
Abstract
The ubiquitous iron oxides and oxyhydroxides on Mars represent intriguing targets in the search for evidence of past microbial life on the red planet. So far, no studies have systematically investigated microbial fossils in fluviolacustrine ferricrete composed entirely of nanophase goethite; thus, our understanding of biosignatures in such deposits is limited. Here, we report exceptionally well-preserved microfossils in laminated nanophase goethite from the Miocene McGraths Flat Lagerstätte, New South Wales, Australia. Using scanning electron microscopy and energy dispersive spectroscopy, we identified a diverse range of fossil morphotypes, including bacteria and fungi. Syngenetic forms embedded in the sedimentary matrix and post-depositional endolithic colonizers were recognized. Our findings establish nanophase goethite as an excellent preservation medium for microfossils, capable of preserving subcellular details across multiple generations of microbial communities, and highlight nanophase goethite as a promising target for future Mars exploration missions.
Introduction
The ubiquitous iron oxides and oxyhydroxides on Mars, responsible for its characteristic red hue, represent intriguing targets in the search for evidence of past microbial life (Schröder et al., 2016; Amils et al., 2024; Tabata et al., 2024). The detection of these iron oxides in Martian weathering profiles supports the notion of a warm and wet past, conditions that favor the emergence of life (Carter et al., 2015). The abundance of iron on the surface of Mars is due to its largely basaltic (8–10 wt% Fe) crust (McSween et al., 2009). Throughout Mars’ early history, particularly during the warm and wet Noachian (>3.7 Ga) and Hesperian (3–3.7 Ga) periods, and possibly during more recent episodic groundwater flows, this vast source of mafic silicate minerals has been extensively altered, leaching iron(II) into fluviolacustrine settings (Fassett and Head, 2005; Ehlmann et al., 2011; Michalski et al., 2013; Carter et al., 2015; Wordsworth et al., 2021). Given that Mars likely never developed extensive oceans (Di Achille and Hynek, 2010; Carr and Head, 2015), hydrological and weathering processes have been largely land-based, which has contributed to Fe-oxyhydroxide formation (Carter et al., 2015; Loche et al., 2024; Mandon et al., 2024). Consequently, Mars has developed iron-rich fluviolacustrine deposits, perhaps comparable to those that preserve microfossils on Earth.
On Earth, the formation of nanophase iron oxides and oxyhydroxides is linked to biogeochemical iron cycling, which involves microbial processes that either directly control nucleation and mineral growth (biologically controlled mineralization) or indirectly influence the local microenvironment (biologically induced mineralization) (Konhauser, 1998; Schröder et al., 2016; Liu et al., 2024). Moreover, biogeochemical iron cycling, in addition to abiotic oxidative processes, contributes to silicate rock weathering (Fisk et al., 2006) and the formation of iron-rich deposits (e.g., laterite and fluviolacustrine ferricrete) that are recognized as excellent media for the fossilization of microbial life (Levett et al., 2020c; McCurry et al., 2022). However, the existence of iron biogeochemical cycling on Mars remains speculative, with several abiotic iron oxidation pathways proposed (Nixon et al., 2012; Nie et al., 2017; Kikuchi and Shibuya, 2021; Tabata et al., 2024). Nevertheless, the widespread preservation of microfossils in iron-rich deposits on Earth suggests that similar biosignatures could also be preserved on Mars (Konhauser, 1998; McCurry et al., 2022; Amils et al., 2024).
The Miocene McGraths Flat Lagerstätte presents a unique opportunity to examine microfossils in iron-oxyhydroxide-rich (goethite) sedimentary rocks. The site is well known for its exceptional preservation of soft-tissue details from a wide variety of rainforest flora and fauna, such as the ultrastructure of setae from spiders and insects, neuronal tissues in spiders, nanocoatings on the wings and eyes of insects, melanosomes in fish and birds, and stomata in leaves (McCurry et al., 2022; Moulds et al., 2022; Cantrill et al., 2023; Djokic et al., 2023; McCurry et al., 2023; Richardson et al., 2023; Baranov et al., 2024; Frese et al., 2024; Djokic et al., 2025). In addition to macrofossils, the site preserves a diverse microfossil assemblage that includes pollen, spores, fungi, and bacterial structures. The fossils are preserved within a 30–50 cm thick unit of laminated nanophase goethite (hereafter, ferricrete) that was deposited in an iron-rich oxbow lake (McCurry et al., 2022; Djokic et al., 2025). Although organic material is absent, the high fidelity of preservation at McGraths Flat could aid in the identification of microbial structures in other iron-rich sedimentary deposits on Earth and potentially Mars.
To date, no studies have systematically documented microbial fossils from fluviolacustrine ferricrete composed entirely of nanophase goethite (McCurry et al., 2022; Djokic et al., 2025). Previous reports of microfossils from iron-rich deposits include weathering profiles (Levett et al., 2020c), gossans (Williams et al., 2015), iron- and sulfide-rich river deposits such as Río Tinto, Spain (Amils et al., 2024), bog iron deposits (Crerar et al., 1979), modern and ancient iron-bearing microbial tufa mats (Potter-McIntyre et al., 2014), Chocolate Pots hot springs, Yellowstone National Park, United States (Parenteau and Cady, 2010), and iron-rich marine deposits, such as the Gunflint Formation (Rasmussen et al., 2021). Thus, McGraths Flat provides the first opportunity to study microfossils in a goethite-rich sedimentary deposit that formed in an iron-rich, sulfide-poor lake environment (McCurry et al., 2022; Djokic et al., 2025).
Here, we report on a range of exceptionally well-preserved microbial fossils in sedimentary nanophase ferricrete. We used scanning electron microscopy–energy dispersive spectroscopy to assess their biogenicity and timing of formation (paragenesis). A diversity of microbial fossils, including bacteria and fungi, are preserved with remarkable fidelity, which demonstrates the capacity of nanophase goethite to capture a broad range of microbial structures. Our findings indicate that both syngenetic microbial forms, deposited during sediment deposition, and post-depositional colonizers (endoliths) are preserved. This case study supplements previous studies that highlight nanophase iron-rich sedimentary rocks as promising targets in the search for microfossils on Mars.
McGraths Flat taphonomy and paleoenvironment
McGraths Flat, a Miocene fossil site (c. 11–16 Ma) located near Gulgong, New South Wales, Australia, exhibits the exceptional soft-tissue preservation of a rainforest community. The fossils are preserved in laminated, nanophase iron-oxyhydroxide (goethite) suggested to have been deposited in an oxbow lake in a seasonally wet rainforest, with iron sourced from nearby basalts (McCurry et al., 2022; Djokic et al., 2025). Stratigraphic, petrographic, and geochemical analyses support this model, demonstrating that the iron derived from local Eocene basalts capping nearby hilltops (remnants of the regional Liverpool Range Basalt province) (Djokic et al., 2025). The iron was likely liberated during intense basalt weathering under warm and wet conditions that coincided with the Miocene Climatic Optimum (Djokic et al., 2025). Under such conditions, decomposing vegetation would have produced fulvic and humic acids that acidified groundwater and facilitated reductive dissolution of Fe(II) from the basaltic bedrock (Djokic et al., 2025). The iron-rich groundwater was then transported into the oxbow lake, where it precipitated to form very thin beds that coated and later fossilized a wide range of organisms. Trace element signatures (V, Cr, Co, and Ni) and rare-earth element patterns in the ferricrete closely match those of the local basalts, which further supports the proposed iron source and sink relationship (Djokic et al., 2025).
The depositional environment is thought to have developed as a stratified lake that supported fish and aquatic insects in oxic surface waters (Baranov et al., 2024; Djokic et al., 2025; McCurry et al., 2025), while iron-rich bottom waters were likely oxygen-limited based on the lack of scavenging and well-preserved articulated fossils and at least mildly acidic given that abundant iron precipitation generates abundant protons (Djokic et al., 2025). Acidic, oxygen-limited bottom waters would have favored biotic Fe oxidation that led to exceptional preservation in iron-oxyhydroxides (Djokic et al., 2025). Primary Fe(III)-oxyhydroxide precipitation likely occurred through both abiotic and biotic pathways, including aerobic and anaerobic microbial Fe(II) oxidation across the lake’s redoxcline. While there is possible evidence for microbial mat formation at McGraths Flat (Djokic et al., 2025), direct evidence of Fe-oxidizing microbial biosignatures has not yet been identified.
Iron isotope signatures (δ56Fe = −0.51 to +0.14‰) from McGraths Flat ferricrete indicate initial reductive Fe mobilization followed by complete Fe oxidation, consistent with Fe isotope data from modern fluviolacustrine systems (Djokic et al., 2025). The system was predominantly ferruginous (iron-rich and sulfur-poor), conditions likely controlled by the absence of carbonates, sulfates, and sulfides in the surrounding near-surface bedrock. These geological and geochemical conditions favored extensive Fe(III)-oxyhydroxide precipitation, lacking S or CO3 mineral formation, and resulted in the deposition of a 30–50 cm thick fossiliferous unit of laminated nanophase goethite (Djokic et al., 2025).
Precipitation of Fe-oxyhydroxides, rather than secondary oxidative replacement, such as pyrite weathering to goethite, is supported by a high degree of ultrastructural preservation, complete lack of pseudomorphs after other potential mineral precursors (e.g., cubic pyrite), and high Th/U ratios (Djokic et al., 2025). The widespread and rapid precipitation of nanophase Fe(III)-oxyhydroxides created ideal conditions for ultrastructural soft-tissue preservation of both macrofossils and microfossils (McCurry et al., 2022; Moulds et al., 2022; Cantrill et al., 2023; Djokic et al., 2023; McCurry et al., 2023; Richardson et al., 2023; Baranov et al., 2024; Frese et al., 2024; Djokic et al., 2025). Several preservation styles are documented to have occurred contemporaneously with iron deposition at McGraths Flat: External molds that formed around organisms and permineralization of intracellular spaces and/or microbial decay-mediated authigenic mineralization (Djokic et al., 2025).
Microbial biosignatures in nanophase iron
Nanophase iron-bearing minerals, including goethite (α-FeOOH), ferrihydrite (poorly crystalline FeOOH), lepidocrocite (γ-FeOOH), schwertmannite (Fe8O8(OH)6SO4), and hematite (Fe2O3), represent effective preservational media for microbial biosignatures on Earth and potentially Mars (Pierson and Parenteau, 2000; Fernández-Remolar et al., 2005; Amils et al., 2007; Fernández-Remolar et al., 2008b; Parenteau and Cady, 2010; Parenteau et al., 2014; Potter-McIntyre et al., 2014; Hays et al., 2017; Levett et al., 2020a). These iron mineral phases form through abiotic and microbial oxidation processes that trigger Fe oxidation and precipitation to preserve a range of morphological and carbonaceous microbial biosignatures across diverse settings. Below is a summary of non-marine settings where bacteria and microfossils are associated with nanophase iron minerals.
Weathering profiles (duricrusts)
Ferruginous duricrusts (canga) capping iron ore deposits in the Serra Sul, Carajás region, Brazil, preserve bacterial microfossils. The duricrusts are composed of hematite and goethite-cemented breccia ± Ti, Cr, and Al that formed during extensive laterite weathering under a wet, tropical climate (Levett et al., 2020c). Rod-shaped microbial fossils (1–5 µm long) are preserved as Fe and Al mineralized cell envelopes within goethitic matrix surrounding weathered breccia clasts (Levett et al., 2019, 2020b, 2020c). Iron-oxidizing microbes, including Sideroxydans, Azoarcus, and Gallionella species, and some iron-reducers (e.g., Geobacter species, <5% of the total sequences detected), have been identified from biofilms in iron-rich, oxygenated, circumneutral seeps that flow through canga duricrusts at the margins of lakes (Levett et al., 2020b).
Gossan
Gossans form through weathering of massive sulfide deposits, facilitated by microbial oxidation under acidic groundwaters, and they can contain both extant and fossilized microorganisms within the oxidized material. While gossans are predominantly composed of hematite, jarosite, and goethite, they almost exclusively preserve microbial fossils in iron-oxyhydroxides. For example, Fe-oxyhydroxides preserve nonbranching filamentous forms in the Brick Flat Gossan, Iron Mountain, California, United States (Williams et al., 2015); filamentous, coccoidal, and rod-shaped forms ± exopolymeric substances (EPSs) in gossan from the Lavrion District, Attica, Greece (Karaevangelou, 2016); and filamentous and coccoidal forms in late Miocene to Oligocene gossan from Río Tinto Basin, Spain (Fernández-Remolar and Knoll, 2008; Fernández-Remolar et al., 2021a). In the Río Tinto system, underground ferric oxides show clear microbial-like textures and organic–mineral associations, interpreted as reflecting active biogenic mediation during Fe oxidation and subsequent diagenetic replacement (Fernández-Remolar et al., 2021b; 2021c). Although the fossilized microorganisms in these gossans are not classified, it has been suggested that they represent fossilized chemolithotrophs, such as iron-oxidizing bacteria or fungi (e.g., Fernández-Remolar and Knoll, 2008; Williams et al., 2015; Karaevangelou, 2016; Fernández-Remolar et al., 2021a).
Río Tinto, Spain
The Río Tinto basin in southwestern Spain is an acidic, ferric iron- and sulfate-rich riverine system inhabited by diverse acidophilic prokaryotic and eukaryotic microorganisms (Amils et al., 2007). The sulfate- and iron-rich waters derive from the biooxidation of pyrite (FeS2) within the Iberian Pyrite Belt (Amils et al., 2007). These waters predominantly contain three acidophilic bacterial genera that oxidize and/or reduce iron (Leptospirillum, Acidiphilium, and Acidithiobacillus), along with a broad range of other bacteria, algae, fungi (yeast and filamentous), and protists (Fernández-Remolar et al., 2005; Amils et al., 2007).
The most effective preservation medium for microbial structures at Río Tinto occurs in nanophase Fe(III)-oxyhydroxides rather than sulfate minerals (Fernández-Remolar et al., 2005; 2008a). Precipitation processes linked to seasonal evaporation promote the formation of amorphous Fe-oxyhydroxides and schwertmannite that coat organic materials and microbial biofilms (Fernández-Remolar et al., 2005; 2008a; Amils et al., 2024). Modern to several million-year-old terrace deposits demonstrate the long-term stability of these iron-rich biosignatures (Fernández-Remolar et al., 2005; Fernández-Remolar and Knoll, 2008), including degraded organic compounds in ancient terrace horizons (Fernández-Remolar and Knoll, 2008). Early diagenetic processes transform amorphous Fe-oxyhydroxides into crystalline goethite while preserving original nanophase textures and morphological details (Fernández-Remolar and Knoll, 2008).
Iron mineralization encapsulates filamentous and coccoidal bacteria, algae filaments, fungal hyphae, plant tissues, and even insect cuticle (Fernández-Remolar et al., 2005). Such mineralization is often mediated by fungal and bacterial activity, which promotes Fe-oxyhydroxide precipitation (Amils et al., 2024). The iron-rich sediments frequently preserve vesicular textures interpreted as gas bubbles generated by heterotrophs (CO2), algae (O2), or methanogenic archaea (CH4) (Fernández-Remolar et al., 2005).
Acidic bog iron rivers from the New Jersey Coastal Plain
Bog iron deposits formed under acidic rivers and swamps in the southern New Jersey Coastal Plain, United States, contain irregular distributions of unconsolidated to massive limonite intermixed with sands and silts (Crerar et al., 1979). The only Fe mineral identified is goethite. Fe is mobilized from an Fe-rich substrate under acidic groundwaters and transported toward oxygenated, acidic surface waters. Fe-oxidizing bacteria, including Thiobacillus ferrooxidans, Leptothrix ochracea, Crenothrix polyspora, Siderocapsa geminata, and Metallogenium sp., mediate Fe mineral precipitation. Fossilized bacterial forms include sheaths (0.65 µm thick) that resemble Leptothrix and twisted filaments that resemble Metallogenium.
Iron-rich tufa mats
Iron-rich tufa mats (carbonate-rich microbial terrace deposits) formed in the past ∼100,000 years occur along alkaline, groundwater-fed cold springs at the Ten Mile Graben, southeastern Utah, United States (Potter-McIntyre et al., 2014). These CO2− and hydrocarbon-charged springs precipitate aragonite and iron-oxyhydroxides at circumneutral pH. Iron-oxyhydroxides precipitate despite low dissolved iron concentrations (<0.14 mmol/L), which suggests microbial rather than abiotic Fe oxidation (Potter-McIntyre et al., 2014). Modern mats preserve segmented stalks encrusted by amorphous Fe-oxyhydroxides and framboids interpreted as pseudomorphed pyrite. Stalk forms in older tufa exhibit higher degrees of diagenesis. The presence of unstable ferrihydrite in the ∼100,000-year-old tufa mats suggests it may serve as a potential biosignature, as organic polymers might be hindering recrystallization processes (Cornell and Schwertmann, 1996; Potter-McIntyre et al., 2014).
Iron-rich hot springs
Chocolate Pots hot springs, Yellowstone National Park, United States, is an anoxic, slightly acidic, bicarbonate-buffered, mid-temperature hot spring (37–54°C) with high dissolved Fe(II) concentrations (∼5–9 mg/L) (Pierson and Parenteau, 2000; Parenteau and Cady, 2010; Parenteau et al., 2014). In the subsurface, oxidation of volcanic sulfide gas drives sulfuric acid generation. These acidic solutions mix with alkali-chloride waters to produce an acid-sulfate-chloride spring, an intermediary to typical hot spring end members (e.g., acid-sulfate versus alkali-chloride springs) (Parenteau and Cady, 2010). In the anoxic vent waters, aqueous Fe(II) is oxidized by oxygen from oxygenic cyanobacteria, precipitating primary siliceous nanophase iron-oxyhydroxides that mineralize diverse phototrophic microbial communities (Parenteau and Cady, 2010). Extant and fossil cyanobacterial mats are preserved, including (1) millimeter-scale mats dominated by rods of Synechococcus, filaments of Chloroflexus and Rosieflexus (green nonsulfur filamentous anoxygenic phototrophs) growing at ∼50°C, (2) several millimeter-thick gelatinous mats and streamers that contain Pseudoanabaena filaments, (3) spring surface mats of Oscillatoria princeps filaments associated with metallic films, growing in ∼40°C quiet water pools, and (4) Oscillatoria that form dendritic structures, growing in high-energy outflow channels and terracettes (Pierson and Parenteau, 2000; Parenteau and Cady, 2010; Parenteau et al., 2014). Recent studies demonstrate lipid biomarker preservation in lithified microbial mat cores (Parenteau et al., 2014), showing that 85% of the original carbon content is destroyed by lithification (Parenteau et al., 2014). These investigations highlight how iron chelates organic compounds and inhibits cellular autolytic enzymes upon cell death, which prevents complete early organic decomposition and explains why iron is an effective preservation medium.
Methods
Sample collection
The samples analyzed in this study were collected during field sampling at McGraths Flat, conducted from 2017 to 2021. The collected material consisted of bulk rock samples that were subsequently split along their bedding planes using standard geological tools, including crack hammers and chisels. Samples were chosen based on the evenness of their surface to provide optimal conditions for high-resolution imaging and analysis. Fossils and rock samples are housed in the Palaeontology Collection of the Australian Museum in Sydney, New South Wales, Australia (accession prefix AM F).
Scanning electron microscopy–energy dispersive spectroscopy analyses
High-resolution scanning electron microscope (SEM) imaging was conducted using a FEI Quanta 650F variable-pressure field-emission SEM (Fig. 1). The instrument was operated at an accelerating voltage of 15 kV with working distances ranging from 12 to 15 mm. We acquired secondary electron (SE) images using an Everhart–Thornley detector to evaluate sample topography and surface morphology. For the coupled scanning electron microscopy–energy dispersive spectroscopy analyses (Fig. 2), we used a Hitachi S3400 SEM operated at 15 kV with working distances between 10 and 15 mm. Samples were prepared by desiccation in a vacuum oven at 100°C for a minimum of 12 h prior to analysis. We conducted energy dispersive spectroscopy on a range of representative specimens to determine their elemental composition, as not all microfossil types could be relocated for analyses (e.g., Fig. 1H–K). We used this data to establish a correlation between SE image contrast and chemical composition, enabling compositional interpretation of microfossils that could not be analyzed by energy dispersive spectroscopy. In SE images, iron-rich microstructures confirmed by energy dispersive spectroscopy consistently appear gray to bright gray, whereas carbon-rich material is dark gray to black (Figs. 1 and 2 cf. Supplementary Fig. S1). While contrast in SE images is typically attributed to topography or charging effects rather than true compositional differences (Goldstein et al., 2017), this systematic correspondence between SE imaging and energy dispersive spectroscopy data supports an iron-rich composition across all observed microfossils.

Microfossil preservation at McGraths Flat.
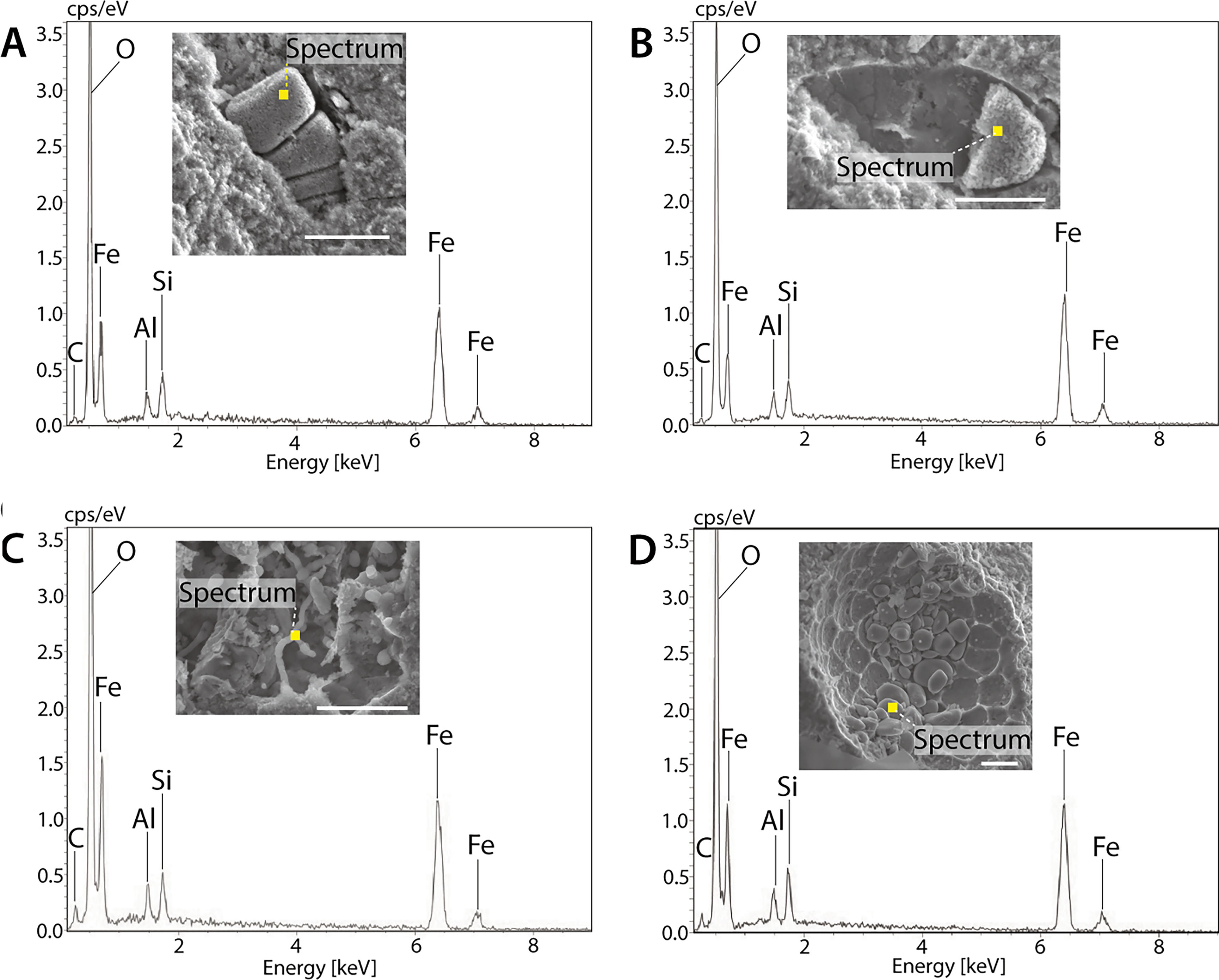
Scanning electron microscopy–energy dispersive spectroscopy analysis of microfossils from McGraths Flat. All microfossils consist largely of goethite (α-FeOOH) based on strong signals for Fe and O; much weaker signals were detected for Al and Si, which is likely due to signals from the matrix or an Al substitution for Fe in goethite (see also Djokic et al., 2025).
A diverse array of microfossil morphotypes with varied preservation context, referring to the position and relationship of potential microbial structures relative to the sedimentary matrix, have been identified at McGraths Flat. Seven morphotypes (I–VII) are described below:
Macro-filamentous structures with elongated, chain-like forms up to 10 µm wide and 100 µm long (Fig. 1A and Supplementary Fig. S2). These consist of individual cylindrical or barrel-shaped segments (approx. 5 × 5 µm) that form chains and branched structures composed of goethite. Where segments are missing, the external mold is visible. Rod-shaped microstructures, measuring 0.5–2 µm in length and 0.25–0.5 µm in diameter, with a 5:1 aspect ratio. These goethite microrods form internal molds (steinkerns) of relatively larger microfossils, including spores, pollen, and fungi (Fig. 1B) (see also Djokic et al., 2025). Radiating, shield-shaped goethite structures situated on the surface of a palynomorph (Fig. 1C). Yeasts, 5–10 µm in diameter, that display budding scars. Some yeasts are completely composed of goethite, whereas others show partial mineralization (Fig. 1D) (see also McCurry et al., 2022: fig. 3C). Nanofilamentous structures 0.5–1 µm thick and up to 5 µm long that occupy spherical cavities (Fig. 1E). Branching microfilaments approx. 1 µm thick and up to 100 µm long (Fig. 1F). These occupy irregular-shaped cavities. In some cases, the filaments protrude from smooth films to honeycomb-like matrices that coat the edge of cavities (Figs. 1G and 2C; Supplementary Fig. S3). Nanobeaded chains in linear arrangements that are up to 10 µm long and 1 µm wide (Fig. 1H–K). These Fe-rich structures occupy cavity space in larger microfossil molds. They are also associated with a network of 1 µm wide track marks (Fig. 1H–K).
Microfossil morphotypes at McGraths Flat fall into three general preservation categories:
Category one (morphotypes I and II) is characterized by external molds, sometimes containing internal molds, that are embedded within the laminated, nanocrystalline goethite matrix. Some specimens only preserve external molds due to the displacement of internal molds from their original position (Figs. 1A and 2A, B). Internal molds are composed of either homogeneous goethite infill (Fig. 1A) or consist of rod-shaped goethite microstructures (Fig. 1B). Rod-shaped goethite microstructures partially to completely fill internal molds in several different microfossil taxa (Figs. 1B and 2B) (see also Djokic et al., 2025). Category two (morphotypes III, V, VI, and VII) are superficial structures preserved above the surface of the rock. These lack external molds and do not form three-dimensional templates of larger microfossils. They typically occupy interstitial spaces such as pores and fractures or cavities produced from the external molds of larger microfossils up to 30 µm in diameter. Category three (morphotype IV) are yeast and yeast-like fossils that share characteristics from categories one and two. They are composed of goethite and exhibit either partial mineralization by nanoscopic goethite grains or complete homogeneous goethite mineralization similar to category one (Figs. 1D and 2D); however, they are superficial structures that lack fossil molds like category two.
Energy dispersive spectroscopy analysis of representative microfossils revealed their composition to be primarily iron (Fe) and oxygen (O), indicative of goethite; weak signals for aluminum (Al) and silicon (Si) likely stem from the surrounding matrix or possibly substitution for Fe by Al in goethite (see also Djokic et al., 2025). No organic carbon has been detected in the microfossils to date (Fig. 2 cf. Supplementary Fig. S1).
The exceptional micro- to nanoscale preservation at McGraths Flat enables detailed morphological analysis of its microfossil assemblage (Figs. 1 and 2), which highlights the capacity of nanophase goethite to capture and retain small and delicate structures. Below, we evaluate the biogenic origins (biogenicity) and timing of formation (paragenesis) of microfossils at McGraths Flat and discuss why nanophase goethite represents a promising target in the search for fossilized life on Mars.
Biogenicity and paragenesis of McGraths Flat microfossils
Detailed morphological preservation and context provide initial assessments of biogenicity and paragenesis (Westall, 1999; Tomescu et al., 2016); however, interpreting the origins of microfossils requires careful consideration, as abiotic processes can mimic simple microbial morphologies (García Ruiz et al., 2017; Muscente et al., 2018). In the following sections, we used multiple complementary characteristics to differentiate biogenic structures from abiogenic minerals (e.g., Fig. 1 cf. Supplementary Fig. S5). These include microscopic dimensions within the range of known microorganisms (e.g., bacteria are typically 0.5–10 µm in size); structures indicative of soft organic bodies (signs of plastic deformation, such as curled, coiled, and rounded structures); reproductive structures such as budding scars; preservation of fossilized biofilm components, that is, EPSs; and signs of behavioral processes, such as boring and/or etching patterns that cross-cut sedimentary laminae and are indicative of endolithic bacteria (Schopf, 1983; Konhauser, 1998; Westall, 1999; Westall et al., 2000; Cady et al., 2003; McLoughlin et al., 2007; Westall et al., 2015; Tomescu et al., 2016; Levett et al., 2019).
While geochemical biosignatures, such as carbon or specific trace elements (e.g., Gangidine, 2020), have not been detected in McGraths Flat microfossils, the exceptional preservation makes morphological assessment particularly valuable. The lack of any detectable carbon at McGraths Flat is likely due to post-depositional degradation of organic matter. This process is supported by observations in other iron-rich environments where very early diagenesis destroys most organics (85%), despite iron’s high potential as a preservation medium in the initial stages of fossilization (Parenteau et al., 2014). Preservation of the internal fossil molds at McGraths Flat suggests that degradation-resistant structures, such as sporopollenin in cell walls of pollen or chitin in cell walls of fungi, likely survived initial fossilization and degraded during a later stage of diagenesis (Figs. 1A, B and 2A, B) (Djokic et al., 2025). The relatively young age (c. 11–16 Ma) and exceptional preservation of McGraths Flat make the morphological features preserved more reliable indicators of biogenicity than in older, more altered deposits.
It is also important to consider when the McGraths Flat microbial organisms formed, such as during initial deposition or post-depositionally. For example, in the 1.8 Ga Gunflint Formation, microfossils are found embedded within amorphous silica, which suggests they were incorporated into the sediment prior to lithification (Rasmussen et al., 2021). In contrast, endolithic bacteria, that is, those that live within rock, display superficial and cross-cutting relationships with the substrate, such as inhabiting cavities and pore spaces (cryptoendolith), or fractures (chasmendoliths), or boring/etching the rock (euendoliths) for metabolic gain (Friedmann and Weed, 1987; McLoughlin et al., 2007). For paragenesis, we assessed whether the structures interpreted as microfossils formed (1) during original sediment deposition (syngenetic), (2) sometime after deposition as microbial endoliths (post-depositional), or (3) from artifacts associated with sample collection and analyses (contamination). The following sections discuss these factors for each morphotype, grouped by their interpreted timing of formation.
Syngenetic microfossils formed during the deposition of McGraths Flat
Macro-filamentous structures, nanorods, and radiating shield-shaped structures exhibit exceptional three-dimensional preservation with microscopic detail (Fig. 1A–C), which provides insight into both their biological origin and syngenetic formation.
Most macro-filamentous structures are interpreted as cyanobacteria that were original inhabitants of the McGraths Flat paleolake. These structures exhibit segmented and branching morphology and are of a size that matches that of cyanobacteria (blue-green algae), a group of photosynthetic microorganisms commonly found in freshwater lake environments (Dvořák et al., 2017). Evidence for their syngenetic formation (buried during initial sedimentation) includes the preservation of external mold impressions in the goethite matrix (Figs. 1A and 2A). The gap between the structures and the sedimentary matrix likely represents the original cell wall that no longer exists (see also Djokic et al., 2025). In extant cyanobacteria, cell wall thickness can range from 10 to 700 nm (Hoiczyk and Hansel, 2000), which is consistent with the dimensions observed at McGraths Flat.
The rod-shaped microstructures likely represent fossilized rod-shaped bacteria that colonized organic remains in the lake sediments. Their bacterial origin is supported by several features, including their uniform nanoscopic size, rod shape with rounded extremities, and plastic deformation that contrasts with abiotic minerals. The abundance, hundreds to thousands, of these rods that compose the internal molds of a variety of microfossil taxa supports microbial colonization (Figs. 1B and 2B) (Djokic et al., 2025). Colonization by the bacteria and subsequent mineralization by iron-oxyhydroxide, possibly mediated by the bacteria themselves, is proposed to have formed the internal molds (steinkerns) of the original soft tissue in various palynofossils (Djokic et al., 2025). At McGraths Flat, the spores’ outer sporopollenin walls are now largely absent, exposing partial to complete internal molds in several different specimens (Figs. 1B and 2B) (Djokic et al., 2025). The reason for the post-depositional loss of these typically resistant outer wall structures is unknown, though it could be the result of microbial degradation (Elsik, 1971; Holcová et al., 2020) (cf. discussion on nanobead chain-like structures).
Radiating, shield-shaped structures are interpreted as fungal germlings that grew on, and possibly degraded, the palynomorph during deposition of the original sediment (Fig. 1C). The structures’ size (8 µm in diameter) and interwoven filaments (hyphae) resemble fruiting bodies (ascomata) of epiphyllous fungi that are typically 8–40 µm in diameter (Conran et al., 2016; see Fig. 2M–T, Lange, 1969; Bianchinotti et al., 2020). Similar lobate ichnofossils (e.g., Fascichnus sp.) have previously been associated with borings produced by the cyanobacterium Hyella stella in a gastropod shell (Glaub et al., 2007); however, H. stella is significantly larger (measuring tens of microns) than the germlings described here. The astrobiological implications of distinguishing between fungal and bacterial fossils are significant. On Earth, fungi did not evolve until approximately 1.5 Ga (Loron et al., 2019). By this time, Mars is thought to have become dry, cold, and largely inhospitable to life (Davila and Schulze-Makuch, 2016). Thus, any fungi discovered on Mars, even if fossilized, probably represent terrestrial contamination (Eigenbrode et al., 2021; Simões et al., 2023). The association with a syndepositional palynomorph, combined with epiphyllous fungi’s preference for warm, humid conditions, aligns with the inferred early-mid Miocene paleoenvironment of McGraths Flat. At this time, mesic rainforests flourished west of the Great Dividing Range, before aridification of the Australian interior (Lange, 1969; Conran et al., 2016; McCurry et al., 2022). The epiphyllous fungi colonized the palynomorph before it was transported into the McGraths Flat oxbow lake, where it became buried and fossilized.
Endolithic microfossils
Branching microfilaments and nanobead chains exhibit morphological characteristics and preservation context that support an endolithic origin (Fig. 1F–K). Branching microfilaments preserved at McGraths Flat match the size and morphology of endolithic fungal hyphae (Figs. 1F, G and 2C) (Moore et al., 2020). While the observed structures share similarities with chytrid rhizoids, the notable absence of an apophysis (a swelling from which rhizoids typically emerge) argues against this taxonomic interpretation (Laundon et al., 2020). Some microfilaments are enveloped by what appears to be fossilized microbial EPSs (Figs. 1G and 2C; Supplementary Fig. S3). EPS is an extracellular substance that can contain a range of biopolymers, such as cellulose, and is known to engulf the secreting microorganism (Westall et al., 2000; Flemming and Wingender, 2010). In one instance, putative fossilized EPS appears to coat the margins of an irregularly shaped void space (Fig. 1G), which we interpret as evidence for a bioetching front. This interpretation is based on the spatial association of fungal material with putative EPS that coats an irregularly shaped cavity space. Li et al. (2021) showed that fungal bioetching occurs through the excretion of organic (oxalic) acid and biomechanical forces and documented an association of EPS with fungal hyphae. Endolithic fungi are known to secrete organic acid to dissolve rock substrates for nutrient acquisition (Sterflinger, 2000), while producing abundant EPS to regulate water loss and mitigate cell wall damage caused by changing temperature and moisture levels in subsurface environments (Omelon, 2016). The association of putative fungal hyphae with EPS that coats the margins of irregular-shaped cavities, combined with the fungi’s ability for rock dissolution, suggests that the branching microfilaments represent post-depositional endolithic fungi that occupied McGraths Flat ferricrete.
The nanobead chain-like structures at McGraths Flat consist of uniform, nanoscopic coccoidal units arranged in chains (Fig. 1H–K), consistent with the form and known dimensions of bacteria such as streptococci (Patterson, 1996). Each coccoidal unit has a deflated appearance, which suggests dehydration of the cytoplasm (Westall, 1999). Their Fe-rich composition further suggests that they are fossilized (Figs. 1H–K and 2; cf. Supplementary Fig. S1; see also Methods section). Bacteria that resemble streptococci have previously been reported from diverse settings, including pollen (Ambika Manirajan et al., 2016), lake sediments (Sherer et al., 1992), and lithified rock (Westall and Folk, 2003; Mezzasoma et al., 2022; Hidalgo-Arias et al., 2023). Thus, the bacteria at McGraths Flat could have been transported into the lake via organic debris, such as pollen grains, inhabited the anaerobic lake sediments during deposition, and/or colonized the sediments after lithification. Multiple lines of evidence suggest an endolithic origin. First, the microfossil cell diameters match the widths of nearby trails that cross-cut and connect to fractures within the sedimentary matrix (Fig. 1H–K); such features are typical of euendolithic microborings (McLoughlin et al., 2007). While these trails could represent degradation patterns similar to those observed in microbially altered pollen and chitinous microfossils (Elsik, 1971; Holcová et al., 2020), their position on the rock substrate rather than on remnant cell walls (notably absent) supports post-depositional euendolithic colonization. Alternatively, they could represent ancestral remnants of organisms that entered the lake prior to lithification, or cryptoendoliths that colonized existing cavities after the original euendolithic boring inhabitants departed. Nevertheless, their repeated association with the microborings in two separate locations and size (width) correlation suggests that these fossilized bacteria created the structures. Similar coccoidal forms have been documented in other iron-rich endolithic settings worldwide, for example, in magnetite (Fe3O4) grains from the Archean Isua Greenstone Belt (Westall and Folk, 2003) and within volcanic rocks from Antarctica (Hidalgo-Arias et al., 2023). The McGraths Flat microfossils are interpreted here to represent endoliths in fluviolacustrine ferricrete. This deposit would have provided Fe and perhaps other metals to support the metabolism of such colonizers (McLoughlin et al., 2007; Djokic et al., 2025).
Although endolithic bacteria are well-documented in modern land-based environments (Friedmann and Weed, 1987; Glaub et al., 2007; Walker and Pace, 2007; Hidalgo-Arias et al., 2023), their fossil record is sparse. So far, studies of endolithic fossils have primarily focused on oceanic igneous rocks, with no reports from iron-rich fluviolacustrine sedimentary deposits (Friedmann and Weed, 1987; Ivarsson et al., 2021, and references therein; McLoughlin et al., 2011). Thus, our findings expand the endolithic fossil record.
Syngenetic or endolithic microfossils?
Among the diverse microfossil assemblage preserved at McGraths Flat, nanofilaments and yeasts or yeast-like fossils could have formed either during deposition or post-depositionally as endoliths (Figs. 1D and 2D). The nanofilamentous goethite structures exhibit characteristics indicative of fossilized bacterial filaments, including submicron size, and plastic deformation indicative of soft organic bodies (Tomescu et al., 2016; Oh et al., 2023). Their position within spherical cavities of varying size and spatially associated with cyanobacterial filaments (Fig. 1A, E and Supplementary Fig. S2) provides some ecological context. It suggests that these cavities could represent fossilized molds of algal-derived lipid droplets (Peramuna and Summers, 2014). Alternatively, they could be the remnants of oxygen bubbles produced through photosynthetic activity (Sjöberg et al., 2020). Lipid droplets and oxygen bubbles commonly occur within microbial mats (Peramuna and Summers, 2014; Sjöberg et al., 2020). In the case of lipid droplets, the contents could have served as an energy source for microbial communities. Likewise, oxygen bubbles could have generated sharp redox gradients in the surrounding anoxic sediment that delivered reduced ions, such as Fe(II) or Mn(II), and reducing agents, such as organic matter, via percolating fluids (e.g., Brune et al., 2000; Borer et al., 2018). Such niche interfaces are common throughout sedimentary environments and could create ideal conditions for microbes to generate energy (Brune et al., 2000). Cyanobacterial mats likely developed in the photic zone near the lake surface, and their EPS could have provided structural integrity to maintain bubble stability (Sjöberg et al., 2020). EPS is indicated in the preservation of bubbles throughout the geological record (Djokic et al., 2017). Thus, these bubbles may have remained stable long enough in submerged mats to be exploited by bottom-dwelling microbes, despite the oxygen-limited bottom waters inferred at McGraths Flat paleolake (Djokic et al., 2025). Alternatively, the position of the nanofilaments on rock surfaces within void spaces suggests an endolithic origin.
Yeasts previously documented as syngenetic, indigenous microfossils at McGraths Flat display uniform sizes and diagnostic reproductive structures, such as attachment scars and buds, consistent with modern yeast (Tofalo and Suzzi, 2016; McCurry et al., 2022). While some yeast-like structures lack clear budding scars (Fig. 2D), all yeasts and yeast-like structures (e.g., Fig. 1D and Supplementary Fig. S4) contrast with inorganic goethite precipitates that exhibit variable sizes, botryoidal or spicular textures, and growth inward from the edge of cavities (Supplementary Figs. S5 and S6) (Dhaiveegan et al., 2016: fig. 6C, Westall, 1999). However, determining whether McGraths Flat’s yeast and yeast-like structures represent syngenetic fossils or later colonizers is challenging, as yeasts are known to inhabit both lake sediment (Brandao et al., 2010) and endolithic environments (Gorbushina and Krumbein, 2000). The distribution of the yeast colonies, found predominantly in void spaces that include pores, fissures, and fossil molds, suggests an endolithic origin. While microbial fermentation and subsequent CO2 production could explain the presence of yeasts in spherical cavities (Fig. 1D), it would not explain the yeast-like structures in elongate fissures (Supplementary Fig. S4). The yeast-like structures within such fissures, accompanied by palynomorphs, are likely the result of post-depositional washdown into the rock substrate (Riding, 2021). The palynomorphs within the fissures differ morphologically from syngenetic palynomorphs (cf. Figs. 1B and 2B) (see also McCurry et al., 2022; Djokic et al., 2023), further supporting a washdown origin.
Implications for the search for life on Mars
Exceptionally well-preserved microfossils at McGraths Flat advance our understanding of iron-rich terrestrial deposits that preserve microbial biosignatures. Using scanning electron microscopy–energy dispersive spectroscopy, we show that nanophase goethite can mineral-template a range of microbial biopolymers, including those found in bacteria (peptidoglycan) and fungi (chitin) at McGraths Flat. Goethite appears to have also fossilized associated microbial products, such as EPS (Fig. 1G). The preservation quality via goethite rivals that of other high-fidelity preservation media, such as silica, phosphate, and amber (Poinar, 1992; Dornbos, 2011; Ruff and Farmer, 2016). These observations further highlight goethite’s capacity to preserve a variety of microbial biopolymer structures, consistent with the understanding that organic materials provide negatively charged surfaces for metal ion binding (Leo and Barghoorn, 1976; Konhauser, 1998; Mustoe, 2017). Goethite preserves multiple generations of microbial communities at McGraths Flat, from the original lake inhabitants to post-depositional endoliths, though discriminating between these populations presents challenges. In the search for life on Mars, discriminating between syngenetic and endolithic generations is important (Oh et al., 2023) to accurately assess the age of potential Martian microfossils. Moreover, the harsh Martian environment, characterized by subzero temperatures, extreme aridity, and intense ultraviolet radiation, has led researchers to suggest that any microbial signatures would be predominantly endolithic, having taken refuge within the protective mineral matrix of rocks (Davila and Schulze-Makuch, 2016).
Methodological considerations
The study of microfossils at McGraths Flat highlights several important methodological considerations for future Mars exploration in iron-rich rocks. Scanning electron microscopy has proven invaluable to investigations of iron-rich terrestrial fossil sites (this study; Djokic et al., 2023; Wierzchos and Ascaso, 2001); this suggests that Martian missions should consider adopting similar high-resolution imaging techniques. Although SEM instruments have been designed for Mars rovers, they have not yet been deployed (Thaisen et al., 2010; Shukla and Ray, 2017). This may be in part due to the view that thin section petrography is the only reliable method of determining syngeneity and indigeneity (Walker and Pace, 2007), which currently requires sample return to Earth, as is planned (Muirhead et al., 2020). While petrographic analysis can help determine whether microfossils are embedded in the rock matrix rather than fractures (Walker and Pace, 2007; Tomescu et al., 2016), this method does not provide a three-dimensional view of putative microbial morphology and the depositional context. Fossilized endoliths could still be mistaken for syngenetic microfossils if the enclosing fracture/cavity goes unidentified. Fossilized endoliths may lack detectable carbon and mimic the chemistry of their host rock due to fossilization via mineral coatings sourced from a dissolution of the surrounding substrate (Konhauser, 1998; Wierzchos et al., 2005). Moreover, the opaque iron-rich mineralogy can conceal iron-mineralized microfossils under transmitted light microscopy. While reflected light readily allows observation of some concealed features such as mineral clasts (Djokic et al., 2025), microfossil analysis under these analytical conditions for iron-rich rocks is challenging. Therefore, complementary analytical techniques such as scanning electron microscopy may be useful for investigating microfossils in iron-rich deposits.
Formation and stability of goethite as a preservation medium
While McGraths Flat formed under rainforest conditions that have not existed on Mars, research on the site advances our understanding of nanoscopic Fe-oxyhydroxide as an ideal medium for preserving small-scale (subcellular) biological structures. McGraths Flat represents an iron-rich, sulfide-poor lacustrine setting, a preservational context that contrasts with other terrestrial fossil sites where microfossils are preserved in nanophase iron-oxyhydroxide, including the highly acidic (acid mine drainage), iron- and sulfur-rich Río Tinto (river) in Spain (Fernández-Remolar et al., 2005; Fernández-Remolar and Knoll, 2008), riverine bog iron (Crerar et al., 1979), laterite weathering profiles (Levett et al., 2020a), or acid-sulfate-chloride Chocolate Pots hot springs of Yellowstone National Park, United States (Parenteau and Cady, 2010). Collectively, these examples demonstrate how nanophase iron (goethite) forms and preserves microscopic life across a variety of depositional settings. Rather than being tied to specific conditions, preservation in nanophase iron-oxyhydroxide occurs across a broad spectrum of temperature, pH, hydrology, bedrock, and geomorphological regimes on Earth, which suggests similar preservational potential in nanophase iron-oxyhydroxide on Mars.
On Earth, goethite typically forms either through Fe(II) reacting with oxygen at the surface or via microbial oxidation of Fe(II), as part of a complex biogeochemical cycling of iron (Kappler et al., 2021). Initially, unstable polymorphs of nanoparticulate Fe(III)-oxyhydroxides, such as ferrihydrite or lepidocrocite, co-precipitate or chelate with organics in streams, creeks, lakes, and weathering profiles (Parenteau et al., 2014). These unstable polymorphs transform within a few days or years to stable goethite (Herbert, 1995; Konhauser, 1998; Fernández-Remolar et al., 2005; Fernández-Remolar and Knoll, 2008; Broughton, 2021). Even through this recrystallization, the original nanoparticulate size and thus fine-scale details of fossilized organisms are preserved, resulting in stable, low solubility mineral entombment (Fernández-Remolar et al., 2005; Fernández-Remolar and Knoll, 2008).
Goethite is stable under a wide range of conditions, dehydrating to hematite (Fe2O3) only at temperatures above approximately 250°C (Cornell and Schwertmann, 2003). This stability contrasts with other minerals that commonly act as preserving media, such as silica, carbonate, and phosphate. Opaline silica, despite its capacity for rapid preservation of fine-scale details in environments like hot springs (Ruff and Farmer, 2016), is prone to recrystallization to quartz even at low temperatures given enough time (Hesse, 1989). This process alters morphological information and destroys organics (Teece et al., 2020). Similarly, aragonite that forms skeletal and stromatolitic structures readily converts to calcite in the presence of water (Pamela Reid et al., 2003; Janiszewska et al., 2018), which causes the destruction of any fine-scale details (Janiszewska et al., 2018). Phosphatization can preserve extraordinarily fine-scale detail but displays a bias toward tissues that resist decay and are naturally rich in phosphate, such as bone (Dornbos, 2011; Schiffbauer et al., 2014). Phosphatization typically occurs either at the sediment–water interface or around decaying tissues in aqueous microenvironments. These restricted zones of phosphate mineralization result in thin and highly heterogeneous preservation patterns. Furthermore, multiple phases of phosphatization can occur over time, making the overall likelihood of good preservation quite low (Dornbos, 2011; Schiffbauer et al., 2014). Thus, the long-term preservation quality and potential of nanophase goethite rivals that of other high-fidelity preservation media, such as silica, carbonate, phosphate, and amber (Poinar, 1992; Dornbos, 2011; Ruff and Farmer, 2016). The binding affinity of iron to organics, the ability of iron to preserve subcellular details of labile tissues, and the relatively stable nature of nanophase goethite make such iron-rich rocks relatively promising targets in the search for high fidelity morphological preservation of life on Mars, as has previously been suggested (e.g., Fernández-Remolar and Knoll, 2008; Parenteau et al., 2014; Hays et al., 2017).
Future research
Several avenues for future research could enhance our understanding of microfossil preservation in the iron-rich deposits of McGraths Flat to inform detection of past life on Mars. High-resolution trace element analyses of these iron-rich microfossils, including transmission electron microscopy, nanoscale secondary ion mass spectroscopy, and isotopic studies of both fossil and matrix, could support biogenicity and reveal further taphonomic details (e.g., McLoughlin et al., 2007, and references therein; McLoughlin et al., 2011; Levett et al., 2019; 2020a). Other advanced analytical techniques, such as Mössbauer and micro-Raman spectroscopy or time-of-flight–secondary ion mass spectroscopy, may provide complementary phase identification and biogenicity assessment of the iron-rich microbial structures. However, these analyses generally require very flat polished surfaces and a relative abundance of material to feasibly detect organics. To date, no organics have been detected in McGraths Flat. Further, crystallographic studies could investigate the precursor mineral phase of the rod-shaped structures currently interpreted as fossil bacteria (Fig. 1B) (Djokic et al., 2025). Studies of aquatic microorganisms and endoliths from iron-rich environments would provide valuable modern analogs. Such research is particularly important considering that subsurface life likely represents Earth’s largest biomass, yet the literature on palaeolithobiology remains limited, particularly for iron-rich substrates (Whitman et al., 1998; McLoughlin et al., 2007; Walker and Pace, 2007; Omelon, 2016; Tomescu et al., 2016). Additionally, researchers should exercise caution when identifying syngenetic microfossils in Earth’s ancient record, as our understanding of endolithic fossil formation and preservation pathways remains limited (McLoughlin et al., 2007; Tomescu et al., 2016; Ivarsson et al., 2021). Implementing these research approaches could significantly improve our ability to recognize and interpret potential biosignatures in iron-rich rocks on Mars and thus could strengthen future life detection missions.
Conclusions
The assemblage of microfossils preserved at McGraths Flat demonstrates that iron-oxyhydroxides (such as goethite) are excellent at preserving microbial life. The site contains exceptionally well-preserved microfossils in which nanophase goethite has mineral-templated various microbial biopolymers, including those found in bacteria (peptidoglycan) and fungi (chitin), in addition to associated microbial products such as EPS (various natural biopolymers). This highlights the capacity of goethite to preserve a wide range of microbial organisms. The site preserves syngenetic and endolithic microbial generations. Although Mars’ geological history largely rules out fungal (eukaryotic) evolution, these exceptionally well-preserved fossils demonstrate the effectiveness of goethite to preserve a diverse set of microbial remains. Unsurprisingly, morphology and preservation context show that microfossils embedded in the matrix are syngenetic, while it is more difficult to establish the timing of formation for fossils that occupy cavities, unless there is direct evidence of an endolithic origin (such as microborings or etchings). These findings enhance our understanding of microbial paleobiology and establish preliminary observations for discriminating microfossils in ferruginous sedimentary rocks on Earth and potentially Mars.
Footnotes
Acknowledgments
The authors acknowledge the traditional custodians of the land and waterways on which McGraths Flat is located, the Wiradjuri Nation People. Nigel McGrath for access to the fossil site. The Etheridge family descendants (M.R.M.); Australian Museum Research Institute, Australian Museum Trust (M.R.M.); and Australian Research Council (ARC) Linkage Project: LP210301049 (M.R.M., P.V., M.F., and P.S.). They acknowledge the scientific and technical assistance of Microscopy Australia, especially from the Centre for Advanced Microscopy, ANU (jointly funded by the ANU and the Australian Federal Government). They thank Frank Brink and Felipe Kremer for technical assistance. They thank Kliti Grice, Jochen Brocks, and Bonnie Teece for communications on organic preservation in iron and McGraths Flat. P.M.S. published with the permission of the Chief Geoscientist and Head of the Geological Survey of NSW. The use of the AI language model Claude [claude.au] and Grammarly was used to identify grammatical and readability issues, which were reviewed with human oversight and incorporated as appropriate.
Author’s Contributions
T.D., P.M.S., M.R.M., and M.F.: Writing—original draft preparation. T.D.: Conceptualization. T.D. and M.F.: Methodology. T.D. and M.F.: Visualization. T.D., P.M.S., J.O., V.A.K., M.F., J.R.H., D.J.C., R.R., and M.R.M.: Investigation. T.D., P.M.S., J.O., V.A.K., M.F., J.R.H., D.J.C., R.R., and M.R.M.: Writing—reviewing and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Etheridge family descendants (MRM); Australian Museum Research Institute, Australian Museum Trust (MRM); and Australian Research Council (ARC) Linkage Project: LP210301049 (MRM, PV, MF, PS).
Supplemental Material
Associate Editor: Kathleen A. Campbell
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.