
Abstract
To address the chemistry responsible for life’s emergence, we examine the simple chemicals accessible on early Earth and the processes that could have transformed them. We focus on carbon reduction as a process central to the direct precursors of life. Two main approaches for the emergence of life’s building blocks and their associated functions are discussed. One approach investigates chemical routes for forming life’s building blocks via processes that worked differently from those of extant life, referred to as “unrestricted” prebiotic chemistries. The other approach establishes a direct mechanistic connection between prebiotic building blocks and extant biochemistry, referred to as “lifelike” prebiotic chemistries. Significant gaps remain in our understanding of early Earth’s conditions, which makes it difficult to constrain the possible locations and mechanisms of life’s emergence and the sources of precursors. Although numerous differences remain between the main origin-of-life hypotheses, consideration of the geochemistry of carbon reduction is a common denominator of these historically divided hypotheses.
1. Introduction
To understand the processes that gave rise to life, we must first determine what precursor molecules for building block synthesis early Earth likely provided. By precursors, we refer to any small, geochemically derived molecules that serve as the reagents to produce the building blocks of life, such as carboxylic acids, amino acids, nucleobases, lipids, and sugars. There are many gaps in our knowledge of Earth’s conditions at the time of life’s emergence. Consequently, the essential details concerning where and how life emerged remain a mystery, which includes the identity and sources of precursors.
Over the decades of origin-of-life (OoL) research, several hypotheses have been formulated and reframed to shape the research field: from primordial soup to the ribonucleic acid (RNA) World, information-first or genetics-first and metabolism-first, heterotrophic and autotrophic origins—just to name a few (Gilbert, 1986; Wächtershäuser, 1997; Anet, 2004; Schönheit et al., 2016; Gutekunst, 2018). Some of these hypotheses have overlapping components, and many have led to subhypotheses, each with their own chemical, geochemical, and biochemical variations.
In general, these interrelated hypotheses focus either on routes to a self-replicating information system (often in polymer form) from environmental building blocks or the emergence of self-replicating proto-metabolism from building blocks produced by an autocatalytic network. These hypotheses are typically based on, or at least connected to, a certain physicochemical locale, for example, warm little ponds or hydrothermal vents (Preiner et al., 2020a). The environmental settings for these hypotheses are, in turn, driven by a focus on certain precursor molecules (e.g., hydrogen cyanide [HCN], simple organic acids, and ammonia [NH3]). In other words, the precursors, reactions for obtaining them, and processes that transform them into more complex building blocks often define the hypothesis (Andreani et al., 2023; Saladino et al., 2012; Benner et al., 2012; Sousa and Martin, 2014; Rauscher and Moran, 2022). Furthermore, the types of chemistry available in the physicochemical locale (homogeneous catalysis, surface-enhanced catalysis, and condensation reactions without metals) are closely associated with their respective OoL scenario.
Given that early Earth’s environmental conditions are not well constrained, extant Earth environments such as vents, ponds, and hot springs often serve as guides. The fact that many hypotheses focus on only one environment has led to some of the misunderstandings between researchers who investigate different OoL hypotheses. Certain chemical characteristics are limited to specific environments, while others are more widespread. For example, hydrogen (H) may be abundant in both land-based and deep-ocean settings, and wet–dry cycling exists in hydrothermal vents, as well as on land (Andreani et al., 2023; Klein et al., 2020; Šimonis et al., 2026).
Considering the current knowledge gaps, it seems prudent to remain agnostic and avoid defining the environmental conditions of early Earth too stringently: for example, by focusing solely on alkaline hydrothermal vents and excluding vents with other fluid geochemistries/pH or imagining highly orchestrated environmental conditions characterized by a certain type of “working” laboratory prebiotic chemistry system. We aim to present a hypothesis-neutral review of the geochemical pathways leading to key precursor molecules and their chemical pathways into life’s building blocks. Simultaneously, we provide a nuanced overview of the two main scientific approaches to life’s emergence.
Here, we reframe the debate and propose that the core conceptual distinction between the various prebiotic scenarios is ultimately not based on environmental setting, but something else. Namely, it is a matter of whether life’s building blocks, such as amino acids and nucleotides, originated from prebiotic chemistries that worked differently than extant biochemistry (“unrestricted prebiotic chemistries,” hereafter referred to as unrestricted chemistries) or whether life’s building blocks originated from prebiotic chemistries that resembled extant biochemistry (“lifelike prebiotic chemistries,” hereafter referred to as lifelike chemistries). Whereas hypotheses focusing on unrestricted chemistries are not constrained by known biological synthesis pathways (Powner et al., 2010; Gibard et al., 2018; Poddar et al., 2024), those proposing lifelike chemistries assume that biochemistry today maintains mechanisms from its emergence and rely on a direct connection between the chemistry of life and prebiotic chemistry. How the various OoL hypotheses sort into these two categories is shown in Fig. 1A.
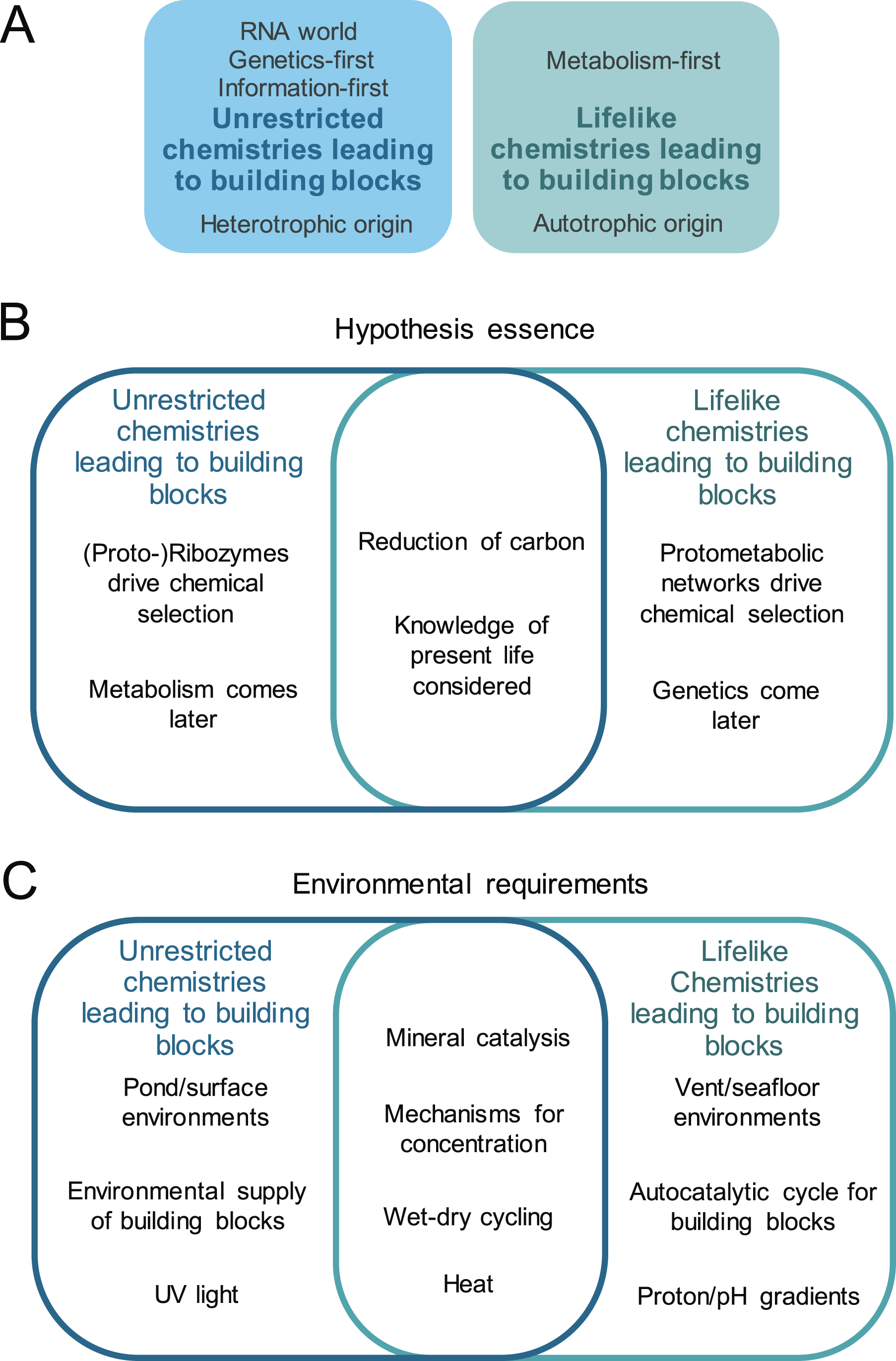
We reframe common origin-of-life hypotheses as “unrestricted” prebiotic chemistries and “lifelike” prebiotic chemistries that lead to the formation of essential building blocks for life.
There are many research angles to the emergence of life, although the reduction of carbon (C) is central to them all; no widely accepted OoL scenario operates exclusively on oxidized C. In fact, relative to Earth as a whole, biological matter is enriched in C, H, and nitrogen (N) while depleted in oxygen (O) and metals (McKay, 2004). From the RNA World to metabolism-first, from primordial soups and lightning to hydrothermal vents, these hypotheses all require reduced C as a key step to building block synthesis. Nevertheless, the most abundant C source on early Earth was carbon dioxide (CO2), C’s most oxidized form (Johansen et al., 2024; Zahnle et al., 2007).
No preserved geological proxies for the levels of Hadean atmospheric CO2 exist, but calculations have proposed that up to 100 bar of CO2 was present in Earth’s atmosphere (Sleep, 2010). It seems likely that CO2 reacts with the seafloor and is slowly subducted into Earth’s mantle over time (Zahnle, 2006). More recent modeling studies suggest Earth’s secondary atmosphere, which existed for 5–200 million years, was also dominated by CO2, H2, and N2. This is because of (1) early CO2 outgassing, (2) the water-gas shift reaction, which efficiently oxidizes carbon monoxide (CO) to CO2, (3) hydrogen gas (H2) depletion due to solar extreme ultraviolet (XUV) sputtering, and (iv) decreased H2 outgassing from impacts (Johansen et al., 2024; Pearce et al., 2022b).
How CO2 was reduced in these early atmospheres and into what C compound(s) are where things get interesting for emergence-of-life hypotheses. The most straightforward route is hydrogenation, that is, CO2 reacting with H2, which on an early Earth could have been produced by hydrothermal rock alteration, radiolysis, mechanoradicals, impact degassing, and volcanic degassing (Klein et al., 2020; Wogan et al., 2023). When CO2 and H2 react, the equilibrium lies on the side of reduced carbon compounds. Thus, one could expect them to react spontaneously to produce methane (CH4), water, and heat (Shock, 1990). But due to kinetic barriers (Shock, 1990; Maden, 2000), this reaction is extremely slow without catalysts such as mineral surfaces. The product distribution also depends on the pH, solvent (or no solvent), temperature, type and structure of the mineral catalyst, reaction time, and the ratio between CO2 and H2 (Peters et al., 2023; McCollom and Seewald, 2006; Beyazay et al., 2023a; Beyazay et al., 2023b; Preiner et al., 2020b; Prakash et al., 2022; Wogan et al., 2023; Zahnle et al., 2020). The reduction usually undergoes two-electron removal steps and can lead to products that include CO, formate, formaldehyde, methanol, acetate, ethanol, pyruvate, alkenes, and alkanes. Also, electron sources other than H2 are feasible and include iron sulfides (Huber and Wächtershäuser, 1998; Nan et al., 2024), pH gradients (Hudson et al., 2020), and native metals (Varma et al., 2018; Beyazay et al., 2023b). CO2 can also be transformed into organic compounds via photocatalysis. Examples of this include radical atmospheric chemistry to generate formaldehyde (Cleaves, 2008) and combinations of photolytic and mineral catalysis (Senanayake and Idriss, 2006). Independent of the initial reduction mechanism, it has been shown that certain reduced C compounds can react to form more complex molecules (with two or three C atoms or higher). This occurs either by reacting with other reduced C compounds (often but not exclusively via further reduction steps) or with molecules that bear elements other than C, H, or O, that is, nitrogen (N)-, sulfur (S)-, or phosphorus (P)-containing compounds. When it comes to surface-catalyzed reactions, Fischer–Tropsch-type reactions, which involve CO as the carbon source and H2 as the electron source, are thought to be mostly responsible for the formation of long-chained C compounds (Peters et al., 2023; Riedel et al., 2003). The Sabatier process (CH4 from CO and H2) is a well-known representative of these types of reactions (Li et al., 2019). Also, the formose reaction, which is the formation of sugars from formaldehyde (Breslow, 1959; Haas et al., 2020; Tran et al., 2023), should be listed here since formaldehyde (CH2O) is a simple one-C compound and an important intermediate in CO2 reduction. Oxidation reactions can follow the initial reduction to form new products and reaction partners (e.g., methane oxidation). The formation of the carbon precursor HCN depends on reducing conditions, that is, mainly starting from CH4 as a carbon source (Diefenbach et al., 1999).
What types of reduced C compounds and routes of catalysis are considered “plausible” within the emergence-of-life community depends mostly on how one answers the following question: Does the source of C—and the manner in which the source is transformed into precursors and building blocks—resemble the C sources and transformation mechanisms of the last universal common ancestor (LUCA)? This separation is most apparent when considering the classical OoL hypotheses, most notably metabolism-first versus genetics-first. The distinction proposed here (unrestricted vs. lifelike chemistries to building blocks), however, offers an inclusive approach toward life’s origins. Commonalities of these different approaches with respect to the essence of each hypothesis and known environmental requirements are shown in Fig. 1B, C.
When considering the hypotheses of life’s emergence via lifelike chemistries, one can consider them as a mechanism for harnessing (or dissipating) the free energy stored in the form of a redox gradient between CO2 in Earth’s atmosphere/hydrosphere and geochemically provided electrons (e.g., H2) in its mantle. Depending on the catalysts and environmental conditions, a complex series of reactions would have emerged in a stepwise fashion and ultimately led to the chemical reaction that is life (Martin, 2011). The core concept of the lifelike chemistry approaches is that reactions that lead to life are still imprinted in parts of life’s metabolic core and, by extension, would have also existed in LUCA.
Alternatively, when considering the hypotheses for life’s emergence via unrestricted chemistries, the connection between life as we know it and prebiotic reactions is not a direct link with known biochemistries, although the original source of carbon would have been the same. The argument that life has reinvented its chemistry many times in the past 4 billion years makes it difficult to deduce the chemistry that led to it by considering the phylogenetically reconstructed metabolism of LUCA.
Below, we contrast these two different prebiotic approaches to forming life’s building blocks with CO2 as the principal carbon source (Fig. 2).
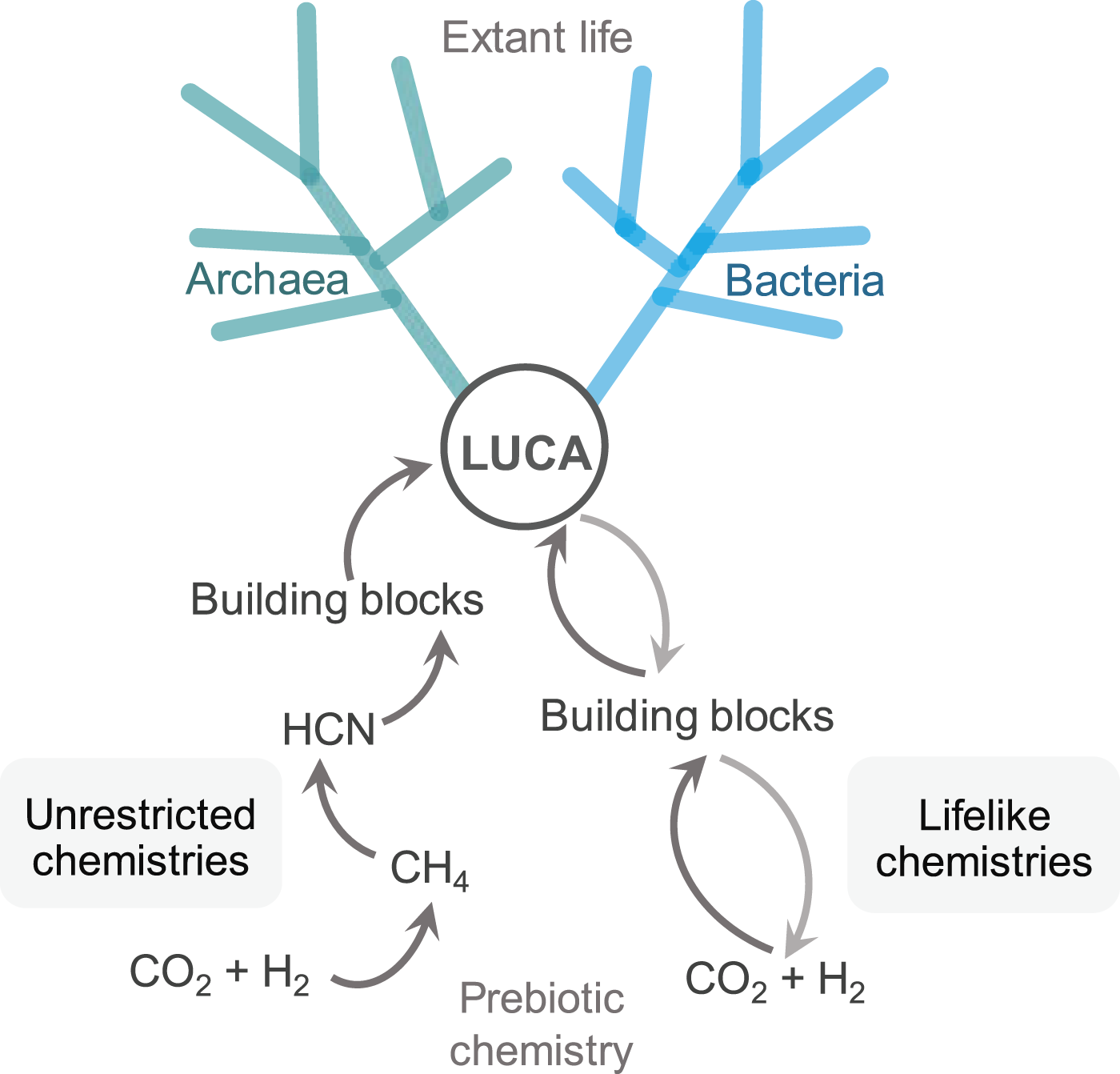
The two opposing prebiotic chemistry approaches—from precursors to building blocks to life. One establishes a path of least resistance between the chemistry of life’s emergence and extant biochemistry, while the other assumes the chemistry of extant life is not connected to the chemistry that led to life’s first building blocks. Partly adapted from Weiss et al. (2016).
The core idea of this approach is to forgo metabolism until later stages of the evolution of life as we know it. Instead, what if a simple life-form could emerge that is sustained by building blocks produced by the environment? This approach assumes that such a life-form would, at minimum, require some sort of genetic information that could be copied. This information is assumed to be a base-pairable polymer that stores sequence information (e.g., nucleic acids such as RNA, deoxyribonucleic acid (DNA), or a functional variant not used by extant life on Earth, “XNA”). Furthermore, many have proposed that the first evolving life-form needed to be encased somehow so that it was separated from the environment. Some options for individuation include fatty acid vesicles (Deamer, 2017; Milshteyn et al., 2018) and coacervates (i.e., phase separation in water; Aumiller et al., 2016; Poudyal et al., 2019).
The most popular version of this OoL approach is the RNA World hypothesis, which suggests that first life used RNA as both an information polymer and an enzyme for catalytic replication (Woese, 1967; Crick, 1968; Orgel, 1968; Gilbert, 1986; Dworkin et al., 2003). The logic behind an RNA world is that the DNA–protein world of today involves very complex mechanisms for replication and function and could not plausibly have emerged spontaneously. It is more reasonable to imagine the DNA–protein world being a product of chemical evolution that started from simpler, more error-prone machinery with less functionality. RNA is a great candidate for composing this simpler life-form, as it is an information polymer with a good capacity to fold onto itself and become catalytic (e.g., Lincoln and Joyce, 2009; Robertson and Joyce, 2014). Moreover, RNA still has a role in life today (e.g., ribosomal RNA, transfer RNA), which makes it easy to envisage its role in early life prior to the DNA–protein world. Researchers who work on the RNA World hypothesis try to experimentally demonstrate and understand pathways for the production of RNA building blocks in early Earth environments (Ponnamperuma et al., 1963; Tian et al., 2005; Ferus et al., 2017; Sebree et al., 2018; Zahnle et al., 2020; Wogan et al., 2023; Pearce et al., 2024) and mechanisms by which RNA can spontaneously become self-replicating (Mizuuchi, Furubayashi, and Ichihashi, 2022; Zorc and Roy, 2024).
In most current versions of this model, HCN is central to the production of RNA building blocks on early Earth. HCN is soluble in water and has a great tendency to react with itself and other small molecules, which makes it an excellent precursor to complex biomolecules (Matthews, 2004). A straightforward example of this is the reaction 5 HCN → adenine (Oro, 1961). Adenine is one of the four base-pairing components of RNA; the others are cytosine, guanine, and uracil. Some organic hazes derived from HCN polymerization contain these four nucleobases (Perrin et al., 2021). Other pathways to RNA building blocks that circumvent the need to produce a nucleobase first have been demonstrated and proposed; however, in all cases, HCN or other nitriles (e.g., cyanoacetylene [HCCCN]) are still involved (Powner et al., 2009; Ritson and Sutherland, 2013; Becker et al., 2019; Yi et al., 2020). If HCN is indeed the precursor to RNA, we must consider its source on early Earth.
HCN is a reduced form of C, which forms favorably in atmospheres that are rich in N2 and reduced C species, such as CH4. In contrast, atmospheres rich in oxidized C, such as CO2, do not favorably form HCN (Pearce et al., 2022b; Johansen et al., 2024). A simple explanation for this is that it is very hard to remove O atoms from CO2, because the C atom holds onto the O atoms with strong double bonds. Conversely, the C in CH4 holds onto H atoms with weaker single bonds, which makes it much easier to remove them; this enables the C to bond with a N atom released from N2 dissociation to form HCN.
Atmospheric models show that there can be a five-order-of-magnitude difference in HCN production in reducing (H2, N2, CH4, H2O) atmospheres compared with oxidizing (CO2, N2, H2O) atmospheres (Pearce et al., 2022b). Models and experiments suggest that HCN forms most abundantly via ultraviolet (UV) photochemistry in atmospheres with greater than a few percent CH4 (Zahnle et al., 2020; Pearce et al., 2022a; Wogan et al., 2023). However, CH4 was not a major species outgassed from volcanoes on early Earth (Trail et al., 2011), and it is highly reactive. This makes it difficult to imagine a scenario where it can increase in concentration in the atmosphere. One promising scenario requires a large (>400 km) asteroid impact that could deliver enough heat (>950 K) to produce thermodynamically significant amounts of CH4 from CO2 and H2 (Wogan et al., 2023). In such a setting, HCN could build up to 10−9 bar in the troposphere (Wogan et al., 2023), where it could rain out into warm little ponds and react to produce the building blocks of RNA Fig. 3.
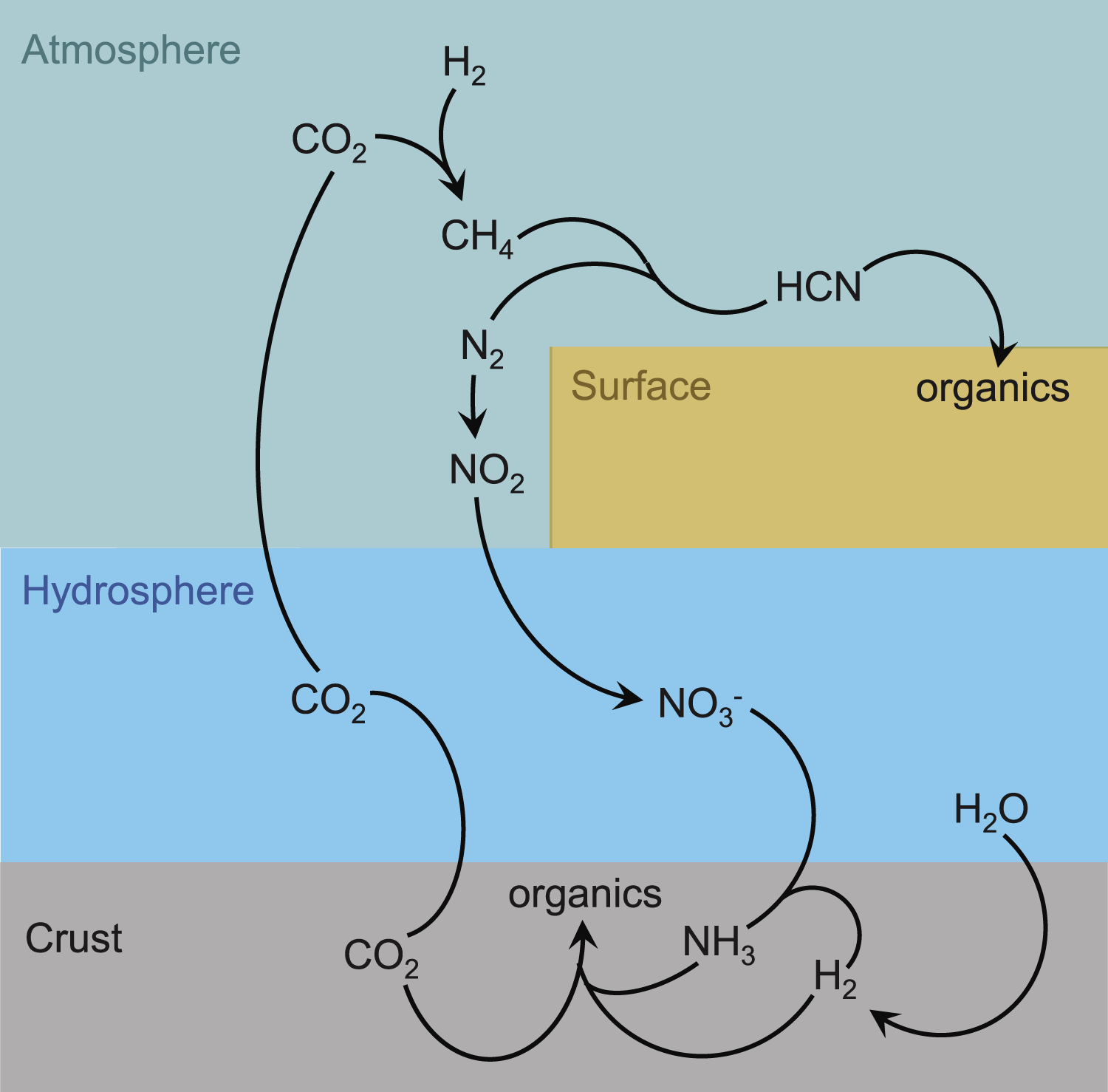
Schematic overview of possible paths of major endogenous carbon and nitrogen sources on an early Earth, starting from a CO2−, H2−, and N2−containing atmosphere.
HCN also polymerizes in the atmosphere to form solid organic haze particles, as observed on present-day Titan with its hazy, orange, opaque atmosphere. This type of atmosphere has also been proposed for early Earth—sometimes referred to as “the pale orange dot” model (Arney et al., 2016). Organic haze is another abundant source of the building blocks of RNA. Various experimental studies have measured nucleobases in simulated organic hazes, with abundances up to tens of parts per million (Pearce et al., 2024; Kawai et al., 2019; Sebree et al., 2018; Hörst et al., 2012). Organic haze particles readily fall out of the atmosphere and could have landed in warm little ponds, where their contents can react and complexify (Pearce et al., 2024).
Considering these two mechanisms for concentrating HCN, it is hard to envisage high local concentrations of RNA building blocks given the various molecular sinks that are present within ponds, such as hydrolysis, UV photodissociation, and seepage through pores. Models suggest that nucleobases produced by atmospherically sourced HCN or from organic haze particles can only reach 0.01–1 μM in ponds with a porous bottom or 1–100 μM in ponds without a porous bottom (Pearce et al., 2022b; Pearce et al., 2024). This is potentially problematic, given that the lowest experimental concentration of nucleobases that have reacted to form nucleosides (nucleobase + ribose) and nucleotides (nucleoside + phosphate) is ∼100 μM (Ponnamperuma et al., 1963). RNA building block production in the large-impact scenario has yet to be modeled; however, if the surface could remain below boiling temperatures in the presence of ∼1% CH4, this hypothesis might lead to more HCN and haze deposited in ponds, potentially providing higher nucleobase concentrations (Pearce et al., 2022a).
Another potential solution to the concentration problem is to build up large HCN stockpiles in salts that can later be released in high concentration when the pond conditions change (Toner and Catling, 2019; Hara and Templeton, 2024). The main candidate for HCN stockpiling is for it to react with iron to form ferrocyanide via the reaction Fe2+ + 6CN− ↔ Fe(CN)64−. At a high concentration, ferrocyanides precipitate from solution as a variety of insoluble and soluble salts (Williams, 1915). These ferrocyanides can later release free cyanide via UV radiation, thermal decomposition, or reactions with dissolved CO or hydrogen sulfide (Arrhenius et al., 1994; Keefe and Miller, 1996; Patel et al., 2015; Hara and Templeton, 2024). However, this hypothesis requires more modeling and experimental demonstration to be widely accepted. Additional mechanisms to slightly boost concentrations of HCN also exist within ponds, such as interactions with membrane vesicles (Chen and Walde, 2010; Himbert et al., 2016).
HCN is also found on a variety of meteorites and comets, which leads to the hypothesis that HCN also could have been delivered exogenously to early Earth (Smith et al., 2019; Todd and Öberg, 2020; Kurosawa et al., 2013). A continuous terrestrial production of crucial precursors, however, seems preferable since a sustained source of the building blocks of life would have been necessary until it was supplanted by a primitive metabolism (which in this model occurs late). Single influxes of HCN precursors and larger building blocks of life to local environments, such as via meteoritic delivery, may provide boosts to local building block concentrations, although they are not sustainable sources in the face of environmental sinks such as hydrolysis, photolysis, dilution, and seepage (Pearce et al., 2022b).
Additional key elements required for the emergence of life include S and P; however, of these two species, only P is necessary for the emergence of RNA, as it is present in the backbone of RNA as phosphate (PO43−) and thus must be available for nucleosides to be phosphorylated to produce an RNA monomer (i.e., a nucleotide). The main P-bearing mineral in Earth’s crust is apatite, though it is poorly soluble in water (Walton et al., 2021). A more soluble form of P potentially available on early Earth would be schreibersite, which is present in iron meteorites (Gull et al., 2015) and can be produced on Earth’s surface during cloud-to-ground lightning (Pasek et al., 2017) or hypervelocity impacts (Pasek, 2017). The other key element for present-day life on Earth is S, and it is present in two of the 20 proteinogenic amino acids (methionine and cysteine), as well as in some vitamins (e.g., biotin and thiamine) and coenzymes (e.g., coenzyme A, lipoic acid) required for metabolism. The outgassing of S from volcanoes would have been primarily in the form of sulfur dioxide (SO2), as zircon analyses suggest the mantle would have been oxidized as early as ∼4.35 billion years ago (Trail et al., 2011).
The constraint of depending on unrestricted chemistries en route to life inevitably introduces limitations that must be addressed and depend on processes that must be demonstrated in the laboratory. Some issues that currently lack clear solutions are (1) the lack of a high-yielding pathway to ribonucleotides, (2) demonstration that a stable self-replicating RNA system can exist, and (3) explanation of how a self-replicating information system evolved from sourcing building blocks from the environment to producing them metabolically. Indeed, although there are ideas as to how metabolism emerged following RNA-based chemistry (e.g., the retrograde hypothesis; Horowitz, 1945; Lazcano and Miller, 1999; Scossa and Fernie, 2020), this connection is still an area of great speculation.
The second approach to explaining the emergence of life is to identify a “path of least resistance” that connects the core biochemistry of LUCA’s metabolism to the geochemistry responsible for the emergence of the first living systems. This is challenging because our understanding of LUCA’s physiology (Weiss et al., 2016; Moody et al., 2024) is disconnected from the HCN-dependent prebiotic chemistry described in Section 3. The chemical gap between prebiotic chemistry and LUCA’s biochemistry has classically been considered a minor inconvenience to synthetic chemistry approaches to abiogenesis (e.g., Wu and Sutherland [2019] argue that evolution would close the gap between unrestricted life chemistries in the prebiotic world and LUCA, given enough time). In contrast, this chemical gap is seen as one of the main problems that concerns evolutionary biologists working on origins topics (Harrison et al., 2023; Mrnjavac et al., 2024). This brings forward the fundamental question: Are there nonenzymatic mechanisms analogous to those conserved in biological systems that can explain the production of reduced biomolecules on early Earth? In this section, we present examples of lifelike prebiotic chemistry giving rise to such biomolecules.
To date, we cannot close the knowledge gap between prebiotic chemistry and the biochemistry of LUCA. And a complete reinvention of metabolism over millions of years seems possible: there are many examples in Earth’s biological history of lineages that abandon an ancient trait that is detrimental for their fitness (Carscadden et al., 2023); this has often resulted in the presence of vestigial organs or genes. However, by definition, the universally conserved core of metabolism (Weiss et al., 2016; Wimmer et al., 2021) suggests that the intimate functioning of life’s intermediary C metabolism is ancient and too important to change. The lifelike approaches to abiogenesis propose that life arose due to geochemistry analogous to its metabolic core.
The metabolism of Earth’s modern biosphere is mainly based on the biochemistry of CO2, although some living organisms convert reduced C in the form of CH4 to CO2 (Farr et al., 2023; Nitschke and Russell, 2013). The lifelike approaches stem from a common origins hypothesis under hydrothermal conditions; however, CO2 fixation as a starting reaction seems currently more consistent with LUCA’s physiology (Moody et al., 2024). It makes sense to wonder whether prebiotic chemistry based only on the geochemistry of CO2 could have been responsible for life’s emergence.
Reviewing recent work that has explored lifelike prebiotic chemistry, we can see that CO2 and H2 have been demonstrated to spontaneously react to form intermediary carbon metabolites, that is, simple organic compounds central to metabolism such as formate, acetate, pyruvate, and Krebs cycle constituents (i.e., ketoacids). This has been demonstrated using iron and nickel alloys and minerals under crustal (partial) pressures (Varma et al., 2018; Preiner et al., 2020b; Rauscher and Moran, 2022), as well as under milder conditions using naturally occurring pH and redox gradients to bypass endergonic barriers (Hudson et al., 2020). These organic compounds and the required catalysts have widespread roles in cellular biochemistry, particularly in acetogens and methanogens (Russell and Martin, 2004; Braakman and Smith, 2012; Martin, 2020). Also, the lifelike prebiotic chemistry of amino acids is relatively straightforward and takes place by reductive amination (e.g., Barge et al., 2019; Maughan and Miller, 1999; Kaur et al., 2024a) or transamination (e.g., Nakajima et al., 1975) of ketoacids, just like in the analogous biological mechanisms.
The biologically analogous prebiotic chemistry of amphiphiles (necessary for the assembly of membranes) has often been related to that of organosulfur compounds, since that resembled the biological mechanism mediated by acetyl-CoA (McMurry and Begley, 2005), arguably the most important biological thioester. Prebiotic thioesters also could have played a role in energy metabolism similar to that which organophosphates played later on (Goldford et al., 2017). Prebiotic thioesters such as methyl thioacetate are readily formed under simulated alkaline hydrothermal conditions (Whicher et al., 2018; Kitadai et al., 2021). However, a prebiotic connection of fatty acid synthesis to the condensation of thioesters has not yet been demonstrated. Also, lifelike prebiotic chemistry of nucleotides and their constituent parts has only recently been experimentally explored in an extensive manner. For instance, the synthesis of sugars emulating the biological gluconeogenesis pathway was only demonstrated within this decade (Messner et al., 2017). One challenge has been the production of a specific set of sugar species, since a wide range of sugars are formed from formaldehyde in the (nonbiologically analogous) formose reaction (Breslow, 1959; Camprubi et al., 2022; Colón-Santos et al., 2019; Haas et al., 2020). On the lifelike nucleotide synthesis end (Harrison and Lane, 2018), recent work includes the synthesis of nucleotide biological precursors, such as phosphoribosyl pyrophosphate (Akouche et al., 2017) and orotate (Yi et al., 2022).
A key challenge in finding parallels between life’s biochemical mechanisms and the chemical mechanisms of its putative geochemical origins is showing how simple reactions that start from very simple chemical species (e.g., CO2 and H2) could undergo a stepwise increase in complexity. Theoretical work has shown that there potentially exist minimal, self-sustaining (autocatalytic) chemical networks connected to CO2 and H2, which means that a theoretical roadmap of increasing self-production (i.e., reproduction) exists (Xavier and Kauffman, 2022; Wimmer et al., 2021; Xavier et al., 2020)—a feat that could further promote increases in molecular and network complexity (Peng et al., 2022). However, so far experimental verification of this type of abiogenic autocatalysis is extremely challenging, although initial steps have been taken (Zimmermann et al., 2024; Yu et al., 2024). The challenges reflect two factors: (1) the often very large set of molecules that network autocatalysis experiments would require, which are analytically challenging to manage, and (2) the fact that the parameter space to explore is virtually infinite. But there are new movements in chemistry, among them the push for experimental chemputation, that aim to push the boundaries of experimental automatization to speed up the exploration of chemical space (e.g., Asche et al., 2021). In any case, a sturdier experimental backing is essential to connect life’s biochemical mechanisms to its abiotic roots.
Hypotheses that focus on lifelike prebiotic chemistries have been criticized (Wu and Sutherland, 2019), for instance, for focusing narrowly on (marine) hydrothermal settings for the OoL, like serpentinizing systems (water–rock interaction system in Earth’s oceanic crust), due to the abundance of H2 and dissolved CO2 (Lang et al., 2010; Konn et al., 2015). Such settings pose challenges to explaining the concentration and polymerization of monomers, such as amino acids and nucleotides. Can water-rich environments promote monomer polymerization when chemical theory posits that the reverse reaction, hydrolysis, should be favored (Ross and Deamer, 2016)? It turns out problems such as these are not as stringent as one might think; the sponge-like structure of some hydrothermal edifices favors the convection of hydrothermal fluids due to thermal gradients, which promotes the concentration of monomers in colder pockets, a phenomenon known as thermophoresis (Matreux et al., 2024). Other solutions for the “water problem” within hydrothermal settings have been proposed (Gagrani and Baum, 2025; Westall et al., 2018; Lee et al., 2024; do Nascimento Vieira et al., 2020; Andreani et al., 2023).
Furthermore, the source of fixed N in this scenario is oxidized atmospheric N species (NOx) from lightning (Wong et al., 2017), which are water soluble and can be reduced to NH3 in environments rich in H2 such as serpentinizing systems (Shang et al., 2023; Dörr et al., 2003; Gao et al., 2025; Summers and Chang, 1993). Even more than that, nitrate (NO3−), with the help of minerals, can be directly used for prebiotic amino acid synthesis (Chimiak et al., 2024). Also, nitrides (N3−) embedded in minerals were shown to incorporate N into organic compounds during CO2 fixation (Beyazay et al., 2023c). Environments where CO2, H2, and NH3 are colocalized with metal catalysts have great potential to synthesize biomolecules via reaction mechanisms similar to those used by life as we know it (Kaur et al., 2024b; Rauscher and Moran, 2022).
When it comes to the source of S, apart from the aforementioned volcanic outgassing, several mineral sources (e.g., iron sulfides such as pyrite or greigite) or hydrothermally reduced sources like hydrogen sulfide (H2S) can also be considered in prebiotic chemistry scenarios (Mojzsis, 2007). Phosphorus, the final element to complete CHNOPS, was for a long time considered almost impossible to retrieve from its “only significant source,” the mineral apatite (Schwartz, 2006). However, reducing conditions, such as those found in serpentinizing or other rock–water interaction systems, could have produced different active P species, such as phosphite, that enabled organo-P bond formation (Walton et al., 2023; Bryant et al., 2010; Gull et al., 2023). There is also geological evidence that hydrothermal fluids were rich in dissolved P (Rasmussen et al., 2021).
While the emergence of primary metabolism is inherent in lifelike chemistry OoL scenarios, explanations for the informational part of life remain challenging. When focusing on metabolism-first hypotheses, one needs to explain how a self-replicating chemical network can constitute a precursor of an extant heredity mechanism (RNA/DNA). How the composition (and thus heredity) of such a network could undergo adaptive change remains a difficult question to address experimentally. Could the exchange of energy and matter with its environment take precedence and dictate the composition of an emerging network replicator? If so, this replicator’s composition could not pass on the information from the previous generation, since it would simply follow the basic drivers of its environment (Vasas et al., 2010). In contrast, if the composition of the next generation is sufficiently biased by the composition of the previous one, then autocatalytic network replicators would, in principle, be able to evolve and become gradually more complex despite lacking formal genetic polymers (Vasas et al., 2010). Theoretical work that suggests these primitive hereditary mechanisms could have played an important role before the advent of genetic polymers has been prolific (e.g., Lancet et al., 2018; Katla et al., 2025; Markovitch and Krasnogor, 2018). Demonstrating the evolvability of such systems experimentally remains a crucial step to be undertaken to connect life’s intricate biochemical mechanisms with geochemistry.
Limitations and Opportunities of Early Building Blocks for Life
Is there a way to decide, based on current information, which of the two different ways to approach prebiotic chemistry, unrestricted life versus lifelike chemistries, is more likely? The answer is no. Each approach has its advantages and disadvantages. For now, the former lacks a comprehensive connection between primary metabolism and heredity, while the latter lacks convincing mechanisms for RNA molecules to later evolve a primary metabolism. And while individual parts of each hypothesis may be falsifiable (e.g., can molecule X be synthesized under Y conditions?), the hypotheses as a whole are malleable and can often be rescued by proposing new geochemical conditions. This makes it very hard to rule out either approach with absolute certainty. Furthermore, new catalysts and novel chemistries are being found on a regular basis, a recent example is in the work of Rout et al. (2025).
With all the different ways to generate biomolecules from simple precursors, one should remember that we ultimately need to explain how they can be actively replenished within a prebiotic system. Solving the problem of getting to a certain set of biomolecules—for example, from ketoacids to thioesters or nucleotides—does not explain how a self-sustaining and evolvable network of chemical reactions was generated. In order to self-sustain on both an informational and material level, biomolecules would eventually have to influence and promote their own synthesis (Ganti, 2003; Xavier et al., 2020; Peng et al., 2022). This has not been shown for any of the hypotheses presented here.
The three authors of this article each have their preferred hypotheses. These preferences were influenced by differences in scientific background and by advisors and other scientists in the field. Different experiences and insights, however, should not limit the potential for scientific exchange, and such exchanges need to be proactively established.
It is likely that the truth lies somewhere between the main approaches; for example, it could be that some molecules needed to be part of (proto)metabolism from its beginnings, while others could be supplied by the environment until life found a way to reproduce them independently. Unraveling life’s origins thus requires that these communities maintain open-mindedness and collaborate more closely in an attempt to converge toward a solution.
Authors’ Contributions
E.C.: Conceptualization, writing—original draft, and writing—review and editing. B.K.D.P. and M.P.: Conceptualization, writing—original draft, writing—review and editing, and visualization.
Footnotes
Acknowledgments
The authors want to thank the peer reviewers for their thorough comments, which significantly improved the contents of this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
E.C. thanks the National Aeronautics and Space Administration, the National Science Foundation, the U.S. Department of Agriculture, and the University of Texas System for funding. B.K.D.P. thanks the Heising-Simons Foundation and Purdue University for funding. M.P. thanks the Max Planck Society, the International Max Planck Research School “Principles of microbial life,” the European Research Council (101221521), and the Human Frontiers Science Program (RGEC29/2025) for funding. The authors want to thank the peer reviewers for their thorough comments, which significantly improved the contents of this article.
Associate Editor: David A. Baum