
Abstract

We are searching for life in many places with many techniques. Beyond the solar system, transit spectroscopy may find exoplanet atmospheres produced by life (Schwieterman et al., 2018). Radio or optical Search for Extraterrestrial Intelligence (SETI) may succeed. Maybe we’ll find technosignatures: Dyson spheres or industrial atmospheric pollution. On Mars we’re chasing down methane, “leopard spots,” and microbially induced sedimentary structures. We’re looking for organic molecules on Titan and in gas-phase jets from Europa and Enceladus. Trying to understand the origins of life on Earth, we analyze carbon 12/13 ratios and fossil stromatolites. We’re using metagenomics to search for ever-more extreme extremophiles and for ever-more deeply rooted branches in the phylogenetic tree of life. We have analyzed hundreds of meteorites and material from carbonaceous asteroids Bennu and Ryugu. Such searches tend to look for specific molecules: Amino acids, nucleobases, and lipids. Yet, fundamental statistical properties of the chemical organization of life may be more diagnostic than the contingent details of our biomolecules (McKay, 2004). Measures of molecular ecology or complexity or diversity have been suggested to identify life (Sharma et al., 2023; Saeedi et al., 2025; Yoffe et al., 2026). Identifying such biosignatures in small, contaminated multibillion-year-old samples is a tough row to hoe. Thus, any new technique is welcome that can help hoe rows.
Enter a new tool—Pareto Optimality (Rodgers et al., 2026). Vilfredo Pareto was a 19th-century Italian economist concerned with inequality. In economics Pareto optimality is a state in which no one person can be made better off without making someone else worse off. To apply this to engineering, no-worse-off people are replaced with no-worse-off task performances. Pareto optimal solutions (also known as the Pareto front) are solutions that cannot improve the performance at any task without decreasing the performance at some other task. This process is also called multiobjective optimization and has proven useful in economics and engineering. Can it be useful in biology? In particular, can it be useful in astrobiological contexts to identify life? That is the question Rodgers et al. (2026, this issue) ask, “Can Pareto Optimality Be Evidence of Life?”
For 4 billion years life on Earth has been designed by selection pressures pushing and pulling in different directions to perform multiple tasks. Rodgers et al. argue that these selection pressures and fundamental physics (Cockell, 2018) produce designs that simultaneously function optimally—in the sense of performing as well as they can at any given task without decreasing the performance at any other tasks. They argue that these optimizations could describe life elsewhere in the universe and, therefore, Pareto optimality could be a useful test for biogenicity. I’m not sure nature cares about inequality or no-worse-offness as much as Pareto did. Doesn’t evolution improve fitness by increasing the performance of an important task even if it decreases the performance of a very minor task? Wouldn’t such cases violate Pareto optimality? (Fig. 1).
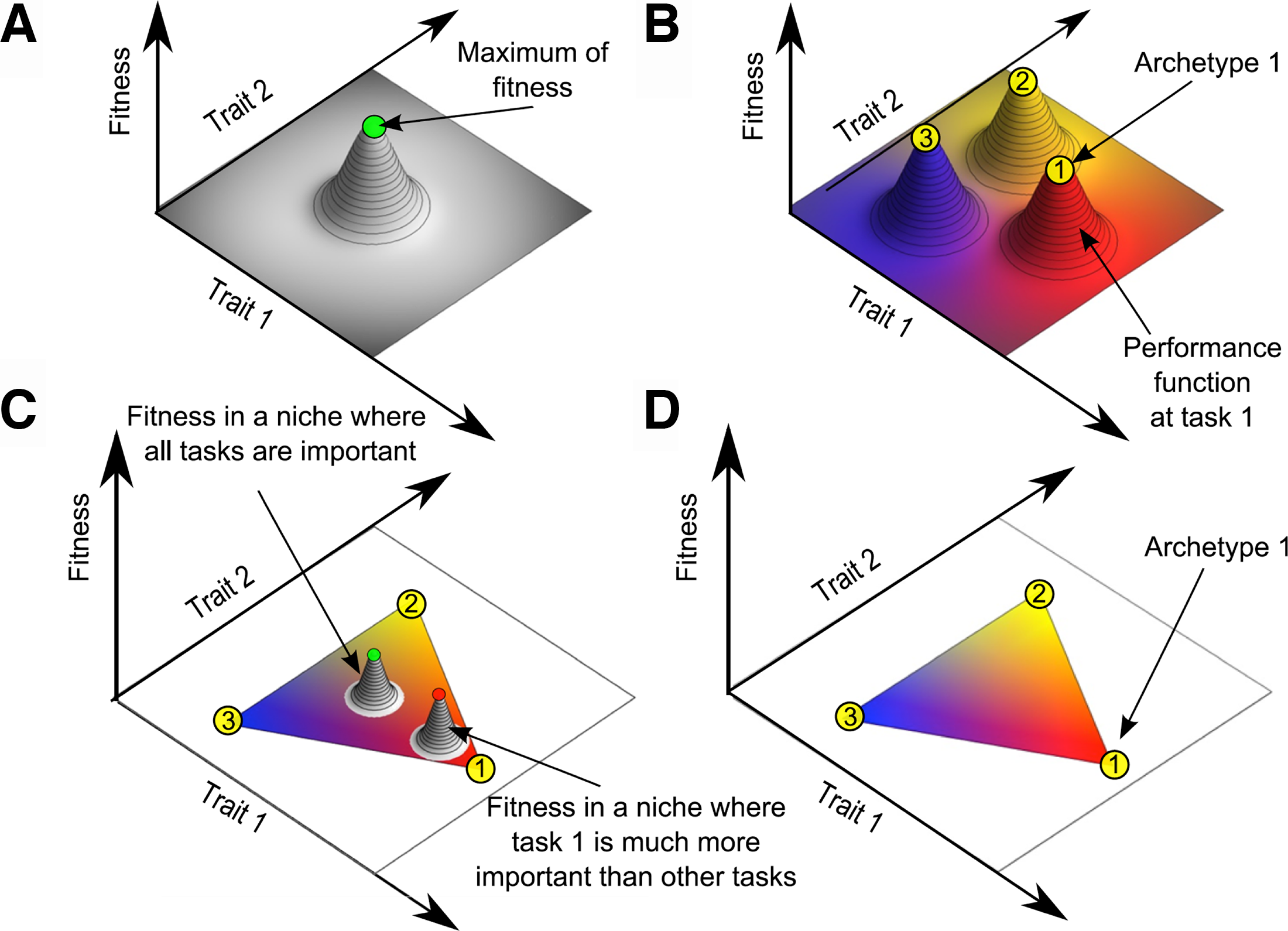
The difference between a conventional adaptive landscape
The figure helps explain how Pareto optimality is different from the conventional fitness peaks of an adaptive landscape (Wright, 1932). Pareto optimality may be a useful concept for engineering and economics where the performance functions are well-defined, but assuming such well-defined performance functions in biological evolution (especially in the context of astrobiology) is much more tenuous (as Rodgers et al. point out). If one makes good guesses about the important, dominant tasks, the Pareto front can be usefully small. But with a large number N of tasks (and multiple ways of performing those tasks), the Pareto front would be enormously N-dimensional. Optimization using hundreds of unimportant, unprioritized tasks will produce a uselessly large Pareto front.
The most important biological tasks can be difficult to identify. Is efficient motility an important task? What is “efficient motility?” Are salmon swimming upstream against a strong current moving efficiently? There seem to be as many ways of moving efficiently in an environment as there are environments to move in: cheetahs, snakes, tuna, jellyfish, impalas, sloths, turtles, moths, bats, unikonts, bikonts, and Pelagibacter ubique. Despite this largely eukaryotic variety, the most fundamental prokaryotic tasks are the ones most likely to be shared by life elsewhere and, therefore, the most likely to be useful in astrobiological Pareto optimization.
When the task or the function to be performed is a given (as defined by economists or engineers), Pareto optimality makes sense. But what if the task is constantly cycling between day and night, cold and hot, wet and dry seasons, times of food scarcity and times of food abundance. Over durations from minutes to billions of years the tasks needed to stay alive change and keep changing.
Economists and engineers can work from a blank slate. Engineers can crumple up an original design, go back to the drawing board, and come up with a newer, better plan. Evolution cannot. Natural selection cannot jettison the accumulated baggage of its evolutionary history. The performance of new tasks is often (or always?) burdened by previous tasks. The blind spot in each of our eyes is produced by the quirky history of our inside-out retinas. Other examples are wisdom teeth, appendices, and the recurrent laryngeal nerve in giraffes. Recent optimizations will always be blurred by the remnant baggage of billions of years of previous optimizations. Like a palimpsest, overwritten again and again, the obsolete optimizations are always embedded in a life form. If optimal designs are signs of biogenicity, so too are suboptimal designs. Such inherited bad designs can undermine assumptions about optimization.
If evolution is predominantly contingent and path-dependent (Gould, 2002), evolved structures will be dominated by billions of years of previous obsolete optimizations. If the dominant selection forces on life on Earth are the quirky self-referentiality of other life forms, then evolution will be highly contingent. Path-dependent baggage and quirky contingency may extend back to the origins of life and the particular accidents of molecular evolution and the choice of biomolecules. Such fundamental unpredictability undermines the ability to identify tasks and apply Pareto optimization.
Alternatively, if evolution is predominantly convergent (Conway Morris, 2003), functional tasks would be more stable and recognizable based on terrestrial analogs. Universal physics and the general similarities between the chemistries of many billions of wet rocky Earth-like planets may converge on a universal Pareto front of amino acids and nucleotides. If so, Pareto optimality could be a useful astrobiological tool.
Many hours are being spent trying to identify precisely what counts as a biosignature. These efforts are redolent of our efforts to define life (e.g., Benner, 2010). Are viruses alive? Are transposons alive? Is the biosphere alive? We keep thinking we know what life is and we keep trying to define it. However, behind any black-or-white test of biogenicity (Is it life or not?) lurks what Dawkins (2011) has called the tyranny of the discontinuous mind. We may be imposing a strict, artificial boundary onto a continuum in which there is no identifiable line between life, proto-life, proto–proto-life, molecular evolution, and the quirks of nonequilibrium chemistry at hydrothermal vents or warm tidal pools. This continuum may also be N-dimensional. The search continues.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Associate Editor: Sherry L. Cady