
Abstract
Peracetic acid (PAA) is extensively used in poultry processing, but its bactericidal mechanism remains poorly understood. Reactive oxygen species (ROS) are linked to bacterial cell death by many bactericidal agents. This study investigated the role of ROS in PAA against Salmonella Typhimurium 14028 using bacterial suspensions treated with PAA alone (20 ppm), PAA combined with 2,2ʹ-dipyridyl (Dip; iron chelator) or with thiourea (ROS scavenger) followed by incubation at 37°C for 1 h. Post-incubation, serial dilutions were plated on Luria-Bertani agar to determine colony-forming units per milliliter. Additionally, eight single-deletion mutants and the wild-type (control) of Salmonella Typhimurium 14028 were tested for survivability after exposure to PAA (treatment; 20 ppm) and hydrogen peroxide (H2O2; control). The mutants were categorized by predicted ROS impact: increase in ROS production (atpC, gnd, nuoG, pta, sdhC, and zwf) and no change in ROS production (negative control; edd and pykA). Finally, rifampicin-based selection assay was utilized to evaluate the mutation rate of the wild-type strain in the presence of PAA (60 ppm), H2O2 (control), and PAA + thiourea. Treatment with 20 ppm PAA significantly (p < 0.05) reduced the viability of Salmonella Typhimurium 14028 by a log10 reduction of 0.34, while co-treatment with thiourea restored counts to control level. The survival of the mutants predicted to increase ROS production was significantly reduced compared with the control mutants and wild-type strain (p < 0.05). Exposure to sublethal PAA led to a 28-fold increase in mutation rate suggesting the production of ROS by PAA was responsible for the observed increase. ROS production is a significant component of PAA’s bactericidal activity against Salmonella Typhimurium, and increasing ROS production might be exploited to enhance PAA-mediated killing of Salmonella Typhimurium. ROS production by PAA can lead to the development of antibiotic resistance in Salmonella Typhimurium when cells manage to escape cell death.
Introduction
Accumulation of reactive oxygen species (ROS) due to a bacteria’s inability to rapidly detoxify can lead to cell death (Schieber and Chandel, 2014; Zhao and Drlica, 2014). In aerobic bacteria, ROS are typically generated as by-products of metabolism. Generally, bacterial cells can mitigate levels of ROS through detoxification mechanisms (Juan et al., 2021). The balance between ROS production and detoxification is critical for maintaining cellular homeostasis and preventing oxidative stress-related damage. Oxidative agents such as chlorine and peracetic acid (PAA) have been commonly used in poultry production (Bauermeister et al., 2008; Cano et al., 2021; Aljuwayd et al., 2025). However, in previous research, we demonstrated that exposure to sublethal concentrations of antimicrobials such as sodium hypochlorite significantly increased the mutation rate in Salmonella Typhimurium 14028 through ROS production and led to kanamycin resistance (Aljuwayd et al., 2024).
ROS, particularly hydroxyl radicals (·OH), are both highly reactive and extremely short-lived—typically in the nanosecond range—and react at nearly diffusion-limited rates with surrounding biomolecules, resulting in minimal diffusion within cells and highly localized oxidative damage (Fasnacht and Polacek, 2021; Jomova et al., 2023; Tripathy and Oelmüller, 2012). Experimentally increasing endogenous ROS levels has been shown to sensitize bacteria such as Escherichia coli to oxidative stress and boost antibiotic efficacy (Brynildsen et al., 2013; Kohanski et al., 2010a; Dwyer et al., 2007; Hébrard et al., 2009; Liu et al., 2008). Such strategies hold promises in addressing antibiotic resistance by increasing the likelihood of cellular death while minimizing the opportunity for mutations (Van Acker and Coenye, 2017). Exploring ROS-mediated mechanisms can further expand understanding of bacterial physiology and create new opportunities for innovative antimicrobial approaches (Paulsen and Carroll, 2013; Miller and Britigan, 1997; Murugaiyan et al., 2022).
Thiourea and 2,2ʹ-dipyridyl (Dip), compounds that modulate ROS levels, were used in the current study to assess the effect of ROS on cellular death and mutation rate. Thiourea is generally used as a scavenger of hydroxyl radicals, the most reactive and damaging ROS. Thiourea neutralizes ROS by reacting with hydroxyl radicals in an oxidative agent such as hydrogen peroxide (H2O2). Using thiourea, Karash and Kwon (2018) demonstrated the bactericidal effect of antibiotics on Salmonella Typhimurium through increased ROS production (Karash and Kwon, 2018). Dip chelates ferrous iron, thereby inhibiting the Fenton reaction, which converts H2O2 into hydroxyl radicals. By limiting this reaction, Dip reduces the intracellular generation of hydroxyl radicals and other ROS. Korobov, Lemkina and Polyudova (2022) illustrated the bactericidal effect of warnerin on Staphylococcus cohnii via ROS accumulation using Dip.
Based on previous reports that oxidative stress, particularly hydroxyl radical formation, plays a central role in the bactericidal action of oxidizing agents against Gram-negative bacteria, we hypothesized that PAA’s antimicrobial activity against Salmonella Typhimurium involves the generation of ROS, including hydroxyl radicals. We anticipated that decreasing ROS levels with Dip or thiourea would protect Salmonella Typhimurium while using mutant strains of specific energy production genes would increase ROS production and oxidative stress susceptibility. We also expected that sublethal PAA exposure would result in a higher mutation rate due to elevated ROS levels compared with treatments with PAA plus thiourea or Dip and control groups. The objectives of this study were to: (1) Evaluate the role of ROS in the bactericidal action of PAA by assessing the impact of ROS suppression with thiourea and Dip on the survivability of Salmonella Typhimurium under PAA treatment. (2) Investigate the effect of gene deletions on ROS production and bacterial survivability by comparing the survivability of different Salmonella Typhimurium mutants with single-gene deletions that either increase or have no impact on ROS production when exposed to PAA and H2O2. (3) Determine the mutation rate of Salmonella Typhimurium exposed to sublethal concentrations of PAA using a rifampicin-based selection assay.
Materials and Methods
Effect of PAA on Salmonella Typhimurium reduction via ROS
In the current study, we used ROS-modulating agents thiourea and Dip (Sigma-Aldrich, St. Louis, MO) in combination with PAA to evaluate the role of ROS on the bactericidal action of PAA (Sigma-Aldrich) against Salmonella Typhimurium 14028 wild-type strain. Salmonella Typhimurium was cultured by inoculating one colony into Luria-Bertani (LB) broth and incubated overnight at 37°C with vigorous shaking (300 rpm) under aerobic conditions. The prepared culture (10 µL) was inoculated into 10 mL LB broth and incubated for 16 h under the same conditions. The culture was washed twice with phosphate-buffered saline (PBS; pH 7.4) and pelleted. The pellet was resuspended in 10 mL PBS, and 1 mL aliquot was distributed into three tubes. Six respective treatments were administered: (1) No treatment, (2) PAA alone (20 ppm), (3) PAA with Dip (20 ppm and 500 µM, respectively), (4) PAA with thiourea (20 ppm and 150 mM, respectively), (5) Dip alone (500 µM, reagent control), and (6) thiourea alone (150 mM, reagent control). The cultures were incubated at 37°C with vigorous shaking (300 rpm) in aerobic conditions. The samples were taken at 1 h of incubation, serially diluted, and plated in duplicates onto LB agar. The plates were incubated at 37°C for 24 h under aerobic conditions. The colonies were then enumerated to determine colony-forming units per milliliter (CFU/milliliter). PBS was used as the suspension medium for all treatments to eliminate growth-related variables and focus on the direct bactericidal effects of PAA. This also ensured controlled redox conditions during ROS modulation experiments involving thiourea and Dip. The PAA concentration of 20 ppm was used for bactericidal and ROS-modulation experiments. Figure 1a describes an overview of the experimental procedure.
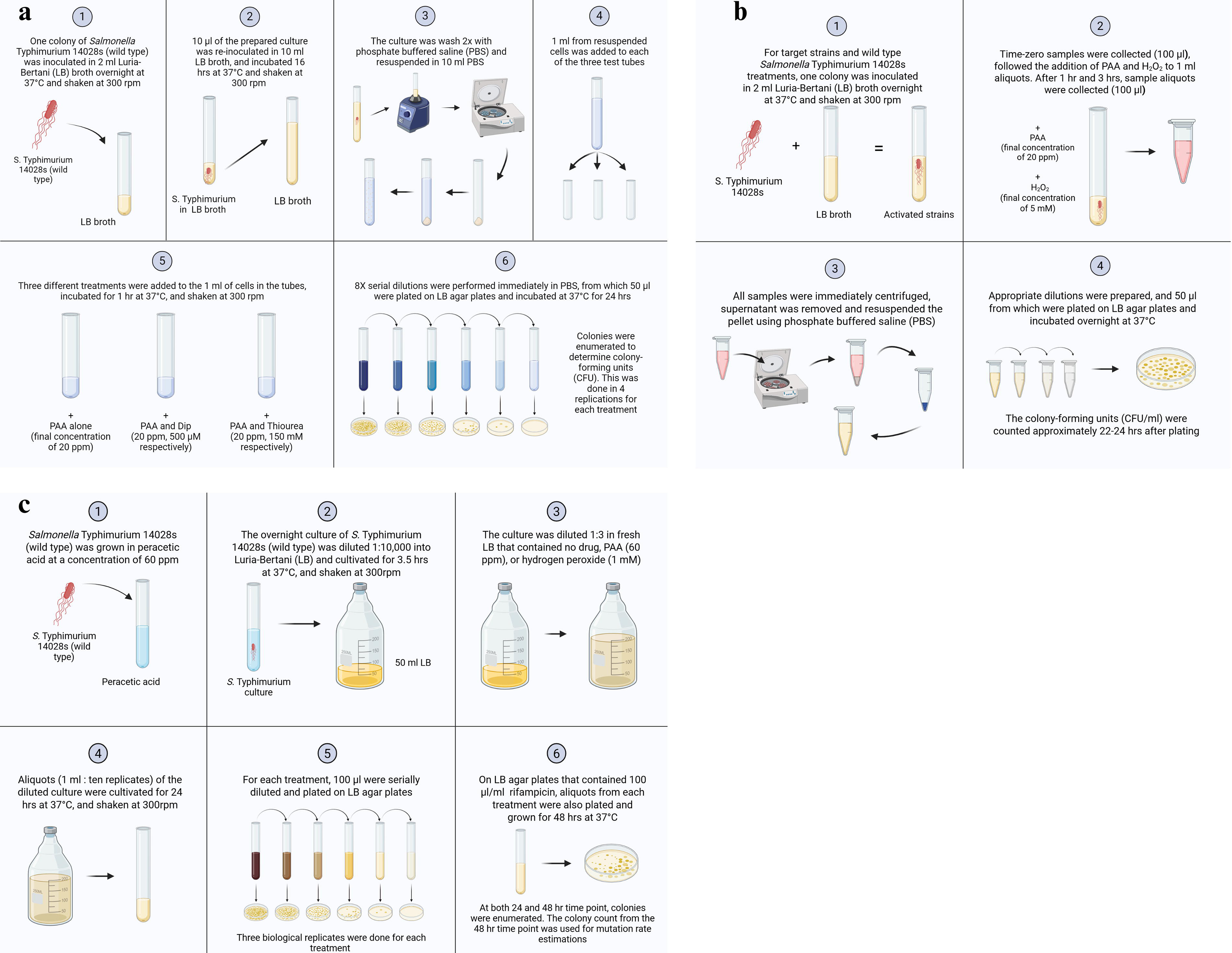
Overview of experimental design.
Effect of PAA and H2O2 on Salmonella Typhimurium mutants and wild-type strain
We assessed the survivability of eight Salmonella Typhimurium 14028 single-gene deletion mutants to investigate the role of ROS in the bactericidal action of PAA. These mutants were predicted to modulate ROS production in the presence of PAA (Biodefense and Emerging Infections [BEI], Manassas, VA; Table 1). Eight Salmonella Typhimurium 14028 mutants and a wild-type strain were cultured in LB broth and incubated overnight at 37°C with vigorous shaking (300 rpm). The experiments involving the mutant strains were conducted under the same conditions and the same protocol as described for the wild-type Salmonella Typhimurium. This includes culture preparation, PBS washing, PAA exposure, and incubation conditions. The fresh cultures were transferred into treatments at 107–108 CFU/mL and incubated aerobically at 37°C at 300 rpm. After time-zero samples were collected, 1 mL of each culture was aliquoted into 15 mL test tubes containing PAA and H2O2 (20 ppm and 5 mM H2O2 final concentration, respectively). The samples were collected at 1 and 3 h and centrifuged (8,385 × g). The pellets were resuspended in PBS and serially diluted. 50 µL of dilutions were plated on LB agar and incubated overnight at 37°C aerobically for 24 h. The colonies were then enumerated to assess CFU/mL. Survival percentages for each strain were calculated relative to its own time-zero average CFU/mL. The identity of all mutant strains was confirmed by PCR using gene-specific primers flanking the targeted deletion, as described in the BEI Resources documentation. Figure 1b describes an overview of the experimental procedure.
List of the Single Deleted Genes Mutants Used in This Study Along with Their Synthesized Protein, Protein Functionality, and Source Reference
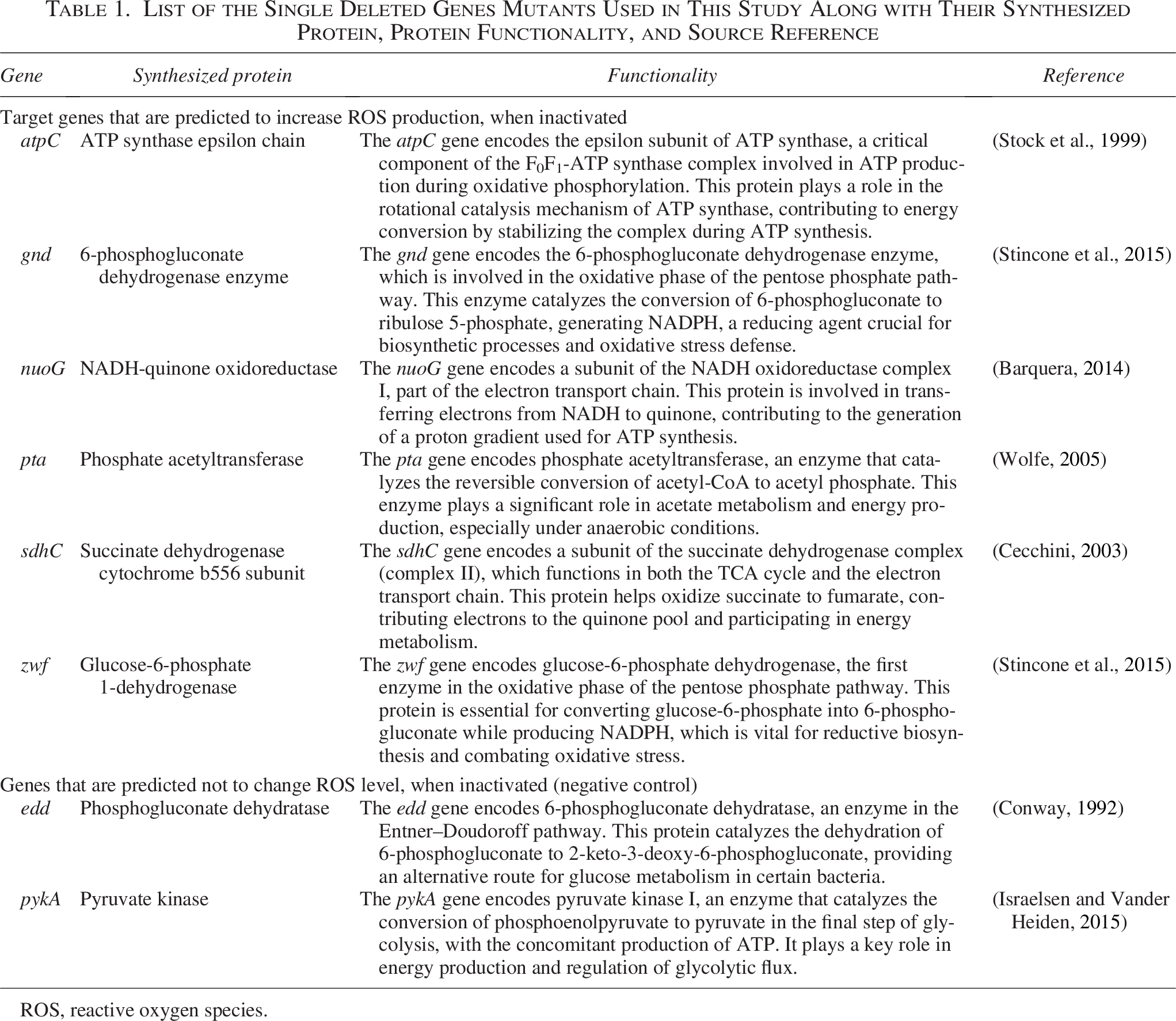
ROS, reactive oxygen species.
Determination of mutation rate
We used a rifampicin-based selection assay to determine the mutation rate of wild-type Salmonella Typhimurium 14028 exposed to sublethal concentrations of PAA (60 ppm), as our preliminary study showed it induced a 60% reduction in bacterial survival. The PAA concentration was chosen based on a level that induced a detectable effect on growth or survivability within the first 6 h following PAA addition. This concentration was used only for mutation rate determination. Later, culture density was recovered to reach near the untreated colony density 24 h after treatment. This allowed a comparison between mutation frequencies for cultures of similar densities following treatment with a PAA or H2O2. Mutation rates were assessed after 24 h using a rifampicin-based selection assay (Giraud et al., 2001). An overnight culture of Salmonella Typhimurium was diluted 1:10,000 in LB broth and incubated for 3.5 h at 37°C at 300 rpm. The 3.5-h incubation allowed Salmonella Typhimurium to reach the logarithmic growth, during which active DNA replication maximizes the likelihood of mutation occurrence. This standardized growth phase ensures that mutation rates were assessed under optimal and comparable conditions across experimental treatments, enhancing the reliability and validity of the results (Giraud et al., 2001). The culture was diluted 1:3 in fresh LB broth, PAA (60 ppm), or H2O2 (1 mM). Aliquots with 10 (1 mL) replicates for each treatment were incubated at 37°C at 300 rpm for 24 h. Each sample was serially diluted and plated on LB agar containing 100 µg/mL rifampicin (Fisher Scientific, Pittsburgh, PA) to determine the number of CFU/mL. The plates were incubated under aerobic conditions at 37°C for 48 h. Colonies were enumerated at 48 h, and mutation rates were estimated. The colony counts from 10 replicates were used in the MSS maximum-likelihood method to calculate the mutational events per culture (Rosche and Foster, 2000; Sarkar, Ma and Sandri, 1992). The mutation rate was determined by dividing the mutational events per culture by the number of bacteria plated (Rosche and Foster, 2000). The change in mutation rate was estimated for all treatments and untreated Salmonella Typhimurium control. Three biological replicates were used for each treatment. Figure 1c describes an overview of the experimental procedure.
Statistical analysis
All experiments were conducted with a minimum of three independent biological replicates. For bactericidal assays, results are reported as mean log10 CFU/mL ± standard deviation. For ROS-modulation assays, results are expressed as survivability percentages relative to the untreated control, calculated from mean CFU counts (log10 CFU/mL). For mutation rate determination, mutation frequencies were calculated using the MSS maximum-likelihood method (Rosche and Foster, 2000; Sarkar, Ma and Sandri, 1992), and mutation rates were obtained by dividing the estimated mutational events per culture by the number of viable cells plated. Differences in mean mutation rates, log reductions, and survival percentages were evaluated via one-way analysis of variance, followed by Tukey’s post-hoc test for multiple comparisons. Statistical significance was determined at p < 0.05. All analyses were conducted using JMP software (JMP®, Version JMP Pro-17; SAS Institute Inc., Cary, NC, 1989–2023).
Results
Effect of PAA on Salmonella Typhimurium reduction via ROS
The role of ROS in the bactericidal action of PAA against Salmonella Typhimurium 14028 wild-type strain was assessed by comparing the reduction in Salmonella Typhimurium counts following treatment with PAA and a combination of PAA with either Dip or thiourea against a control group with no treatment. PAA alone (20 ppm) significantly reduced Salmonella Typhimurium counts to 8.96 log10 CFU/mL (relative to the untreated control, 9.3 log10 CFU/mL; p < 0.05; Fig. 2). The addition of Dip (500 µM) to PAA (20 ppm) significantly mitigated the reduction in Salmonella Typhimurium 14028 to 9.08 log10 CFU/mL, compared with the untreated control (p < 0.05; Fig. 2). When thiourea (150 mM) was used in combination with PAA (20 ppm), the bactericidal effect of PAA was abolished entirely, with no significant difference from the untreated control group. The treatment of thiourea alone (150 mM) as a reagent control did not show any bactericidal activity, while Dip (500 µM) as a reagent control showed some reduction in bacterial survival (p > 0.05; Fig. 2). In our study, we verified that 150 mM thiourea did not inhibit bacterial growth, and its inclusion abolished the bactericidal effect of PAA, supporting its role as an effective ROS scavenger under the tested conditions.
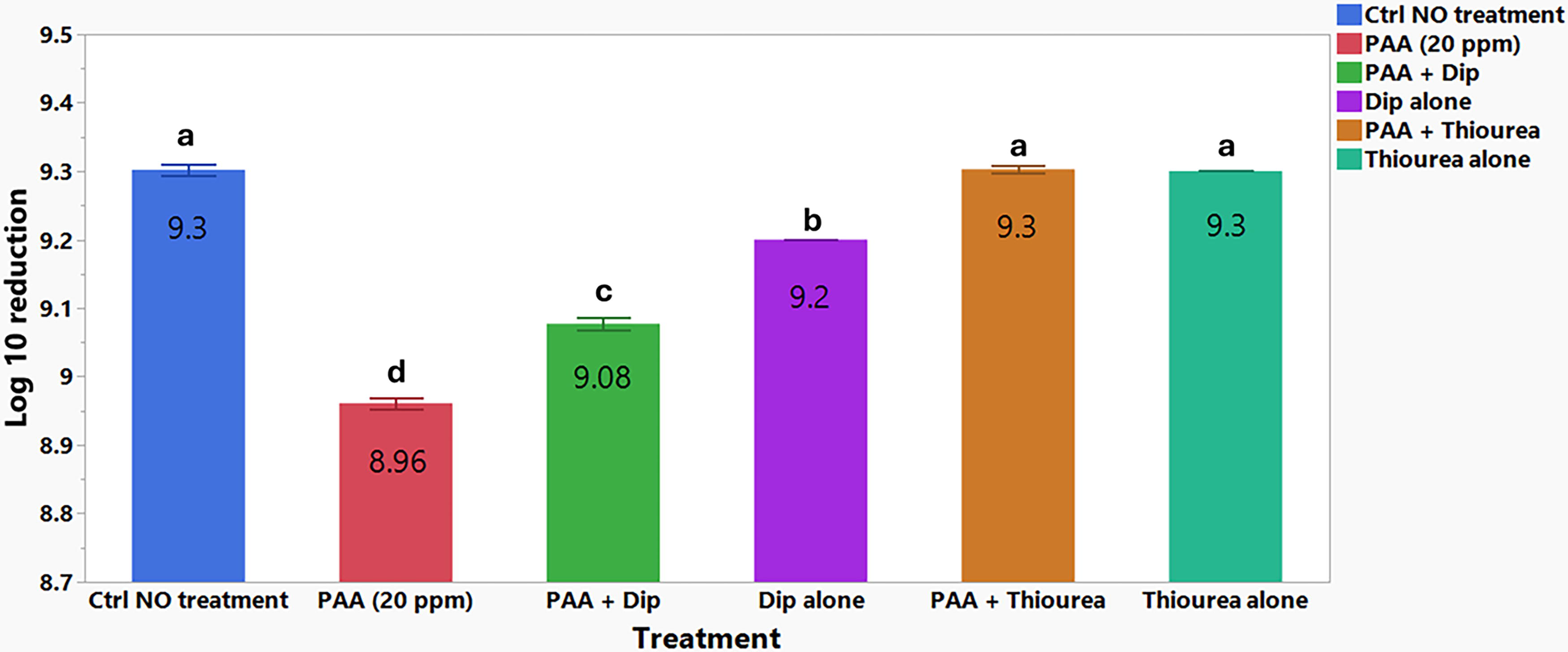
Log10 survivors of Salmonella Typhimurium 14028 following treatment with PAA alone (20 ppm) and PAA combined with compounds that modulate ROS production, Dip (2,2ʹ-dipyridyl, 500 µM) and thiourea (150 mM), and reagent controls alone Dip (2,2ʹ-dipyridyl, 500 µM) and thiourea (150 mM) aerobically for 1 h at 37°C. The letters a–d denote significant pairwise differences between the treatments (p < 0.05). PAA, peracetic acid; ROS, reactive oxygen species.
Effect of PAA and H2O2 on Salmonella Typhimurium mutants and wild-type strain
To evaluate the impact of ROS production on the survivability of Salmonella, growth of ROS-producing mutants of Salmonella was compared with the wild-type strain when treated with PAA (20 ppm) and H2O2 (5 mM). The samples were treated for 1 and 3 h, and mutant survival was compared based on their designated categories. The mutants (Table 1) were grouped into two categories. Target mutants were predicted to increase ROS production, enhancing oxidative stress and cell death susceptibility. This group included the mutants atpC, gnd, nuoG, pta, sdhC, and zwf. Negative controls were predicted to produce the wild-type level ROS, including the mutants edd and pykA. Target mutant survivabilities were significantly reduced at both 1 and 3 h of treatment with PAA (20 ppm) compared with the wild-type strain and 0 h (p < 0.05; Fig. 3). When treated with H2O2 (5 mM), Salmonella Typhimurium mutant survivability was also reduced, though not always significantly compared with the control group and 0 h. Negative control survivabilities were slightly reduced after treatment with PAA and H2O2 similarly to the control group at the 1-h treatment; however, the reduction was less at the 3-h treatment in comparison with the control group (wild-type group) (p > 0.05; Fig. 3).
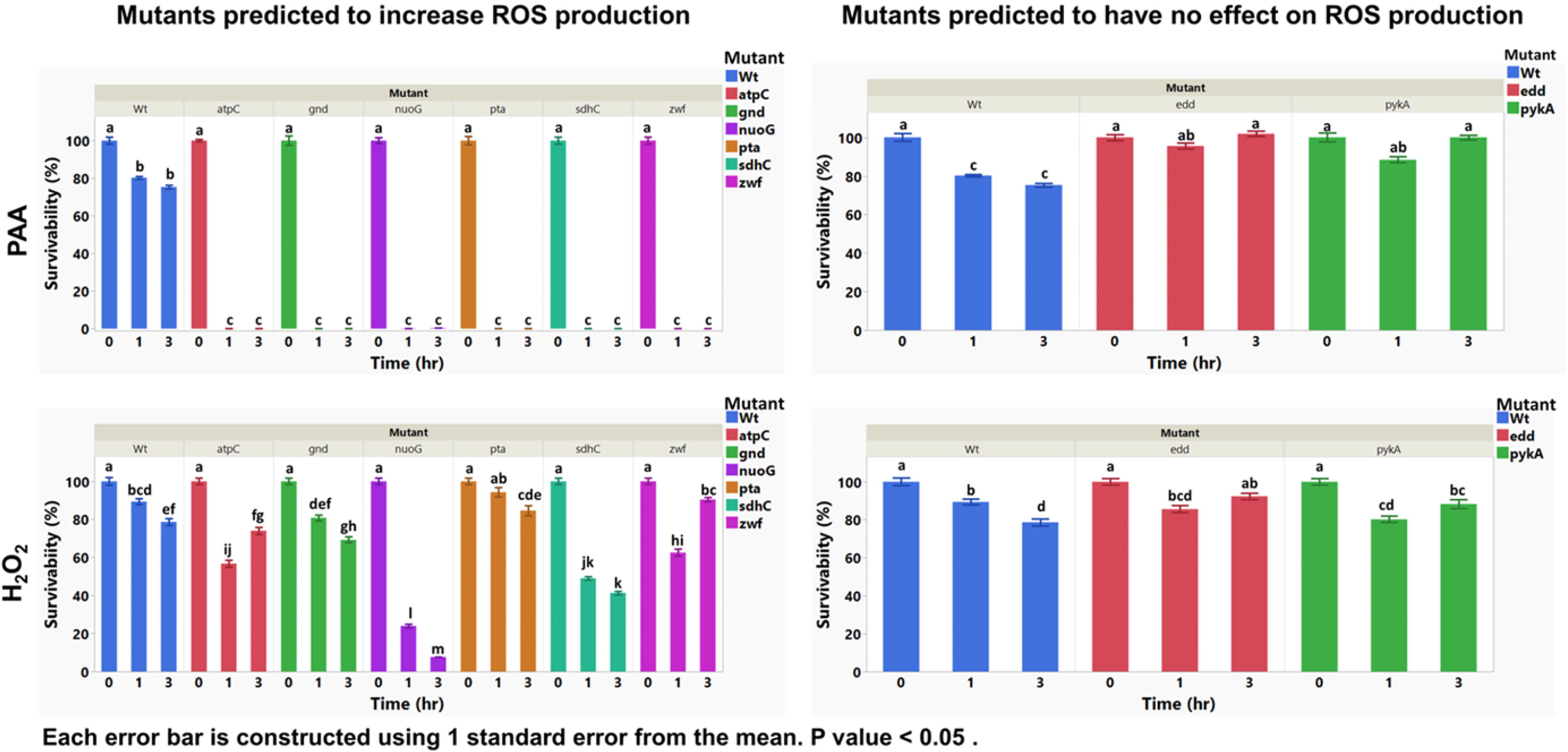
Percent survival of ROS production-associated mutants of Salmonella and a wild-type strain (Wt) when treated with PAA (20 ppm) and H2O2 (5 mM) after 1 and 3 h of treatment. The letters a–m denote significant pairwise differences between the treatments (p < 0.05). Survival percentages for each strain were calculated relative to its own time-zero average CFU/mL. CFU, colony-forming unit; H2O2, hydrogen peroxide; PAA, peracetic acid; ROS, reactive oxygen species.
Determination of mutation rate
The mutation rates of the wild-type Salmonella strain were compared under different conditions to assess ROS impact on mutagenesis. The treatments included PAA, H2O2, and PAA combined with thiourea. Significant differences in the fold change of mutation rates were observed between PAA and control groups (Fig. 4). Treatment with PAA (60 ppm) resulted in an approximately 28-fold increase in the mutation rate compared with the no treatment control group. However, when thiourea (150 mM) was added to PAA, the fold change in the mutation rate was significantly reduced to about 7-fold (p < 0.05). Although H2O2 (1 mM) significantly increased the mutation rate in Salmonella (∼22-fold) compared with the negative control group, it was still significantly lower than the mutation rate observed with PAA (p < 0.05; Fig. 4).
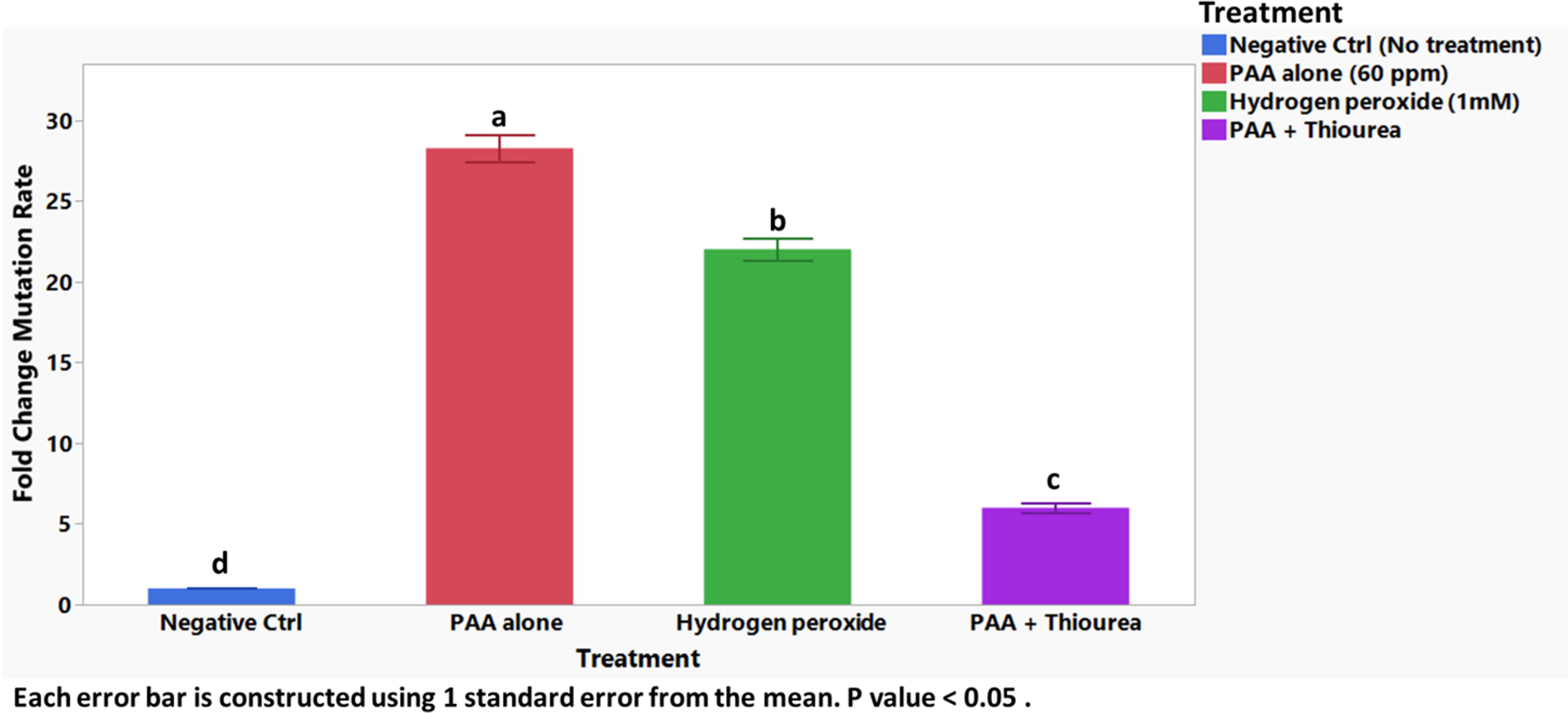
Fold change mutation rates of Salmonella Typhimurium 14028 exposed to sublethal concentrations of PAA (60 ppm), H2O2 (1 mM), PAA + Thiourea, and negative control after 24 h using rifampicin-based selection assay. The letters a–c denote significant pairwise differences between the treatments (p < 0.05). H2O2, hydrogen peroxide; PAA, peracetic acid.
Discussion
The findings from the current study indicate the importance of ROS in antimicrobial oxidative agent antimicrobial activities. Kohanski et al. (2010b) demonstrated that bactericidal antibiotics induce cellular damage by producing hydroxyl radicals via the Fenton reaction, leading to cell death. Similarly, Touati (2000) demonstrated the importance of ROS production on bacterial death during antimicrobial application. The authors reduced the bactericidal efficacy of H2O2 using an iron chelator that inhibits the formation of hydroxyl radicals. Imlay (2008) and Dwyer et al. (2014) showed ROS’s wide range of functions in microbial physiology and a lethal role for antibiotic mechanisms. Similarly, in the current study, we observed that oxidative stress induced by PAA involves mechanisms associated with the generation and accumulation of ROS (Fig. 2).
The susceptibility of specific mutants to PAA and H2O2 revealed that gene deletions predicted to increase ROS production resulted in significantly higher sensitivity to PAA-induced oxidative stress (Fig. 3). Disruptions in these pathways are potentially linked to increased ROS production through multiple mechanisms: (1) Inefficient electron transfer in the electron transport chain (nuoG, sdhC, atpC) disrupts the normal flow of electrons, causing electron leakage and superoxide formation (Stock et al., 1999; Barquera, 2014; Cecchini, 2003). (2) Reduced NADPH production and antioxidant defense, due to deletions in genes involved in the pentose phosphate pathway (gnd, zwf), impairs the bacterium’s ability to detoxify ROS and maintain redox balance (Stincone et al., 2015). (3) Metabolic imbalances caused by disrupted acetate metabolism (pta) alter cellular respiration and exacerbate oxidative stress (Wolfe, 2005). Negative control mutants (edd and pykA), predicted to produce ROS levels similar to the wild-type strain, exhibited survivability patterns closer to the wild-type under both treatments. While slight reductions in survivability were observed at the 1-h mark, their resilience at 3 h suggests that oxidative defenses remained intact.
The mutation rate significant increases observed with PAA (Fig. 4) indicates ROS can cause DNA damage, leading to mutations that may contribute to antibiotic resistance. The reduction in mutation rates with the addition of thiourea underscores the central role of hydroxyl radicals in this process. This is consistent with previous studies demonstrating its effectiveness in scavenging hydroxyl radicals and mitigating oxidative damage in E. coli without adversely affecting cell viability (Davies et al., 2009; Chueca, Pagan and Garcia-Gonzalo, 2014; Liou et al., 2015; Aljuwayd et al., 2025). Taken together, these results emphasize the critical role of ROS in mediating PAA’s bactericidal action. They also underscore the importance of specific metabolic pathways in determining bacterial susceptibility to oxidative stress. This study contributes to a deeper understanding of how disruptions in energy metabolism and redox balance heighten bacterial vulnerability, providing potential targets for enhancing the efficacy of oxidative stress-based antimicrobial treatments. Although this study provides insights into the role of ROS bactericidal PAA activity against Salmonella Typhimurium, several limitations must be acknowledged. These in vitro experiments do not fully replicate the complexities of food processing environments where organic matter and other factors could influence the effectiveness of PAA, and future research should include matrices such as poultry meat. Additionally, the study focused on a single strain of Salmonella Typhimurium and single-gene deletion mutants which do not account for potential genetic interactions or compensatory mechanisms that could influence ROS production. Exploring the response of multi-deleted gene mutants, additional strains/serotypes of Salmonella as well as other foodborne pathogens would help assess whether the observed ROS-mediated killing mechanism occurs more broadly. Finally, the current study did not explore other bacterial responses, such as DNA repair or protein oxidation. Further research on these molecular pathways would provide more understanding of how ROS influences bacterial survival and mutation.
Conclusions
The current study underscores the critical role of ROS in the bactericidal action of PAA and potential mutagenesis in Salmonella Typhimurium. Our findings highlighted oxidative stress as a significant mechanism of action in the bactericidal activity of PAA on Salmonella Typhimurium. However, our findings also indicated that sublethal exposure to PAA increased the mutation rate in Salmonella Typhimurium, inducing antibiotic resistance. Due to PAA’s ability to induce mutagenesis in Salmonella Typhimurium endogenous ROS modulation should be considered as a complementary strategy to enhance the bactericidal effects of oxidative agents such as PAA. Understanding and manipulating ROS dynamics could lead to more effective antimicrobial treatments, especially for Salmonella in certain poultry products (CDC, 2023; USDA, 2023).
Authors’ Contributions
Funding acquisition, methodology, project administration, resources, validation, and writing of the original draft was done by Y.M.K. and M.K.W.A. Conceptualization and supervision were done by Y.M.K. Data curation, formal analysis, software, visualization, and investigation were done by M.K.W.A. Writing, review, and editing of final draft were done by M.K.W.A., E.G.O., S.C.R., M.J.R., and Y.M.K.
Footnotes
Acknowledgments
The eight single-deletion mutants of Salmonella Typhimurium 14028 used in this study were obtained through the NIH Biodefense and Emerging Infections Research Resources Repository (BEI resources), National Institute of Allergy and Infectious Diseases, National Institutes of Health. The authors thank Haley Tarcin for article formatting and construction of ![]() .
.
Funding Information
No funding was received for this article.
Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that influence the work reported in this article.