
Abstract
Negative-pressure wound therapy (NPWT) is one of the most advanced therapeutic methods in the treatment of various hard-to-heal acute and refractory chronic wounds. Recent emerging evidence points to a role of the microRNA-mediated regulation of angiogenesis in ischemic tissues, and a series of microRNAs associated with angiogenesis have been successively identified. In this study, we found that miR-195 expression was significantly upregulated and the microvessel density (MVD) was increased in granulation tissue collected 7 days after NPWT compared with those in the pre-NPWT tissue. Moreover, the expression of NLRX1, the potential target gene of miR-195, was down-regulated in post-NPWT compared with that in pre-NPWT tissue. Significant negative correlations were detected between miR-195 and NLRX1 expression levels (r = −.856, P < .001) and between NLRX1 expression and MVD (r = −.618, P < .05), whereas miR-195 expression was positively correlated with MVD in the granulation tissue (r = .630, P < .05). In summary, NPWT may suppress NLRX1 expression through the upregulation of miR-195 expression, thus efficaciously promoting angiogenesis in the granulation tissue to enhance wound healing.
The past 2 decades have witnessed significant developments in novel treatments for hard-to-heal acute and chronic wounds, such as negative-pressure wound therapy (NPWT) and silver dressings. NPWT is a process in which a wound is covered with a vacuum (air-tight) dressing to which negative pressure is applied. It has become a widely used and efficacious treatment, which has been demonstrated to initiate a series of biological effects on a closed surgical incision, reduce the wound area, promote healing, relieve edema, and stimulate angiogenesis in the granulation tissue.1-3 These combined effects of NPWT may contribute to faster and stronger healing with a decreased risk of infection of hard-to-heal wounds.
Angiogenesis refers to the process by which an organism forms new capillaries from preexisting microvessels. 4 Mounting evidence5,6 has shown that angiogenesis promotes physiological processes such as embryonic development, endometrial tissue revascularization, and tissue repair. In addition, angiogenesis plays a key role in hypertrophic scar formation, malignant tumor growth, diabetic peripheral neuropathy, and the initiation and progression of other diseases.
MicroRNAs (miRNAs) are small noncoding RNAs, 19 to 25 nucleotides long, that are known to regulate several protein-coding genes in both plants and animals. Through perfect or imperfect complementary base-pairing with the 3′ untranslated region of target mRNAs, miRNAs can cause target mRNA degradation or translation suppression, thus regulating the expression of the target genes. This subsequently affects cell proliferation, differentiation, migration, and apoptosis, contributing to the processes of angiogenesis, 7 wound repair, or tumor growth and metastasis. Recent studies have indicated that miRNAs can regulate angiogenesis through regulation of the expression of vascular endothelial growth factor (VEGF), 8 heparin-binding growth factor, 9 and matrix metalloproteinases, and are capable of influencing various cellular processes such as cell growth and proliferation by regulating signaling pathways such as the RPS6KB1/HIF-1a, 10 PIK3C2α/Akt/HIF-1α, 11 and TGF-β/SMAD pathways. 12 NOD-like receptor (NLR) family member X1 (NLRX1) is a recently discovered member of the NLR family, which is localized to the mitochondria, and has been validated to exert multiple functions, including the regulation of immune and antiviral responses. 13 Accumulating evidence suggests that NLRX1 can induce changes in the production of reactive oxygen species in living cells, which is relevant for many diseases. 14 For example, in arthritis, the production of NOX-induced reactive oxygen species promotes endothelial cell migration, angiogenic tube formation, and secretion of several proangiogenic mediators. 15
However, the functions of miRNAs and NLRX1 in angiogenesis and the underlying mechanism remain unknown. In our previous study, we found that NLRX1 was closely linked to angiogenesis and emerged as a potential target gene of the miRNA miR-195 through prediction with TargetScan, MiRanda, and PicTar. To better understand this angiogenesis regulation mechanism and the links to the healing effects of NPWT, in the present study, we evaluated the expression levels of miR-195 and NLRX1, and the change in the microvessel density (MVD) in the granulation tissue of patients collected before and after NPWT. We further investigated the association of miR-195 and NLRX1 expression with wound angiogenesis and the correlations among the 3 variables.
Material and Methods
Tissue Samples
The set of tissue samples was collected from 6 patients with refractory wounds for at least 4 weeks at the Third Xiangya Hospital of Central South University from March 2014 to March 2015. All samples were handled according to the ethical and legal standards and approved by the Research Ethics Committee of Affiliated Cancer Hospital of Xiangya School of Medicine. All patients provided informed consent prior to surgery.
Systemic Therapy
In view of the poor local blood supply and undernourished skin in the refractory wounds, as well as poor bodily functions and comorbidities that most of the patients experienced, adjuvant therapies, including nutrition improvement, anemia correction, and blood glucose control, were administered preoperatively.
Administration of NPWT
Following routine debridement to remove the necrotic tissue and disinfection of the periwound area, an NPWT device (V.A.C. negative pressure suction device, KCI, San Antonio, TX) was used for the application of negative pressure. The V.A.C. dressing was cut so as to suit the dimensions and shape of each wound, and then pieced together. For deep wounds and sinus tracts, the V.A.C. dressing was placed onto an internal drainage tube, whereas for shallow or flat wounds, the V.A.C. dressing was placed on an external drainage tube. During the procedure, the adequacy of membrane sealing was assessed, and effective negative pressure was indicated by the collapse of the V.A.C. dressing and appearance of the drainage tube in the dressing. The vacuum was generally maintained at 75 to 125 mm Hg, and intermittent or continuous suction was adopted according to the dimensions and position of the wounds.
Experimental Grouping and Specimen Preparation
The wound color, exudation, secretion and margins, and the periwound skin of patients were observed and recorded before redressing at the first day before NPWT (pre-NPWT group, hereinafter referred to as the Control group) and the seventh day after NPWT (NPWT group), respectively. Subsequently, an approximately 1 × 1 × 1 cm3 full-thickness skin section was cut from the wound margin of each patient using tissue scissors. On collection of tissue specimens, the patients and their family members were notified, and informed consent forms were signed. Each tissue block was divided into 2 equal portions. For the first portion, the epidermis and adipose tissues were removed, and then the tissue was divided into 100-mg blocks, frozen at −80°C, and then transferred to a liquid nitrogen refrigerator for storage until use in reverse transcription-polymerase chain reaction (RT-PCR) and western blot analyses. The other portion was fixed in formaldehyde, embedded in paraffin, and sectioned for use in hematoxylin-eosin staining and immunohistochemistry (IHC).
Determination of miR-195 and NLRX1 mRNA Expression Levels by RT-PCR
Total RNA from each tissue specimen was extracted using the TRIzol method and then reverse-transcribed into cDNA for use as templates for PCR. The procedure was carried out in accordance with the instruction manual of the TaKaRa RT-PCR Kit (TaKaRa Bio Inc, Tokyo, Japan), using the following amplification program: initial denaturation for 5 minutes at 94°C, 30 cycles of denaturation for 30 seconds at 94°C, annealing for 30 seconds at 60°C, and extension for 30 seconds at 72°C, and a final extension for 6 minutes at 72°C. Quantitative fluorescence PCR analysis was then directly performed on the synthesized cDNA solutions using the following primers: miR-195 sense 5′-CGTAGCAGCACAGAAAT-3′, antisense 5′-GTGCAGGGTCCGAGGT-3′; U6 small nuclear RNA (endogenous control for miR-195) sense 5′-CTCGCTTCGGCAGCACA-3′, antisense 5′-AACGCTTCACGAATTTGCGT-3′; NLRX1 sense 5′-GGCATCAGGACGGAGGAGGAGT-3′, antisense 5′-GGGCACGGTGAACACGAAGG-3′; and β-actin (endogenous control for NLRX1) sense 5′-CATCCTGCGTCTGGACCTGG-3′, antisense 5′-TAATGTCACGCACGATTTCC-3′. The experiments were repeated thrice, and the 2−ΔΔCT method was used to calculate the relative expression levels of miR-195 and NLRX1 from the experimental data.
Determination of NLRX1 Protein Expression Levels in Tissue Samples With Western Blot
The protein of the tissue samples was extracted with RIPA cell lysis buffer according to the manufacturer’s instructions (Shanghai Bioeasy Biotechnology Co Ltd, Shanghai, China). A BCA assay kit (Beijing Zhongshan Golden Bridge Biotechnology Co Ltd, Beijing, China) was used to determine the protein concentration. The Bio-Rad Gel Imaging System was used for image acquisition, and Quantity One Image Analysis software was used to analyze the NLRX1 (Semmerfeld Technology Co Ltd, China) and β-actin (endogenous control) protein bands. NLRX1 expression was calculated as the relative expression level to that of β-actin (NLRX1/β-actin).
Determination of NLRX1 Expression and MVD in Tissue Samples With IHC
Samples were fixed in 4% formaldehyde, embedded in paraffin, and serially sectioned into 4-µm sections. IHC testing was performed with rabbit anti-human NLRX1 and mouse anti-human CD31 antibodies (Santa Cruz Biotechnology, Santa Cruz, CA), followed by horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G (Beijing Zhongshan Golden Bridge Biotechnology Co Ltd,) secondary antibody, using the EliVision method in strict accordance with the instruction manual of the assay kit (Fuzhou Maixin Biotech Co Ltd, Fuzhou, China). The specimens were stained with diaminobenzidine, counterstained with hematoxylin, washed with distilled water, restored to blue color with phosphate-buffered saline, sealed, and examined under a microscope. Images from the microscope were captured as normal computer images. Determination of count data from the results for all indicators was performed using a double-blind method by 2 senior pathologists. Observations were made at 400× magnification, and results for observation indicators were recorded. Five high-power fields were examined for each section, and paired double-blind counting was performed.
NLRX1 is expressed primarily in the cell cytoplasm, and positively stained cells exhibit a yellow brown color. The positive-stained cell proportion was scored as follows: 0 (⩽5% positive cells), 1 (6% to 25% positive cells), 2 (26% to 50% positive cells), and 3 (>51% positive cells). 14 Staining intensity was graded according to the following criteria: 0 (colorless), 1 (light yellow), 2 (yellow brown), and 3 (brown). The products of the aforementioned 2 scores were calculated and classified as (−) (product ⩽3) and (+) (product ⩾4).
Determination of the MVD was performed according to the method reported by Weidner et al. 15 The yellow-/brown-stained vascular endothelial cells indicated positive expression of CD31. Each yellow-/brown-stained endothelial cell or endothelial cell cluster was counted as 1 microvessel, and each branch structure was also counted as 1 microvessel as long as the structures were not connected. For each section, examination was performed under low power (100×), and the region with the highest MVD was selected. Subsequently, microvessel counts were performed in 5 high-power fields (400×), and the mean value was considered as the MVD for that section.
Statistical Analysis
Statistical analysis was performed using SPSS 16.0. The paired t test was used for statistical analysis, with P < .05 indicating statistically significant differences. Associations between variables were tested using the nonparametric Spearman’s rank correlation coefficient.
Results
Gross Observations of Wounds and Wound Healing Status
Before treatment, there was little granulation tissue detected in the wound beds. The wounds showed a relatively darker appearance accompanied by necrotic tissue. A significant amount of purulent discharge was present, and swelling of the periwound tissue was prominent. At 7 days after NPWT, there was an increased amount of granulation tissue observed in the wounds, which had a finely granular, bright red appearance and was prone to bleeding with touch. No necrotic tissue or purulent discharge was observed in the wounds, the wound sizes were reduced, and swelling of the periwound tissue was significantly alleviated (Table 1). In particular, the wounds of patients 1 and 2 had completely healed after receiving post-NPWT autologous skin grafts. For patients 3 and 4, infections were significantly alleviated after NPWT, and the wounds had healed without festering within 2 weeks after flap transplantation. For patients 5 and 6, the infections were also significantly alleviated after NPWT, and the wounds had healed without tibial bone exposure after flap transplantation and autologous skin grafting.
Baseline Characteristics of the Patients.
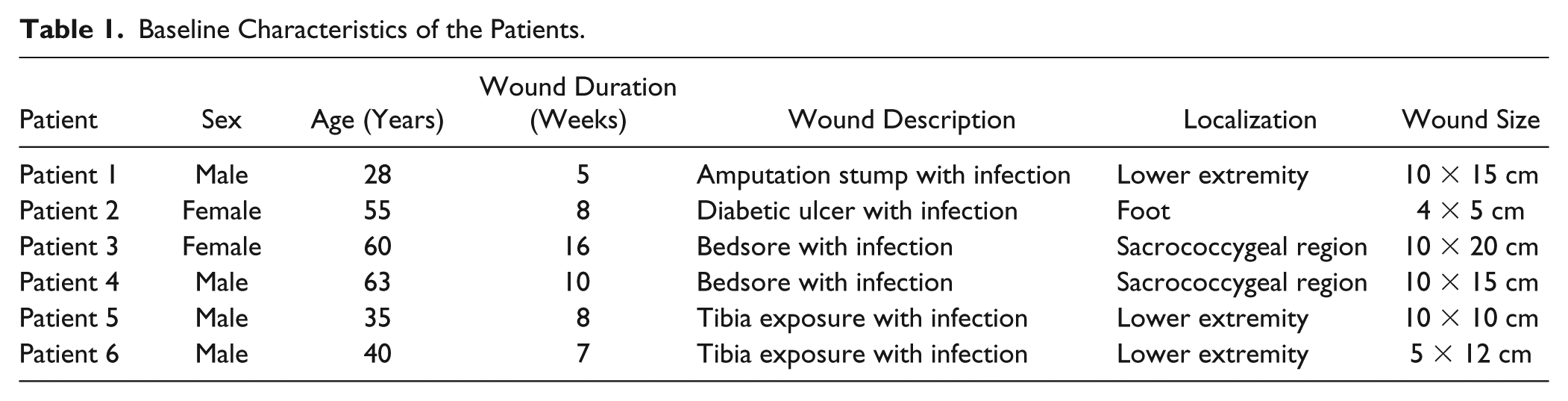
miR-195 Is Upregulated and NLRX1 Is Downregulated in the Granulation Tissue After NPWT
The RT-PCR assay results demonstrated that miR-195 was expressed at low levels in the granulation tissue before NPWT and then significantly increased after NPWT (t = 5.898, P < .001; Figure 1), indicating that NPWT might promote miR-195 expression in wounds.
Relative expression levels of miR-195 (a) and NLRX1 mRNA (b) in wound biopsies from patients in the NPWT (negative-pressure wound therapy) and control groups. ***P < .001.
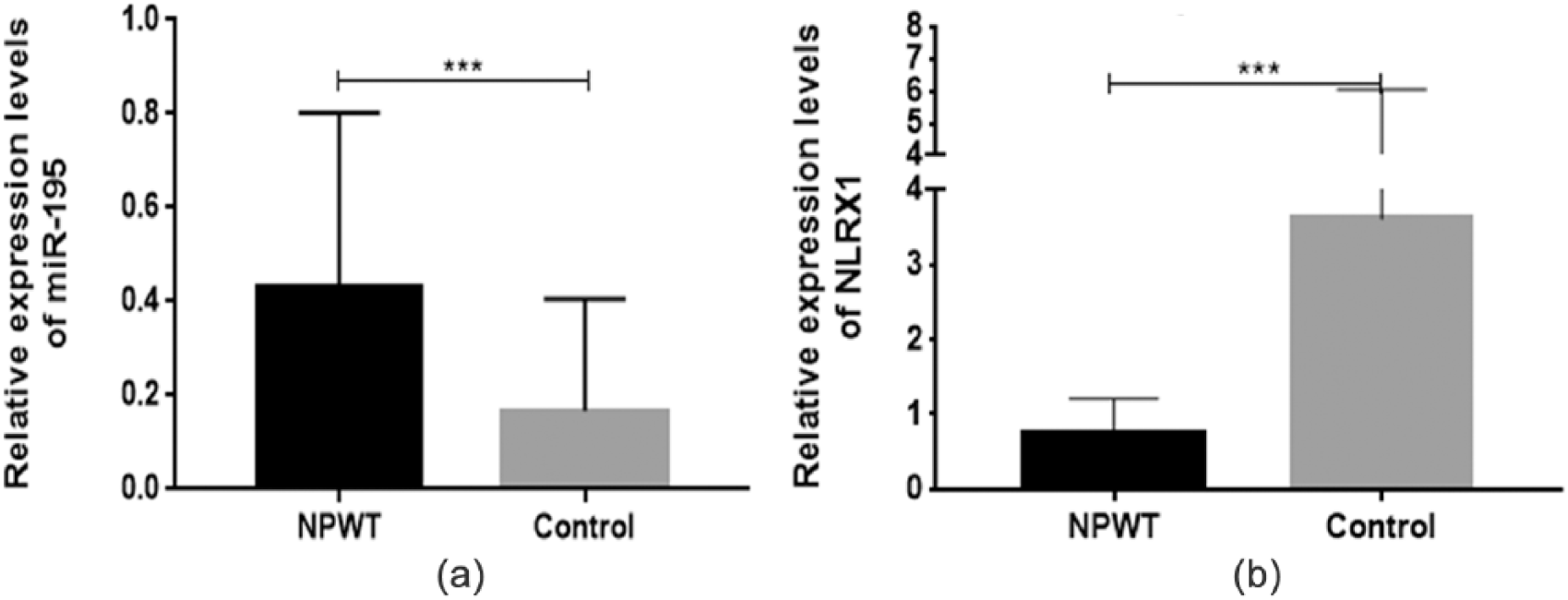
By contrast, NLRX1 mRNA was expressed at higher levels in the granulation tissue before NPWT and was significantly decreased after NPWT (t = 5.235, P < .001; Figure 2). Similarly, western blotting analysis showed that the expression level of NLRX1 protein in the granulation tissue after NPWT were significantly lower than the pre-NPWT levels (P < .05; Figure 2). These results indicated that NPWT may have a suppressive effect on NLRX1 expression in wounds.
Western blot analysis of NLRX1 protein expression in wound biopsies from patients in the NPWT (negative-pressure wound therapy) and control groups. Data are expressed as mean ± standard deviation. *P < .05.
IHC Results
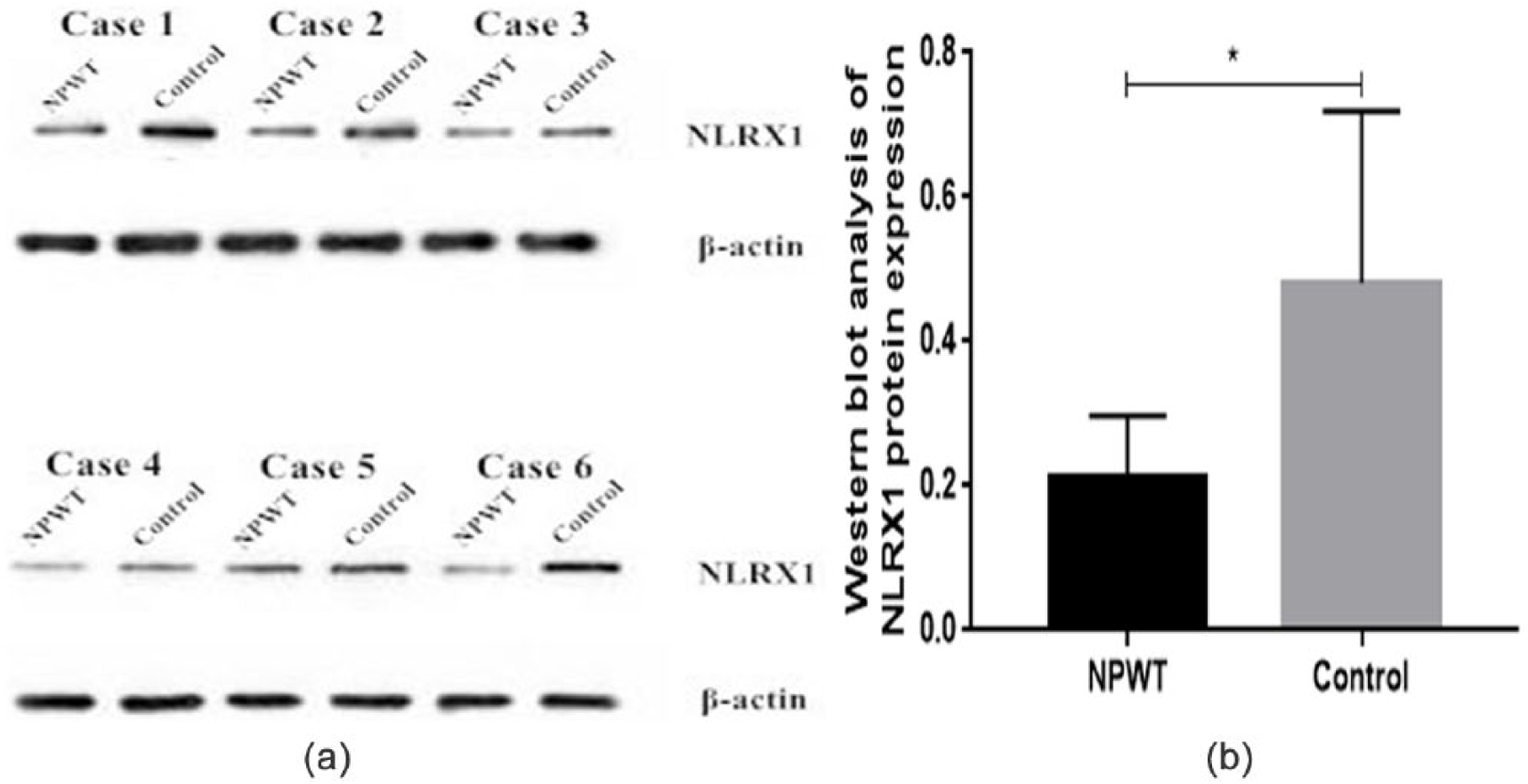
In the granulation tissue, NLRX1 is primarily expressed in the cytoplasm, and positive-stained cells exhibit a yellow/brown color. NLRX1 is also primarily expressed in the cytoplasm of normal tissue; however, the positively stained cells exhibit a lighter color, and some cells appear colorless. All 6 patients showed a (+) score of NLRX1 IHC before NPWT, whereas only 1 patient (16.67%) had an overall (+) score after NPWT. Indeed, the expression levels of NLRX1 in the granulation tissue were significantly lower (P < .05) compared with the pre-NPWT expression levels (Figure 3).
Immunohistochemical assessment of NLRX1 expression in wound biopsies from patients in the NPWT (negative-pressure wound therapy) and control groups (magnification, 100×).
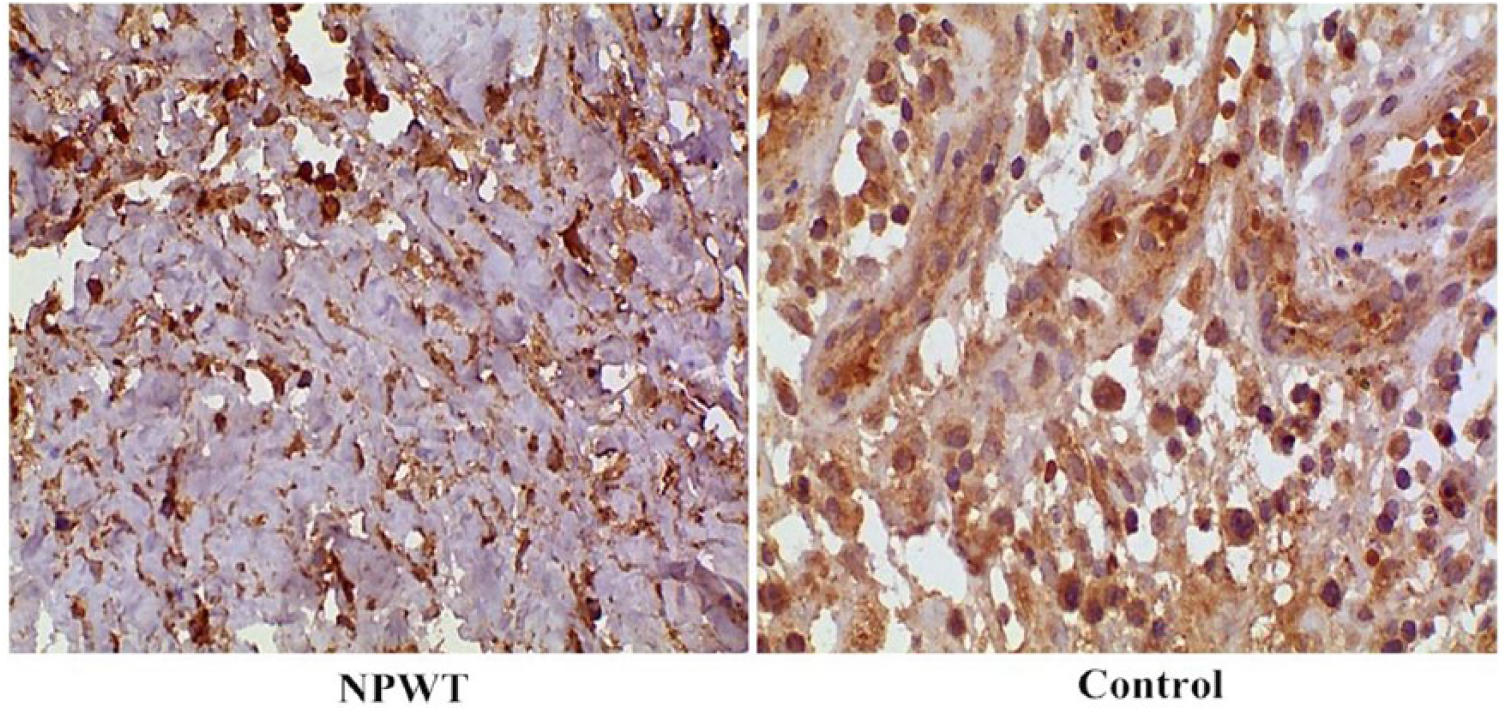
CD31-positive vascular endothelial cells exhibit a yellow/brown color after staining, with the densest staining occurring in the margins of granulation tissue. A portion of the new blood vessels in the granulation tissue exhibited tubular structures, while the other portion of endothelial cells shows a dotted or linear distribution. Figure 4 shows representative images of cells from the granulation tissue demonstrating increased staining for CD31 (a marker for MVD) after NPWT. Indeed, there was a significant increase (t = 13.82, P < .01) in the number of microvessels in the granulation tissue at 7 days after NPWT (12.03 ± 2.20) compared with that before NPWT (5.83 ± 1.73). These results suggest that NPWT may contribute to angiogenesis of the wound.
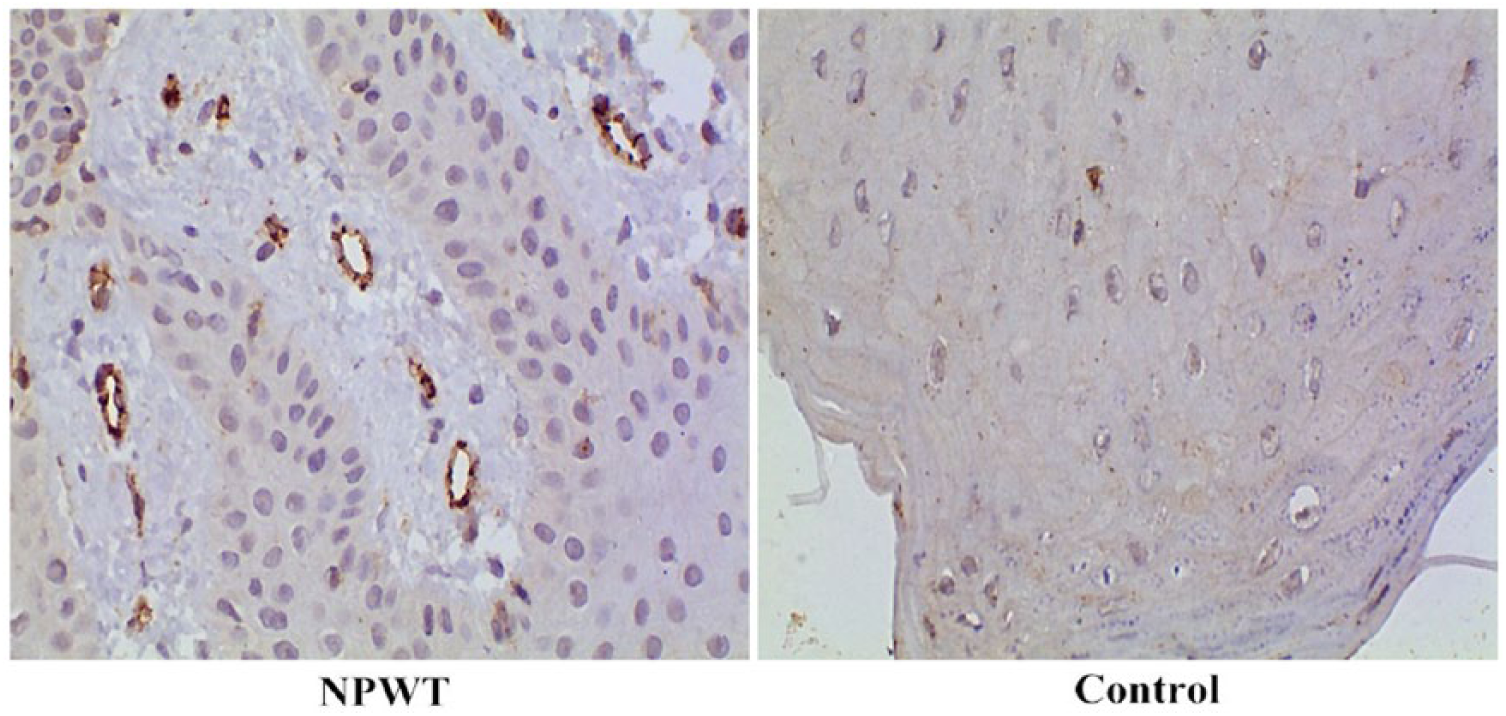
Immunohistochemical assessment of CD31 (a marker of microvessels) expression in wound biopsies from patients in the NPWT (negative-pressure wound therapy) and control groups. The brown color denotes positive staining. Representative images for each group are shown (magnification, 100×).
Correlations of miR-195, NLRX1, and MVD
Correlations among miR-195, NLRX1, and MVD in the granulation tissue of wounds were assessed using Spearman’s rank correlation coefficient. The results indicated that NLRX1 expression levels were negatively correlated to miR-195 levels (r = −.856, P < .001) and MVD (r = −.618, P < .05), whereas miR-195 expression and MVD were positively correlated (r = .630, P < .05) in the granulation tissue.
Discussion
Many factors contribute to the delayed healing or nonhealing of wounds, and the most important factor is arguably impaired or delayed angiogenesis. 4 Indeed, the process of angiogenesis, which involves the migration and proliferation of residual vascular endothelial cells to form new blood vessels, is a key stage in wound healing. Although the actual mechanism by which NPWT promotes wound healing remains unclear, it may involve the combined effects of the reduction of local edema and reduction of wound exudate accumulation. To explore this mechanism in greater detail, we examined the effects of wound healing and expression level changes in the granulation tissue in patients with various types of hard-to-heal wounds before and after receiving NPWT. NPWT significantly promoted wound healing in all cases, which was accompanied by an increase in the MVD in the wounds. This finding indicated that NPWT can effectively promote wound angiogenesis and improve local wound microcirculation, thus facilitating wound healing. Moreover, the various stages of angiogenesis are regulated by specific genes and molecules. Therefore, clarifying the influences of NPWT on the expression of angiogenesis-associated regulatory genes in chronic wound tissue will help gain an in-depth understanding of the treatment mechanism of NPWT, and may provide new targets for future clinical treatments of refractory chronic wounds.
Based on their biological functions, miRNAs can be categorized into angiogenesis-promoting miRNAs (eg, miR-21, miR-31, miR-27b, miR-126, miR-Boa, let-7f, miR-17-92, miR-210, miR-378, and miR-296) and angiogenesis-inhibiting miRNAs (eg, miR-15/16, miR-20, miR-92a, miR-214, miR-221/222, miR-320, and miR-328). miR-195 is an important member of the miR-15/16/195/424/497 family. Its existence was first predicted and proven in mice, 16 and then miR-195 of the same origin was subsequently found in human cells. 17 Although miR-195 has been associated with angiogenesis, the specific regulatory mechanisms were not elucidated. Wang et al 9 showed that miR-195 could inhibit VEGF expression by binding to the 3′ untranslated region of VEGF mRNA, thereby affecting the VEGF/VEGFR signaling pathways and inhibiting angiogenesis. MiR-195 was also shown to downregulate the expression of FGF1/CDC42, resulting in the inhibition of vascular smooth muscle cell proliferation and migration as well as the reduction of neointimal formation, thereby affecting angiogenesis. 18 Another study showed that miR-195 participated in cardiac remodeling via upregulation of the TGF-β1/Smads signaling pathway. 19 The results of the present study further expand the roles of miR-195 by demonstrating significant upregulation of miR-195 expression in the fresh granulation tissue with a high blood vessel density, suggesting a link between high levels of miR-195 expression and angiogenesis promotion.
Moreover, NLRX1 expression was shown to be associated with angiogenesis, with a significant decrease in NLRX1 expression levels in fresh granulation tissue accompanied by high MVD after NPWT. The activity of nuclear factor-kappa B (NF-κB) is associated with angiogenesis, which also regulates the transcription of VEGF, a key member of the family of angiogenic factors. In a study of malignant glioblastoma in a nude mouse model, Xie et al 8 found that blockade of the NF-κB signaling pathway resulted in the suppression of VEGF expression, which subsequently inhibited glioblastoma growth and angiogenesis. In a review by Tabruyn et al, 20 the new role of NF-κB in angiogenesis was discussed in detail, highlighting that VEGF expression is regulated by NF-κB. Moreover, Shibata et al 21 indicated that VEGF/VEGFR may be the downstream target of the NF-κB signaling pathway and that NF-κB mediates the angiogenic ability of breast tumors. In addition, many studies 22 have proven that NLRX1 can inhibit NF-κB activity or relevant NF-κB signaling pathways. One study indicated that NLRX1 could inhibit the downstream NF-κB pathway by interacting with TRAF6, 23 and Allen et al 24 showed that NLRX1 could interfere with the RIG-I/MAVS signaling pathway, thereby inhibiting the activation of downstream NF-κB and affecting its functions. Based on these previous findings and the present results, it can be inferred that NLRX1 may have an indirect inhibitory effect on angiogenesis through inhibition of NF-κB; however, further studies are required for elucidating the actual effect and the specific mechanisms. Overall, the present results indicate a negative correlation between miR-195 and NLRX1 expression levels, which further validates the prediction that NLRX1 may be a potential target of miR-195. However, further experimental studies are required for confirmation.
In conclusion, the results of this study indicate that NPWT may suppress the expression of NLRX1 by inducing the upregulation of miR-195 expression, and thereby promoting wound angiogenesis to ultimately promote wound healing. However, a larger sample size is required to validate the results obtained in this study, and further studies are needed to determine the specific molecular mechanism.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: (1) National Science Foundation of Hunan Province (2015JJ4053); (2) Hunan Health and Family Planning Commission Research Project (B2016-126, 2015-40); (3) Science and Technology Million Project of Inner Mongolia Medical University (YKD2017KJBW016); and (4) Inner Mongolia Autonomous Region Natural Science Fund (2018BS08009).