactions within the internal environments of stable coacervates (Oparin, 1986).
Contemporary understanding of liquid–liquid phase separation has revealed that coacervation processes underlie the formation and function of membrane-less eukaryotic organelles, including nucleoli, stress granules, and the so-called P-bodies (Hyman and Brangwynne, 2012; Yewdall et al., 2021). The biomolecular condensates play crucial roles in cellular organization and regulation, validating core aspects of Oparin’s original insights on compartmentalization without membranes.
Current reassessments of coacervates in prebiotic contexts have demonstrated their remarkable versatility. Recent studies have shown that coacervates can serve as sophisticated enzymatic microreactors, concentrating substrates and enzymes while providing molecular crowding effects that enhance reaction rates (Mason et al., 2017; Drobot et al., 2018). Advanced coacervate systems now include synthetic polymer-based, peptide-based, and nucleic acid-based variants; each offers distinct advantages for biotechnological applications. Notably, dipeptide coacervates have emerged as particularly promising artificial membraneless organelles, capable of biorthogonal catalysis and cellular uptake for therapeutic applications (Cao et al., 2024).
Together with liposomes (Deamer and Pashley, 1989) and coacervates, porous rocks, clays and other chemically active surfaces (Bernal, 1944), emulsion droplets, DNA and RNA self-replicative origami systems (Joyce and Szostak, 2018), and self-organizing assemblages of RNA and DNA that are formed in the absence of membranes (Todisco et al., 2018) indicate that a wide variety of systems may have formed in the primitive environment that could have played key roles in the evolution of systems that led to the first cellular systems. In retrospect, many of Oparin’s results of decades of work on coacervates may appear naïve. However, throughout those years, Oparin defended the idea of precellular evolution to insist that the abiotic synthesis of organic compounds was a necessary, but not sufficient, condition for the emergence of life.
Can Life Be Synthesized?
“There still remains, however, the problem of the artificial synthesis of organisms but for its solution a very detailed knowledge of the most intimate, internal structure of living things is essential,” wrote Oparin (1936) in the closing pages of his 1936 book.
Even the synthesis of comparatively simple organic combinations can be accomplished only when one possesses a more or less complete understanding of the atomic arrangements of their molecules. This, of course, would apply even more so in the case of such complex systems as organisms. We are still too far removed from such a comprehensive knowledge of the living organism to even dream of attempting its chemical synthesis. For the present research into the origin of life must, therefore, be restricted to studies of a purely analytical character.
We are faced with a colossal problem of investigating each separate stage of the evolutionary process as it was sketched here. We must delve into the properties of proteins and learn the structure of colloidal organic systems, enzymes, protoplasmic organization, and so on. The road ahead is hard and long, but without doubt it leads to the ultimate knowledge of the nature of life. The artificial building or synthesis of living things is a very remote, but not an unattainable goal, along this road.
Despite Oparin’s optimism, the artificial synthesis of life is still a remote target (Porcar and Peretó, 2018). There are major gaps in our current narratives of the emergence of life. The transition from autocatalytic chemical networks to heritable information systems is a poorly understood process that remains an open issue. The spatial organization of prebiotic chemistry presents unresolved questions about how molecular complexity emerges in heterogeneous environments. Computational models of autocatalytic network research exhibit huge gaps in experimental validation of theoretical frameworks. Finally, the integration of multiple organizational levels (from molecular autocatalysis to compartment formation to protogenetic systems) represents a major conceptual challenge.
Charles Darwin wrote in 1863 that “[i]t is mere rubbish thinking at present of the origin of life; one might as well think of the origin of matter….” It is not a phrase to despair about because, as Bernal (1944) stated,
[T]his does not mean that we should accept wild hypothesis of the origin of life or of matter, which simply conceal ignorance, but rather that we should attempt almost from the outset to produce careful and logical sequences in which we can hope to demonstrate that certain stages must have preceded certain others, and from these partial sequences gradually built up one coherent history. There are bound to be gaps where this cannot be done, but until the process is attempted, these gaps cannot be located, nor can the attempt be made to fill them up…
Conclusions
No molecule is alive by itself, but the awareness of the role of nucleic acids led to major misunderstandings and to blatant expressions of biological reductionism in which chance was given an ontological role. Following the in vitro synthesis of DNA achieved by Kornberg and his group (Adler et al., 1958), Muller stated that “those who define life as I do will admit that the most primitive forms of things that deserve to be called living had already been made in the test tube by A. Kornberg” (Muller, 1960). As summarized in Figure 3, for Muller (1926, 1960), the origin of life was the result of a chance appearance in the primitive waters of the planet of a gene that, several decades later, he exchanged with a living DNA molecule.
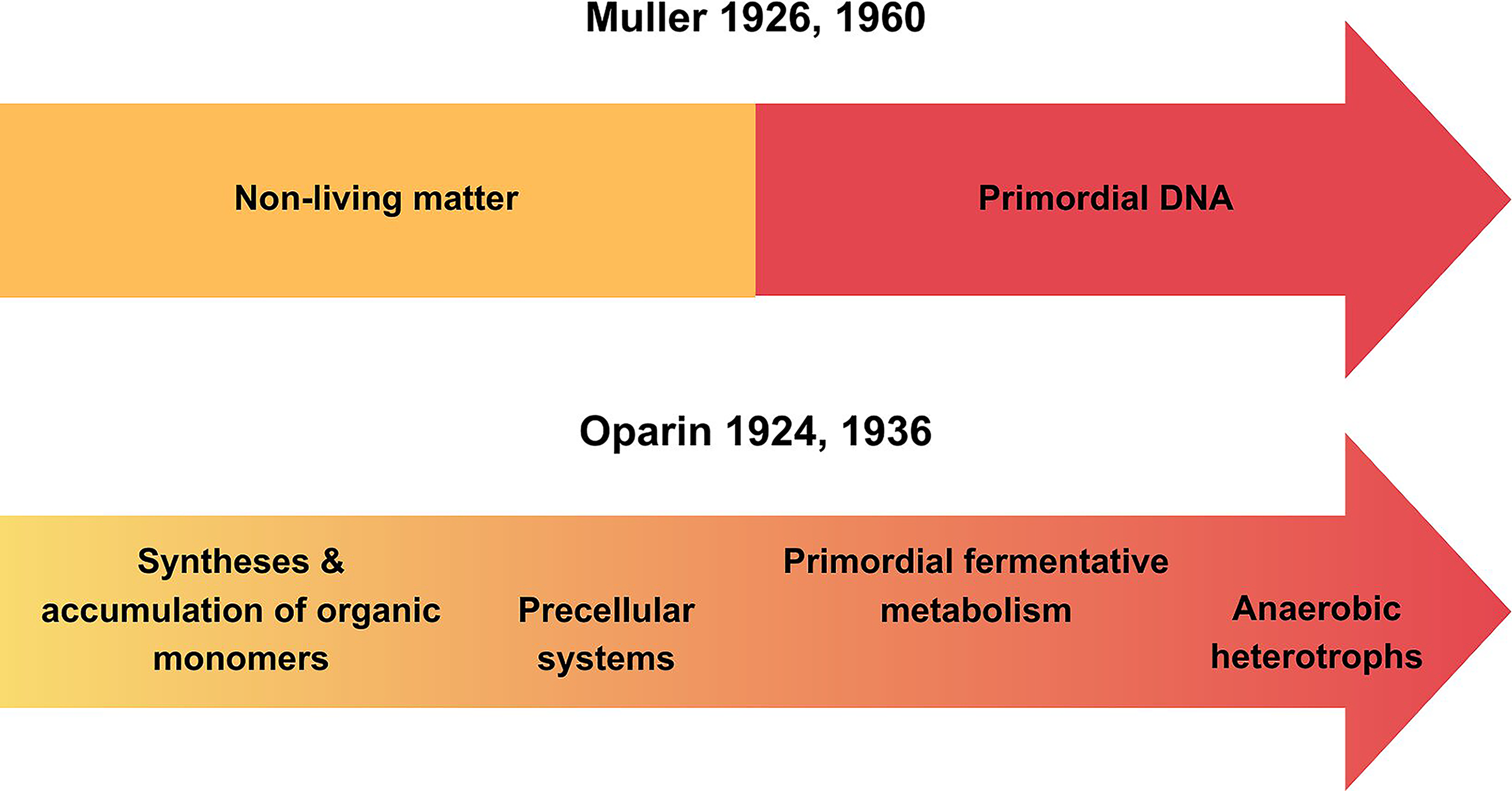
Contrasting models for the emergence of life. The upper arrow illustrates Muller’s reductionist model that assumes a direct transition from non-living matter to a primordial gene or DNA. The lower arrow depicts Oparin’s stepwise scenario, in which life is the outcome of gradual events starting with the syntheses and accumulation of organic compounds, followed by precellular systems in which primordial fermentative metabolism emerges, from which the first anaerobic heterotrophs evolved.
Life depends on the interactions and functional integration that exist between its molecular components and between the entire system and its environment. These interactions are the result of the evolution of the system as a whole. In other words, the development of increasingly refined and close correlations between the components of living systems is the outcome of a historical process that apparently took place very rapidly in evolutionary terms. What Monod (1970) failed to understand and Oparin (1972) appreciated very well is that the structure and functional properties of the molecular components of cells, along with the subcellular systems they form and their complex adaptive properties, can only be understood as the historical results of a combination of physicochemical constraints and natural selection. This reflects the contingent nature of biological evolution.
However, the history of the biosphere is not a mere sequence of chance events. Natural selection can overcome contingent effects to some extent, but chemical and biochemical constraints may be of greater importance than we generally recognize. To understand the nature of life, we must recognize both the limits imposed by the laws of physics and chemistry and the contingency of history. Biology is a historical science, not a subdiscipline of physics.
Cells consist of individual chemical components and systems of interacting molecules. Many of these subcellular units are functional modules, and even when isolated, their biochemical and physicochemical properties, as well as those of their individual components, are retained. Therefore, it is not surprising that some of these components exhibit, on their own, properties that we associate with living beings, such as catalysis, replication, or self-organization.
It is easy to recognize that the biochemical properties of subcellular systems and their components (such as nucleic acid replication, enzyme catalysis, and membrane-forming properties) are the historical result of the evolutionary amplification of the basic physicochemical properties that their simpler molecular predecessors also possessed. In general terms, it can be said that all living entities share the same basic molecular processes, biochemical composition, and biomolecular structures. The differences that separate them are not the components that constitute them but the evolutionary history that shaped them.
The extraordinarily refined functional and structural properties of different biological molecules, which Oparin (1957, 1972), for lack of adequate terms, called “intentionality” or “purposefulness,” could not have appeared spontaneously on early Earth. The structure and functional properties of the molecular components of cells, along with the subcellular systems they form and their exquisite interaction, can only be understood through the combination of their physical and chemical characteristics and their complex adaptive interactions that reflect historical processes of evolution at the subcellular level. These refined properties and interactions are the result of the evolution of the system as a whole, and their basic features must have been established during the poorly defined period that connects the prebiotic phase of synthesis and accumulation of organic compounds with the first biological entities.
Footnotes
Acknowledgments
This work is based in part on the many conversations that one of us (A.L.) had the opportunity to enjoy with A.I. Oparin many years ago and is indebted to the late Professor Loren Graham, with whom I had several exchanges on the life and work of A.I. Oparin and the complex times he lived. He also thanks Prof. Dr. Uwe Hossfeld and Dr. Georgy Levit (Friedrich-Schiller Universität Jena) for useful conversations on the scientific biography and contributions of Ernst Haeckel. The results discussed here are based on previously published material (Lazcano, 2016, 2024a, ![]() ).
).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Associate Editor: Manasvi Lingam