
Abstract
Following the Viking experiments in 1976, many of the original inferences of biological metabolism have been replicated via abiotic mechanisms we now know are plausible on Mars’ surface. While in many cases subsequent experiments have cast doubt on whether Viking truly detected life, numerous other studies since Viking have greatly expanded our knowledge of life’s limits and microbial metabolism. In particular, increased characterization of Earth’s subsurface has revealed the astounding complexity and adaptability of life, highlighting chemically based metabolisms as potentially strong targets for future life detection missions. Over the same time frame, we have gained knowledge of putatively more habitable regions in Mars’ subsurface, relative to the original Viking lander surface sites, that could host similar organisms. In this review, we discuss the wealth of knowledge concerning the habitability of zones across Mars’ surface/subsurface, and we suggest specific microbial metabolisms that should be targeted in future life detection missions based on laboratory and field studies under analogous conditions on Earth and with consideration of recommendations from the larger Astrobiology community. The ability to leverage these advancements in subsurface research toward the incorporation of increased specificity in future life detection efforts is additionally discussed in the context of current Mars subsurface mission progress and planetary protection and defense concerns.
Introduction
Fifty years since the original experiments, debate continues over the role of biological metabolism versus abiotic interference in the Viking 1976 lander life detection mission (McKay et al., 2025). Despite disagreements on abiotic versus biotic processes measured in the original approach (Klein, 1978), the life detection experiments performed as part of Viking stand to this day as robust tests of core biological metabolism. Specifically, the three primary experiments performed by the original Viking lander, including the Labeled Release Experiment, the Gas Exchange Experiment, and the Pyrolytic Release Experiment, correspond to three ubiquitous characteristics of terrestrial microbiology: transformation of fixed carbon (heterotrophy), generation of chemical disequilibrium (kinetic production of metabolic byproducts), and fixation of organic carbon from inorganic sources (phototrophy or chemotrophy) (Biemann et al., 1977; Levin and Straat, 1977; Oyama and Berdahl, 1977). Despite the advantageous design of Viking’s experiments to test these biologically fundamental processes, this same strength in generality led to weaknesses in the eventual interpretation of its results, particularly in the context of an abiotically harsh and highly oxidizing martian surface (Lasne et al., 2016).
Much has been learned about the range of metabolisms and extreme environments capable of supporting life on Earth that could better inform experiments and locations of future life detection missions in the nearly 50 years since Viking (Rothschild and Mancinelli, 2001). This acquired research especially highlights Earth’s subsurface as a suitable host for a complex diversity of chemotrophic life, spanning a wide variety of environments analogous to potential subsurface zones on Mars (Tarnas et al., 2021). In the 1980s and 1990s, an early and very important observation in this field saw the discovery of highly diverse life active in and around hydrothermal vents at the seafloor (Corliss et al., 1979; Jannasch, 1983; Baross and Hoffman, 1985; Deming and Baross, 1993; Wirsen et al., 1993; Kelley et al., 2002). This work expanded the range of conditions known to support life and demonstrated that complex cellular communities could flourish off the concentrated redox products of water–rock interaction with a chemolithoautotrophic base of the food chain. Characterization of life in the subsurface continued with discoveries of vast, deeply buried microbial ecosystems across Earth, expanding our known distribution of life to kilometer-scale depths (D’Hondt et al., 2004; Chivian et al., 2008; Ahonen et al., 2010; Payler et al., 2019; Ford et al., 2024; Nisson et al., 2024). Examples of such settings include high-energy-flux hydrothermal systems, frozen permafrost soils, deeply buried pelagic marine sediments, deep oceanic basalts, and deep terrestrial aquifers that have been stable for billions of years (Kelley et al., 2002; Michalski et al., 2013; Orcutt et al., 2013; Michalski et al., 2018; Onstott et al., 2019).
Since Viking, the range of known metabolisms has also expanded. It now includes anaerobic methanotrophy (Hoehler et al., 1994), respiration of an increasingly large array of metals and complex carbon compounds (Amend and Shock, 2001), and even the direct extracellular transfer of electrons (Hernandez and Newman, 2001). The adoption of direct sequencing of DNA from environmental samples revealed the existence of many deep branches on the tree of life that had been missed by the traditional microbial culturing techniques (Rappé and Giovannoni, 2003; Lloyd et al., 2018). The expansion of the known range of analogous habitats (including the marine and terrestrial subsurfaces) and the range of microbial metabolic strategies that have happened since the Viking missions ended suggests that the possibilities for life on Mars are even more expansive than was known at the time of those missions. These findings also allow us to evaluate life in a metabolic context beyond broad classifications (e.g., heterotrophy, phototrophy, and/or autotrophy) and instead consider and target specific metabolic strategies and substrates that we now know support life in analogous subsurface environments of Earth (Merino et al., 2019; Beaver and Neufeld, 2024).
In this review, we highlight advances over the last 50 years in laboratory and field-based microbiology and how these can inform future approaches to life detection on Mars in the context of the past Viking experiments as landmark detection efforts for microbial metabolism. Specific consideration is given to environments of the subsurface and how terrestrial analog work including laboratory and field studies supports this region as a good target for future life detection missions on Mars. To frame this discussion, we pose the following three primary questions:
What makes the subsurface a strong candidate for future life detection efforts on Mars, relative to the original surface setting explored by the Viking experiments? Has recent research revealed more likely microbial metabolisms and tolerances that could persist under martian conditions, and are there one or more metabolisms that seem most plausible? How does knowledge gained from laboratory and field studies of subsurface microbiology over the past 50 years guide our decisions moving forward with regard to life detection on Mars?
Building off the initial surface-based approach of Viking, we can revisit our search for extant life on Mars and beyond with an improved selection of environmental context, a refined focus on metabolic targeting, and a greater understanding of how to mitigate and interpret abiotic interference in life detection experimentation.
There are numerous environmental factors that may impact habitability at the martian surface (Clark, 1998; Stoker et al., 2010; Schuerger et al., 2012), including low water availability in the regolith (Grant, 2004; Tosca et al., 2008), extreme fluctuations in temperature (Spanovich et al., 2006), and high levels of radiation/oxidant exposure (Hassler et al., 2014). It has been suggested that the martian subsurface, at various depths, likely provides secluded, more habitable environments relative to the martian surface (Onstott et al., 2019; Stamenković et al., 2019). Perhaps the three most important characteristics of a habitable subsurface environment on Mars are the availability of (1) liquid water at habitable temperatures, (2) protection from the cosmic and ionizing radiation that pummels the surface of the planet, and (3) an energy source for metabolism (Fig. 1). These factors are discussed in greater detail below, supporting regions of the martian subsurface as beneficial alternatives for future life detection experiments.
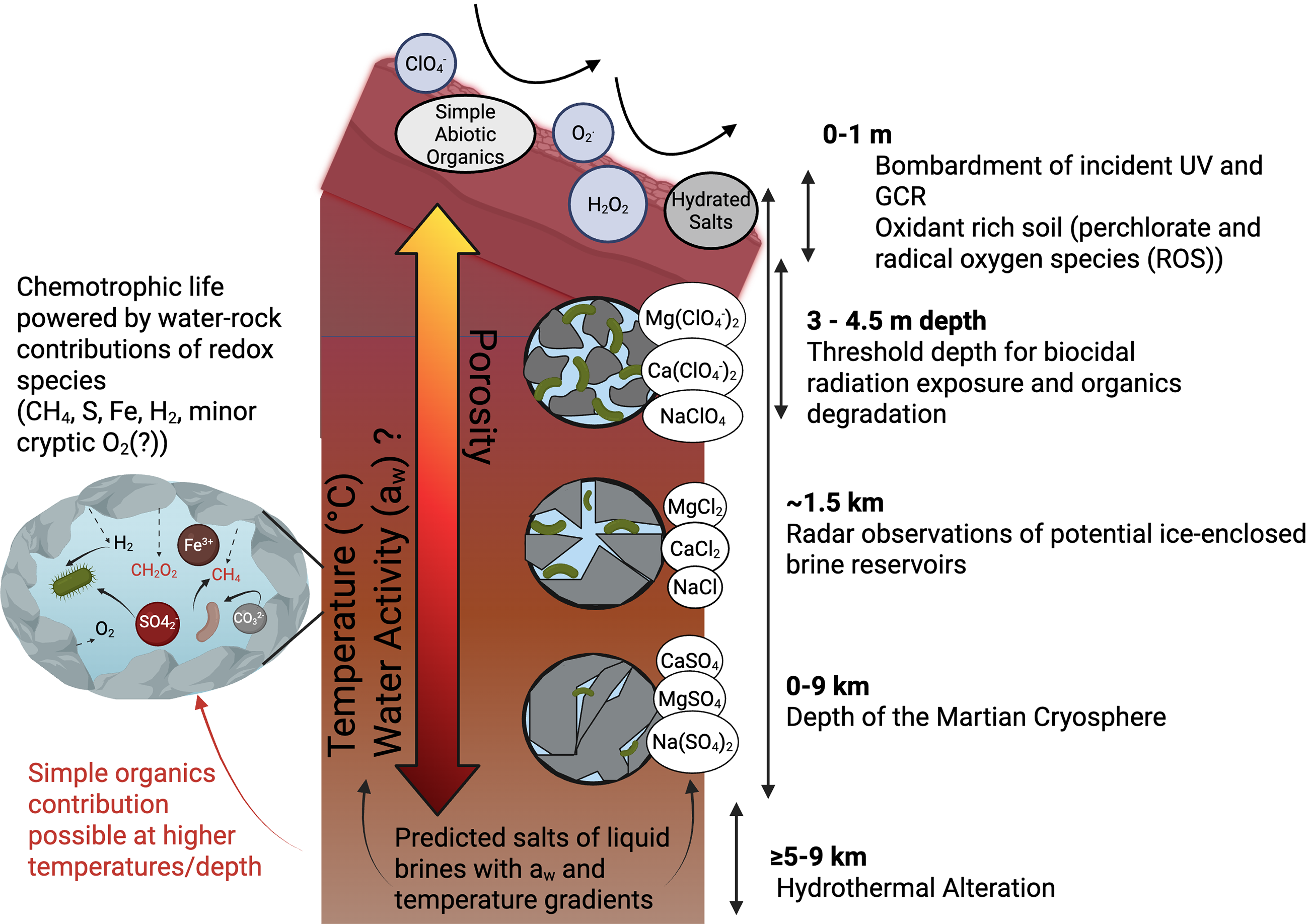
Physical and chemical gradients predicted to dictate habitability across depth in the martian subsurface. These include (1) constraints on water; although water activity (aw) gradients are not yet known for Mars, predicted liquid brine stability depends on primary ionic composition with perchlorate-rich brines transitioning into chloride- and sulfate-enriched brines with depth and temperature (Chevrier and Slank, 2024; Stevens and Cockell, 2023). It is important to note that porosity is expected to decrease with depth, which may also influence water availability, with any potential brines increasingly localized to pore spaces and fractures (Michalski et al., 2018). (2) Surface radiation exposure is expected to subside to lower background radionuclide decay around 3–5 m depth. (3) If liquid water is present at depth, chemotrophic microbial communities could utilize the products of water–rock interaction including inorganic electron donors/acceptors and simple abiotic organics. Image created using biorender.com.
Transient liquid water may be available on the surface of Mars in the form of recurring slope lineae (McEwen et al., 2014; Ojha et al., 2015). The intermittent nature of these brines, however, implies that they would not constitute stable or long-term habitats on the surface, even if upwelled from subsurface aquifers (Stillman et al., 2016; Abotalib and Heggy, 2019; Liu et al., 2025). More stable water in the near-surface or subsurface is expected to vary with location and phase (liquid water or ice) (Boynton et al., 2002; Chevrier and Slank, 2024; Wright et al., 2024). Unbound water deposits are estimated to account for 20–34 m global equivalent layer (GEL) water stored in near-surface regions (within hundreds of micrometers of the surface and including the polar ice caps; Carr and Head, 2015; Carr and Head, 2019). A much larger amount of water (130–260 m GEL) is expected to be tied up in, or required for the formation of, hydrated mineral deposits on Mars (Wernicke and Jakosky, 2021). While these are global estimates, water-enriched locales have also been identified, particularly at Meridiani Planum with an average of 7 wt % water equivalent hydrogen (WEH) as measured for the region by the neutron spectrometer on Mars Odyssey (Feldman et al., 2004; Clark et al., 2005). For further context in units of WEH, global estimates suggest equatorial ranges between 2 and 10 wt % WEH with averages between 5 and 7 wt % WEH (Feldman et al., 2004; Maurice et al., 2011; Wernicke and Jakosky, 2021).
Exactly how water availability extends into the martian subsurface from surface/shallow subsurface enriched regions likely also depends on major ionic composition and geothermal temperatures. For instance, under a modern martian geothermal gradient (10°C/km from Michalski et al., 2013,2018), any near-surface pockets of liquid would harbor extremely low water activities (aw) bordering the limits of life (∼0.49 aw; Paris et al., 2023; Doran et al., 2024) with a primary composition of chloride and/or perchlorate salts that allow for eutectic brine formation ≤−20°C (Chevrier and Slank, 2024; Chevrier, 2025). This includes relatively shallow subsurface depths (∼1.5 km) where previous Mars Advanced Radar for Subsurface and Ionosphere Sounding observations identified a putative liquid brine pocket (Picardi et al., 2005; Orosei et al., 2018; Lauro et al., 2020). As temperatures increase in the deeper subsurface (≥−20°C at ∼3 km depth, ≥0°C at ∼5 km depth; Michalski et al., 2013,2018), brines could increase in concentration of sulfate-dominated salts (Chevrier et al., 2024), also offering an environment of lower toxicity (Rzymski et al., 2024) and beneficial redox species abundance (e.g., iron and sulfate) at depth. While life has been inferred to maintain (very slow) metabolic activity down to −40°C (Price and Sowers, 2004), with this limit subsequently argued to be closer to −28°C to −20°C (Clarke et al., 2013; Doran et al., 2024), the combined water availability and increased temperatures lend to greater habitability likely in Mars’ deeper subsurface.
Radiation
The inability to confidently assign a biological origin to the Viking experiments has since been attributed to a strong abiotic background, likely due to the presence of oxidizing species in the surface regolith (Lasne et al., 2016; McKay et al., 2025). Radiation exposure on the surface of Mars is strong, particularly given the planet’s thin atmosphere and the lack of an ozone layer (Kuhn and Atreya, 1979; Cockell et al., 2000). Additionally, from the lack of a magnetic field, ionizing radiation such as galactic cosmic radiation (GCR) (Saganti et al., 2004) and solar energetic particles (Hassler et al., 2014) also reach the martian surface. This exposure increases oxygen (O2) radical formation through the irradiation of hydrogen peroxide (H2O2) and perchlorates (Davila et al., 2013; Zhang et al., 2022), forming damaging superoxide species and excess molecular O2 (Yen et al., 2000; Georgiou et al., 2007). For instance, the abiotic formation of O2 in the original Gas Exchange Experiment has been observed in cryptic O2 production from perchlorate irradiation (Quinn et al., 2013) and has been proposed in the abiotic dismutation of H2O2 in semi-aqueous environments on early Earth (He et al., 2021). In addition to ionizing radiation, ultraviolet (UV) irradiation has been found responsible for the abiotic formation of simple organic species on mineral surfaces, such as those putatively observed in the Pyrolytic Release Experiment (Hubbard, 1979; Poch et al., 2014). It has since been suggested that the decomposition product of perchlorate, hypochlorite, could have acted to oxidize organics, including the amino acid alanine, added to the martian regolith in the Labeled Release Experiment, resulting in observed labeled carbon dioxide (CO2) formation (Quinn et al., 2013). Although none of these explanations are considered conclusive today, the potential for abiotic influence highlights the challenges imposed by the martian surface on life detection. Constraints have been proposed to 3 m or greater for the survivability of microbial cells in a lower radiation martian subsurface (∼1 μGy/day vs. 480 μGy/day on the surface) (Dartnell et al., 2007; Hassler et al., 2014). At this depth, microbial cells would be shielded from GCR and UV from the surface, with primary exposure at depth from crustal radionuclide decay (Dzaugis et al., 2018). Subsurface life may also need to adapt to toxicity from perchlorate exposure (Rzymski et al., 2024) that could be transferred to depth via aqueous dissolution and fluid migration in the shallow subsurface (Cull et al., 2010). While similar processes may have moved perchlorates into the deeper subsurface, with subsurface perchlorate brines predicted on Mars (Lauro et al., 2020), specific mechanisms and concentrations are not yet known.
Energy
The range of energy metabolisms that are present in subsurface environments is highly dependent on the availability of geochemical substrates, a phenomenon that has been observed across several subsurface field studies on Earth. For instance, one can track a metabolic shift in subsurface communities distributed across a subduction zone and see sulfur-based respirations early in subduction (where the downgoing tectonic plate is still releasing ancient seawater) transitioning to more iron and nitrogen cycling stimulated by inorganic carbon (e.g., carbon monoxide [CO] and/or CO2) and hydrogen (H2) release in later near-volcanic subduction zones (Fullerton et al., 2021; Rogers et al., 2023). In marine sediments, life is largely heterotrophic, metabolizing recalcitrant ancient organic matter using sulfate, iron, or CO2 as terminal electron acceptors (D’Hondt et al., 2004; Biddle et al., 2006). In the case of the deep terrestrial subsurface, organisms can be found in ancient aquifers harboring very low microbial biomass, supported by the products of Earth’s slow radioactive decay (Lin et al., 2005; Chivian et al., 2008). These strong ties between microbial localization and metabolic substrate availability emphasize the need to characterize geochemical availability in subsurface environments on Mars.
Chemolithoautotrophs are one such metabolic group, represented by relatively simple metabolisms and likely among the first metabolic guilds to arise on Earth (Battistuzzi et al., 2004; Ueno et al., 2006; Shen et al., 2009; Bontognali et al., 2012), with the same trajectory potentially having occurred in Mars’ early history, if life evolved there. Organisms in this category, such as methanogenic archaea, require only simple inorganic substrates such as H2, CO, and CO2 as energy and/or carbon sources (Borrel et al., 2013), whereas sulfate-reducing bacteria can use H2 as an electron donor and sulfate, sulfite, or thiosulfate as electron acceptors (Tarnas et al., 2021; Zhou et al., 2025). Substrate availability to fuel these metabolisms on Mars includes abundant calcium, magnesium, and iron sulfates throughout the regolith (Lane et al., 2004; Arvidson et al., 2005; Bibring et al., 2005; Gendrin et al., 2005; Bibring et al., 2006; Squyres et al., 2012; Arvidson et al., 2014; Nachon et al., 2014; Kronyak et al., 2019; Bishop et al., 2025). Hydrogen, for use as a potential electron donor, is present in the martian atmosphere and proposed in the subsurface through a variety of mechanisms including serpentinization and radiolysis (Oze and Sharma, 2005; Schulte et al., 2006; Sherwood Lollar et al., 2014; McMahon et al., 2016; Tarnas et al., 2018; Tarnas et al., 2021; Nisson et al., 2024; Tosca et al., 2025). Simple organic sources of carbon could serve as an additional option for chemotrophic life to use as an electron donor, with the formation of simple hydrocarbons via Fischer–Tropsch type synthesis alongside transition metal catalyzed acetate and formate in radiolytic and/or serpentinizing subsurface systems (Sherwood Lollar et al., 2002; McCollom and Seewald, 2007; Dzaugis et al., 2018; Sherwood Lollar et al., 2021). However, the proposed fluids in these studies supporting the abiogenic formation of these organics only harbor temperatures as cool as 25–100°C, with the actual formation of organics likely occurring before total cooling. This would largely constrain availability of similar compounds to depths below the cryosphere (≥5–9 km estimated around the martian equator by Clifford and Parker, 2001, and Clifford et al., 2010) (Fig. 1).
Microbial Activity in the Martian Subsurface—Energy Metabolism and Stress Defense
If a future life detection mission on Mars were to focus on the subsurface, experiments to test extant metabolic activity should be redesigned to better target strategies performed by life in analogous subsurface settings on Earth. Fortunately, the past 50 years of research into microbial metabolic function under simulated martian conditions and into the terrestrial and marine subsurface regions of Earth, have garnered a wealth of information on light-independent, chemotrophic-based life for catabolic energy gain. Chemotrophic organisms are known to survive under low-energy conditions and adopt defense strategies for dealing with high abiotic stress (Jones, 2018) that are still a concern for the martian subsurface as detailed above. In many cases, the most ubiquitous metabolic strategies are also the most phylogenetically robust; they appear across deeply rooted lineages in the tree of life (Moody et al., 2025). With regard to energy metabolism, this includes what are considered the oldest metabolisms on Earth in methanogenesis, arising prior to 3.6 billion years ago (Ga) (Battistuzzi et al., 2004; Lyons et al., 2024), and various sulfur- and iron-utilizing strategies (Blank, 2004; Philippot et al., 2007; Ilbert and Bonnefoy, 2013; Ward et al., 2021; Mateos et al., 2023). Importantly, these energy-yielding catabolic metabolisms are used by many of the same microorganisms living in extreme conditions and employing similar anabolic (energy-burning) stress-defense metabolisms to combat physical and chemical constraints of the subsurface (Hoehler, 2007). Here, we review the primary energy-yielding metabolic and energy-burning stress defense categorizations of life found in Earth’s subsurface, as revealed through decades of laboratory and field-analog research since Viking. Additionally, we build on decades of previous literature that has highlighted the potential for chemosynthetic and anaerobic metabolisms on Mars, suggesting metabolisms with specific relevance to the martian subsurface (Boston et al., 1992; Shock, 1997; Fisk and Giovannoni, 1999; Mancinelli, 2000; Chapelle et al., 2002; Kotelnikova, 2002; Mancinelli, 2003; Jepsen et al., 2007; Westall, 2013; Westall et al., 2013; King, 2015; Westall et al., 2015; Checinska Sielaff and Smith, 2019; Onstott et al., 2019; Atri, 2020; Rucker et al., 2023; Butturini et al., 2025).
Candidate energy metabolisms for life in the subsurface
Methanogenesis
Methanogenesis is considered one of the oldest metabolisms on Earth, with some estimates suggesting methanogens arose between 3.78-4.11 Ga, with the origin of life on Earth occurring prior to 4.1 Ga (Battistuzzi et al., 2004). More specifically, hydrogenotrophic methanogenesis is considered to be the most ancestral pathway for methanogenesis, with estimates suggesting it arose at least 3.46 Ga (Ueno et al., 2006; Lyons et al., 2024). The ability for methanogens to use simple, inorganic substrates (CO, CO2, H2) for energy and metabolism and their ability to exist in extreme subsurface environments (Kotelnikova and Pedersen, 1997; Kotelnikova, 2002; Moser et al., 2005; Inagaki et al., 2015), shielded from harmful ionizing and UV radiation, have suggested that these archaea may be model organisms for both past and present life on Mars (Boston et al., 1992; Chapelle et al., 2002; Paulchamy et al., 2024; Butturini et al., 2025). However, the presence of methane (CH4) (Krasnopolsky et al., 1997; Formisano et al., 2004; Geminale et al., 2008; Mumma et al., 2009; Geminale et al., 2011; Webster et al., 2015; Webster et al., 2018; Webster et al., 2021) in the martian atmosphere is not only controversial (e.g., Lefèvre and Forget, 2009; Zahnle et al., 2011; Lefèvre, 2018; Viscardy et al., 2025) but its abiotic versus biotic origin is also debated (Atreya et al., 2007; Atreya et al., 2011). Nevertheless, considering that most of the CH4 in Earth’s atmosphere is due to biogenic processes (Schoell, 1988), the presence of CH4 in the martian atmosphere represents an exciting possibility for extant or extinct life on Mars and has fueled decades of research on methanogen growth and survivability under extreme conditions.
Numerous laboratory studies have explored the effect of individual or combined martian environmental conditions on methanogen growth and survivability (e.g., Kral et al., 2004; Morozova et al., 2007; Morozova and Wagner, 2007; Chastain and Kral, 2010; Kral et al., 2011; Kral and Altheide, 2013; Kral et al., 2014; Schirmack et al., 2014; Sinha and Kral, 2015; Mickol and Kral, 2017; Sinha et al., 2017; Mickol et al., 2018a; Mickol and Kral, 2018; Mickol et al., 2018b; Sinha and Kral, 2018; Maus et al., 2020; Harris et al., 2021; Sinha et al., 2021; Harris and Schuerger, 2025). For instance, freeze/thaw experiments including osmotic and thermal stresses (Serrano et al., 2019) indicated that even thermophilic species can return to a metabolically active state following drastic temperature changes, given sufficient nutrients and time (Mickol et al., 2018a; Mickol et al., 2018b), which could be reconciled with specific climatic shifts in Mars’ history. Methanogen responses to low temperatures include adjusting membrane fluidity and activating cold-shock response genes (among other strategies; Cavicchioli, 2006; Mickol et al., 2018a; Mickol et al., 2018b; Serrano et al., 2019; Paulchamy et al., 2024). Additional defense strategies by Methanosarcina species, specifically, include robust aggregate formation, protecting interior cells in the face of harsh conditions (Morozova et al., 2007; Morozova and Wagner, 2007). These seemingly resilient methanogenic archaea have also been found to utilize nutrients from martian regolith simulants for growth (Kral et al., 2004; Schirmack et al., 2015; Sinha and Kral, 2015; Craig et al., 2017). Further support for the possibility of biogenic methanogenesis on Mars has been seen in theoretical modeling: Sauterey et al. (2022) and Rucker et al. (2023) found energetic support for the metabolism on Noachian Mars. Under modern martian subsurface conditions, however, Sholes et al. (2019) estimated a supported methanogen biomass nearly 105 times less than that of Earth, which emphasizes the need for life detection measurements adapted for low density and/or slower growing life.
Sulfur metabolism
Sulfur-metabolizing bacteria can refer to sulfate/sulfite reducers, sulfur/sulfide oxidizers, and thiosulfate and/or elemental sulfur disproportioners (Zhou et al., 2025). Similar to methanogenesis, sulfate reduction is considered one of the most ancient metabolisms on Earth, with evidence for sulfate reduction by sulfate-reducing bacteria present on Earth as early as 3.4–3.5 billion years ago (Ohmoto et al., 1993; Shen et al., 2001; Ueno et al., 2008; Shen et al., 2009; Roerdink et al., 2012; Roerdink et al., 2016; Mateos et al., 2023). Support for the existence of sulfur metabolisms on Mars mainly results from the interpretation of Mars-analog environments and microbially mediated reactions on Earth (Pontefract et al., 2017; McMahon et al., 2020; Oliver et al., 2022; Macey et al., 2023). For example, using spatially resolved ion microprobe (secondary ion mass spectrometry) measurements, McMahon et al. (2020) determined that sulfate within gypsum (calcium sulfate) veins of possible Cenozoic age (<66 million years ago) was likely the result of microbial sulfate reduction based on the high fractionation of the sulfur and the presence of pyrite surrounding the veins. The authors suggest that similar metabolisms could have occurred in the martian subsurface, with microbial sulfate reduction coupled to the oxidation of CH4 and/or H2, where CH4 and H2 could be produced abiotically through serpentinization or radiolysis, or by methanogenesis (McMahon et al., 2020).
There is also the potential that redox (dis)equilibriums enable sulfur-dependent microbial metabolism such as sulfide oxidation and sulfate reduction (Lefticariu et al., 2010; Norlund et al., 2010; Macey et al., 2020; Tarnas et al., 2021; Moreras-Marti et al., 2022; Wong et al., 2022; Hurowitz et al., 2025). From radiolysis alone, Tarnas et al. (2021) estimated that the martian subsurface could support a similar number of sulfate-reducing bacteria as the deep subsurface on Earth (ca. 105 cells/kg rock; Onstott et al., 2019), with the possibility for higher cell densities in regions with concentrated sulfide deposits. Further investigation of sulfate deposits is considered key to improving our understanding of Mars’ hydrogeological and potentially habitable history (Figueroa et al., 2024; Ettehadi et al., 2025; Sellam et al., 2025; Tino et al., 2025; Yeşilbaş et al., 2025). Importantly, the detection of organic matter on Mars (Eigenbrode et al., 2018; Stern et al., 2022) and large fractionation in sulfur isotopes measured at Gale crater (Franz et al., 2017) both lend credence to the possibility of past microbial sulfate reduction, as well as the potential for thermochemical sulfate reduction of organic matter (Cai et al., 2022). As opposed to methanogens, laboratory experiments exposing sulfur-metabolizing bacteria to martian conditions are relatively sparse. Using the anaerobic psychrophile Desulfotalea psychrophila, Mosquera et al. (2023) demonstrated sulfate reduction under Mars-relevant concentrations of calcium, magnesium, and iron sulfates, as well as low temperature (0°C, −5°C). However, data shown represent only endpoint analyses; growth metrics over time would provide stronger evidence for the ability of sulfur-metabolizing species to thrive on past or present Mars.
Iron metabolism
On Earth, the utilization of iron as an electron donor [ferrous iron—Fe2+] and as an electron acceptor [ferric iron—Fe3+], in the metabolic processes of iron oxidation and reduction, respectively, is ubiquitous across many subsurface chemotrophic communities (Straub et al., 2001; Casar et al., 2021). The former typically occurs in highly reducing environments that harbor acidic conditions, such as acid mine drainage sites, where ferrous iron from pyrite is oxidized by chemolithotrophic microbes to form secondary ferric iron oxides such as hematite (Price et al., 2018; Floyd et al., 2019; Fifer and Wong, 2024). Río Tinto is considered one such highly acidic site on Earth that is regarded as a martian habitability analog, since it contains similar iron oxide and sulfur-containing evaporite mineral deposits to those of Meridiani Planum (Fernández-Remolar et al., 2005; Souza-Egipsy et al., 2006). Alternatively, iron reduction involves the anaerobic implementation of ferric iron as an electron acceptor. Because this ferric iron is typically in the form of insoluble ferric (hydr)oxides, this strategy requires extracellular electron transfer (Boston et al., 1992; Nealson and Cox, 2002; Shaffer et al., 2025). Both iron oxidation and reduction are relatively old in terms of metabolic phylogeny (≥2.4 Ga; Ilbert and Bonnefoy, 2013), and if chemotrophic life evolved on Mars at some point, iron is considered a strong potential candidate to support such metabolism, given the planet’s iron-enriched chemistry (Price et al., 2018). This is particularly due to observations of ferrihydrite, various other iron oxides, and minor pyrite minerals in regions of the martian surface that may extend to the subsurface (Wong et al., 2022; Valantinas et al., 2025).
Other anaerobic strategies
Due to the complexity and diversity of chemotrophic life found in Earth’s subsurface, other anaerobic strategies must be noted for Mars in addition to the primary anaerobic metabolisms detailed above. For instance, if abiotic or biotic CH4 is present, then we must consider the potential for anaerobic methanotrophy/CH4 oxidation (Marlow et al., 2014; Guerrero-Cruz et al., 2021), as well as acetogenesis, which can also occur through conversion of CO2 and H2 (Boston et al., 1992). Additionally, in several extreme environments on Earth, microbial communities have been found that can utilize typically toxic metals such as arsenic and selenium as alternative electron acceptors (Maiers et al., 1988; Zecchin et al., 2021). Given that these metals have also been reported in the martian regolith (Gellert et al., 2018; Wang et al., 2025), albeit at minor concentrations, they could supplement energy metabolism in lieu of O2 availability. Metal utilization from solid substrates (such as iron oxides mentioned above) can additionally involve the transfer of electrons extracellularly in some microbial species, which could help shape redox networks and connect habitable zones (up to millimeter scale) within the subsurface (Shi et al., 2016; Yang et al., 2021). Finally, some microbial species are also known to reduce perchlorate/chlorate directly, which could be an advantageous strategy among the abundant perchlorate salts on Mars (Coates and Achenbach, 2004; Oren et al., 2014).
Potential for cryptic oxygen-fueled aerobic metabolism
Aerobic metabolism is largely overlooked as a feasible life-supporting strategy on Mars, since the atmosphere contains low O2 (0.13% atmospheric composition relative to 21% on Earth; Luo et al., 2016; Gupta et al., 2024). Additionally, the major O2 production pathway here on Earth, oxygenic photosynthesis (Dismukes et al., 2001), is likely nonviable on the Red Planet’s freezing and high radiation surface (McKay, 2010), although it has been suggested as a potential pathway in martian snow and ice (Khuller et al., 2024). Oxygenic photosynthesis, however, is not the only potential O2 production mechanism, with several “cryptic” O2 production pathways since identified on Earth that could be sustained via similar mechanisms on Mars (Ruff et al., 2024). Both biotic and abiotic mechanisms (not including phototrophy) have been modeled and/or identified in both terrestrial and aquatic settings of Earth’s surface and subsurface (Ruff et al., 2022,2023, 2024). Examples include the production of O2 from radionuclide decay (e.g.,238U, 232Th, 40K) (Sauvage et al., 2021; Nisson et al., 2024), electrolysis via polymetallic nodule generated currents (Sweetman et al., 2024), or biotic dismutation pathways for chlorite, nitrous oxide, and peroxides/superoxides (Ettwig et al., 2012). Some studies have proposed the ability for O2 production on planetary surfaces via irradiation of ice or sulfur dioxide (SO2) with space-originating charged particles (e.g., Teolis et al., 2017; Wallner et al., 2022; Szalay et al., 2024) that could occur given similar compositions found on modern or past Mars (Titus et al., 2003; Khayat et al., 2015; Chevrier et al., 2024). While the exact mechanisms and quantifications of O2 production on Mars’ surface and in the subsurface are not yet known, these proposed processes mean we cannot discount O2 as a potential electron acceptor (including in the oxidation of CH4, sulfur, and iron compounds mentioned previously). Furthermore, O2 has been observed to exhibit seasonal variations in the martian atmosphere (Trainer et al., 2019), and while the mechanism is also unknown, this seasonality has been suggested as a potential biosignature similar to CH4 (Yung et al., 2018).
Candidate stress defense strategies for life in the martian subsurface
High salt
Halophilic microorganisms, which thrive in regions on Earth with naturally high salinity, have been another focus for astrobiology studies due to the existence (or predicted existence) of brines throughout various sites within our solar system, for example, Mars, Europa, Enceladus, and Ceres (Mancinelli et al., 2004; Oren, 2014; Renno et al., 2021; Nathues et al., 2022; Chevrier and Slank, 2024; Perl et al., 2025). With relevance to Mars, many studies have focused on the tolerance of halophilic microorganisms to oxidizing and chaotropic perchlorate and chlorate salts, and UVC-induced oxidizing derivatives (Wadsworth and Cockell, 2017), that are abundant in the surface regolith (Youngblut et al., 2016; Gault and Cockell, 2021; Fig. 1). A vast collection of studies has demonstrated the tolerance of many different microorganisms to high concentrations of perchlorates (Oren et al., 2014; Heinz et al., 2019; Heinz et al., 2020; Fischer et al., 2024; Zbeeb et al., 2025), extending to some of the strongest candidate metabolic guilds under martian conditions such as methanogenic archaea (Shcherbakova et al., 2015; Kral et al., 2016; Serrano et al., 2019; Harris et al., 2021). Specifically, the intracellular accumulation of salts (such as potassium chloride) and/or the biosynthesis of organic compatible solutes have been observed as defense strategies across halotolerant bacteria, archaea, and eukarya to diminish the chaotropic effect of perchlorates (Heinz et al., 2022; Liu et al., 2024; Díaz-Rullo and González-Pastor, 2025). However, in cases where perchlorates can be used directly in anaerobic metabolism as electron acceptors, incomplete biological perchlorate reduction can occur, producing more harmful perchlorate derivatives (such as chlorite) and a lower overall microbial tolerance (Park and Marchand, 2006; McAdam and Judd, 2008; Ren et al., 2021). In the case of subsurface perchlorate brines, microorganisms would be expected to adopt one or more of these strategies.
Low pressure
The growing subfield of low-pressure (<500 mbar) microbiology (Kral et al., 2011; Fajardo-Cavazos et al., 2012; Schirmack et al., 2014; Waters et al., 2014; Waters et al., 2015; Mickol and Kral, 2017; Mickol and Kral, 2018; Schwendner and Schuerger, 2020; Waters et al., 2021) has expanded our understanding of the biocidal properties (e.g., hypobaria, UVC, desiccation, and (per)chlorates) of the martian surface and astrobiological potential of the subsurface (Verseux, 2020). More specifically, hypopiezotolerant microorganisms are microbes capable of surviving and growing in the low-pressure conditions expected near the martian surface and shallow subsurface, that is, <10 mbar (Kral et al., 2011; Fajardo-Cavazos et al., 2012; Schuerger et al., 2013; Schirmack et al., 2014; Waters et al., 2014; Waters et al., 2015; Schuerger and Nicholson, 2016; Fajardo-Cavazos et al., 2018; Mickol and Kral, 2018; Schwendner and Schuerger, 2020; Waters et al., 2021; Schuerger and Kelley, 2025). Interestingly, several Carnobacterium spp. strains have been observed to thrive in low-pressure conditions comparable to Mars, even though hypobaric conditions are thought to disrupt cell membrane integrity (Nicholson et al., 2013; Miller et al., 2023). In other cases, however, low pressure (ca. <50 mbar) appears to be fairly biocidal, potentially due to compounding factors, with many low-pressure studies limited in duration due to the instability of liquid in microbiology growth media under Mars-like pressures (Schuerger and Nicholson, 2006; Mickol and Kral, 2017; Schuerger et al., 2020; Schwendner et al., 2020). However, experiments can be extended to months under Mars-relevant pressures with a stable brine growth medium (Robinson and Ulrich, 2022). For example, halophilic archaea can thrive or survive in high salinity and perchlorate concentrations in anoxic, low-pressure environments representative of Mars, but they typically require complex carbon sources, such as glucose or yeast extract, which are absent on Mars (Robinson and Ulrich, 2022). Conversely, methanogens can grow on simple Mars-like carbon sources (e.g., CO2) under anoxic, low-pressure (here, ca. 7–500 mbar) conditions (Kral et al., 2011; Schirmack et al., 2014; Mickol and Kral, 2018; Harris and Schuerger, 2025). Results from these and similar analog studies gathered since the Viking mission suggest that extreme conditions on Mars may be tolerable, yet no natural site on Earth perfectly replicates present-day martian environments. This is particularly true with regard to the low pressure of Mars, which has not been found in natural environments on Earth (standard sea-level surface pressure of 1013 mbar), unlike other stressors considered (e.g., salinity, redox substrate availability), which stipulates that terrestrial life has never faced evolutionary pressure to evolve to low pressure conditions (perhaps unless an atmospheric ecosystem is considered; Waters et al., 2014; Waters et al., 2015; Mickol and Kral, 2017). Future experiments involving Earth’s microbes under laboratory-simulated martian environmental conditions will be crucial for improving our understanding of the potential for extant life, as well as for the development of containment systems that aim to culture potential extant martian microorganisms (Schwendner and Schuerger, 2020).
Extreme energy limitation
Even if extant life were present in the martian subsurface, the detectability of such life may still be exceedingly difficult. In most of Earth’s subsurface, the energy delivery rate, or power, is too low to allow even a single cell division (Bradley et al., 2020). Since subsurface sediments have been buried for thousands to millions of years, without ever experiencing enough power to fuel cell division, it is possible that individual cells have extremely long lifespans (Hoehler and Jørgensen, 2013; Lever et al., 2015). These aeonophiles, or organisms that make use of long time periods in the same way that thermophiles make use of high temperatures, are better studied through their biosignatures or metabolic activity rather than through direct interrogation of cell proliferation in the environment (Lloyd and Steen, 2025). Although Earth’s aeonophiles can undergo cell division in subsurface environments when the power is increased slightly (Lloyd et al., 2020) or in carefully designed cultures with limited nutrients (Imachi et al., 2020), they nevertheless grow very slowly. If martian microbial life is similarly power-limited and aeonophilic, it would be important to avoid detection methods that require cell growth. A more comprehensive look at the specific experiments performed by the Viking mission, their outcomes, and possibilities for improvement can be found elsewhere (Calomiris, 2026). These recommendations for target metabolisms and discussion of likely microbial adaptation strategies do not imply the presence of thriving ecosystems in the subsurface. Instead, they suggest that where life as observed across extreme constraints on Earth can survive and maintain active metabolism, the same may be true for analogous subsurface regions along Mars’ geothermal gradient (Fig. 2). While interpretations of the Viking experiments remain controversial, our understanding of the likelihood of extant life on Mars has advanced substantially since those early missions, making the search for extant life a priority objective for Mars science moving forward.
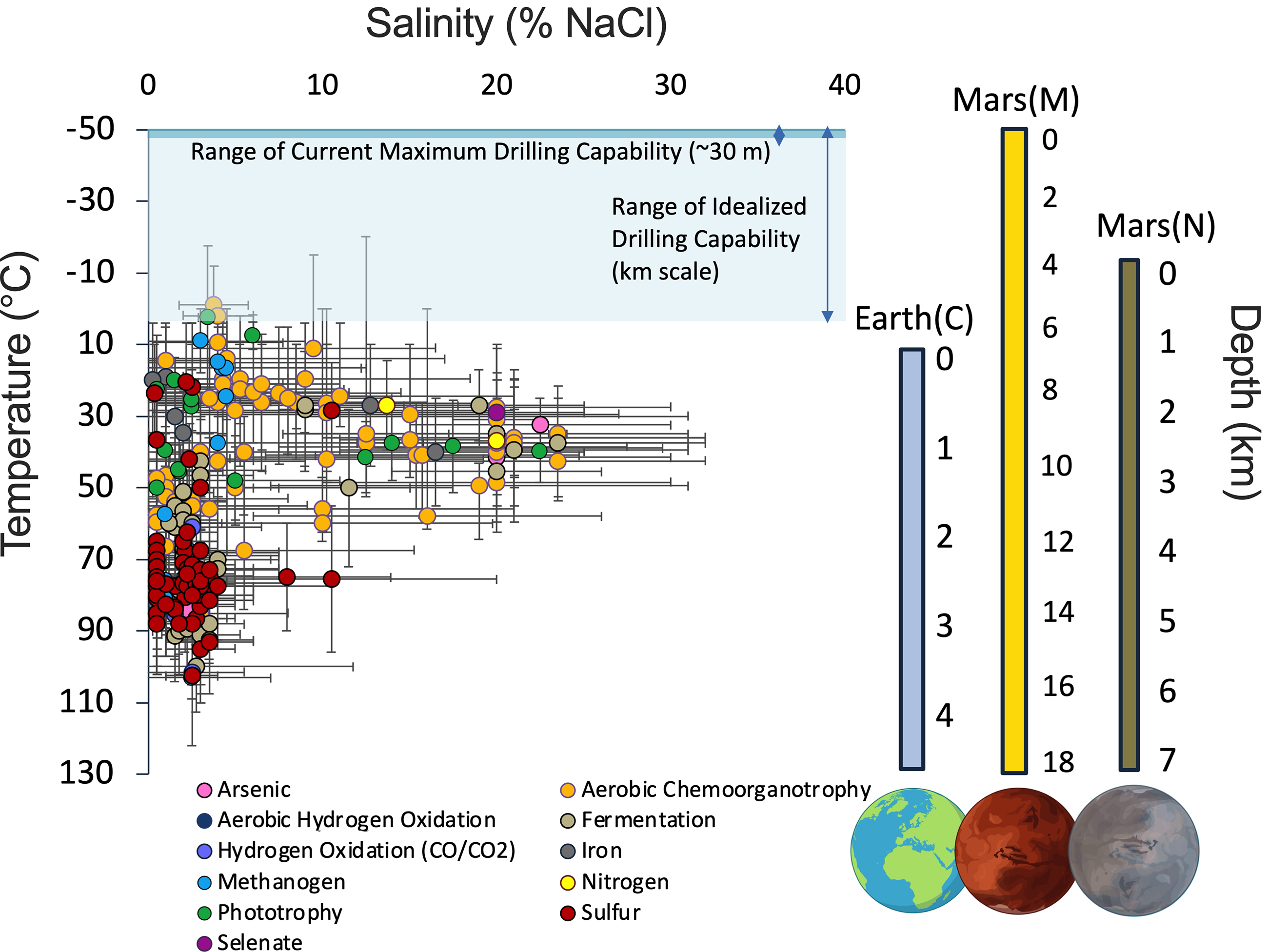
Distribution of microbial extremophiles identified across temperature and salinity (% NaCl) constraints on Earth and grouped by primary energy metabolism. Error bars represent the minimum and maximum of temperature or salinity for the organism’s defined growth range or survival range, where available. Organisms utilizing putative martian metabolisms identified in this study are highlighted, including methanogenesis and sulfur and iron guilds. Geothermal gradients for Earth’s continental crust (C), modern Mars (M), and Noachian Mars (N) from Michalski et al. (2013,2018) are presented to the right for comparison. Depth ranges for current proposed drilling technologies relative to idealized depths for modern Mars are shown with dark and blue highlighted boxes, respectively. Data presented here and associated references are included in Supplementary Data.
Recommended regions to target in the martian subsurface—reconciling habitability and practicality
As we move toward the future of life detection efforts since the original Viking mission, it becomes essential not only that we target specific metabolic activity but also that we design our experimentation with feasibility and accessibility in mind. However, this requires acknowledging a clear discrepancy between depths we can practically access given current drilling technologies and idealized depths we could target for the best chance to find life as we know it within temperature space (Fig. 2). For instance, of the current proposed missions (e.g., Mars Life Explorer [MLE] and ExoMars Rosalind Franklin Rover) that target the martian subsurface, drilling capabilities are often projected to access meter-scale depths at most (Williams and Muirhead, 2021; Da Poian et al., 2022; Phillips‐Lander, 2024), with no technologies needed to reach kilometer depths presently existing (National Academies of Sciences, Engineering, and Medicine, 2025). While concepts of drilling technologies to extend further into the crust (e.g., rotary-percussive based drill systems potentially reaching down to 100 m) are being matured (Tosi et al., 2026a), the ability to reach kilometer-scale depths remains heavily constrained by the required complexity, mass, and/or power of these systems (Tosi et al., 2026b). This poses the essential question: Does our current knowledge of microbial habitability (presented in this review) align with limitations of future drilling missions? Given the constraints of current and near-future mission capabilities, we propose focusing exploration on the shallow subsurface, extending to depths of up to 30 m, as this represents a more realistic target in light of current plans for subsurface exploration on Mars (Altieri et al., 2023; National Academies of Sciences, Engineering, and Medicine, 2023). This depth, while not overlapping with survival distribution ranges of chemotrophic microbes on Earth (Fig. 2), could still be valuable to investigate potential habitable microenvironments or regions where extinct life may have existed in a more favorable Noachian geothermal gradient. Considering the context of this proposed depth and geothermal gradients for Mars, we additionally propose future targeting of specific chemolithotrophic metabolisms, including (at the least) methanogenesis, sulfur utilization, and aerobic heterotrophy, that could be complemented, subsequently, by other anaerobic heterotrophic or autotrophic strategies that typically persist at higher temperatures and lower salinities (Fig. 2).
Considerations for planetary protection and a future human presence on Mars
In the interest of detecting truly indigenous extant life on Mars, it is essential we acknowledge challenges that may arise with planetary protection. Traditionally, planetary protection guidelines have maintained our best efforts to encourage the sterility of robotic explorers sent to destinations within our solar system (Smith et al., 2017; Coustenis et al., 2023). However, robotic missions still largely avoid “special regions” as defined by the Committee on Space Research, even though these locales are considered the most likely to harbor conditions conducive to extant life, at ≥−28°C and water activity ≥0.5 (Kminek et al., 2010; Olsson-Francis et al., 2023). This is because best efforts to fully sterilize spacecraft are often incomplete, and the surface of Mars may not completely deactivate transplanted Earth microorganisms (Kerney and Schuerger, 2011; Hallsworth, 2021). Fortunately, field and laboratory studies on Earth have demonstrated that the bioburden of a human mission to Mars may not be overtly hindering to life detection efforts (Schuerger and Lee, 2015). Nevertheless, as the human element is introduced to regions on Mars where we reasonably believe life may exist, it will be necessary not only to protect the pristine environments on Mars but also to protect the human explorers themselves. If extraterrestrial life is discovered on Mars, it will initially be treated as hazardous, similar to the treatment of deadly pathogens on Earth until we prove otherwise (Craven et al., 2021; Carrier et al., 2022).
The question becomes not solely “How do we protect Earth and Mars?” but “How do we safely perform microbiology on Mars?” Specifically, in high-containment laboratories on Earth, a primary concern when working with dangerous pathogens is aerosolization, which may lead to a loss of containment and human infection (Ravnholdt et al., 2025). Astrobiodefense efforts will need to establish new procedures and protocols for biosafety on Mars, where the ambient low pressure on the surface (Hourdin et al., 1993; Withers, 2012) may promote rapid boiling, evaporation, and aerosol generation from liquid-containing samples taken by astronauts from the shallow subsurface, posing a potential immediate biosafety hazard. Moreover, attempts to culture extant life would occur under medium- to high-vacuum conditions, which may also cause the aerosolization of martian microorganisms (Schwendner and Schuerger, 2020). Astrobiodefense guidelines should focus on developing new biosafety procedures for human operations on Mars, enabling life detection experiments while ensuring the safety of both Mars astronauts and life back on Earth upon their return. High-containment experiments involving extant martian life, or samples reasonably thought to contain martian biology, may be best suited for operations on Mars or in high containment facilities housed on the Moon (Moxley and Ricciardi, 2026). Such measures may be ultimately necessary to minimize biological risk and maintain public trust in this new era of space exploration (Bimm et al., 2025).
Future recommendations in the context of proposed life detection efforts
The larger Astrobiology community has seen the development of several key strategies working to refine the best recommendations for life detection since the original Viking mission. Some of the first contexts came from the goals of the Mars Exploration Program Analysis Group (MEPAG), seeking to define habitability of martian environments throughout the planet’s history while searching for signs of past or present life following evidence for liquid water (MEPAG Next Decade Science Analysis Group, 2008). More recently, the 2023–2032 Decadal Survey (“Origins, World, and Life”) offers an increased emphasis on the detection of extant life, with a strong recommendation for using multiple independent lines of evidence, such as independent biosignatures of metabolism in agreement with other geochemical, isotopic, morphological, and/or spatial analyses (National Academies of Sciences, Engineering, and Medicine, 2019; National Academies of Sciences, Engineering, and Medicine, 2023). Groups aimed at refining these goals include the Mars Extant Life Conference Group (Carrier et al., 2020), which recommended best target locations as “refugia” environments (e.g., caves, deep subsurface, ices, and salts) with a focus on geologically informed biosignature detection. Furthermore, current efforts of the Search for Life Science Analysis Group have developed preliminary recommendations to target (1) the presence of cell-like structures, (2) observation of cellular processes, and (3) the presence of chemical compounds indicative of biosignatures of life, specifically in the context of dynamic fluid and thermal environments within martian mid-latitude ice sheets (Carrier and Scharf, 2025).
Generally, these groups have similar major recommendations to target subsurface ice and detection of extant metabolic activity through multiple detection/biosignature approaches, while implementing agnostic analytical techniques as best possible. These have helped inform the current Mars Life Explorer (MLE) mission concept, which remains the National Aeronautics and Space Administration’s primary recommended drilling-based mission to explore the shallow subsurface of Mars, for detection of diverse organic, geochemical, and morphological biosignatures within an agnostic framework (Williams and Muirhead, 2021). Finally, goals for human exploration have been shaped by the more recent Human Exploration Science Strategy and the proposed Moon to Mars framework (Goodliff et al., 2023; National Academies of Sciences, Engineering, and Medicine, 2025). These strategies emphasize a science-driven approach to Mars exploration while prioritizing the development of off-planet architecture and robotic partnerships to facilitate human efforts and potentially preserve some sterility (a particular concern when/if considering access to special regions). This includes leveraging the Moon itself as an initial testbed for human science and exploration under challenging low-gravity and atmospheric conditions, which will also be encountered on Mars (albeit to different degrees).
Considering these current recommendations in the field and the knowledge we have gained since Viking with regard to terrestrial subsurface microbial life and metabolism, alongside increased characterization of Mars’ surface and subsurface conditions, we recommend the following:
The subsurface of Mars should be the primary target for future life detection efforts, with a goal to access as great a depth, and as close to special regions, as possible.
The deepest feasible depth given current proposed drilling technologies of up to ∼30 m should still be targeted despite being outside the range of known terrestrial chemotrophic metabolism (Fig. 2), while continuing to invest and develop deep drill technologies toward kilometer-scale reaching beyond the cryosphere. Even shallow subsurface investigations could reveal niche, refuge environments as previously emphasized by the Mars Extant Life Group. Detection of extant life through metabolic activity measurements should target chemotrophic redox metabolism and lower porosity regions that may host fracture systems enriched in useful redox/organics species.
Specific targets should include methanogenesis, sulfur metabolism, iron metabolism, and aerobic metabolism, respectively. Life detection efforts should be designed under the expectation that brine-hosted, slow-growing microorganisms are the most likely extant target on current Mars.
Subsurface life on Earth often exists in low-biomass, halophilic systems where turnover and energy flux are limited due to increased investment in stress defense. Metabolic turnover may be exceedingly slow under multiple potential sources of environmental stress on Mars. Implementation measures for planetary protection and human protection must be prioritized, with potential sampling measures employed on the Moon in practice for future Mars missions.
As we move toward future human exploration, the development of planetary protection and biosafety protocols is essential. These could be improved with preliminary implementation in low gravity and low atmospheric conditions, potentially leveraging proposed Moon exploration as an initial testbed.
The idealized version of any scientific endeavor is rarely the eventual reality. As astrobiologists, our primary focus is on the search for life beyond Earth. However, as with the Apollo missions to the Moon, the primary motivation for a human mission to Mars may not always include the same objectives and considerations as life detection efforts. The Astrobiology community will need to balance the desire for pristine science in extraterrestrial environments with the realities of an eventual human presence. In the face of imminent private missions to Mars and terraforming efforts (Coleine et al., 2025; DeBenedictis et al., 2025; Gaviraghi, 2025), the Astrobiology community will need to act expeditiously to define planetary protection and contamination protocols before these missions destroy any extant life. It is imperative that, while astrobiologists continue to push for preservation of planetary and moon settings that may hold the greatest potential for life detection efforts, the community also prepares for colonization and the potential contamination that may follow. The sheer diversity of life on Earth, including the unique and expansive variety of microbial metabolisms possible on our planet, makes it difficult to dismiss the possibility of extant life on Mars.
Authors’ Contributions
D.M.N.: Conceptualization (equal with R.L.M.), data curation (equal with R.L.M.), supervision (lead), visualization (lead), writing—original draft (equal), and writing—review and editing (equal). R.L.M.: Conceptualization (equal with D.M.N.), data curation (equal with D.M.N.), supervision (supporting), visualization (supporting), writing—original draft (equal), and writing—review and editing (equal). A.R. and K.G.L.: Conceptualization (supporting), data curation (supporting), supervision (supporting), visualization (supporting), writing—original draft (equal), and writing—review and editing (equal).
Footnotes
Acknowledgments
This work has greatly benefited not only from the discussions and intellectual contributions of the ∼30 participants in this Viking special issue but also from researchers across global institutions that have contributed to the numerous advancements in microbial subsurface research over the decades since Viking. This review is dedicated to highlighting and continuing the appreciation of the original Viking experiments and future life detection efforts shared by the larger Astrobiology community.
Author Disclosure Statement
The authors report no conflicts of interest with this work.
Funding Information
D.M.N.’s work on this paper was supported by an appointment to the NASA Postdoctoral Program at Ames Research Center, administered by Oak Ridge Associated Universities under contract with NASA. K.G.L. acknowledges NSF grants OCE-1948720; OCE-2448137, and OCE-2152551. Additionally, A.R. acknowledges this material is based upon work supported by the National Science Foundation Graduate Research Fellowship Program under Grant No DGE-2236414. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.